
Potential Biomedical Application of Enzymatically Treated Alginate/Chitosan Hydrosols in Sponges—Biocompatible Scaffolds Inducing Chondrogenic Differentiation of Human Adipose Derived Multipotent Stromal Cells

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Enzyme/Alginate/Chitosan Hydrosols and Sponges
2.3. Measurements of Porosity
2.4. Swelling Ability
2.5. Cell Populations
2.6. The Isolation and Propagation of hASCs
2.7. Propagation of RAW264.7
2.8. Phenotypic Characterization of hASCs Using Flow Cytometry
2.9. Determination of Multipotent Properties of hASCs
2.10. Proliferation Assay of hASCs and RAW264.7 in Cultures with Alginate/Chitosan Hydrosols in Sponges
2.11. Measurement of DNA Synthesis in hASCs Cultures with Biomaterials
2.12. Examination of Morphology
2.13. Analysis of hASCs Viability in Cultures with Biomaterials
2.14. Determination of Chondrogenic Character of hASCs
2.14.1. Effectiveness of Chondrogenic Differentiation of hASCs on Alginate/Chitosan Hydrosols Sponges
2.14.2. Detection of Chondrogenic Nodules
2.14.3. Analysis of Chondrogenic Nodules Formation and Extracellular Matrix Elemental Composition
2.15. Analysis of Gene Expression: Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.16. Determination of Cytokines in Supernatants after RAW264.7 Cultures with Biomaterials
2.17. Oxidative Stress Factors in hASCs and RAW264.7 Culture
2.18. Statistical Analysis
3. Results and Discussion
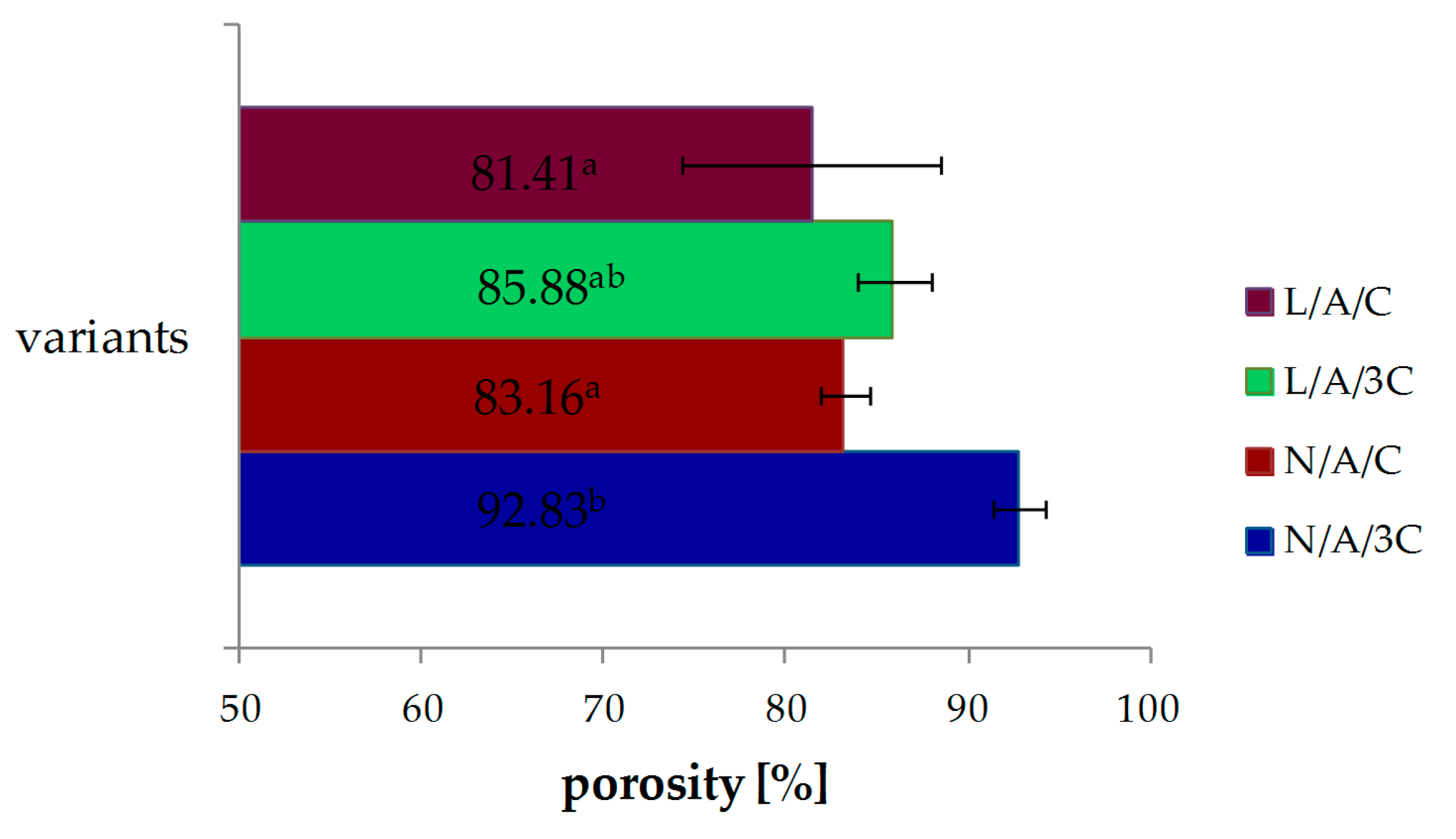
3.1. Porosity
3.2. Swelling Ability
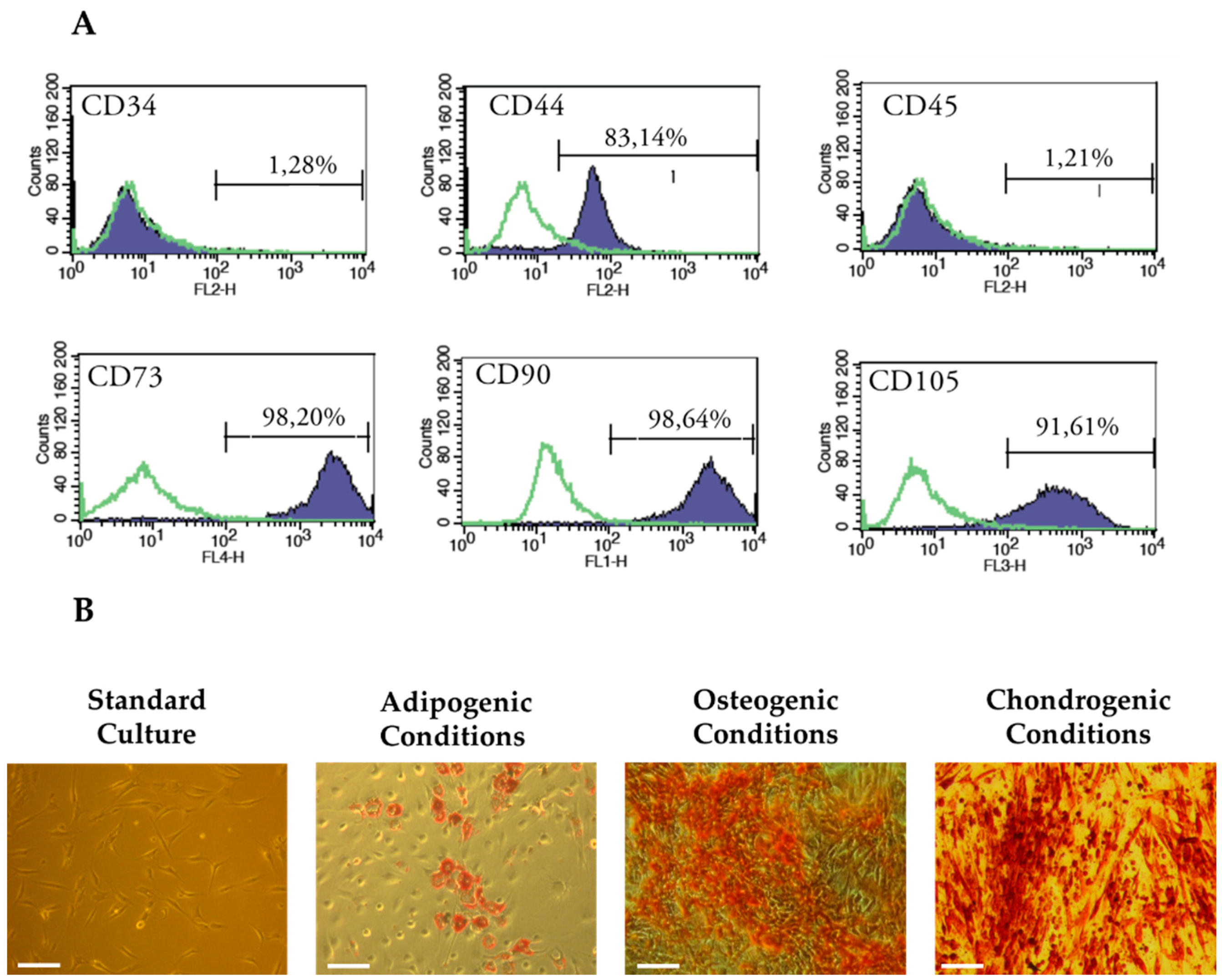
3.3. Characterization of Cells Used in the Experiment
3.4. Results of Biocompatibility Assessment
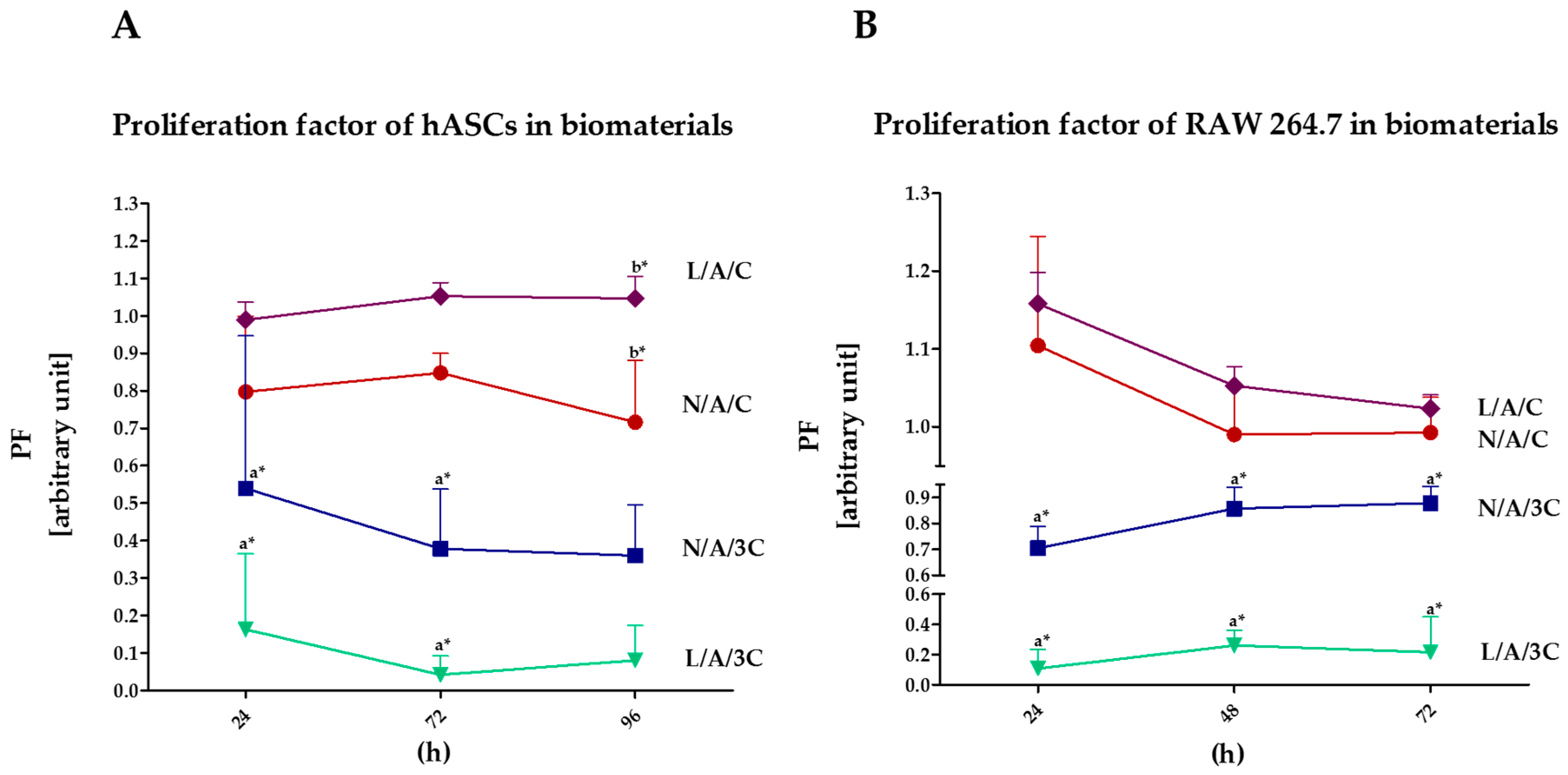
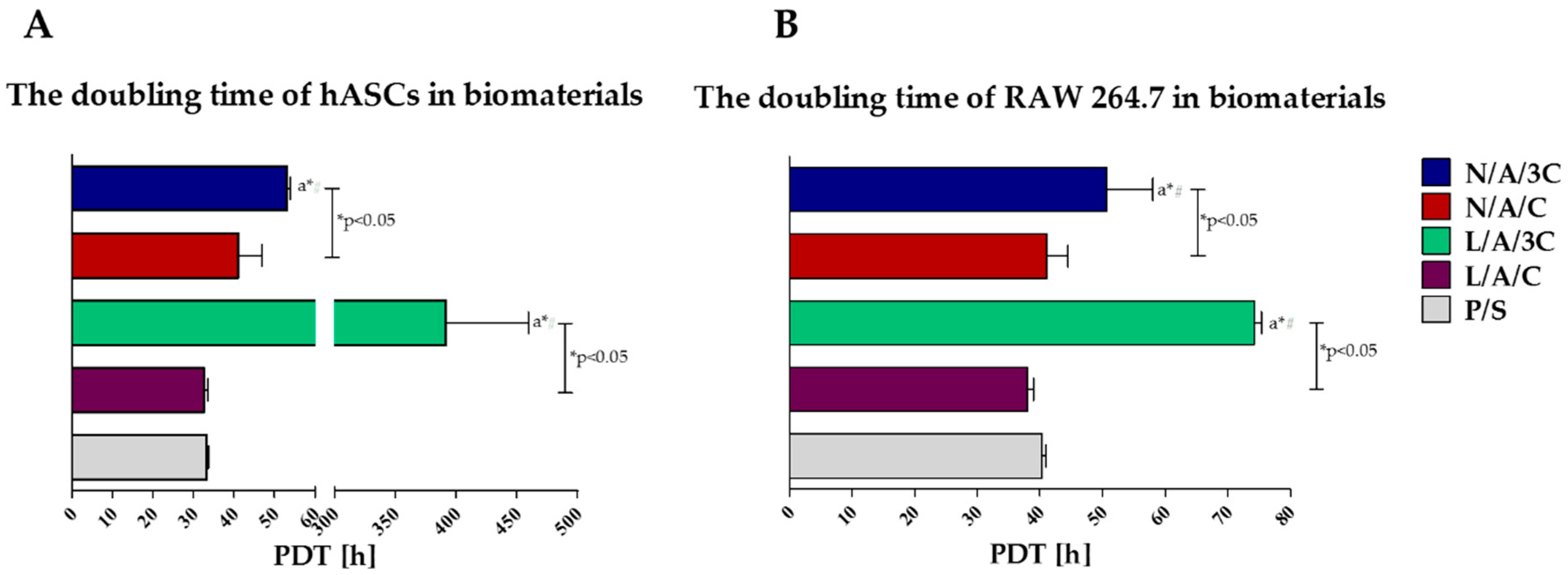
3.4.1. Proliferation of Cells in Cultures with AC Sponges
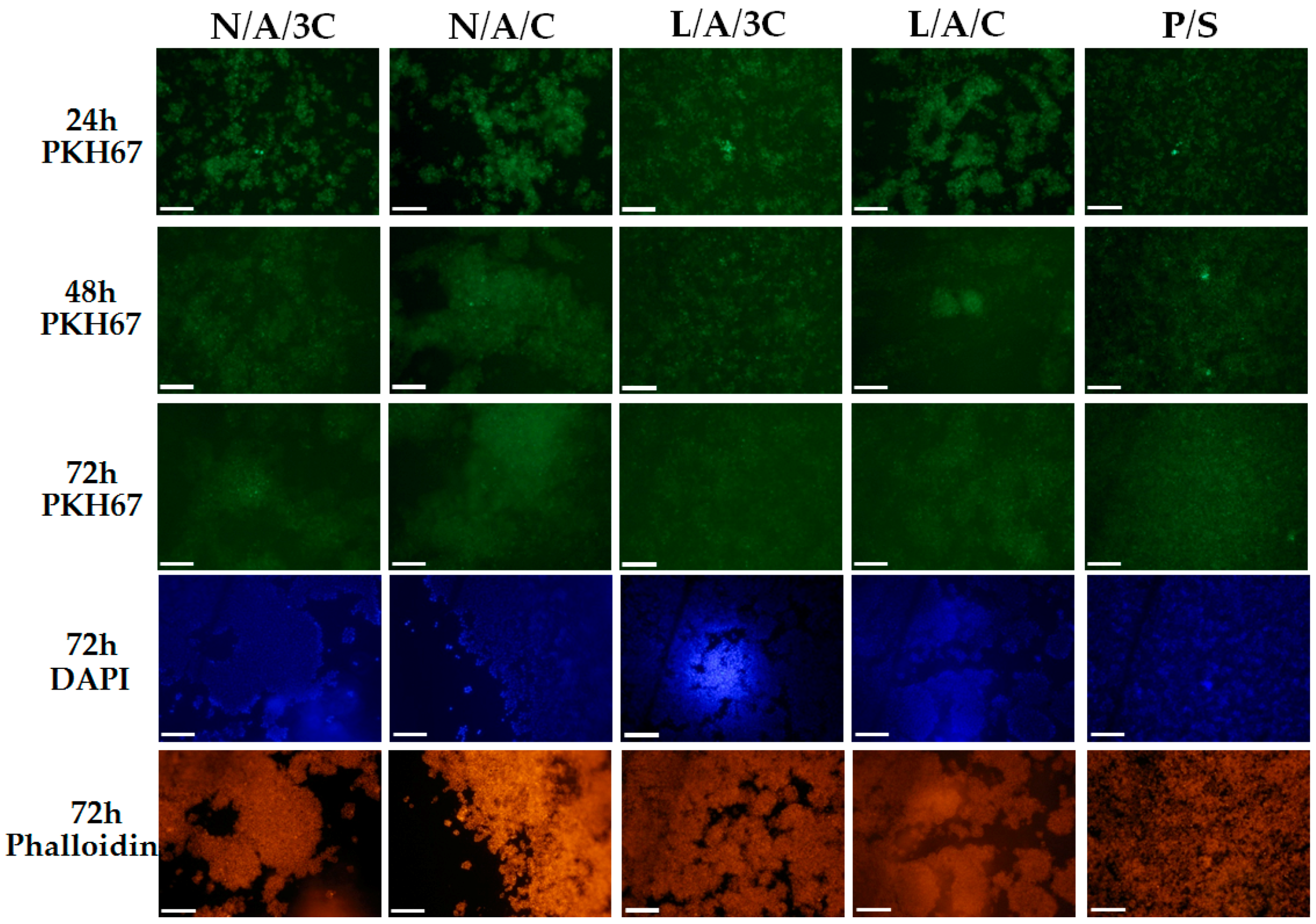
3.4.2. The Influence of AC Sponges on Cells Morphology and Growth Pattern
3.4.3. Cytotoxicity of AC Sponges
3.4.4. Biomaterial-Induced Cytokine Expression in RAW264.7 Monocyte-Macrophage Cell Line
3.4.5. Assays of Oxidative Stress Factors in hASCs and RAW264.7 Cultures with Biomaterials
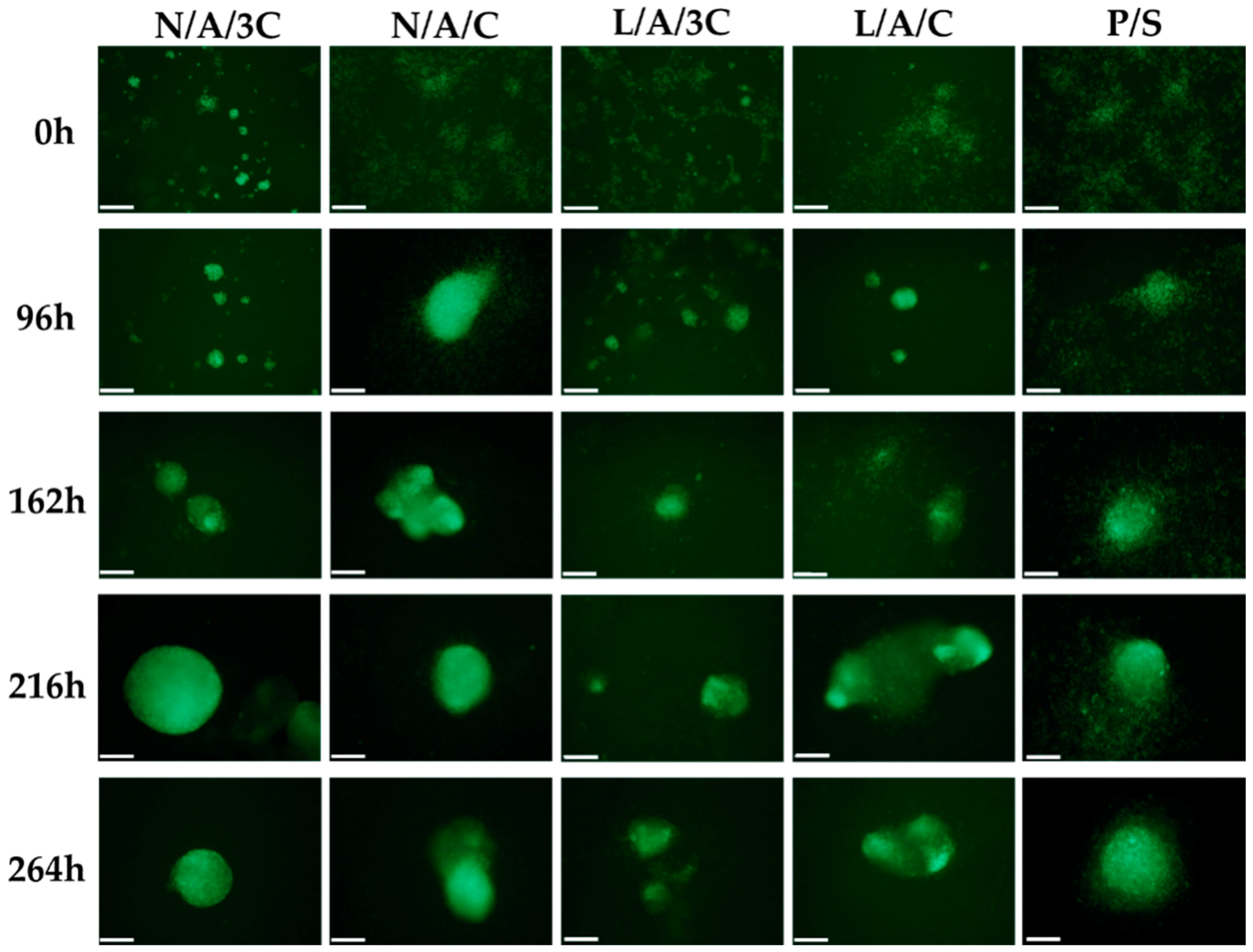
3.5. Chondrogenic Differentiation of hASCs in AC Sponges
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ajibade, D.A.; Vance, D.D.; Hare, J.M.; Kaplan, L.D.; Lesniak, B.P. Emerging Applications of Stem Cell and Regenerative Medicine to Sports Injuries. Orthop. J. Sports Med. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, K.G.; Berry, C.; Bartunek, J. Myocardial Repair and Regeneration: Bone Marrow or Cardiac Stem Cells? Mol. Ther. 2012, 20, 1102–1105. [Google Scholar] [CrossRef] [PubMed]
