
Enhancing Production of Medium-Chain-Length Polyhydroxyalkanoates from Pseudomonas sp. SG4502 by tac Enhancer Insertion



1
International Cooperation Research Center of China for New Germplasm Breeding of Edible Mushrooms, Jilin Agricultural University, Changchun 130118, China
2
Jilin Province Key Laboratory of Fungal Phenomics, Jilin Agricultural University, Changchun 130118, China
3
School of Life Science and Technology, Changchun University of Science and Technology, Changchun 130022, China
4
Tianjin Institute of Industrial Biotechnology, Chinese Academy of Sciences, Tianjin 300308, China
5
Key Laboratory of Wetland Ecology and Environment, Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun 130102, China
*
Author to whom correspondence should be addressed.
Polymers 2023, 15(10), 2290; https://doi.org/10.3390/polym15102290
Submission received: 7 April 2023
/
Revised: 9 May 2023
/
Accepted: 10 May 2023
/
Published: 12 May 2023
Abstract
:Pseudomonas sp. SG4502 screened from biodiesel fuel by-products can synthesize medium-chain-length polyhydroxyalkanoates (mcl-PHAs) using glycerol as a substrate. It contains a typical PHA class II synthase gene cluster. This study revealed two genetic engineering methods for improving the mcl-PHA accumulation capacity of Pseudomonas sp. SG4502. One way was to knock out the PHA-depolymerase phaZ gene, the other way was to insert a tac enhancer into the upstream of the phaC1/phaC2 genes. Yields of mcl-PHAs produced from 1% sodium octanoate by +(tac-phaC2) and ∆phaZ strains were enhanced by 53.8% and 23.1%, respectively, compared with those produced by the wild-type strain. The increase in mcl-PHA yield from +(tac-phaC2) and ∆phaZ was due to the transcriptional level of the phaC2 and phaZ genes, as determined by RT-qPCR (the carbon source was sodium octanoate). 1H-NMR results showed that the synthesized products contained 3-hydroxyoctanoic acid (3HO), 3-hydroxydecanoic acid (3HD) and 3-hydroxydodecanoic acid (3HDD) units, which is consistent with those synthesized by the wild-type strain. The size-exclusion chromatography by GPC of mcl-PHAs from the (∆phaZ), +(tac-phaC1) and +(tac-phaC2) strains were 2.67, 2.52 and 2.60, respectively, all of which were lower than that of the wild-type strain (4.56). DSC analysis showed that the melting temperature of mcl-PHAs produced by recombinant strains ranged from 60 °C to 65 °C, which was lower than that of the wild-type strain. Finally, TG analysis showed that the decomposition temperature of mcl-PHAs synthesized by the (∆phaZ), +(tac-phaC1) and +(tac-phaC2) strains was 8.4 °C, 14.7 °C and 10.1 °C higher than that of the wild-type strain, respectively.
1. Introduction
Polyhydroxyalkanoates (PHAs) have attracted much interest from the scientific community due to their physiochemical properties, including biodegradability and biocompatibility, which are similar to those of petrochemical-based plastics. This intracellular polyester can be synthesized by a variety of bacterial strains under certain deficiency conditions [1] and can serve as a carbon source and energy reserve in bacterial cells [2,3]. PHAs are named according to the number of carbon atoms in their monomeric units: PHAs with C3 to C5 monomer units are commonly called short-chain-length (scl) PHAs, and those with C6 to C14 monomer units are called medium-chain-length (mcl) PHAs [4,5]. Compared with scl-PHAs, mcl-PHAs exhibit elastomer- or adhesive-like properties. Additionally, the scl and mcl copolymerized PHAs have been demonstrated to have better properties and application prospects [6]. PHA synthase (PhaC) is a kind of key enzyme in the PHA synthesis pathway as it determines the monomer structure, composition and molecular weight of PHAs [7]. PhaC can be divided into four categories, namely type I to IV, according to their primary structure, subunit composition and specificity to substrates with different chain length monomers [5,8]. Particularly, type II PhaC is the only polymerase that can synthesize mcl-PHAs from alkane (C6 to C14) precursors. The type II PHA synthases gene locus contains two genes, phaC1 and phaC2, encoding PHA synthase that mainly polymerize the thioesters of coenzyme A (CoA) containing medium- and long-chain PHA monomers to mcl-PHA through β-oxidation or the de novo biosynthesis of fatty acids [9]. Research has shown that PhaC1 and PhaC2 synthases from different Pseudomonas sp. strains generate different products through different synthetic pathways. Guo et al. found that engineered bacterium Pseudomonas mendocina C7C1 with overexpressing phaC1 from P. mendocina NK-01 could accumulate 9.77% mcl-PHA, while P. mendocina C7C2 with overexpressing phaC2 from the same bacterial strain could produce only 3.67% mcl-PHA. They also found that PhaC1 had higher catalytic activity than that of PhaC2 and was the major PHA synthase for mcl-PHA synthesis in P. mendocina [10]. Regarding the phaC gene cluster of Pseudomonas sp., the phaZ gene was found located between the phaC1 and phaC2 genes. It has depolymerase activity and can degrade mcl-PHA synthesized by PhaC1 and PhaC2 into aliphatic and aromatic monomers to provide energy for organisms [11,12,13].
To date, studies have shown that numbers of microorganisms can synthesize scl- or mcl-PHAs, but only Pseudomonas sp. can accumulate them, due to the presence of two PhaC polymerases, PhaC1 and PhaC2, which have different substrate specificity. Moreover, the incorporation of medium- and long-chain fatty acids into PHA could alter the brittleness and crystallinity of scl-PHAs, making them more elastic. Scl-mcl-PHAs with good elasticity have great application prospects in various industries, such as medicine, packaging and others [14]. However, it is difficult to produce mcl-PHA at a large scale due to the low synthetic efficiency of the strains. The production efficiency of scl-mcl-PHAs could potentially be improved using genetically engineered bacteria generated by deleting or enhancing PHA synthesis-related genes at the molecular level. Two genetic engineering methods have been reported: enhancing the expression of PhaC enzymes and deleting the PHA depolymerase-encoding gene phaZ located between phaC1 and phaC2. Akihiko Kondo et al. constructed a recombinant strain by knocking out the depolymerase (phaZ) gene and overexpressing genes of poly (3-hydroxyalkanoate) (PHB) biosynthesis to further improve production for PHB [15]. It was reported that the engineered bacteria constructed by overexpressing phaC1 or phaC2 genes can improve the synthesis efficiency of mcl-PHA. José G.C. Gomez et al. reported that engineered Pseudomonas containing PHA synthases of Aeromonas spp. can successfully produce P(3HB-co-HAMCL) [16]. Nevertheless, recombinant plasmid containing the target gene is unstable and easily lost during the subculture of the host cells. Furthermore, a tac enhancer derived from a functional hybrid of trp and lac promoters [17] has been reported to be able to control the expression levels of the target gene [18,19]. For example, Zhang et al. constructed a recombinant strain by integrating a tac enhancer on chromosomes to improve L-ornithine production. The recombinant strain had L-arginine production of 25 g/L, which was 63.4% higher than that of the original strain (15.3 g/L) [20]. Fukui et al. demonstrated that the expression of gfp under the control of a Ptac promoter was 1.5- to 2-fold higher compared to that of Plac and PphaC [21]. The above studies indicate that increasing the expression of the target gene using the tac enhancer could improve the yield of the product. Nonetheless, the use of a tac enhancer to enhance the expression of PhaC to improve mcl-PHA synthesis in Pseudomonas sp. strains has not been verified [22,23,24].
