

Different Drought Tolerance Strategy of Wheat Varieties in Spike Architecture

 , , , , and
, , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Field Sites Conditions
2.2. Plant Material
2.3. Statistical Data Analysis
3. Results
3.1. Effect of the Photoperiod Sensitivity on Yield Characteristics
3.1.1. Growing season 2019/2020
3.1.2. Growing Season 2020/2021
3.2. Effect of Variety on Yield Characteristics at the Stressed Site
3.2.1. Growing Season 2019/2020
3.2.2. Growing Season 2020/2021
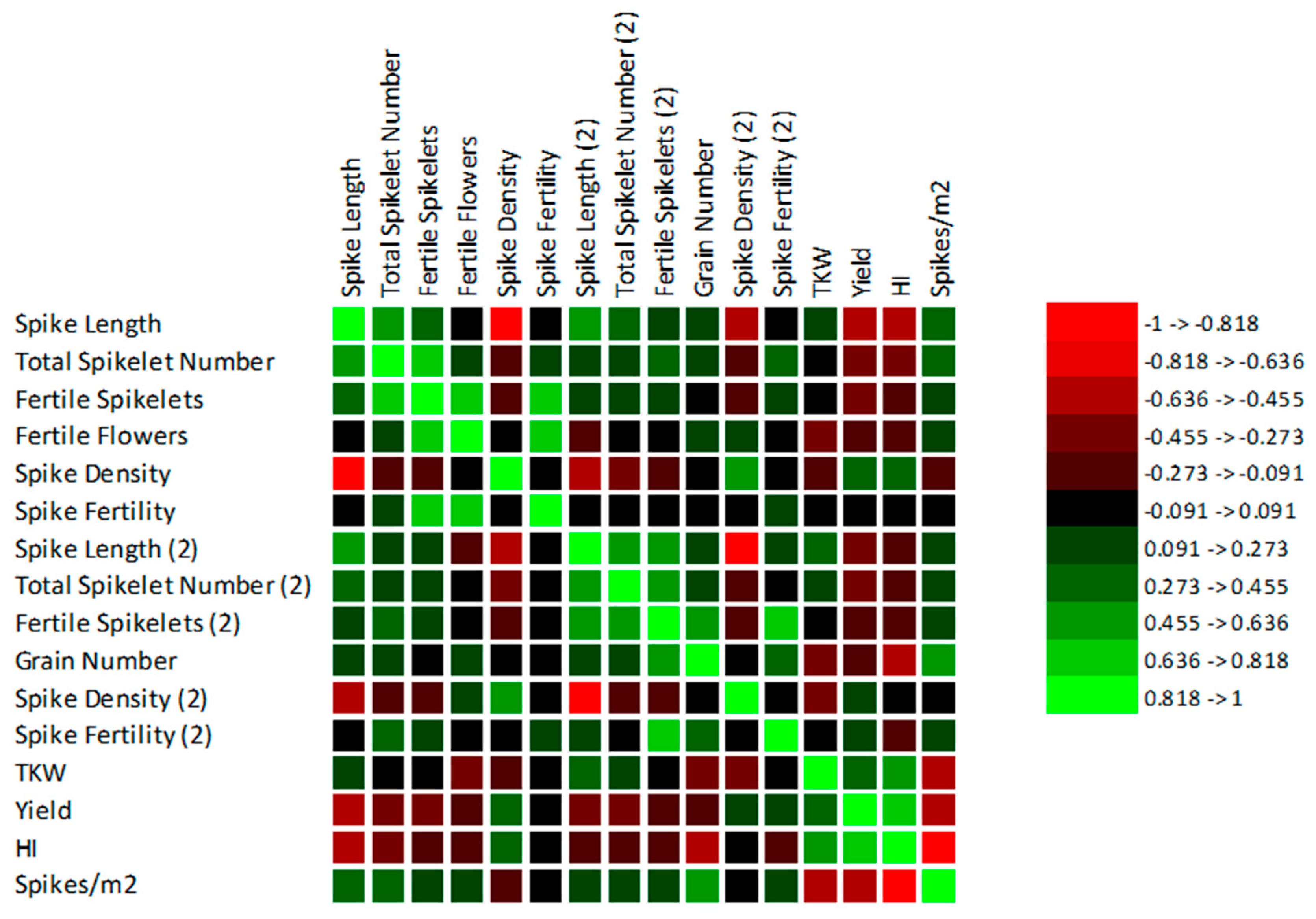
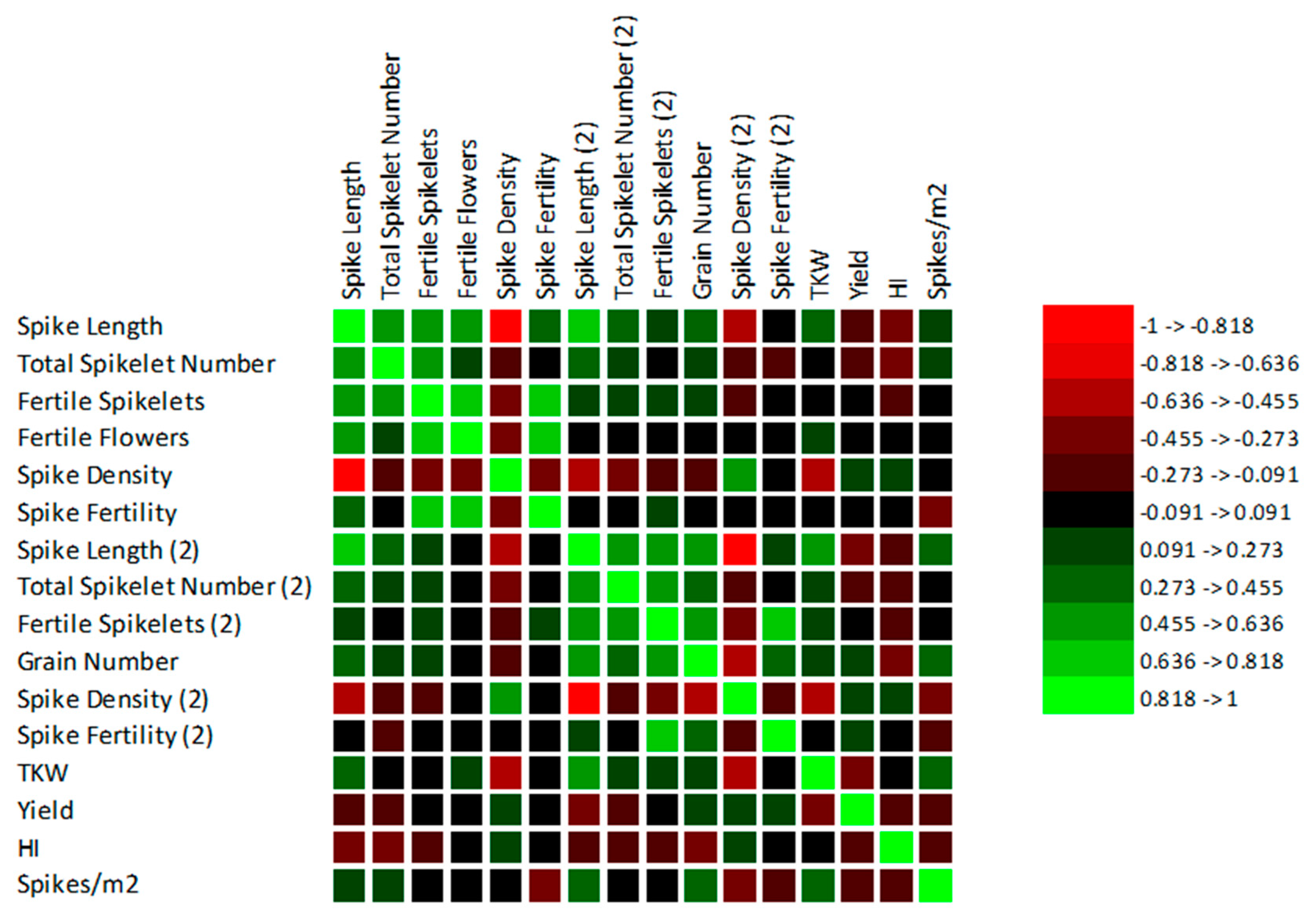
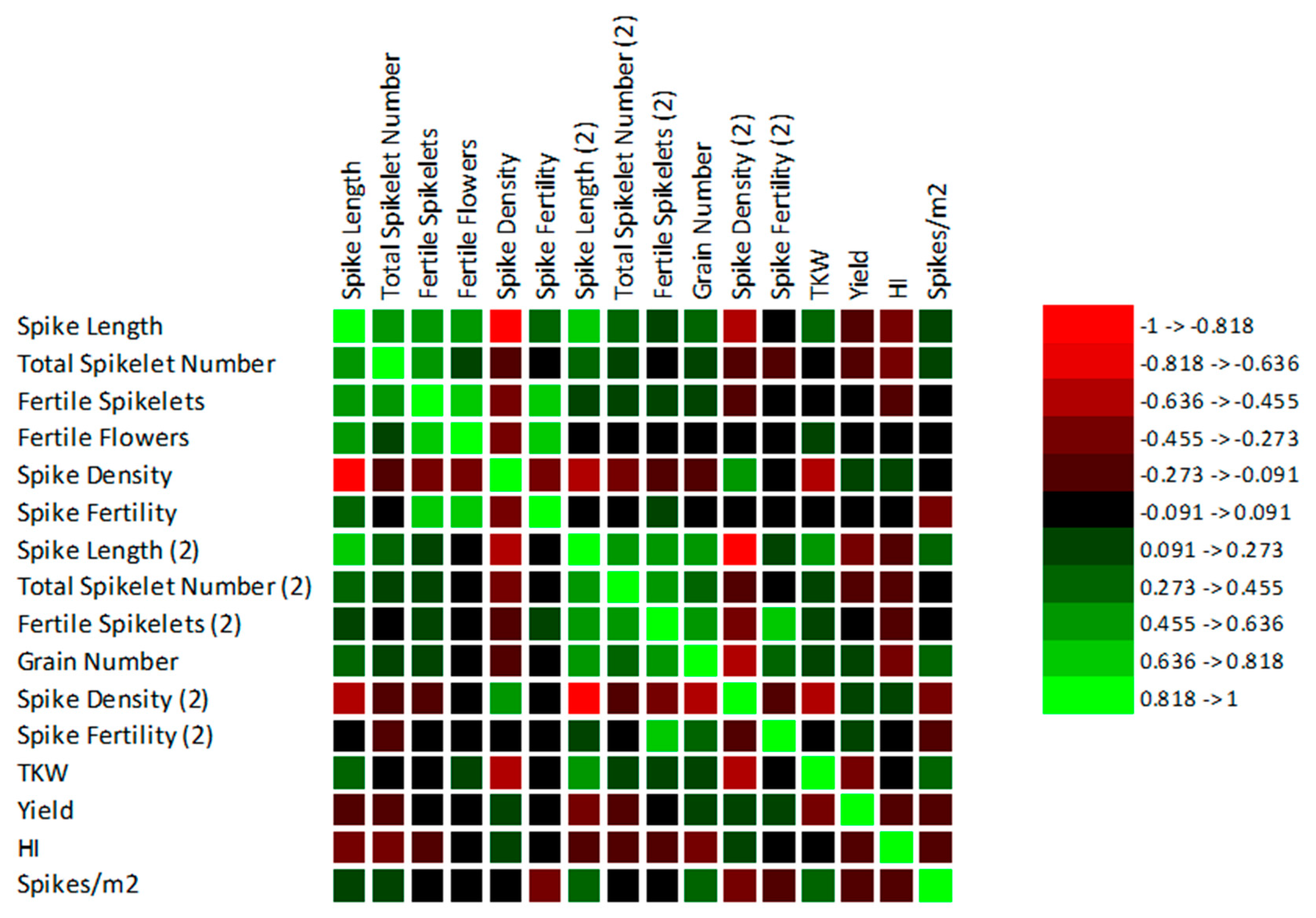