- Carolan, P.J.; Melton, D.A. New findings in pancreatic and intestinal endocrine development to advance regenerative medicine. Curr. Opin. Endocrinol. Diabetes Obes. 2013, 20, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dimitriou, R.; Jones, E.; McGonagle, D.; Giannoudis, P.V. Bone regeneration: Current concepts and future directions. BMC Med. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- NIAMS. Available online: http://www.niams.nih.gov/ (accessed on 20 April 2016).
- Mollon, B.; Kandel, R.; Chahal, J.; Theodoropoulos, J. The clinical status of cartilage tissue regeneration in humans. Osteoarthr. Cartil. 2013, 21, 1824–1833. [Google Scholar] [CrossRef] [PubMed]
- Savkovic, V.; Li, H.; Seon, J.-K.; Hacker, M.; Franz, S.; Simon, J.-C. Mesenchymal stem cells in cartilage regeneration. Curr. Stem Cell Res. Ther. 2014, 9, 469–488. [Google Scholar] [CrossRef] [PubMed]
- Arnhold, S.; Wenisch, S. Adipose tissue derived mesenchymal stem cells for musculoskeletal repair in veterinary medicine. Am. J. Stem Cells 2015, 4, 1–12. [Google Scholar] [PubMed]
- Bai, X.; Alt, E. Myocardial regeneration potential of adipose tissue-derived stem cells. Biochem. Biophys. Res. Commun. 2010, 401, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Marconi, S.; Castiglione, G.; Turano, E.; Bissolotti, G.; Angiari, S.; Farinazzo, A.; Constantin, G.; Bedogni, G.; Bedogni, A.; Bonetti, B. Human adipose-derived mesenchymal stem cells systemically injected promote peripheral nerve regeneration in the mouse model of sciatic crush. Tissue Eng. Part A 2012, 18, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- Marycz, K.; Toker, N.Y.; Grzesiak, J.; Wrzeszcz, K.; Golonka, P. The Therapeutic Effect of Autogenic Adipose Derived Stem Cells Combined with Autogenic Platelet Rich Plasma in Tendons Disorders in Horses in vitro and in vivo Research. J. Anim. Vet. Adv. 2012, 11, 4324–4331. [Google Scholar]
- Toker, N.Y.; Nicpoń, J.; Marycz, K.; Grzesiak, J.; ŚMieszek, A. The Advantages of Autologus Adipose Derived Mesenchymal Stem Cells (AdMSCs) over the NSAIDs Application for Degenerative Elbow Joint Disease Treatment in Dogs—Twelve Cases. Kafkas Univ. Vet. Fak. Derg. 2014, 20, 345–350. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.I.; Ye, F.; Tarantal, A.F. Comparison of growth and differentiation of fetal and adult rhesus monkey mesenchymal stem cells. Stem Cells Dev. 2006, 15, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Harris, D.T. Comparison of human mesenchymal stem cells derived from adipose and cord tissue. Cytotherapy 2013, 15, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Abumaree, M.; Al Jumah, M.; Pace, R.A.; Kalionis, B. Immunosuppressive properties of mesenchymal stem cells. Stem Cell Rev. 2012, 8, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Camussi, G.; Deregibus, M.C.; Bruno, S.; Cantaluppi, V.; Biancone, L. Exosomes/microvesicles as a mechanism of cell-to-cell communication. Kidney Int. 2010, 78, 838–848. [Google Scholar] [CrossRef] [PubMed]
- Mathivanan, S. Exosomes and shedding microvesicles are mediators of intercellular communication: How do they communicate with the target cells? J. Biotechnol. Biomater. 2012, 2, 4. [Google Scholar]
- Biancone, L.; Bruno, S.; Deregibus, M.C.; Tetta, C.; Camussi, G. Therapeutic potential of mesenchymal stem cell-derived microvesicles. Nephrol. Dial. Transplant. 2012, 27, 3037–3042. [Google Scholar] [CrossRef] [PubMed]
- Marycz, K.; Śmieszek, A.; Grzesiak, J.; Donesz-Sikorska, A.; Krzak-Roś, J. Application of bone marrow and adipose-derived mesenchymal stem cells for testing the biocompatibility of metal-based biomaterials functionalized with ascorbic acid. Biomed. Mater. 2013, 8, 065004. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Jin, X.; Cong, Y.; Liu, Y.; Fu, J. Degradable natural polymer hydrogels for articular cartilage tissue engineering. J. Chem. Technol. Biotechnol. 2013, 88, 327–339. [Google Scholar] [CrossRef]
- Iwasaki, N.; Kasahara, Y.; Yamane, S.; Igarashi, T.; Minami, A.; Nisimura, S. Chitosan-Based Hyaluronic Acid Hybrid Polymer Fibers as a Scaffold Biomaterial for Cartilage Tissue Engineering. Polymers 2010, 3, 100–113. [Google Scholar] [CrossRef]
- Pati, F.; Jang, J.; Ha, D.H.; Won Kim, S.; Rhie, J.W.; Shim, J.H.; Kim, D.H.; Cho, D.W. Printing three-dimensional tissue analogues with decellularized extracellular matrix bioink. Nat. Commun. 2014, 5, 3935. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Choi, Y.J.; Shim, J.H.; Park, J.H.; Cho, D.W. Development of a 3D cell printed structure as an alternative to autologs cartilage for auricular reconstruction. J. Biomed. Mater. Res. B Appl. Biomater. 2016. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed]