The presence of the phaZ gene on the PHA synthesis gene cluster might be one of the reasons for the low efficiency of mcl-PHAs-producing strains containing the Class II PHA synthase genes [25]. The recombinant strain P. putida KTMQ01 generated by knocking out the PHA depolymerase phaZ gene enhanced the accumulation of mcl-PHA by 20% of cell dry weight (CDW, titer) compared to the mutant P. putida KT2442 [26]. However, it has been reported that the yield of mcl-PHA produced by recombinant P. putida KT2440 was reduced by 13.9% compared to that produced by the wild-type strain under carbon-limited conditions [27]. The differences in mcl-PHA yield may be derived from different synthetic pathways, PhaC expressions and metabolic balances between different Pseudomonas sp. strains, even after the PHA depolymerase phaZ gene is deleted.
Thus, this study aims to obtain mcl-PHA with high yields and better thermal stability properties from genetically engineered strains. The strains were generated by inserting a tac enhancer on the upstream of phaC1 or phaC2 genes to enhance their expression. As an alternative, the phaZ gene was deleted from the PHA gene cluster to inhibit PHA degradation in SG4502. Three recombinant strains, +(tac-phaC1), +(tac-phaC2) and (∆phaZ), were constructed, and their mcl-PHA synthesis ability was compared using the liquid fermentation method. Gene expression was evaluated using real-time quantitative PCR (RT-qPCR) to explain the differences between the expression of mcl-PHA from the phaC1, phaC2 and phaZ genes in the three recombinant strains. The composition, structure and molecular weight of the products from the three recombinant strains were determined by hydrogen nuclear magnetic resonance spectroscopy (1H-NMR) and gel permeation chromatography (GPC). Moreover, the thermal properties of the products were determined by differential scanning calorimetry (DSC) and thermogravimetric analysis (TGA).
2. Materials and Methods
2.1. Strains and Plasmids
Pseudomonas sp. SG4502 (NCBI: txid547985) was provided by a laboratory at Hokkaido University, Japan. Escherichia coli DH5α and E. coli JM109 were preserved in the laboratory. Plasmids pUC19 and pCDFDuet-1 were purchased from MiaoLing Company (Wuhan City, China). Plasmid pK18 (containing the strong tac enhancer) was provided by Professor Guangming Xiong, Kiel University, Germany. All strains and plasmids used in this study are listed in Table S1.
2.2. Reagents and Media
Tryptone, yeast extract, NaCl, glycerol, Nile red, dimethyl sulfoxide (DMSO), agarose, absolute ethanol, agar powder, NaOH, Na2EDTA·2H2O, Tris and glacial acetic acid were purchased from Beijing Chemical Plant (Beijing, China). Ampicillin, Kanamycin, Streptomycin, chloramphenicol and tetracycline were purchased from Baotaike (Shijiazhuang, China). X-gal and IPTG were purchased from Shanghai QianChen Biotechnology Co., Ltd. (Beijing, China). All chemical reagents were of analytical grade. The concentration of antibiotics used in this study was as follows: Campicillin = 100 mg·mL−1, Ckanamycin = 50 mg·mL−1, Cstreptomycin = 50 mg·mL−1, Ctetracycline = 50 mg·mL−1 and Cchloramphenicol = 25 mg·mL−1.
A Lysogeny Broth (LB) medium was prepared by vigorously mixing 1.0 g of sodium chloride, 1.0 g of tryptone and 0.5 g of yeast extract in 100 mL of ultrapure water using a magnetic stirrer [28]. The medium was used to cultivate wild-type SG4502 cells (45 °C, 180 rpm).
In the pre-culture of all recombinant bacteria (45 °C, 180 rpm), a nutrient broth (NB) was used. The broth was prepared by thoroughly mixing 24.0 g of NB broth and 3 g of yeast extract in 1 L of ultrapure water. Additionally, 1.5 g of agar powder was added to solidify the NB medium.
An inorganic salt medium (MS) was prepared by dissolving 3.6 g of Na2HPO4, 1.5 g of KH2PO4, 1.0 g of NH4Cl, 0.2 g of MgSO4·7H2O and 1 mL of trace elements in 1 L of ultrapure water. The bottle was sealed with a sealing film after the solution was clarified [29]. After a carbon source was added, PHA for fermentation and cultivation (45 °C, 180 rpm).
All the above media were divided into tubes and sterilized by autoclaving at 121 °C and 1 atm for 20 min.
2.3. Experimental Methods
2.3.1. Construction of Recombinant Strain ∆phaZ
The PHA depolymerase-encoding gene phaZ located between the phaC1 and phaC2 genes was knocked out from the genome of wild-type SG4502 by inserting the smr gene into the phaZ gene to interrupt its transcription. The construction process of ∆phaZ is shown in Figure 1 and has been described previously [25]. Using the genomic DNA of wild-type SG4502 as a template, PCR amplification of the pC1ZC2 (C1-Z-C2) fragment was carried out using HS DNA Polymerase (TOYOBO, Osaka, Japan), an upstream primer, pC1ZC2-F, and a downstream primer, pC1ZC2-R. The amplified fragment contained about 500 bp of phaC1, the full-length sequence of phaZ and about 500 bp of phaC2; the total sequence length was 2185 bp (C1-Z-C2). pC1ZC2 (C1-Z-C2) was digested by the HindIII and EcoRI restriction enzymes and then inserted into pUC19 plasmid treated with the same restriction enzymes using EasyGeno Rapid Recombinant Cloning Kit (TIANGEN, Beijing, China). Then, the recombinant plasmid pUC19-C1ZC2 was transferred into E. coli JM109, and colonies containing the recombinant plasmid pUC19-C1ZC2 were screened using blue-white screening. An smr gene fragment cloned from pCDFDuet-1 using Smr-F and Smr-R primers was inserted into the phaZ sequence of the recombinant plasmid pUC19-C1ZC2 using the SacII restriction enzyme to prevent the translation of the phaZ gene. Afterwards, the reaction solution was transferred into E. coli JM109 using the heat shock method, and bacterial colonies containing recombinant plasmid pUC19-C1ZC2-smr were screened using Ampicillin and Spectinomycin resistance screening. Moreover, the recombinant plasmid was sequenced by Jilin KuMei company (Changchun, China); after that, it was transferred into competent cells of wild-type SG4502 by electroporation to knockout the phaZ gene through homologous recombination. HindIII and EcoRI dual digestion reactions were used to identify the deletion mutant. The recombinant strain ∆phaZ was obtained after the transcriptional expression of phaZ was inhibited by smr insertion. The results were validated by PCR identification.