3.3. Correlations between the Observed Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilhite, D.; Glantz, M. Understanding the Drought Phenomena: The Role of Definitions. Water Int. 1985, 10, 111–120. [Google Scholar] [CrossRef]
- Kiliç, H.; Yağbasanlar, T. The Effect of Drought Stress on Grain Yield, Yield Components and some Quality Traits of Durum Wheat (Triticum turgidum ssp. durum) Cultivars. Not. Bot. Hort. Agrobot. Cluj. 2010, 38, 164–170. [Google Scholar] [CrossRef]
- Huang, J.; Zhuo, W.; Li, Y.; Huang, R.; Sedano, F.; Su, W.; Dong, J.; Tian, L.; Huang, Y.; Zhu, D.; et al. Comparison of three remotely sensed drought indices for assessing the impact of drought on winter wheat yield. Int. J. Digit. Earth 2018, 13, 504–526. [Google Scholar] [CrossRef]
- Khatiwada, A.; Neupane, I.; Sharma, B.; Bhetwal, N.; Pandey, B. Effects of Drought Stress on Yield and Yield Attributing Characters of Wheat: A Review. Agriways 2020, 8, 115–121. [Google Scholar] [CrossRef]
- Sarto, M.V.M.; Sarto, J.R.W.; Rampim, L.; Bassegio, D.; da Costa, P.F.; Inagaki, A.M. Wheat phenology and yield under drought: A review. Aust. J. Crop Sci. 2017, 11, 941–946. [Google Scholar] [CrossRef]
- Samaniego, L.; Thober, S.; Kumar, R.; Wanders, N.; Rakovec, O.; Pan, M.; Zink, M.; Sheffield, J.; Wood, E.F.; Marx, A. Anthropogenic warming exacerbates European soil moisture droughts. Nat. Clim. Chang. 2018, 8, 421–426. [Google Scholar] [CrossRef]
