
Vermicompost: Enhancing Plant Growth and Combating Abiotic and Biotic Stress
by
 , , , and
, , , and
Sami ur Rehman
 ,
,
Federica De Castro
 ,
,
Alessio Aprile
* ,
,
Michele Benedetti
* and
Francesco Paolo Fanizzi
Department of Biological and Environmental Sciences and Technologies (DiSTeBA), University of Salento, Via Monteroni, I-73100 Lecce, Italy
*
Authors to whom correspondence should be addressed.
Agronomy 2023, 13(4), 1134; https://doi.org/10.3390/agronomy13041134
Submission received: 31 March 2023
/
Revised: 12 April 2023
/
Accepted: 14 April 2023
/
Published: 16 April 2023
(This article belongs to the Section Soil and Plant Nutrition)
Abstract
:Extensive application of agrochemicals for crop production and protection has negatively affected soil health, crop productivity, and the environment. Organic amendments have been proposed as an efficient alternative for enhancing soil and plant health. Vermicompost amendment offers a sustainable approach to plant nutrition, improving soil health and fertility. This review aims to provide key insights into the potential of vermicompost to boost crop production and protect crops from biotic and abiotic stresses without harming the environment. The role played by earthworms in improving organic matter decomposition, soil fertility, and soil microorganisms’ activity is also discussed here. The value of vermicompost is its promotion of plant growth based on its enrichment with all essential nutrients, beneficial microbes, and plant growth hormones. This review analyzes how vermicompost regulates plant growth and its role in mitigating abiotic stresses such as soil salinity and drought, as well as biotic stresses such as diseases and insect pests attack. The beneficial effects of hormones and humic substances present in vermicompost are also discussed in this review. In fact, due to its properties, vermicompost can be a good substitute for chemical fertilizers and pesticides and its usage could contribute to producing healthy, contaminant-free food for the growing population without negatively affecting the environment.
1. Introduction
Since the “Green Revolution” in the 1960s, intensive use of chemical fertilizers significantly increased food production to meet the demands of the growing population. The food requirements of 50% of the population are met using chemical fertilizers for crop production [1]. However, this has led to environmental pollution and health issues due to agrochemical residues in food commodities [2,3,4]. The harmful effects of chemical fertilizers have moved scientists’ focus toward “green substitutes” with low environmental impact [5]. Among these, vermicompost is an appealing alternative to conventional chemical fertilizers. Vermicomposting is a non-thermophilic process that transforms organic waste materials into valuable fertilizer through the combined action of worms and mesophilic microbes [6]. This method is more effective for organic waste degradation than composting because waste material passes through the earthworm’s gut, excreting casts at temperatures between 10–32 °C. Vermicomposting involves physical processes such as fragmentation, aeration, and the turning over of wastes, as well as biochemical processes such as enzyme digestion, waste material transformation, and enrichment. Earthworms (EWs) modify the physicochemical and biological status of organic matter (OM), reducing the C/N ratio, increasing surface area, exposing more sites for microbial action, and enhancing decomposition [7]. Vermicomposting is considered a sustainable approach to waste management and agricultural production. As a result, it minimizes chemical fertilizers’ application to soil, reducing their harmful effects and the amount of waste directed to landfills [8].
Vermicompost (VC) is a peat-like organic fertilizer with high nutritional contents, aeration, porosity, and water-holding capacity, prepared by the joint action of earthworms and microbes. In addition to organic waste management, VC is recognized as an effective plant growth promoter [9]. Nitrogen (N), phosphorus (P), Potassium (K), and other micronutrients become more available due to microbial activities in VC [10]. Chemical analysis of VC reveals higher levels of N (5 times), K (7 times), and Ca (1.5 times) than the topsoil at a depth of 15 cm in which plants grow [11,12]. The average nutrient contents in VC include N (1.5–2%), P (1.8–2.2%), K (1–1.5%), and organic carbon (0.15–17.98%), along with other micronutrients required for plant growth such as zinc (Zn), magnesium (Mg), and iron (Fe) [13]. Vermicompost is considered a long-term supplier of these macro and micronutrients in a readily available form to plants [14]. Furthermore, vermicompost is enriched with soil microbiota such as nitrogen-fixing and phosphorus-solubilizing organisms [15]. Derivatives of vermicompost, such as VC leachates, humic substances, and phytohormones, are the key stimulators of plant growth promotion. VC leachate is an organic fertilizer consisting of liquid nutrients collected after the water passes through a heap of VC [16]. Vermi-tea (VT) is a water extract of solid VC from which microbes, soluble nutrients, and other beneficial elements are transformed into liquid form [17]. Humic substances (HS) are effective plant biostimulants and can be used on plants to enhance growth and yield by increasing nutrient uptake [18]. HS comprise humic acid, fulvic acid, and humin. HS stimulate lateral roots development and extension, alleviate stress, and improve soil properties and microbial structure [19]. Phytohormones such as auxin, gibberellic acid, and cytokinin have been observed in compost processed by EWs, which promote crop growth [6,20,21].