- Yamane, S.; Iwasaki, N.; Majima, T.; Funakoshi, T.; Masuko, T.; Harada, K.; Minami, A.; Monde, K.; Nishimura, S.-I. Feasibility of chitosan-based hyaluronic acid hybrid biomaterial for a novel scaffold in cartilage tissue engineering. Biomaterials 2005, 26, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Mano, J.F.; Silva, G.A.; Azevedo, H.S.; Malafaya, P.B.; Sousa, R.A.; Silva, S.S.; Boesel, L.F.; Oliveira, J.M.; Santos, T.C.; Marques, A.P.; et al. Natural origin biodegradable systems in tissue engineering and regenerative medicine: present status and some moving trends. J. R. Soc. Interface 2007, 4, 999–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghahramanpoor, M.K.; Hassani Najafabadi, S.A.; Abdouss, M.; Bagheri, F.; Baghaban Eslaminejad, M. A hydrophobically-modified alginate gel system: utility in the repair of articular cartilage defects. J. Mater. Sci. Mater. Med. 2011, 22, 2365–2375. [Google Scholar] [CrossRef] [PubMed]
- Lubiatowski, P.; Kruczynski, J.; Gradys, A.; Trzeciak, T.; Jaroszewski, J. Articular cartilage repair by means of biodegradable scaffolds. Transplant. Proc. 2006, 38, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Titan, A.L.; Stafford, M.; Zheng, C.H.; Levenston, M.E. Variations in chondrogenesis of human bone marrow-derived mesenchymal stem cells in fibrin/alginate blended hydrogels. Acta Biomater. 2012, 8, 3754–3764. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, T.; Song, K.; Yao, L.; Ge, D.; Bao, C.; Ma, X.; Cui, Z. Culture of neural stem cells in calcium alginate beads. Biotechnol. Prog. 2006, 22, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Suzuki, Y.; Tanihara, M.; Ohnishi, K.; Hashimoto, T.; Endo, K.; Nishimura, Y. Reconstruction of rat peripheral nerve gap without sutures using freeze-dried alginate gel. J. Biomed. Mater. Res. 2000, 49, 528–533. [Google Scholar] [CrossRef]
- Hashimoto, T.; Suzuki, Y.; Suzuki, K.; Nakashima, T.; Tanihara, M.; Ide, C. Review: Peripheral nerve regeneration using non-tubular alginate gel crosslinked with covalent bonds. J. Mater. Sci. Mater. Med. 2005, 16, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Szarek, D.; Marycz, K.; Lis, A.; Zawada, Z.; Tabakow, P.; Laska, J.; Jarmundowicz, W. Lizard tail spinal cord: a new experimental model of spinal cord injury without limb paralysis. FASEB J. 2016, 30, 1391–1403. [Google Scholar] [CrossRef] [PubMed]
- Marycz, K.; Meissner, J.M.; Grzesiak, J.; Śmieszek, A.; Kasprzyk, I. Sphingosine-1-Phosphate Enhance Osteogenic Activity of Multipotent Stromal Cells Cultured in Biodegradable 3D Alginate Hydrogels. J. Biomater. Tissue Eng. 2016, 6, 85–97. [Google Scholar] [CrossRef]
- Zimoch-Korzycka, A.; Kulig, D.; Jarmoluk, A.; Marycz, K.; Matuszczak, W. Study of Enzymatically Treated Alginate/Chitosan Hydrosols in Sponges Formation Process. Polymers 2016, 8, 8. [Google Scholar] [CrossRef]