2.3.2. Construction of phaC1 and phaC2 Gene-Overexpression Recombinant Strains
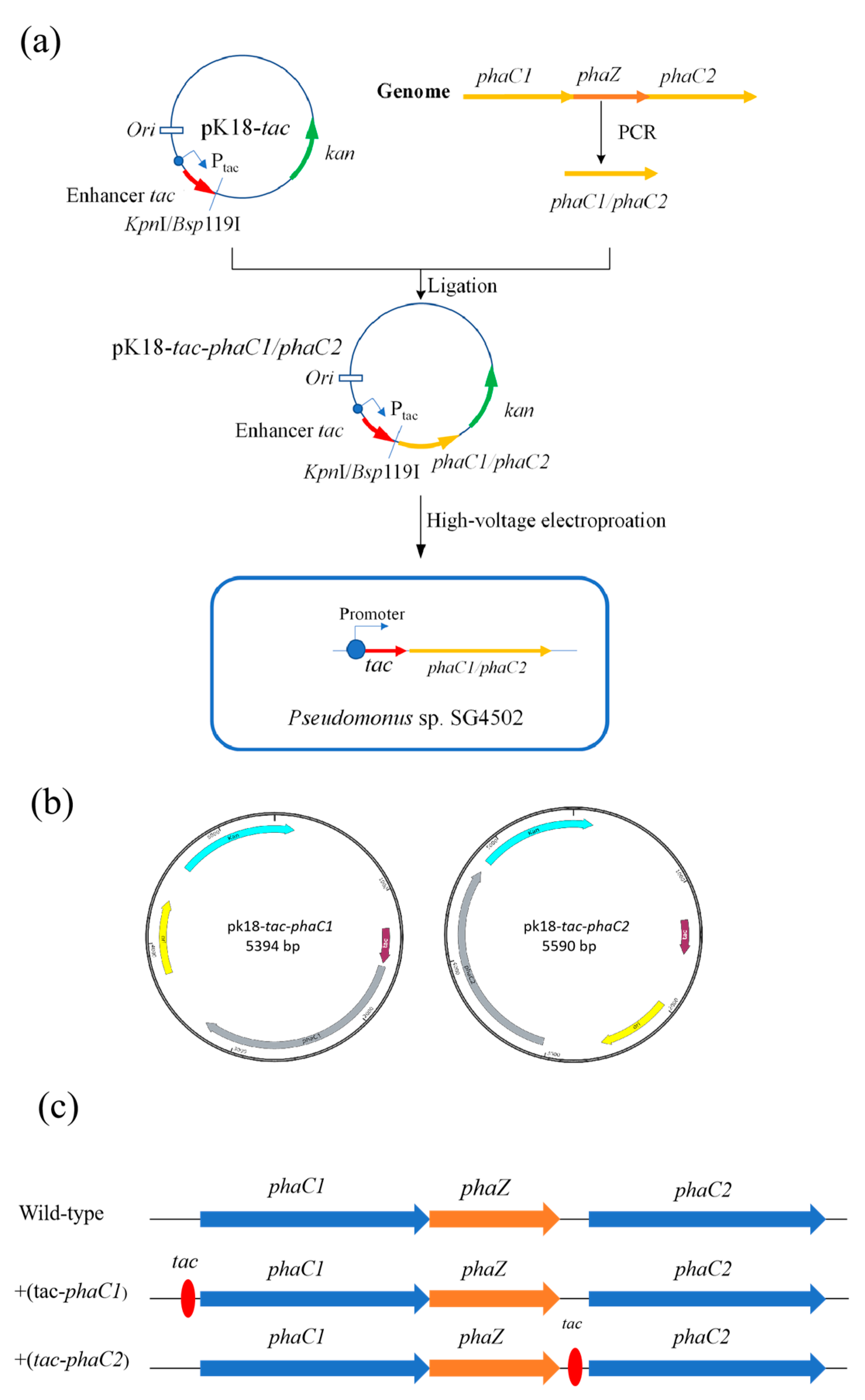
In this study, pK18 plasmid containing the strong tac enhancer was used to overexpress PHA polymerases PhaC1 and PhaC2, from SG4502. The construction process of the recombinant plasmids is shown in Figure 2a. The phaC1 and phaC2 genes were amplified by PCR from the genomic DNA of wild-type SG4502 extracted using a Bacterial Genomic DNA Extraction Kit (TIANGEN, Beijing, China). Upstream and downstream primers (listed in Table S2) of phaC1 (phaC1-F phaC1-R) and phaC2 (phaC2-F phaC2-R), respectively, were used for PCR amplification. Subsequently, the amplified fragments of the phaC1 and phaC2 genes were inserted into the vector pK18-tac at KpnI and Bsp119I enzyme digestion sites, repectively, by the NEBuilder HiFi DNA assembly method using an EasyGeno Rapid Recombinant Cloning Kit (TIANGEN, Beijing, China). The relative position of phaC1/phaC2 and tac in pK18-tac-phaC1 and pK18-tac-phaC2 are shown in Figure 2b. The position of the tac promoter is presented in Figure 2c. In the wild-type, the phaC1 and phaZ genes were closely connected, and then there was a spacer sequence between phaZ and phaC2. The tac promoter was in the upstream of phaC1 and phaC2 in +(tac-phaC1) and +(tac-phaC2), respectively. The recombinant plasmids were transferred into E. coli DH5α using the heat shock method for plasmid replication, and the plasmid was then extracted from the E. coli DH5α cells. Desalted and dried recombinant plasmid (5 μL) extracted from E. coli DH5α in the previous step was added into 100 μL of competent wild-type SG4502 cells. The mixture was transferred into an electric shock cup, in which it was treated with a voltage of 1900 V and a shock time of 5 ms. Moreover, the shock resistance was 200 Ω after the tube was placed on ice for 20 min [30]. The bacterial liquid was collected by centrifugation and then resuspended and smeared on a plate containing medium with different antibiotics (Kan, 50 mg·mL−1), and the plate was incubated at 37 °C until colonies were observed. After that, the recombinant strains containing tac-phaC1 and tac-phaC2 were screened by PCR using q-phaC1-F, q-phaC1-R, q-phaC2-F and q-phaC2-R primers. Finally, the recombinant plasmids were sequenced by Jilin KuMei company (Changchun, China).
2.3.3. Fermentation of Recombinant Strains and Extraction of Products
Wild-type and recombinant strains were inoculated in 100 mL of NR medium and cultured overnight to prepare a seed culture. The seed culture was inoculated into 100 mL of MS medium containing 1% sodium octanoate and cultured at 45 °C and 180 rpm for 72 h. Three parallel experiments were conducted for each strain. The bacterial cells were collected by centrifugation at 4 °C at 5000 rpm for 10 min and then washed thrice with distilled water. The CDW of the bacteria was weighed after being freeze-dried (FD-1B-80). Then, PHA was extracted using the chloroform-methanol precipitation method, as has been described elsewhere [31].
2.3.4. Nile Red Fluorescent Staining of Recombinant Strains
Nile red is a lipophilic fluorescent dye that quickly, sensitively and reliably detects lipid components in cells; it emits a fluorescent signal after binding to lipid substances such as wax esters, triacylglycerols and various fatty acids [32]. To macroscopically investigate the accumulation of PHA in different strains, the cellular contents of PHA in the culture described in Section 2.3.3 were tracked at different time points (6, 12, 24, 36, 48 and 72 h) using Nile red fluorescence staining. At each time point, 1 mL of bacterial liquid was withdrawn, centrifuged and then washed twice with distilled water. Next, the bacterial cells were resuspended in ultrapure water (1.5 mL) and then mixed with 3 μL of Nile red solution. The mixture was incubated at 37 °C in the dark for 15 min, and its fluorescence emission at 530 nm was determined using a Biotek’s fluorescence assay microplate reader to initially determine the PHA synthesis ability of wild-type SG4502, ∆phaZ mutant, recombinant +(tac-phaC1) and +(tac-phaC2) strains.
2.3.5. RT-qPCR of phaC1, phaC2 and phaZ Genes in Different Strains
The strains were cultured in 50 mL of NB medium at 45 °C and 180 rpm for 72 h. Then, the bacteria were collected by centrifugation at 12,000 rpm at 4 °C for 2 min and then diluted to 1 × 109 CFU/mL. The total RNA of the four strains was extracted using a cultured cell/bacterial total RNA extraction kit (TIANGEN, Beijing, China) and used as a template in the reverse transcription reaction to synthesize cDNA. The cDNA was used in the real-time measurement of the transcription levels of the phaC1, phaC2 and phaZ genes using q-phaC1-F/R, q-phaC2-F/R and q-phaZ-F/R primer pairs, respectively (Table S2).