Vermicompost has been shown to have a broad range of effects on abiotic and biotic stresses, including ameliorating soil salinity [22], mitigating drought stress [23], and controlling insect pests and diseases [24]. Soil amendment with vermicompost has been found to suppress the attacks from Aproaerema modicella (a leaf miner) in the groundnut field [13]. Applying vermicompost tea (VT) has been shown to suppress Fusarium moniliforme, thereby controlling foot rot disease in rice [25]. VT also contains phenolic substances that make plant tissues unpalatable to insect pests [26]. Numerous studies have examined vermitechnology as waste management, vermicompost production, and vermicompost’s nutrient characteristics [27]. However, further investigation is needed to understand its role as a plant growth regulator and biostimulator for plants under stress conditions. This review aims to explain the potential of vermicompost as an outstanding plant growth regulator, and also addresses the questions that how vermicompost combats plant stresses such as water scarcity, soil salinity, insect pests, and disease, highlighting its function an eco-friendly organic fertilizer and biopesticide in comparison to other commonly used agrochemicals (Figure 1).
2. Earthworm Impact
Due to their priming effects, earthworms serve as ecological mediators influencing soil structures and microbial activities [28]. They are considered among the largest soil species in tropical and temperate regions, accounting for 40–90% of soil macro-fauna [29]. EWs are classified into three main niche groups: epigeic, endogeic, and anecic. Anecic species inhabit deep mineral layers, endogeic species are active in the uppermost mineral layers, and epigeic species feed on surface litter. They promote organic matter decomposition by enhancing microbial activity and population in the soil [30]. EWs such as Eisenia fetida and Eudrilus sp. have been found to increase bacterial diversity during the early stages of vermicomposting [31,32]. EWs play a vital role in breaking down OM and converting macro and micronutrients [33]. The comprehensive roles of EWs in OM decomposition, microorganisms’ activity, soil structure, and nutrient availability are discussed here.
2.1. Soil Structure
Among different soil characteristics, its structure is the most important parameter determining fertility. For example, an ideal soil structure improves water-holding capacity, reducing water and soil loss [34]. EWs improve soil structure through humus production, mineral weathering, and the mixing of OM to form soil aggregates. Although it has been demonstrated that E. fetida increases the weathering of kaolinite, biotite, smectite, and anorthite [35], the effect of EWs on mineral weathering still needs to be fully understood. Further studies are required to determine whether the EWs, the microbes in their gut, or both induce an increase in mineral weathering.
A recent study on soil aggregate formation by Al-Maliki and Scullion [36] focused on the relationship between kaolinite and organic material in the presence of EWs. Researchers found that EWs regulate soil structure by improving aggregate stability and stabilizing aliphatic carbon in kaolinite. The burrowing activities of EWs influence the mechanical and hydraulic properties of soil, creating macropores for water percolation in soil profiles and reducing surface runoff [37]. In tropical and temperate soils, the casting and burrowing practices of the anecic EWs control erosion by increasing soil structural stability and porosity [38]. The effect of EWs on soil compaction and loss depends on species type and their interaction with the soil. The long-term interaction with the Eudrilidae family (de-compacting EWs) and Reginaldia omodeoi (compacting EWs) helps maintain soil structure in tropical regions [28]. The cast production in horizontal and vertical burrows by endogeic and anecic EWs influences soil bulk density and porosity. For instance, Rhyacodrilus omodeoi, an anecic earthworm, increased the bulk density from 1.12 to 1.23 g cm−3, and Pontoscolex corethrurus, an endogeic earthworm, reduced the soil porosity from 58% to 53% [39,40].