- Ikeda, T.; Ikeda, K.; Yamamoto, K.; Ishizaki, H.; Yoshizawa, Y.; Yanagiguchi, K.; Yamada, S.; Hayashi, Y. Fabrication and Characteristics of Chitosan Sponge as a Tissue Engineering Scaffold. BioMed Res. Int. 2014, 2014, e786892. [Google Scholar] [CrossRef] [PubMed]
- Archana, D.; Upadhyay, L.; Tewari, R.P.; Dutta, J.; Huang, Y.B.; Dutta, P.K. Chitosan-pectin-alginate as a novel scaffold for tissue engineering applications. Indian J. Biotechnol. 2013, 12, 475–482. [Google Scholar]
- Smieszek, A.; Donesz-Sikorska, A.; Grzesiak, J.; Krzak, J.; Marycz, K. Biological effects of sol-gel derived ZrO2 and SiO2/ZrO2 coatings on stainless steel surface-In vitro model using mesenchymal stem cells. J. Biomater. Appl. 2014, 29, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Śmieszek, A.; Basińska, K.; Chrząstek, K.; Marycz, K. In Vitro and In Vivo Effects of Metformin on Osteopontin Expression in Mice Adipose-Derived Multipotent Stromal Cells and Adipose Tissue. J. Diabetes Res. 2015, 2015, e814896. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.J.; Bae, Y.K.; Kim, M.; Kwon, S.-J.; Jeon, H.B.; Choi, S.J.; Kim, S.W.; Yang, Y.S.; Oh, W.; Chang, J.W. Comparative Analysis of Human Mesenchymal Stem Cells from Bone Marrow, Adipose Tissue, and Umbilical Cord Blood as Sources of Cell Therapy. Int. J. Mol. Sci. 2013, 14, 17986–18001. [Google Scholar] [CrossRef] [PubMed]
- Śmieszek, A.; Czyrek, A.; Basinska, K.; Trynda, J.; Skaradzińska, A.; Siudzińska, A.; Marędziak, M.; Marycz, K. Effect of Metformin on Viability, Morphology, and Ultrastructure of Mouse Bone Marrow-Derived Multipotent Mesenchymal Stromal Cells and Balb/3T3 Embryonic Fibroblast Cell Line. BioMed Res. Int. 2015, 2015, 769402. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- She, Z.; Zhang, B.; Jin, C.; Feng, Q.; Xu, Y. Preparation and in vitro degradation of porous three-dimensional silk fibroin/chitosan scaffold. Polym. Degrad. Stab. 2008, 93, 1316–1322. [Google Scholar] [CrossRef]
- Kumar, P.T.S.; Srinivasan, S.; Lakshmanan, V.-K.; Tamura, H.; Nair, S.V.; Jayakumar, R. Synthesis, characterization and cytocompatibility studies of α-chitin hydrogel/nano hydroxyapatite composite scaffolds. Int. J. Biol. Macromol. 2011, 49, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Karageorgiou, V.; Kaplan, D. Porosity of 3D biomaterial scaffolds and osteogenesis. Biomaterials 2005, 26, 5474–5491. [Google Scholar] [CrossRef] [PubMed]