RT-qPCR was performed using Super Real Fluorescence Quantity Premix Reagent Color Edition (TIANGEN, Beijing, China) on a Mastercycler ep realplex real-time PCR system (Eppendorf, Hamburg, Germany). RT-qPCR conditions were as follows: pre-denaturation at 95 °C for 15 min, followed by denaturation at 95 °C for 10 s and extension at 63.8 °C for 32 s for a total of 40 cycles. Each assay was performed in triplicate. Moreover, the relative gene expression level of the recombinant strains was normalized to that of the wild-type strain, and the calculations were carried out using the 2−ΔΔCt method against the 16S rRNA gene, which was an internal reference.
2.3.6. Compositional and Structural Analysis of Accumulated Products
About 5 to 10 mg of the extracted product was dissolved in 0.5 mL of deuterated chloroform. 1H-NMR spectroscopy of polymer PHA was conducted on a Bruker MSL 400 spectrometer (400 MHz) at 90 °C, a pulse of 4 ms, a spectral width of 3000 Hz and a repetition rate of 4 s.
20 mg of purified PHA product was added into 10 mL of deuterated chloroform and let stand at room temperature for more than 12 h until thoroughly mixed. After being filtered through a 0.22 μm membrane, the sample was subjected to GPC detection using a Shimadzu gel chromatograph at 30 °C. HPLC grade chloroform was used as the mobile phase, and polystyrene was used as the standard sample. The flow rate was 1 mL·min−1, and the injection volume was 50 μL.
2.3.7. Thermal Property Analysis
The melting point and the glass transition temperature of the PHA product were detected on a differential scanning calorimeter using a sample with a mass of about 20 mg. Nitrogen was used as a protective carrier gas, of which the flow rate was set at 50 mL·min−1. The sample was reheated from −50 °C to 220 °C at a heating rate of 10 °C·min−1.
Thermogravimetric data of the PHA product were determined on a synchronous thermal analyzer STA449C (NETZSCH, Bayern, Germany) using about 20 mg of sample. Nitrogen was used as a protective carrier gas; its flow rate was set at 10 mL·min−1. The temperature was increased from 50 °C to 350 °C at a rate of 10 °C·min−1.
3. Results and Discussion
3.1. Construction and Identification of the Knockout Strain ∆phaZ
Firstly, to construct the recombinant strain (∆phaZ), the C1ZC2 gene was amplified by PCR. The results from electrophoresis (Figure S1) showed that there was a clear band at around 2000 bp, which corresponds to the theoretical size (2185 bp). The amplified C1ZC2 gene fragment was digested by the HindIII and EcoRI sites and then inserted into the pUC19 plasmid linearized by the same restriction enzymes, from which the recombinant plasmid pUC19-C1ZC2 was generated, as can be seen in Figure S2. In correspondence with the expected size, the size of the recombinant plasmid was about 4700 bp, indicating that the recombination was successful.
Furthermore, the smr gene was inserted into the phaZ gene to prevent its transcription. The electrophoresis results (Figure S3) of the smr gene cloned from pCDFDuet-1 plasmid showed a band at about 1025 bp, which corresponds to the theoretical size (1000 bp). The cloned smr gene was inserted into the pUC19-C1ZC2 plasmid to construct a pUC19-C1ZC2-smr recombinant plasmid using the SacII enzyme. The electrophoresis (Figure S4) of enzyme digestion products of pUC19-C1ZC2-smr plasmid showed two obvious bands: one was at about 3500 bp, and the other was near 1900 bp. These bands were in line with the theoretical sizes of the plasmid cleaved by the enzyme. Moreover, the sequencing results demonstrated that the similarity between the recombinant plasmid sequence and the ideal sequence was 100% (Figure S5).
The constructed pUC19-C1ZC2-smr was transferred into competent wild-type SG4502 to generate the phaZ-knockout recombinant strain and screened based on Amp and Smr double resistance. PCR results showed that the smr gene could be amplified from the ∆phaZ but not from the wild-type strain (Figure 3). Thus, these results showed that the deletion mutant ∆phaZ was successfully constructed.
3.2. Construction and Identification of Recombinant Strains +(tac-phaC1) and +(tac-phaC2)
PCR amplification results of phaC1 and phaC2 showed two obvious bands at approximately 1700 bp, both of which were consistent with the theoretical sizes of both phaC1 and phaC2 (Figure S6). The recombinant plasmid containing the target gene phaC1/phaC2 was obtained by ligating with pK-18 vector. After phaC1/phaC2 was verified by colony growth, PCR amplification (Figure S7) and sequence similarity (Figure S8a,b), the recombinant plasmid pK18-phaC1/phaC2 was electroporated into competent wild-type SG4502 cells, and the recombinant strains were screened through Kanamycin resistance. The electrophoresis results indicated that the recombinant strains containing the tac enhancer +(tac-phaC1)/+(tac-phaC2) had been successfully constructed (Figure S9).
3.3. Product Accumulation in Recombinant Strains
In order to evaluate the PHA accumulation capacity of the three recombinant strains, cells were stained with Nile red at different time points during cultivation (Figure 4a) [33]. As time went on, the magnitude of fluorescence signal could be ranked as follows: +(tac-phaC2) > ∆phaZ > wild-type SG4502 > +(tac-phaC1). The results above indicate that under the same conditions, the recombinant strains +(tac-phaC2) and ∆phaZ have a better ability to synthesize PHA than wild-type SG4502. Similarly, both the CDW and the product yield of the three strains were compared (Figure 4b). The CDW of +(tac-phaC1), +(tac-phaC2), ∆phaZ and wild-type SG4502 was 0.57, 1.83, 1.44 and 1.18 g/L, respectively. This result showed that the growth rate of all genetically engineered bacteria was higher than that of the wild-type, and the genetic engineering had no effect on the growth of the strains. Additionally, among all strains, the +(tac-phaC2) strain had the highest growth rate. The product yield of +(tac-phaC1), +(tac-phaC2), ∆phaZ, and wild-type SG4502 was 0.013, 0.04, 0.032 and 0.026 g/g, respectively. The products yield was 50.0% lower in +(tac-phaC1) and 53.8% and 23.1% higher in +(tac-phaC2) and ∆phaZ than the wild-type. The product yield was consistent with the results of CDW and Nile red fluorescence staining. When comparing with the wild type, +(tac-phaC2) showed a better proliferation ability and PHA synthesis ability than +(tac-phaC1). This was probably because the tac position in the PHA synthase gene cluster had an influence on gene transcription and protein expression. Additionally, the ∆phaZ strain had a higher production yield than the wild type, which could avoid the degradation of PHA synthesized during the whole culture period. To explain the reasons for the different product yields from different recombinant strains, the transcriptional levels of phaC1, phaZ and phaC2 genes in three strains were investigated.