2.2. Organic Matter Decomposition
Decomposition is a crucial process that regulates nutrient cycling in terrestrial environments through decomposers such as nematodes, protozoa, microbes, and EWs. EWs are efficient members of the decomposers’ community and play a significant role in plant residue decomposition and the turnover of other organic materials [41,42]. EWs considerably impact decomposition by modifying OM and microbes that pass through their gut and are released in the cast [43,44]. During the gut passage, mucus is added, accelerating microbial and enzymatic activities, thereby increasing OM mineralization [45,46]. Through the vertical distribution of grounded material in the soil profile, EWs’ activity increases surface area for microbial colonization and faster decomposition [47]. Organic matter breakdown by EWs is closely related to the material’s chemical properties. The palatability of material for EWs increases with a reduction in the C/N ratio. Ernst et al. confirmed that residue decomposition by EWs was highest in maize litter (C/N: 34.8) compared to Miscanthus litter (C/N: 134.4) [41]. Epigeic EWs inhabit litter, ingest, and transform OM from various habitats such as forest litter and livestock dung. They interact with microbes and other organisms within the habitat, affecting decomposition processes [43,48]. EWs contribute to decomposition processes through fragmentation, incorporation, and mixing organic material into the soil.
2.3. Soil Fertility
Various species of EWs can produce different soil biological profiles and fertility levels. The combined activity of these species ensures the maintenance of soil fertility throughout the soil profile [49,50]. EWs play a vital role in producing soil aggregates and biostructures such as pores for better movement of nutrients and water. They also increase N mineralization by encouraging the microbial population both directly and indirectly [51,52]. EWs effectively contribute to nitrogen recycling and enhance the availability of essential plant nutrients. They consume an abundance of substrate but retain only a tiny portion of it (5–10%); the remaining substrate is excreted as vermicompost, which is enriched in N, P, K, and other micronutrients and beneficial microbes. These microbes increase the nitrogen fixation process, providing more nitrogen in worm casts [38,53]. A meta-analysis of casts reveals higher nutrient contents in casts than soil [54]. EWs are also well-known for increasing phosphorus availability in their casts and burrows [55]. P is an essential nutrient for plants, helping to accumulate and transform energy in the metabolic activities of living organisms. It also promotes seedling growth and crop maturity. Earthworm casts contain more P than the soil without them, so EWs have a positive association with P acquisition in soil [56].
Van Groenigen et al. [54] focused on the increase in soil fertility properties in earthworm casts and coined a new term, “relative cast fertility (RCF)” [54]. RCF explains the relative difference in fertility and soil properties, such as nutrient availability and pH, between the EWs casts and the soil. This difference can result from two key processes: concentration and transformation. Firstly, EWs can enhance the RCF by concentrating existing soil fertility towards earthworm-influenced pools of soil fertility (Figure 2). EWs are selective in choosing their food, and this food selection varies among species [57]. The choice of EWs’ diet centers around the size, age, biochemical properties, and microbial population in the feeding mixture [58,59,60,61]. Secondly, RCF can be increased by the transformation process in the gut or casts, as illustrated in Figure 2. The earthworm gut is home to various microbes effectively involved in several biochemical processes [62,63], which affect soil fertility properties. Due to the transformation process, nutrient availability and pH are usually higher in worm casts than in the soil. For instance, N and P contents are higher in the gut or casts [64,65], and the availability of other nutrients such as Ca, Mg, and K may also increase during gut transit [35].