- Burg, K.J.L.; Porter, S.; Kellam, J.F. Biomaterial developments for bone tissue engineering. Biomaterials 2000, 21, 2347–2359. [Google Scholar] [CrossRef]
- Ma, L.; Gao, C.; Mao, Z.; Zhou, J.; Shen, J.; Hu, X.; Han, C. Collagen/chitosan porous scaffolds with improved biostability for skin tissue engineering. Biomaterials 2003, 24, 4833–4841. [Google Scholar] [CrossRef]
- Chen, Q.Z.; Harding, S.E.; Ali, N.N.; Lyon, A.R.; Boccaccini, A.R. Biomaterials in cardiac tissue engineering: Ten years of research survey. Mater. Sci. Eng. R 2008, 59, 1–37. [Google Scholar] [CrossRef]
- Zhang, Y.; Tse, C.; Rouholamin, D.; Smith, P.J. Scaffolds for tissue engineering produced by inkjet printing. Cent. Eur. J. Eng. 2012, 2, 325–335. [Google Scholar] [CrossRef]
- Griffon, D.J.; Sedighi, M.R.; Schaeffer, D.V.; Eurell, J.A.; Johnson, A.L. Chitosan scaffolds: Interconnective pore size and cartilage engineering. Acta Biomater. 2006, 2, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Lien, S.M.; Ko, L.Y.; Huang, T.J. Effect of pore size on ECM secretion and cell growth in gelatin scaffold for articular cartilage tissue engineering. Acta Biomater. 2009, 5, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.I.; Krebs, M.D.; Bonino, C.A.; Samorezov, J.E.; Khan, S.A.; Alsberg, E. Electrospun chitosan-alginate nanofibers with in situ polyelectrolyte complexation for use as tissue engineering scaffolds. Tissue Eng. Part A 2011, 17, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Khademhosseini, A.; Dehghani, F. Enhancing cell penetration and proliferation in chitosan hydrogels for tissue engineering applications. Biomaterials 2011, 32, 9719–9729. [Google Scholar] [CrossRef] [PubMed]
- Wimardani, Y.S.; Suniarti, D.F.; Freisleben, H.J.; Wanandi, S.I.; Ikeda, M.A. Cytotoxic effects of chitosan against oral cancer cell lines is molecular-weight-dependent and cell-type-specific. Int. J. Oral Res. 2012, 3, e1. [Google Scholar]
- Omar Zaki, S.S.; Ibrahim, M.N.; Katas, H. Particle Size Affects Concentration-Dependent Cytotoxicity of Chitosan Nanoparticles towards Mouse Hematopoietic Stem Cells. J. Nanotechnol. 2015, 2015, e919658. [Google Scholar] [CrossRef]
- Song, J.E.; Kim, A.R.; Lee, C.J.; Tripathy, N.; Yoon, K.H.; Lee, D.; Khang, G. Effects of purified alginate sponge on the regeneration of chondrocytes: In vitro and in vivo. J. Biomater. Sci. Polym. Ed. 2015, 26, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Kurachi, M.; Nakashima, T.; Miyajima, C.; Iwamoto, Y.; Muramatsu, T.; Yamaguchi, K.; Oda, T. Comparison of the activities of various alginates to induce TNF-α secretion in RAW264.7 cells. J. Infect. Chemother. 2005, 11, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Witsell, A.L.; Schook, L.B. Tumor necrosis factor alpha is an autocrine growth regulator during macrophage differentiation. Proc. Natl. Acad. Sci. USA 1992, 89, 4754–4758. [Google Scholar] [CrossRef] [PubMed]