3.4. Transcription Level of phaC1, phaZ, and phaC2 Genes in Recombinant Trains
To explore why the product yields were different, the transcription levels of the phaC1, phaC2 and phaZ genes in the three recombinant strains were investigated by RT-qPCR (Figure 5). Compared with that of wild-type SG4502, the relative transcription level of the phaC1 and phaZ genes in +(tac-phaC1) was higher, while that of the phaC2 gene was slightly lower. For +(tac-phaC2), the relative transcription level of phaC1 and phaC2 was considerably higher, while that of phaZ was lower, compared to that of wild-type SG4502. These results suggest that the insertion position of tac can directly affect the transcription level of genes related to PHA synthesis. When the tac enhancer was inserted at the upstream of phaC1, the transcription level of phaZ (located downstream of phaC1 in the same gene cluster) and phaC1 significantly increased. At the same time, the transcription level of phaC2 located at the downstream of phaZ in the same gene cluster was not more impacted due to its long distance from tac. The transcription level of a gene directly affects its protein expression level. Considering the transcription levels of PHA synthase genes in +(tac-phaC1), the expression levels of PhaC1 and PhaZ proteins should be significantly increased, while that of PhaC2 protein should not drastically change; this can disrupt the balance between the synthesis and degradation of products, causing the product yield to decrease. Our results were consistent with those from a previous study in which the overexpression of phaC caused the expression of phaZ to be higher and the PHA yield to be lower compared with those of the wild type [34,35]. On the other hand, in +(tac-phaC2), the tac enhancer located upstream of phaC2 at a distance from phaZ (which was located between phaC1 and phaC2) enhanced the transcription level of phaC1 and phaC2 but decreased the transcription level of phaZ. Higher transcription levels of phaC1 and phaC2 and a lower transcription level of phaZ can directly affect the expression level of the corresponding proteins, resulting in an increase in the product yield in +(tac-phaC2) compared to that in the wild-type. In brief, the tac enhancer can influence the transcription level of phaC genes and improve the yield of mcl-PHA in genetically engineered bacteria, and the location and distance of tac from the target genes in the same gene cluster play important roles in the change.
Finally, regarding knockout strains ∆phaZ, the transcription level of the phaC1 gene was not affected, while phaC2 was significantly increased and phaZ was almost not transcribed. During the culture process of ∆phaZ, the low expressed PhaZ should greatly reduce the decomposition ability of synthesized mcl-PHA, coupled with the high expression of PhaC2, resulting in a significant increase in the product yield. These results showed that a high transcription level of phaC2 is crucial for mcl-PHA synthesis in wild-type SG4502. Furthermore, in SG4502, PhaC1 was active for short chain fatty acids such as 3HB, 2HB and synthesized scl-PHA [13]. For wild-type SG4502, PHA with different properties could be an effective product by the tendentious change in the transcription or expression level of phaC1/phaC2 based on different substrate specificity using the molecular method.
3.5. Structure, Composition, and Molecular Weight of mcl-PHA
The structure, composition and molecular weight of products synthesized from wild-type SG4502, +(tac-phaC1), +(tac-phaC2) and ∆phaZ were analyzed by 1H-NMR and GPC.
3.5.1. 1H-NMR
The 1H-NMR results for the four samples are shown in Figure 6. The peaks centered at 0.86, 1.28, 1.50, 2.50 and 5.11 ppm corresponded to CH3 (a1, a2, a3), (CH2)n (b1, b2, b3), CH2 (branched chain) (c1, c2, c3), CH2 (main chain) (e1, e2, e3) and CH (d1, d2, d3) of the three components (3HO, 3HD and 3HDD) in the polymers, respectively [11]. This result revealed that the transcription levels of phaC1, phaC2 and phaZ had no impact on the component of monomer units in mcl-PHA synthesized.
The 1H-NMR results indicated that the polymer synthesized by recombinant bacteria using sodium octanoate as the sole carbon source contained the same monomers as wild-type SG4502. Compared to the wild type, the production of copolymers synthesized by +(tac-phaC2) increased by 53.8%, while +(tac-phaC1) decreased by 50.0%. The reason for this is attributed to the substrate specificity of the phaC1 and phaC2 genes. phaC1 has activity on short chain fatty acids, while phaC2 has activity on medium and long chain fatty acids [36]. Using sodium octanoate as the sole carbon source, enhanced transcription of phaC2 leads to an increase in protein expression, which in turn leads to an increase in polymer production synthesized by +(tac-phaC2). However, +(tac-phaC1) with higher transcription of phaC1 and phaZ showed a sharp decrease in the yield of the synthesized copolymer because of the low substrate specificity of PhaC for sodium octanoate. In addition, ∆phaZ are similar to +(tac phaC2); the deletion of phaZ gene leads to a significant increase in phaC2 transcription, resulting in a significant increase in polymer production [13].
3.5.2. GPC
The molecular weight distribution of the synthesized mcl-PHA was determined by GPC (Table 1). The Mn of mcl-PHAs from the +(tac-phaC1), +(tac-phaC2) and ∆phaZ strains was higher than that of the wild-type strain. Theoretically, the PDI of molecular weight indicates the homogeneity of the polymer and is the factor directly affecting its properties. In this study, the PDI of mcl-PHAs from recombinant strains was one-half lower than that of mcl-PHAs from the wild type, indicating that mcl-PHAs from the former were more evenly distributed than those from the latter, which is consistent with the molecular weight PDI of similar polymers reported previously [4,10]. Thus, we may conclude that the change in PDI of mcl-PHAs from three recombinant strains might have a positive impact on their thermal properties.
3.6. Thermal Property of mcl-PHA
3.6.1. DSC
To investigate the influence of PDI on the thermal properties of mcl-PHAs, DSC and TGA were carried out. The melting temperature (Tm) and glass transition temperature (Tg) of mcl-PHAs from +(tac-phaC1), +(tac-phaC2), ∆phaZ and wild-type SG4502 were 62.73 °C and −34.08 °C (Figure 7a), 64.83 °C and −40.90 °C (Figure 7b), 63.94 °C and −36.69 °C (Figure 7c) and 64.67 °C and −38.38 °C (Figure 7d), respectively. These results indicated that the Tg and Tm of mcl-PHA synthesized by the recombinant strains were similar to those of mcl-PHA synthesized by the wild-type strain, which is consistent with the findings reported in the literature [37].