2.4. Soil Microorganisms
EWs positively or negatively impact microbial populations and their ecological diversity. They influence the functions of microbial decomposers by feeding on microorganisms and making adjacent areas more susceptible to microbial attack after organic matter is broken down. Microorganisms such as bacteria, fungi, and protozoa are essential food sources for the EWs. The combined action of EWs and microorganisms accelerates the decomposition of organic matter [66]. The function of microorganisms and organic matter decomposition is affected by gut-associated processes. The composition of gut-associated microbes (GAM) is related to the ingested material. In addition, the number and behavior of the microbes in EWs’ gut differ from those in uninterrupted material. Hong et al. [67] analyzed the community structure of GAM and explained the impact of enzyme-producing microbes on EWs biomass. They detected 57 bacterial clones in EWs gut using PCR-DGGE analysis. Aeromonas hydrophila, Paenibacillus motobuensis, and Photobacterium ganghwense were active in enzyme production. The mixture inoculated with Aeromonas hydrophila and Paenibacillus motobuensis showed the highest survival rate (100%) and increased the EWs’ growth and cast production, demonstrating a symbiotic relationship between EWs and microorganisms. Specific microorganisms respond differently to the gut environment, and the selective impact on microorganisms during the passage through the EWs’ gut has been analyzed [68]. For example, some bacteria became activated during passage to EWs’ gut, while others remain unchanged or are digested, thus reducing their number [62,69]. Monroy et al. [70] observed a 98% decline in coliforms density after pig slurry passed through the gut of Eisenia fetida. Earthworms can modify microbial physiology and stimulate enzymatic activity during the vermicomposting of pig slurry [31,32,50]. Studies on soil and six species of earthworms and fungi showed that EWs process the soil efficiently by increasing the fungal species involved in faster decomposition processes [71,72].
3. Vermicompost and Its Derivatives as a Plant Growth Promoter
Vermicompost enhances crop growth, yield, and quality due to its plant growth-promoting characteristics. Vermicompost stimulates plant emergence because of the availability of essential plant nutrients. The effects of vermicompost and its derivatives on plant growth and yield are summarized in Table 1. Loera-Muro et al. [73] studied the morphology of Rosmarinus officinalis and Mentha spicata plants, as well as microbial count, using vermicompost leachates (VCL) and inorganic fertilizer. Plants treated with VCL showed an increased root length for M. spicata (74 cm) compared to those treated with inorganic fertilizer (38 cm). Bidabadi et al. [74] examined the effect of foliar-applied VCL on pomegranate and observed a significant increase in leaf area, root, and shoot biomass, along with the induction of salt tolerance in pomegranate. VCL also boosts antioxidant enzyme activity and reduces oxidative stress. This liquid partially originates from EWs bodies and is enriched with nutrients, vitamins, amino acids, and various growth hormones such as auxins and cytokines [75]. Seeds soaked in VCL and joint application of VC + VCL led to significant improvement in biochemical parameters (photosynthetic rate, total chlorophyll, transpiration rate) of Withania somnifera [76]. Aslam et al. [77] analyzed the impact of VC, VT, and chemical fertilizer on tomato (Solanum lycopersicum) growth. The results showed that foliar application of 5% VT in tomatoes increased the physiological and morphological parameters. Plants treated with 10% VT reached the flowering stage three weeks earlier than the control and improved protein and soluble sugar contents and the growth of faba beans.