- Kornicka, K.; Marycz, K.; Tomaszewski, K.A.; Marędziak, M.; Śmieszek, A. The Effect of Age on Osteogenic and Adipogenic Differentiation Potential of Human Adipose Derived Stromal Stem Cells (hASCs) and the Impact of Stress Factors in the Course of the Differentiation Process. Oxid. Med. Cell. Longev. 2015, 2015, 309169. [Google Scholar] [CrossRef] [PubMed]
- Marycz, K.; Kornicka, K.; Basinska, K.; Czyrek, A. Equine Metabolic Syndrome Affects Viability, Senescence, and Stress Factors of Equine Adipose-Derived Mesenchymal Stromal Stem Cells: New Insight into EqASCs Isolated from EMS Horses in the Context of Their Aging. Oxid. Med. Cell. Longev. 2016, 2016, 4710326. [Google Scholar] [CrossRef] [PubMed]
- Marędziak, M.; Marycz, K.; Tomaszewski, K.A.; Kornicka, K.; Henry, B.M. The Influence of Aging on the Regenerative Potential of Human Adipose Derived Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 2152435. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Chen, A.; You, H.; Li, K.; Zhang, D.; Guo, F. Proliferation and chondrogenic differentiation of CD105-positive enriched rat synovium-derived mesenchymal stem cells in three-dimensional porous scaffolds. Biomed. Mater. 2011, 6, 015006. [Google Scholar] [CrossRef] [PubMed]
- Tığlı, R.S.; Gümüşderelioğlu, M. Evaluation of alginate-chitosan semi IPNs as cartilage scaffolds. J. Mater. Sci. Mater. Med. 2008, 20, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Lewis, R.; Ferreira-Mendes, A.; Rufino, A.; Dart, C.; Barrett-Jolley, R. Potassium channels in articular chondrocytes. Channels (Austin) 2012, 6, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Parry, N.M.; Phillippo, M.; Reid, M.D.; McGaw, B.A.; Flint, D.J.; Loveridge, N. Molybdenum-induced changes in the epiphyseal growth plate. Calcif. Tissue Int. 1993, 53, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.; Wu, B.M. Biological and mechanical characterization of chitosan-alginate scaffolds for growth factor delivery and chondrogenesis. J. Biomed. Mater. Res. Part B Appl. Biomater. 2015. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme addition (E) | Polymers ratio A:C | Variants (E/A/C) |
|---|---|---|
| (U) | (v/v) | (Code) |