3.6.2. TGA
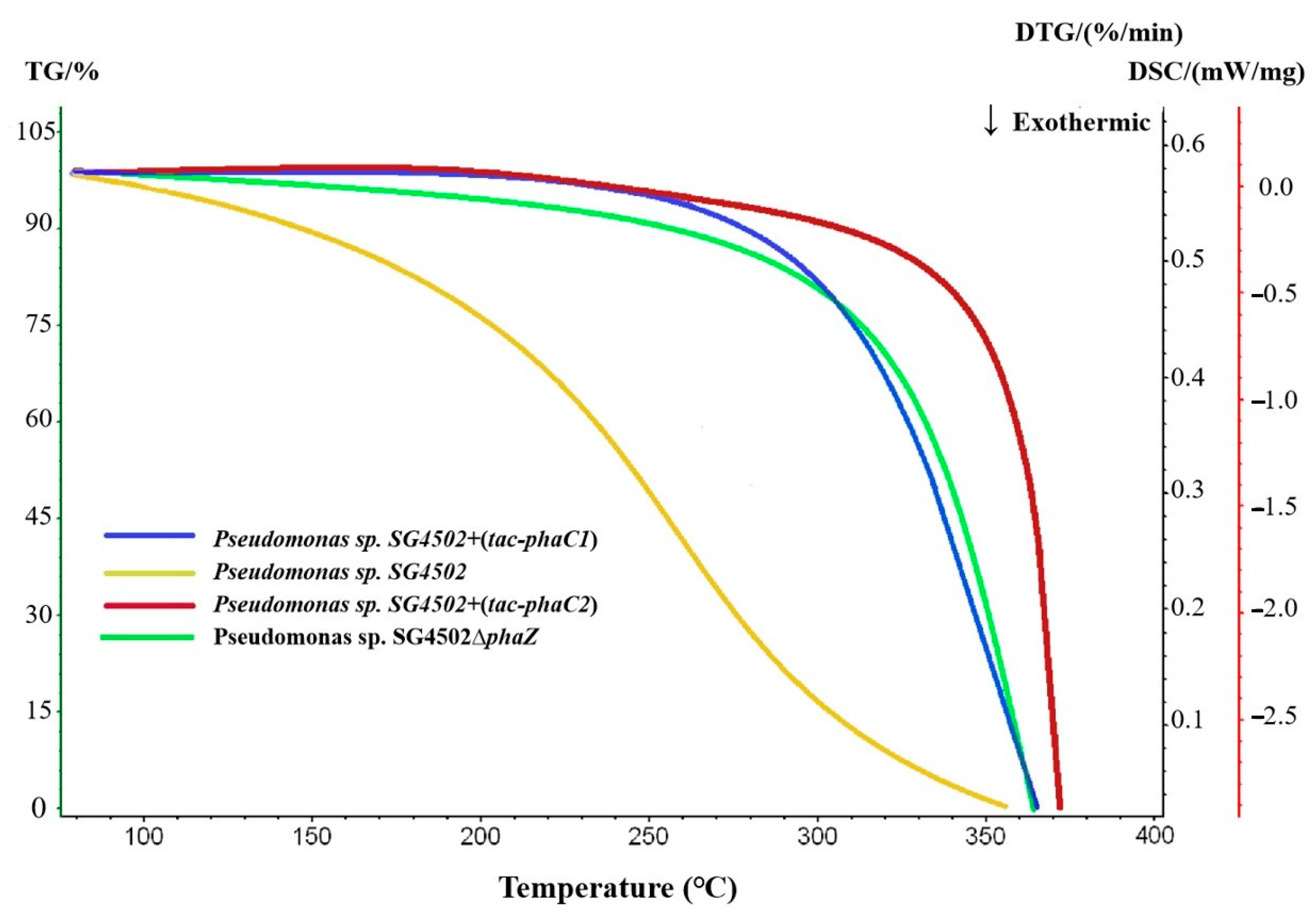
As can be seen in Figure 8, the maximum decomposition temperature of PHA from wild-type SG4502, +(tac-phaC1), +(tac-phaC2) and ∆phaZ was 356.7 °C, 365.1 °C, 371.4 °C and 366.8℃, respectively. Additionally, the decomposition temperature of mcl-PHA synthesized by +(tac-phaC1), +(tac-phaC2) and ∆phaZ was 8.4 °C, 14.7 °C and 10.1 °C, respectively, higher than that of the wild-type strain. Moreover, the decomposition temperature of mcl-PHA from the recombinant strains increased throughout the assay. The increase in thermal stability may be due to the irregular chain length of the copolymer, which could result in a high degree of crystallization of side chains; consequently, the thermal stability of the polymer improved [38]. Combined with the results on molecular weights, the PDI of mcl-PHA from +(tac-phaC2) was lower compared with that of mcl-PHA from wild-type SG4502 and +(tac-phaC1). Thus, it can be inferred that the polymer synthesized by the recombinant strains is more evenly distributed and more stable than that synthesized by the wild-type strain.
4. Conclusions
In sum, the recombinant strains of SG4502, +(tac-phaC2) and ∆phaZ could efficiently improve the yield of mcl-PHA. Compared with the phaZ gene deletion method, the tac insertion method is more efficient in synthesizing products. mcl-PHAs synthesized by +(tac-phaC1)/+(tac-phaC2) and ∆phaZ contain 3HO, 3HD and 3HDD monomer units that are comparable to those from wild-type SG4502. In addition, the PDI of mcl-PHAs from the three recombinant strains was lower than that of mcl-PHAs from the wild-type strain. The Tm and Tg of mcl-PHAs synthesized by all the recombinant strains were within the known Tm and Tg ranges of mcl-PHAs. The decomposition temperature of mcl-PHAs from the recombinant strains was higher than that of mcl-PHAs from the wild-type strain, indicating that they have higher thermal stability. Finally, the tac enhancer insertion method is a simpler and more flexible method for regulating the transcription and expression of the target gene compared to the gene knockout method. Together, this study presents a strategy for producing genetically engineered bacteria that can highly efficiently produce mcl-PHA.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/polym15102290/s1, Figure S1: Electrophoretic results of target fragment C1ZC2. M: DNA Marker GsDL10001, 1, 2: PCR product of C1ZC2 gene; Figure S2: Electrophoretic results of recombinant plasmid pUC19-C1ZC2; Figure S3: PCR electrophoresis results of smr; Figure S4: Double enzyme digestion electrophoresis results of recombinant plasmid pUC19-C1ZC2-smr; Figure S5: The sequencing results of recombinant plasmid pUC19-C1ZC2-smr; Figure S6: Electrophoretic target genes map; Figure S7: Electrophoresis results of PCR identification; Figure S8: Comparison of sequencing results of recombinant plasmid; Figure S9: Electrophoresis results of PCR identification; Table S1: Strains and plasmids used in this study; Table S2: Primers used in this study.
Author Contributions
L.S. and M.W. wrote the manuscript. D.Y. revised the manuscript. Y.L., H.Y. and X.H. supervised the project. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China grant number 31971252, Development and Reform Commission of Jilin Province grant number 2020C028-3, the Strategic Priority Research Program of the Chinese Academy of Sciences grant number XDA28010502 and the Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project grant number TSBICIP-CXRC-006.
Data Availability Statement
Data is contained within the article or supplementary material.
Acknowledgments
Thanks to the Key Laboratory of Wetland Ecology and Environment, Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences for its diversified technology help.
Conflicts of Interest
The authors declare no conflict of interests.