3.1. Humic Substances
Humic substances (HS) can influence nutrient uptake by modifying root anatomy for better nutrient interception, enhancing root exudates, supporting plant–microbe interactions, and altering the expression of transporters involved in nutrient acquisition [89,90]. Direct impacts of HS on plant growth are due to improvements in nutrient uptake, while indirect effects include alterations of physiochemical and biological changes in amended soil [89,91]. Fulvic and humic acids in VC dissolve insoluble minerals in organic matter (OM), making them more accessible to plants, aiding stress tolerance, and promoting growth [92]. HS improved the soil aggregate stability and reduced N leaching and soil erosion. They have a high affinity for organic and inorganic ions in the soil and can form complexes with cations and inorganic phosphorus, preventing leaching and promoting bioavailability for plant uptake. These complexes are formed due to the presence of O-, N-, and S-containing functional groups in the HS structure and are important because nutrient deficiency is linked to the low availability of these metallic cations [93,94]. Humic substances are produced during the humification process of organic matter. Mature VC primarily consists of humified substances produced by the action of EWs, which accelerate the humification process [95]. Dobbs and coworkers [96] evaluated the bioactivity of HS by modifying the root architecture and proton pump activation of maize and tomato. All the VC-derived HS showed more significant bioactivity than the original HS and Potassium permanganate (KMnO4) oxidized, being highly efficient in modifying root architecture and proton pump activation for both test plants. However, no consistent relationship was observed between the molecular sizes of HS and bioactivity. The hydrophobicity index was significantly correlated with the stimulation of the proton pump. It is also proposed that the hydrophobic domain can stabilize bioactive molecules, such as auxins, in HS. The hydrophobic forces can be disrupted by contact with organic acids produced by roots, releasing bioactive compounds from humic material. The application of vermicompost and VC-derived humic acids and fulvic acids improved Cannabis sativa L. growth when applied at 5%, 0.05 mg/mL, and 1%, respectively [97]. Humic acid fertilizers are widely used in soil remediation, crop production, and environmental safety. Humic acid improves soil structure, fertilizer use efficiency, and crop growth and yield, increasing farmers’ income. Gomes et al. [98] evaluated the effect of humic acid derived from vermicompost of sugarcane filter cake on Garcinia mangostana L. seedlings. Results showed increased N, P, K, Mg, Ca, S, Mn, and Zn uptake by shoots and roots of the plant, enhanced chlorophyll (a, b) contents, and decreased carotenoid contents. Similarly, the results of El-Hameid et al. [99] described that VCL with humic acid increased total chlorophyll content, leaf mineral contents, fruit retention, fruit weight, number of fruits, total sugar contents, ascorbic acid, and total acidity in mangoes. The foliar application of humic substances on plants significantly increases their growth and fruit production. This stimulating effect of HS is attributed to humic acids and fulvic acid molecules [100]. Balmori et al. [101] studied the effect of foliar application of humic acid extract from vermicompost on fruit production and the quality of garlic (Allium sativum L.). Results demonstrated the stimulating effect of vermicompost-derived HS on Allium sativum metabolism, increasing bulb caliber, the number of garlic cloves, and fruit quality indices compared to control. The nutrient uptake (N, P, K, Ca, Mg) by plants increased with the use of humic acid derived from VC. Baldotto et al. [102] reported a beneficial effect of humic acids from vermicompost on Ca and Mg accumulation in pineapple seedlings. Correspondingly, Nikbakht et al. [103] also described the positive effect of humic acids on the uptake of Ca and Mg in gerbera seedlings. Hernandez et al. [104] assessed humic substances derived from vermicompost and applied them to lettuce leaves. They concluded that humates applied at 15 mg C L−1 shortened the production cycle and allowed for early harvesting without affecting the lettuce quality and yield. The application of humates enhances protein content and nutrient uptake and stimulates phenylalanine ammonia-lyase and nitrate reductase in lettuce leaves.
3.2. Phytohormones
The increase in plant growth due to vermicompost application is primarily attributed to the continuous availability of macro and micronutrients, along with biological effects related to enzymatic activities and plant hormones [105]. The combined activities of EWs and microbes secrete phytohormones such as auxins, gibberellins, and cytokinin during the vermicomposting process [106]. The presence of cytokinins in VC tea and indole-3-acetic acid (auxins) in VC-derived humic acids was detected by GC-MS [107]. As a result, the highest concentrations of indole-3-acetic acid, gibberellic acid, and cytokinins were 7.37, 5.7, and 2.8 mg kg−1, respectively, in vermicompost (cow manure + leaf litter + tannery waste) [6]. Various phytohormones in garden waste VCL were analyzed using HPLC-MS [108]. The primary hormonal constituents included cytokinins (60% of N6-isopentenyl adenine), indole-3-acetic acid, gibberellins, and brassinosteroids. Brassinosteroids are a group of essential steroid hormones vital for plant growth and development. Their signaling roles promote cell division and expansion and are crucial in reproduction and etiolation [109]. It was reviewed that various biofertilizers could provide plant hormones and hormone-like elements beneficial for organic and conventional plant cultivation [110]. The physiological functions of phytohormones are described in Figure 3.