| N * | 1:3 | N/A/3C |
| 1:1 | N/A/C | |
| 1,000 L | 1:3 | L/A/3C |
| 1:1 | L/A/C |
| Gene | GenBank accession number | Primer sequence | Annealing temperature (°C) | Amplicon size (bp) |
|---|---|---|---|---|
| Human transcripts | ||||
| ACAN | NM_001113455.2 | F: GAGCCTGAAAACCAGACGGA | 570 | 109 |
| R: TCTCCTCTGTTGCTGTGCTG | ||||
| ADAMTS5 | NM_007038.3 | F: TATGACAAGTGCGGAGTATG | 60.8 | 182 |
| R: TTCAGGGCTAAATAGGCAGT | ||||
| Coll-1 | NM_001003090.1 | F: ACCGACCAAGAAACCACAGG | 61.1 | 226 |
| R: GCACGGAGATTCCTCCAGTT | ||||
| Coll-2 | NM_001006951.1 | F: GACAATCTGGCTCCCAAC | 60.3 | 233 |
| R: ACAGTCTTGCCCCACTTAC | ||||
| GAPDH | NM_001003142.2 | F: GATTGTCAGCAATGCCTCCT | 58.0 | 198 |
| R: GTGGAAGCAGGGATGATGTT | ||||
| Mouse transcripts | ||||
| ACTβ | NM_007393.5 | F: CGACGATGCTCCCCGGGCTGTA | 62.0 | 574 |
| R: CTCTTTGATGTCACGCACGATTTCCCTCT | ||||
| p53 | NM_011640.3 | F: AGTCACAGCACATGACGGAGG | 61.0 | 287 |
| R: GGAGTCTTCCAGTGTGATGATGG | ||||
| TNFα | NM_013693.3 | F: ACAGAAAGCATGATCCGCGA | 62.0 | 295 |
| R: CTTGGTGGTTTGCTACGACG | ||||
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimoch-Korzycka, A.; Śmieszek, A.; Jarmoluk, A.; Nowak, U.; Marycz, K. Potential Biomedical Application of Enzymatically Treated Alginate/Chitosan Hydrosols in Sponges—Biocompatible Scaffolds Inducing Chondrogenic Differentiation of Human Adipose Derived Multipotent Stromal Cells. Polymers 2016, 8, 320. https://doi.org/10.3390/polym8090320
Zimoch-Korzycka A, Śmieszek A, Jarmoluk A, Nowak U, Marycz K. Potential Biomedical Application of Enzymatically Treated Alginate/Chitosan Hydrosols in Sponges—Biocompatible Scaffolds Inducing Chondrogenic Differentiation of Human Adipose Derived Multipotent Stromal Cells. Polymers. 2016; 8(9):320. https://doi.org/10.3390/polym8090320
Chicago/Turabian StyleZimoch-Korzycka, Anna, Agnieszka Śmieszek, Andrzej Jarmoluk, Urszula Nowak, and Krzysztof Marycz. 2016. "Potential Biomedical Application of Enzymatically Treated Alginate/Chitosan Hydrosols in Sponges—Biocompatible Scaffolds Inducing Chondrogenic Differentiation of Human Adipose Derived Multipotent Stromal Cells" Polymers 8, no. 9: 320. https://doi.org/10.3390/polym8090320
APA StyleZimoch-Korzycka, A., Śmieszek, A., Jarmoluk, A., Nowak, U., & Marycz, K. (2016). Potential Biomedical Application of Enzymatically Treated Alginate/Chitosan Hydrosols in Sponges—Biocompatible Scaffolds Inducing Chondrogenic Differentiation of Human Adipose Derived Multipotent Stromal Cells. Polymers, 8(9), 320. https://doi.org/10.3390/polym8090320