References
- Schlegel, P.H.G.; Gottschalk, G.; Von Bartha, R. Formation and utilization of poly-β-hydroxybutyric acid by kanallgas bacteria (hydrogenomonas). Nurture 1961, 4787, 463–465. [Google Scholar]
- Tsuge, T.; Hyakutake, M.; Mizuno, K. Class IV polyhydroxyalkanoate (PHA) synthases and PHA-producing Bacillus. Appl. Microbiol. Biotechnol. 2015, 99, 6231–6240. [Google Scholar] [CrossRef]
- Satoh, Y.; Tajima, K.; Nakamoto, S.; Xuerong, H.; Matsushima, T.; Ohshima, T.; Kawano, S.; Erata, T.; Dairi, T.; Munekata, M. Isolation of a thermotolerant bacterium producing medium-chain-length polyhydroxyalkanoate. J. Appl. Microbiol. 2011, 111, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Hyakutake, M.; Tomizawa, S.; Sugahara, I.; Murata, E.; Mizuno, K.; Abe, H.; Tsuge, T. Carboxy-terminal modification of polyhydroxyalkanoate (PHA) via alcoholysis reaction catalyzed by Class IV PHA synthase. Polym. Degrad. Stab. 2015, 117, 90–96. [Google Scholar] [CrossRef]
- Rehm, B.H.A. Polyester synthases: Natural catalysts for plastics. Biochem. J. 2003, 376, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Rhie, M.; Kim, H.; Joo, J.; Cho, I.; Son, J.; Jo, S.; Sohn, Y.; Baritugo, K.; Pyo, J.; et al. Metabolic engineering for the synthesis of polyesters: A 100-year journey from polyhydroxyalkanoates to non-natural microbial polyesters. Metab. Eng. 2020, 58, 47–81. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Shi, M.; Zhang, T.; Li, L.; Li, L.; Xian, M. Natural and engineered polyhydroxyalkanoate (PHA) synthase: Key enzyme in biopolyester production. Appl. Microbiol. Biotechnol. 2017, 101, 7417–7426. [Google Scholar] [CrossRef] [PubMed]
- Tajima, K.; Han, X.; Hashimoto, Y.; Satoh, Y.; Satoh, T.; Taguchi, S. In vitro synthesis of polyhydroxyalkanoates using thermostable acetyl-CoA synthetase, CoA transferase, and PHA synthase from thermotorelant bacteria. J. Biosci. Bioengineer. 2016, 122, 660–665. [Google Scholar] [CrossRef]
- Antonio, R.; Steinbuchel, A.; Rehm, B. Analysis of in vivo substrate specificity of the PHA synthase from Ralstonia eutropha: Formation of novel copolyesters in recombinant Escherichia coli. FEMS Microbiol. Lett. 2000, 182, 111–117. [Google Scholar] [CrossRef]
- Guo, W.; Duan, J.; Geng, W.; Feng, J.; Wang, S.; Song, C. Comparison of medium-chain-length polyhydroxyalkanoates synthases from Pseudomonas mendocina NK-01 with the same substrate specificity. Microbiol. Res. 2013, 168, 231–237. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, J.; Ma, Y.; Chen, G. Engineering biosynthesis of polyhydroxyalkanoates (PHA) for diversity and cost reduction. Metab. Eng. 2020, 58, 82–93. [Google Scholar] [CrossRef]
- Guimarães, T.; Araújo, E.; Hernández-Macedo, M.; López, J. Polyhydroxyalkanoates: Biosynthesis from alternative carbon sources and analytic methods: A short review. J. Polym. Environ. 2022, 30, 2669–2684. [Google Scholar] [CrossRef]
- De Eugenio, L.; Garci, A.; Luengo, J.; Sanz, J.; Roma, N.; Garci, A.; Prieto, M. Biochemical evidence that phaZ gene encodes a specific intracellular medium chain length polyhydroxyalkanoate depolymerase in Pseudomonas putida KT2442: Characterization of a paradigmatic enzyme. J. Biol. Chem. 2007, 282, 4951–4962. [Google Scholar] [CrossRef] [PubMed]
- Pappalardo, F.; Fragala, M.; Mineo, P.; Damigella, A.; Catara, A.; Palmeri, R.; Rescifina, A. Production of filmable medium-chain-length polyhydroxyalkanoates produced from glycerol by Pseudomonas mediterranea. Int. J. Biol. Macromol. 2014, 65, 89–96. [Google Scholar] [CrossRef]
- Chavan, S.; Yadav, B.; Tyagi, R.D.; Drogui, P. A review on production of polyhydroxyalkanoate (PHA) biopolyesters by thermophilic microbes using waste feedstocks. Bioresour. Technol. 2021, 341, 125900. [Google Scholar] [CrossRef]
- Oliveira-Filho, E.R.; Guaman, L.P.; Mendonca, T.T.; Long, P.F.; Taciro, M.K.; Gomez, J.G.C.; Silva, L.F. Production of polyhydroxyalkanoates copolymers by recombinant pseudomonas in plasmid- and antibiotic-free cultures. J. Mol. Microbiol. Biotechnol. 2018, 28, 225–235. [Google Scholar] [CrossRef]
- De Boer, H.; Comstock, L.; Vasser, M. The tac promoter: A functional hybrid derived from the trp and lac promoters. Biochemistry 1983, 80, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Lozano Terol, G.; Gallego-Jara, J.; Sola Martinez, R.; Martinez Vivancos, A.; Canovas Diaz, M.; De Diego Puente, T. Impact of the expression system on recombinant protein production in Escherichia coli BL21. Front. Microbiol. 2021, 12, 682001–682013. [Google Scholar] [CrossRef]
- Lehmeier, B.; Amann, E. Tac promoter vectors incorporating the bacteriophage T7 gene 10 translational enhancer sequence for improved expression of cloned genes in Escherichia coli. J. Biotechnol. 1922, 23, 153–165. [Google Scholar] [CrossRef]
- Zhang, B.; Ren, L.; Yu, M.; Zhou, Y.; Ye, B. Enhanced l-ornithine production by systematic manipulation of l-ornithine metabolism in engineered Corynebacterium glutamicum S9114. Bioresour. Technol. 2018, 250, 60–68. [Google Scholar] [CrossRef]
- Fukui, T.; Ohsawa, K.; Mifune, J.; Orita, I.; Nakamura, S. Evaluation of promoters for gene expression in polyhydroxyalkanoate-producing Cupriavidus necator H16. Appl. Microbiol. Biotechnol. 2011, 89, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Muneer, F.; Rasul, I.; Azeem, F.; Siddique, M.; Zubair, M.; Nadeem, H. Microbial polyhydroxyalkanoates (PHAs): Efficient replacement of synthetic polymers. J. Polym. Environ. 2020, 28, 2301–2323. [Google Scholar] [CrossRef]
- Bugnicourt, E.; Cinelli, P.; Lazzeri, A.; Alvarez, V. Polyhydroxyalkanoate (PHA): Review of synthesis, characteristics, processing and potential applications in packaging. Express Polym. Lett. 2014, 8, 791–808. [Google Scholar] [CrossRef]
- Chen, G.; Chen, X.; Wu, F.; Chen, J. Polyhydroxyalkanoates (PHA) toward cost competitiveness and functionality. Adv. Ind. Eng. Polym. Res. 2020, 3, 1–7. [Google Scholar] [CrossRef]
- Pang, X.; Cui, W.; Liu, L.; Zhang, S.; Lv, J. Gene knockout and overexpression analysis revealed the role of N-acetylmuramidase in autolysis of Lactobacillus delbrueckii subsp. bulgaricus Ljj-6. PLoS ONE 2014, 9, e104829. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Yuan, M.; Liu, F.; Jian, J.; Chen, G. Enhanced production of medium-chain-length polyhydroxyalkanoates (PHA) by PHA depolymerase knockout mutant of Pseudomonas putida KT2442. Bioresour. Technol. 2009, 100, 2265–2270. [Google Scholar] [CrossRef]
- Vo, M.; Ko, K.; Ramsay, B. Carbon-limited fed-batch production of medium-chain-length polyhydroxyalkanoates by a phaZ-knockout strain of Pseudomonas putida KT2440. J. Ind. Microbiol. Biotechnol. 2015, 42, 637–646. [Google Scholar] [CrossRef]