- Moss, R.; Babiker, M.; Brinkman, S.; Calvo, E.; Carter, T.; Edmonds, J.; Elgizouli, I.; Emori, S.; Erda, L.; Hibbard, K.; et al. Towards New Scenarios for Analysis of Emissions, Climate Change, Impacts, and Response Strategies; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2008; pp. 10–34. [Google Scholar]
- Spinoni, J.; Vogt, J.V.; Naumann, G.; Barbosa, P.; Dosio, A. Will drought events become more frequent and severe in Europe? Int. J. Climatol. 2017, 38, 1718–1736. [Google Scholar] [CrossRef]
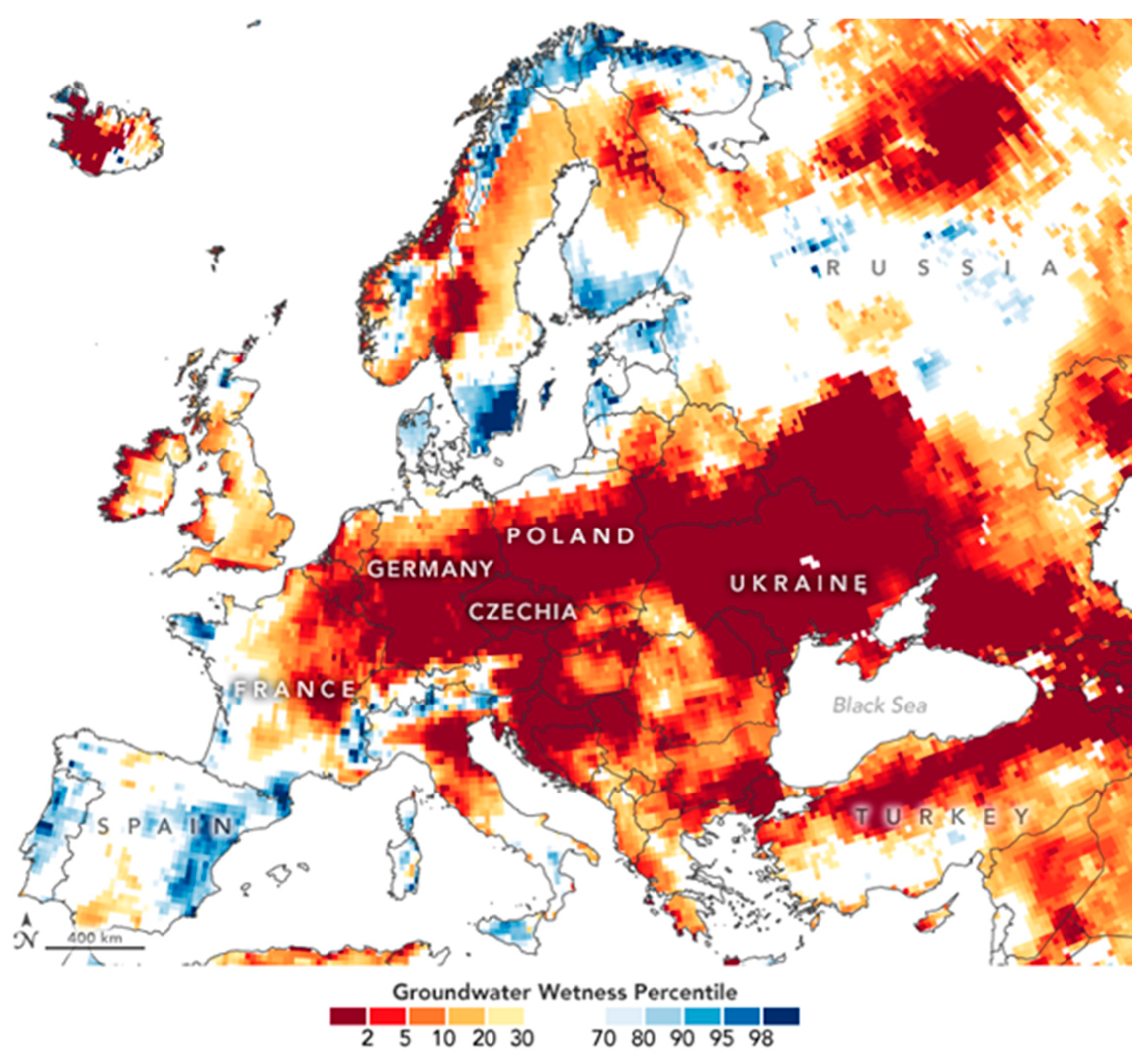
- Dauphin, L. NASA Observatory Image. National Drought Mitigation Center. 2020. Available online: https://earthobservatory.nasa.gov/images/146888/signs-of-drought-in-european-groundwater (accessed on 1 August 2022).
- Slafer, G.A.; Savin, R.; Sadras, V.O. Coarse and fine regulation of wheat yield components in response to genotype and environment. Field Crops Res. 2014, 157, 71–83. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Siddique, K.H.M. Drought Stress in Wheat during Flowering and Grain-filling Periods. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Ahmad, Z.; Waraich, E.A.; Akhtar, S.; Anjum, S.; Ahmad, T.; Mahboob, W.; Hafeez, O.B.A.; Tapera, T.; Labuschagne, M.; Rizwan, M. Physiological responses of wheat to drought stress and its mitigation approaches. Acta Physiol. Plant. 2018, 40, 80. [Google Scholar] [CrossRef]
- Ji, X.; Shiran, B.; Wan, J.; Lewis, D.C.; Jenkins, C.L.D.; Gondon, A.G.; Richards, R.A.; Dolferus, R. Importance of pre-anthesis anther sink strength for maintenance of grain number during reproductive stage water stress in wheat. Plant Cell Environ. 2010, 6, 926–942. [Google Scholar] [CrossRef] [PubMed]
- Pireivatlou, A.S.; Masjedlou, B.D.; Aliyev, R.T. Evaluation of yield potential and stress adaptive trait in wheat genotypes under post anthesis drought stress conditions. Afr. J. Agric. Res. 2010, 5, 2829–2836. [Google Scholar] [CrossRef]
- Khalili, M.; Naghavi, M.R.; Aboughadareh, A.P.; Rad, H.N. Effects of Drought Stress on Yield and Yield Components in Maise cultivars (Zea mays L.). Int. J. Agron. Plant Prod. 2013, 4, 809–812. [Google Scholar]
- Okuyama, L.A.; Federizzi, L.C.; Neto, J.F.B. Plant traits to complement selection based on yield components in wheat. Ciênca Rural 2005, 35, 1010–1018. [Google Scholar] [CrossRef]
- Mirbahar, A.A.; Markhand, G.S.; Mahar, A.R.; Abro, S.A.; Kanhar, N.A. Effect of water stress on yield and yield components of wheat (Triticum aestivum) varieties. Pak. J. Bot. 2009, 41, 1303–1310. [Google Scholar]
- Protić, R.; Protić, N.; Prodanović, R.; Zarić, G.; Hyba, H.H.H.; Mnifid, A.A.; Kharud, M.M.M. Spike Length of Winter Wheat Varieties According to Different Ways of Seed Protection. Rom. Biotechnol. Lett. 2018, 23, 13697–13701. [Google Scholar]
- Rodríguez, D.; Andrade, F.H.; Goudriaan, J. Effects of phosporus nutrition on tiller emergence in wheat. Plant Soil 1999, 209, 283–295. [Google Scholar] [CrossRef]
- Fang, Y.; Du, Y.; Wang, J.; Wu, A.; Qiao, S.; Xu, B.; Zhang, S.; Siddique, K.H.M.; Chen, Y. Moderate Drought Stress Affected Root Growth and Grain Yield in Old, Modern and Newly Released Cultivars of Winter Wheat. Front. Plant. Sci. 2017, 8, 672. [Google Scholar] [CrossRef]
- Zhang, L.; He, X.; Liang, Z.; Zhang, W.; Zou, C.; Chen, X. Tiller development affected by nitrogen fertilisation in a high-yielding wheat productive system. Crop Sci. 2020, 60, 1034–1047. [Google Scholar] [CrossRef]
- Rajala, A.; Hakala, K.; Mäkelä, P.S.; Peltonen-Sainio, P. Drought Effect on Grain Number and Grain Weight at Spike and Spikelet Level in Six-Row Spring Barley. J. Agron. Crop Sci. 2010, 197, 103–112. [Google Scholar] [CrossRef]
- Knezevic, D.; Zecevic, V.; Stamenkovic, S.; Atanasijevic, S.; Milosevic, B. Variability of number of kernels per spike in wheat cultivars (Triticum aestivum L.). J. Cent. Eur. Agric. 2012, 13, 617–623. [Google Scholar] [CrossRef]
- Langridge, P.; Reynolds, M. Breeding for drought and heat tolerance in wheat. Theor. Appl. Genet. 2021, 134, 1753–1769. [Google Scholar] [CrossRef] [PubMed]