- Chen, G. Plastics Completely Synthesized by Bacteria: Polyhydroxyalkanoates; Springer: Berlin/Heidelberg, Germany, 2010; pp. 17–37. [Google Scholar]
- Wang, L.; Wang, Z.; Shen, C.; You, M.; Xiao, J.; Chen, G. Differentiation of human bone marrow mesenchymal stem cells grown in terpolyesters of 3-hydroxyalkanoates scaffolds into nerve cells. Biomaterials 2010, 31, 1691–1698. [Google Scholar] [CrossRef]
- Novikova, L.; Pettersson, J.; Brohlin, M.; Wiberg, M.; Novikov, L. Biodegradable poly-beta-hydroxybutyrate scaffold seeded with Schwann cells to promote spinal cord repair. Biomaterials 2008, 29, 1198–1206. [Google Scholar] [CrossRef]
- Williams, J.; Yoo, J.; Atala, A. Regenerative medicine approaches for tissue engineered heart valves. Princ. Regen. Med. 2019, 59, 1041–1058. [Google Scholar]
- Chen, G.; Wang, Y. Medical applications of biopolyesters polyhydroxyalkanoates. Chin. J. Polym. Sci. 2013, 31, 719–736. [Google Scholar] [CrossRef]
- Vigneswari, S.; Chai, J.; Shantini, K.; Bhubalan, K.; Amirul, A. Designing novel interfaces via surface functionalization of short-chain-length polyhydroxyalkanoates. Adv. Polym. Technol. 2019, 2019, 3831251. [Google Scholar] [CrossRef]
- Arias, S.; Bassas-Galia, M.; Molinari, G.; Timmis, K. Tight coupling of polymerization and depolymerization of polyhydroxyalkanoates ensures efficient management of carbon resources in Pseudomonas putida. Microb. Biotechnol. 2013, 6, 551–563. [Google Scholar] [CrossRef]
- Anjum, A.; Zuber, M.; Zia, K.; Noreen, A.; Anjum, M.; Tabasum, S. Microbial production of polyhydroxyalkanoates (PHAs) and its copolymers: A review of recent advancements. Int. J. Biol. Macromol. 2016, 89, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.K.; Munir, R.I.; Plouffe, J. Polyhydroxyalkanoate (PHA) polymer accumulation and pha gene expression in phenazine (phz-) and pyrrolnitrin (prn-) defective mutants of Pseudomonas chlororaphis PA23. Polymers 2018, 10, 1203. [Google Scholar] [CrossRef]
- Leong, Y.; Show, P.; Ooi, C.; Ling, T.; Lan, J. Current trends in polyhydroxyalkanoates (PHAs) biosynthesis: Insights from the recombinant Escherichia coli. J. Biotechnol. 2014, 180, 52–65. [Google Scholar] [CrossRef]
- Carrasco, F.; Dionisi, D.; Martinelli, A.; Majone, M. Thermal stability of polyhydroxyalkanoates. J. Appl. Polym. Sci. 2006, 100, 2111–2121. [Google Scholar] [CrossRef]
Figure 1.
Construction process of knockout mutant ∆phaZ.

Figure 2.
Overexpression plasmid construction. (a) +(tac-phaC1)/+(tac-phaC2) plasmid construction process. (b) Maps of recombinant plasmids. (c) the position of tac enhancer in the +(tac-phaC1)/+(tac-phaC2).
Figure 2.
Overexpression plasmid construction. (a) +(tac-phaC1)/+(tac-phaC2) plasmid construction process. (b) Maps of recombinant plasmids. (c) the position of tac enhancer in the +(tac-phaC1)/+(tac-phaC2).

Figure 3.
PCR amplification of smr in wild-type SG4502 (1) and the knockout strain ∆phaZ (2).

Figure 4.
(a) Nile red fluorescent staining experiment (Results of Pseudomonas sp. SG4502, Pseudomonas sp. SG4502 + tac-phaC1, Pseudomonas sp. SG4502 + tac-phaC2 and Pseudomonas sp. SG4502∆phaZ were shown in black, red, blue, and green lines, respectively), (b) the CDW and PHA yield of wild-type SG4502 and its recombinant strains.
Figure 4.
(a) Nile red fluorescent staining experiment (Results of Pseudomonas sp. SG4502, Pseudomonas sp. SG4502 + tac-phaC1, Pseudomonas sp. SG4502 + tac-phaC2 and Pseudomonas sp. SG4502∆phaZ were shown in black, red, blue, and green lines, respectively), (b) the CDW and PHA yield of wild-type SG4502 and its recombinant strains.

Figure 5.
RT-qPCR analysis of phaC1, phaC2 and phaZ. Error bars represent standard deviations (three replicates). Results of Pseudomonas sp. SG4502, +(tac-phaC1), +(tac-phaC2) and ∆phaZ are shown in white, light grey, dark grey and black, respectively.
Figure 5.
RT-qPCR analysis of phaC1, phaC2 and phaZ. Error bars represent standard deviations (three replicates). Results of Pseudomonas sp. SG4502, +(tac-phaC1), +(tac-phaC2) and ∆phaZ are shown in white, light grey, dark grey and black, respectively.

Figure 6.
1H-NMR results of culture products of wild-type SG4502 (a), recombinant strain +(tac-phaC1) (b), +(tac-phaC2) (c) and ∆phaZ (d).
Figure 6.
1H-NMR results of culture products of wild-type SG4502 (a), recombinant strain +(tac-phaC1) (b), +(tac-phaC2) (c) and ∆phaZ (d).

Figure 7.
Differential scan heat result of (a) wild-type SG4502, (b) recombinant strain +(tac-phaC1), (c) recombinant strain +(tac-phaC2), (d) knockout strain ∆phaZ. Tm: melting temperature, Tg: glass transition temperature.
Figure 7.
Differential scan heat result of (a) wild-type SG4502, (b) recombinant strain +(tac-phaC1), (c) recombinant strain +(tac-phaC2), (d) knockout strain ∆phaZ. Tm: melting temperature, Tg: glass transition temperature.

Figure 8.
Thermogravimetric analysis results of culture products of wild-type SG4502, +(tac-phaC1), +(tac-phaC2) and ∆phaZ. (Results of wild-type SG4502, +(tac-phaC1), +(tac-phaC2) and ∆phaZ are shown in yellow, blue, red and green lines, respectively).
Figure 8.
Thermogravimetric analysis results of culture products of wild-type SG4502, +(tac-phaC1), +(tac-phaC2) and ∆phaZ. (Results of wild-type SG4502, +(tac-phaC1), +(tac-phaC2) and ∆phaZ are shown in yellow, blue, red and green lines, respectively).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
GPC results of wild-type SG4502 and recombinant strains.
| Strains | Number Average Molecular Weight (Mw, Da) | Weight Average Molecular Weight (Mn, Da) | Polydispersity Index (PDI) Mw/Mn |
|---|---|---|---|
| wild-type SG4502 | 3298 | 15,038 | 4.56 |
| +(tac-phaC1) | 4965 | 13,265 | 2.67 |
| +(tac-phaC2) | 5480 | 13,799 | 2.52 |
| ∆phaZ | 7675 | 19,982 | 2.60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Song, L.; Wang, M.; Yu, D.; Li, Y.; Yu, H.; Han, X. Enhancing Production of Medium-Chain-Length Polyhydroxyalkanoates from Pseudomonas sp. SG4502 by tac Enhancer Insertion. Polymers 2023, 15, 2290. https://doi.org/10.3390/polym15102290
AMA Style
Song L, Wang M, Yu D, Li Y, Yu H, Han X. Enhancing Production of Medium-Chain-Length Polyhydroxyalkanoates from Pseudomonas sp. SG4502 by tac Enhancer Insertion. Polymers. 2023; 15(10):2290. https://doi.org/10.3390/polym15102290
Chicago/Turabian StyleSong, Linxin, Ming Wang, Dengbin Yu, Yu Li, Hongwen Yu, and Xuerong Han. 2023. "Enhancing Production of Medium-Chain-Length Polyhydroxyalkanoates from Pseudomonas sp. SG4502 by tac Enhancer Insertion" Polymers 15, no. 10: 2290. https://doi.org/10.3390/polym15102290
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.