- Rizza, F.; Badeck, F.W.; Cattivelli, L.; Lidestri, O.; di Fonzo, N.; Stanca, A.M. Use of a water stress index to identify barley genotypes adapted to rainfed and irrigated conditions. Crop Sci. 2004, 44, 2127–2137. [Google Scholar] [CrossRef]
- Li, C.; Li, L.; Reynolds, M.P.; Wang, J.; Chang, X.; Mao, X.; Ring, R. Recognising the hidden half in wheat: Root system attributes associated with drought tolerance. J. Exp. Bot. 2021, 72, 5117–5133. [Google Scholar] [CrossRef]
- Rijal, B.; Baduwal, P.; Chaudhary, M.; Chapagain, S.; Khanal, S.; Khanal, S.; Poudel, P.B. Drought Stress Impacts on Wheat and Its Resistance Mechanisms. Malays. J. Sustain. Agric. 2021, 5, 67–76. [Google Scholar] [CrossRef]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Antioxidant responses of wheat plants under stress. Genet. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef]
- Munns, R.; Richards, R.A. Recent Advances in Breeding Wheat for Drought and Salt Stresses. In Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops; Jenks, M.A., Hasegawa, P.M., Jain, S.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 565–585. [Google Scholar]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future Köppen-Geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 180214. [Google Scholar] [CrossRef]
- Rafferty, J.P. (Ed.) Chapter 2, Climatic Classification. In Climate and Climate Change; Britannica Educational Publishing, Rosen Educational Services: New York, NY, USA, 2011; pp. 100–124. [Google Scholar]
- Cane, K.; Eagles, H.A.; Laurie, D.A.; Trevaskis, B.; Vallance, N.; Eastwood, R.F.; Gororo, N.N.; Kuchel, H.; Martin, P.J. Ppd-B1 and Ppd-D1 and their effects in southern Australian wheat. Crop Pasture Sci. 2013, 64, 100–114. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Guo, Z.; Zhao, Y.; Röder, M.S.; Reif, J.C.; Ganal, M.W.; Chen, D.; Schnurbusch, T. Manipulation and prediction of spike morphology traits for the improvement of grain yield in wheat. Sci. Rep. 2018, 8, 14435. [Google Scholar] [CrossRef]
- Mwadzingeni, L.; Shimelis, H.; Rees, D.J.G.; Tsilo, T.J. Genome-wide association analysis of agronomic traits in wheat under drought-stressed and non-stressed conditions. PLoS ONE 2017, 12, e0171692. [Google Scholar] [CrossRef] [PubMed]
- Qaseem, M.F.; Qureshi, R.; Shaheen, H. Effects of Pre-Anthesis Drought, Heat and Their Combination on the Growth, Yield and Physiology of diverse Wheat (Triticum aestivum L.) Genotypes Varying in Sensitivity to Heat and drought stress. Sci. Rep. 2019, 9, 6955. [Google Scholar] [CrossRef] [PubMed]
- Taheri, S.; Saba, J.; Shekari, F.; Abdullah, T.L. Effects of drought stress condition on the yield of spring wheat (Triticum aestivum) lines. Afr. J. Biotechnol. 2011, 10, 18339–18348. [Google Scholar]
- Khyber, J.A.; Soomro, F.; Sipio, W.D.; Baloch, A.W.; Soothar, J.K.; Soothar, M.K.; Ali, Z. Evaluation of Bread Wheat (Triticum aestivum L.) Genotypes forDrought Tolerance through Selection Indices. J. Hortic. Plant Res. 2019, 7, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Wasaya, A.; Manzoor, S.; Yasir, T.A.; Sarwar, N.; Mubeen, K.; Ismail, I.A.; Raza, A.; Rehman, A.; Hossain, A.; EL Sabagh, A. Evaluation of Fourteen Bread Wheat (Triticum aestivum L.) Genotypes by Observing Gas Exchange Parameters, Relative Water and Chlorophyll Content, and Yield Attributes under Drought Stress. Sustainability 2021, 13, 4799. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, M.; Gu, W.; Chen, Z.; Gu, Y.; Pei, L.; Tian, R. Effect of drought on photosynthesis, total antioxidant capacity, bioactive component accumulation, and the transcriptome of Atractylodes lancea. BMC Plant Biol. 2021, 21, 293. [Google Scholar] [CrossRef]
- Litvin, A.G.; van Iersel, M.W.; Malladi, A. Drought Stress Reduces Stem Elongation and Alters Gibberellin-related Gene Expression during Vegetative Growth of Tomato. J. Am. Soc. Hortic. Sci. 2016, 141, 591–597. [Google Scholar] [CrossRef]
- Sangtarash, M.H. Responses of Different Wheat Genotypes to Drought Stress Applied at Different Growth Stages. Pak. J. Biol. Sci. 2010, 13, 114–119. [Google Scholar] [CrossRef]
- Blum, A.; Ramaiah, S.; Kanemasu, E.T.; Paulsen, G.M. Wheat recovery from drought stress at the tillering stage of development. Field Crops Res. 1990, 24, 67–85. [Google Scholar] [CrossRef]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef]
- Arjona, J.M.; Royo, C.; Dreisigacker, S.; Ammar, K.; Villegas, D. Effect of Ppd-A1 and Ppd-B1 Allelic Variants on Grain Number and Thousand Kernel Weight of Durum Wheat and Their Impact on Final Grain Yield. Front. Plant Sci. 2018, 9, 888. [Google Scholar] [CrossRef] [PubMed]
- Würschum, T.; Rapp, M.; Miedaner, T.; Friedrich, C.; Longin, H.; Leiser, W.L. Copy number variation of Ppd-B1 is the major determinant of heading time in durum wheat. BMC Genet. 2019, 20, 64. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, H.; Fujita, M.; Seki, M.; Kojima, H.; Shimazaki, Y.; Matsunaka, H.; Chono, M.; Hatta, K.; Kubo, K.; Takayama, T.; et al. Growth and Yield Properties of Near-Isogenic Wheat Lines Carrying Different Photoperiodic Response Genes. Plant Prod. Sci. 2015, 18, 57–68. [Google Scholar] [CrossRef]
- Díaz, A.; Zikhali, M.; Turner, A.S.; Isaac, P.; Laurie, D.A.; Hazen, S.P. Copy Number Variation Affecting the Photoperiod-B1 and Vernalization-A1 Genes Is Associated with Altered Flowering Time in Wheat (Triticum aestivum). PLoS ONE 2012, 7, e33234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Gianmarco, T.I.; Slafer, G.A.; González, F.G. Photoperiod-sensitivity genes shape floret development in wheat. J. Exp. Bot. 2019, 70, 1339–1348. [Google Scholar] [CrossRef]
- Duggan, B.L.; Domitruk, D.R.; Fowler, D.B. Yield component variation in winter wheat grown under drought stress. Can. J. Plant Sci. 2000, 80, 739–745. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Earliness | Awns | Photoperiod Sensitivity | Country of Origin |
|---|---|---|---|---|
| Balitus | very early | awnless | insensitive (a) | Austria |
| Bohemia | very early | awnless | insensitive (a) | Czech Republic |
| IS Conditor | moderately early | awned | sensitive (b) | Slovakian Republic |
| RGT Sacramento | moderately early | awned | insensitive (a) | France |
| Tobak | semi-late | awnless | sensitive (b) | Germany |
| Tonnage | semi-late | awnless | sensitive (b) | Denmark |
| Tybalt | semi-late | awnless | sensitive (b) | Netherlands |
| Year | 2019/2020 | 2020/2021 | Site | ||
|---|---|---|---|---|---|
| days from emergence | 216 | 243 | 236 | 276 | FIELD 1 |
| Zadoks scale | 64 | 73 | 66 | 91 | |
| days from emergence | 222 | 249 | 235 | 275 | FIELD 2 |
| Zadoks scale | 67 | 80 | 68 | 91 | |
| Traits | 2019/2020 | 2020/2021 | ||||||
|---|---|---|---|---|---|---|---|---|
| FIELD 1 | FIELD 2 | FIELD 1 | FIELD 2 | |||||
| Ppd-D1a | Ppd-D1b | Ppd-D1a | Ppd-D1b | Ppd-D1a | Ppd-D1b | Ppd-D1a | Ppd-D1b | |
| Spike Length * | 105.9 ± 18.9 | 102.1 ± 7.4 | 97.4 ± 16.2 | 92.2 ± 7.3 | 94.5 ± 16.1 | 89.0 ± 9.2 | 91.3 ± 18.1 | 83.7 ± 6.1 |
| Total Spikelet Number | 20.2 ± 1.0 | 20.8 ± 1.4 | 19.1 ± 1.1 | 19.3 ± 1.3 | 19.5 ± 1.1 | 20.2 ± 1.0 | 18.4 ± 1.4 | 18.8 ± 0.8 |
| Fertile Spikelets | 17.7 ± 1.2 | 18.6 ± 1.6 | 16.6 ± 1.6 | 17.2 ± 1.3 | 17.5 ± 1.1 | 18.2 ± 1.4 | 16.3 ± 1.3 | 16.4 ± 1.1 |
| Fertile Flowers | 51.9 ± 8.5 | 57.0 ± 6.5 | 50.8 ± 10.4 | 53.8 ± 9.6 | 50.9 ± 5.2 | 55.3 ± 6.6 | 48.2 ± 7.0 | 48.3 ± 7.4 |
| Spike Density | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 |
| Spike Fertility **** | 87.7 ± 5.2 | 89.0 ± 3.4 | 87.2 ± 5.8 | 89.1 ± 7.1 | 89.4 ± 3.4 | 90.2 ± 4.4 | 88.4 ± 3.2 | 87.5 ± 4.0 |
| Spike Length (2) * | 99.5 ± 11.9 | 85.0 ± 9.8 | 95.1 ± 14.9 | 81.9 ± 5.2 | 86.4 ± 13.8 | 81.3 ± 7.0 | 81.1 ± 13.5 | 77.3 ± 7.2 |
| Total Spikelet Number (2) | 20.1 ± 0.9 | 19.8 ± 1.2 | 18.8 ± 1.1 | 18.5 ± 0.8 | 19.3 ± 1.4 | 19.3 ± 0.9 | 18.4 ± 1.1 | 18.5 ± 1.1 |
| Fertile Spikelets (2) | 17.9 ± 1.0 | 17.6 ± 1.4 | 16.6 ± 1.3 | 15.9 ± 0.9 | 16.9 ± 1.2 | 17.17 ± 1.1 | 15.3 ± 1.1 | 16.0 ± 1.2 |
| Grain Number | 52.5 ± 5.8 | 54.4 ± 6.5 | 48.1 ± 8.3 | 48.8 ± 6.4 | 44.5 ± 4.7 | 49.0 ± 5.1 | 38.3 ± 5.3 | 42.9 ± 6.1 |
| Spike Density (2) | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.2 ± 0.0 |
| Spike Fertility (2) **** | 89.1 ± 5.1 | 88.6 ± 4.9 | 88.4 ± 6.8 | 85.8 ± 4.3 | 87.6 ± 3.9 | 88.9 ± 5.0 | 83.5 ± 4.5 | 86.6 ± 4.5 |
| TKW ** | 42.4 ± 2.8 | 37.9 ± 1.7 | 44.0 ± 1.4 | 43.3 ± 2.0 | 44.1 ± 2.0 | 41.5 ± 1.1 | 43.0 ± 1.8 | 40.1 ± 1.8 |
| spikes/m2 | 836.7 ± 38.6 | 972.5 ± 126.2 | 591.7 ± 37.2 | 549.1 ± 44.0 | 666.7 ± 74.5 | 748.3 ± 64.8 | 449.9 ± 34.6 | 478.6 ± 56.8 |
| HI | 0.4 ± 0.0 | 0.3 ± 0.0 | 0.4 ± 0.0 | 0.4 ± 0.0 | 0.5 ± 0.0 | 0.5 ± 0.0 | 0.4 ± 0.0 | 0.4 ± 0.0 |
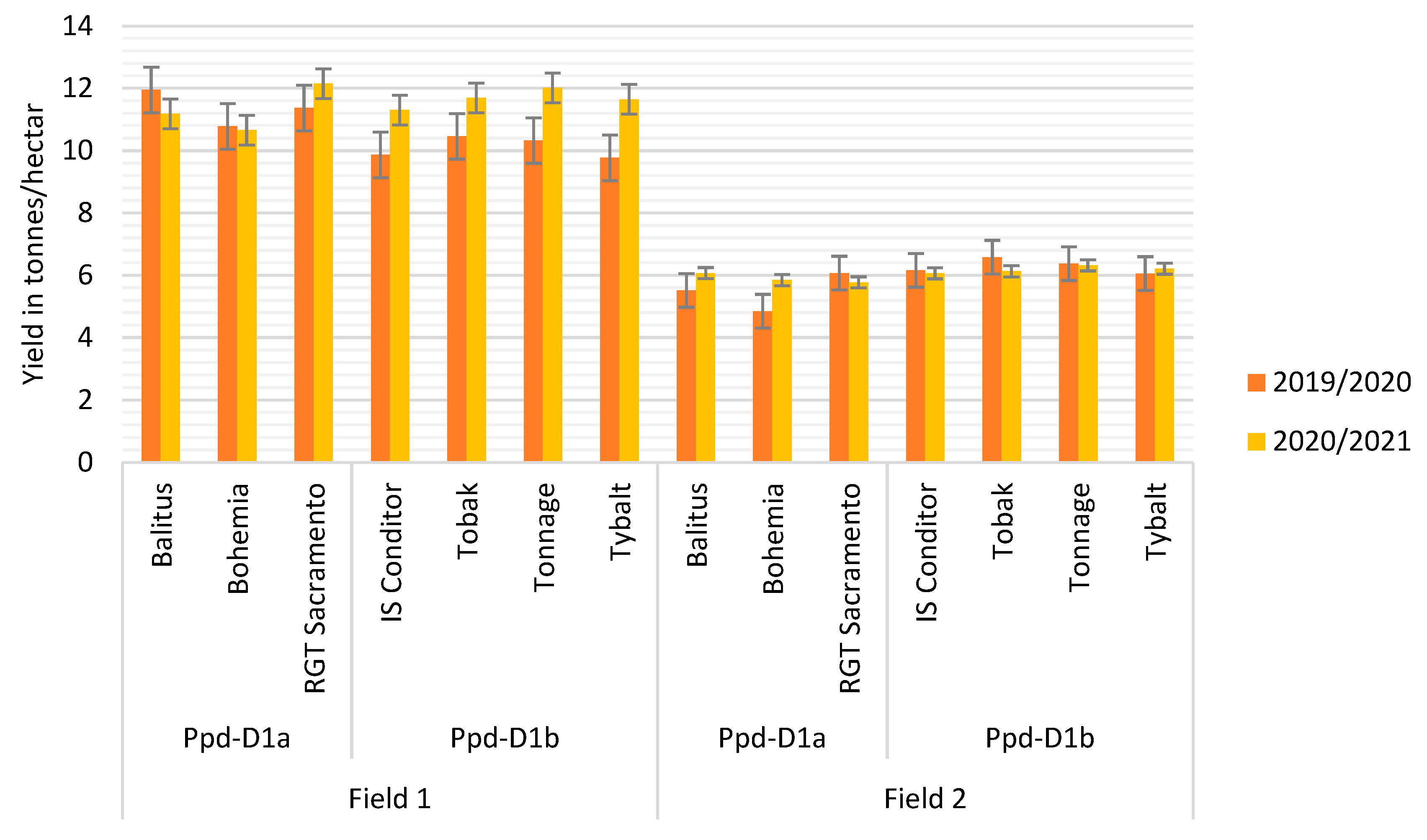
| Yield *** | 11.4 ± 0.5 | 10.1 ± 0.3 | 5.5 ± 0.5 | 6.3 ± 0.2 | 11.3 ± 0.6 | 11.66 ± 0.3 | 5.9 ± 0.1 | 6.1 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frantová, N.; Rábek, M.; Elzner, P.; Středa, T.; Jovanović, I.; Holková, L.; Martinek, P.; Smutná, P.; Prášil, I.T. Different Drought Tolerance Strategy of Wheat Varieties in Spike Architecture. Agronomy 2022, 12, 2328. https://doi.org/10.3390/agronomy12102328
Frantová N, Rábek M, Elzner P, Středa T, Jovanović I, Holková L, Martinek P, Smutná P, Prášil IT. Different Drought Tolerance Strategy of Wheat Varieties in Spike Architecture. Agronomy. 2022; 12(10):2328. https://doi.org/10.3390/agronomy12102328
Chicago/Turabian StyleFrantová, Nicole, Michal Rábek, Petr Elzner, Tomáš Středa, Ivana Jovanović, Ludmila Holková, Petr Martinek, Pavlína Smutná, and Ilja Tom Prášil. 2022. "Different Drought Tolerance Strategy of Wheat Varieties in Spike Architecture" Agronomy 12, no. 10: 2328. https://doi.org/10.3390/agronomy12102328