The application of vermicompost as an alternative to synthetic fertilizers stimulates plant production. Steffen et al. [111] studied the efficiency of VC in flowering and productivity of tomatoes by partially replacing synthetic fertilizers with VC, considering five treatments consisting of 0, 10, 20, 40, and 50% vermicompost substitution with synthetic fertilizer. Vermicompost at application rates of 40 and 50% resulted in early flowering and fruit development in tomatoes, higher fruit mass, and production. Arancon and coworkers [81] investigated the impact of VT on lettuce and tomato growth in a hydroponic system. VT significantly increased lettuce yield at nutrient concentrations of 25% and 50% of the recommended rate and improved tomato yield at 50% of the recommended rate of VC tea. The presence of trace concentrations of auxins, gibberellins, cytokinins, and humic acids in VT are responsible factors that enhance tomato and lettuce yield in a hydroponic system. Other experiments were performed by Sardoei et al. [112], who determined the efficacy of VC on the growth and flowering of marigolds (Calendula officinalis) under glasshouse conditions. The seeds were germinated and grown in 10, 20, 30, 40, 50, and 60% of vermicompost media. VC had optimistic effects on the total parameters compared to control and amended media. The maximum root volume and fresh petal weight were achieved in 30% VC treatment. However, plant performance was most remarkable in 60% VC amended medium. Similarly, Suthar et al. [75] examined the effect of vermiwash on seed the germination, sapling growth, and biochemistry of Trigonella foenumgraecum and Cyamopsis tertagonoloba in a laboratory environment. Four trial solutions—vermiwash (100%), vermiwash (50%), urea (5% solution), and distilled water—were used in the study. The highest germination rate was observed with 50% vermiwash, while maximum plant growth parameters, chlorophyll content, total soluble sugars, total proteins, and starch were detected in plant tissues treated with 100% vermiwash.
3.3. Microbial Activity
Vermicompost possesses various properties, including the ability to increase soil microbial activity by enhancing oxygen availability, regulating soil temperature, improving porosity, infiltration, and nutrient content, and boosting crop yield and quality [113]. VC promotes beneficial microbes and their diversity in the soil. It enhances soil biodiversity by supporting valuable microbes, which, in turn, increase plant growth by producing plant enzymes and hormones and controlling plant pathogens and nematodes. EW burrows coated with VC provide an excellent medium for harboring nitrogen-fixing bacteria in the soil. The growth of gram-negative bacteria was significantly improved after vermicompost treatment [114]. Vermicompost (VC) stimulates microbes to fix nitrogen into mucoprotein, which prevents nitrogen leaching into the soil and reduces the C/N ratio [115]. A study investigating the combined effect of vermicompost and plant growth-promoting rhizobacteria (PGPR) on tomato and spinach quality was conducted by Song et al. [116]. The results showed that VC enhanced the beneficial effects of PGPR on the soil and crops. The synergistic effect of VC and PGPR increased soluble protein in spinach and vitamin C in tomatoes. Karnwal et al. [117] cultured zinc-solubilizing bacteria (ZSB) from VC for numerous plant growth-promoting traits in tomatoes. Thirty isolates from VC were screened for Zn solubilization activity. Significant Zn solubilization (26.8 mg L−1) and Zn contents (2.87 mg/100 g) in fruit were obtained by applying bacterial isolates from VC. Thus, the use of these bacteria may provide adequate Zn availability, which results in better plant growth in a sustainable way. The efficiency of cellulolytic microbes-enriched wheat straw VC on physiological and biochemical aspects of wheat under various soil moisture conditions was investigated by Ahmad et al. [118]. Their results revealed that wheat growth was affected by moisture scarcity conditions, but the use of VC boosted the growth performance. Under drought, VC at 6-ton ha−1 treatment augmented the root and shoot biomass, photosynthetic rate, relative water contents, and antioxidant enzymes in wheat cultivars over the control. Concisely, the use of cellulolytic microbe-enriched VC at 6-ton ha−1 was useful in lessening the harmful impacts of drought on wheat. Vermicompost amendment is reported to boost the total dry matter of tomato plants (24.0%), chickpea plants (65.2%), and onion nuts (12.5%) [119,120]. The application of microbial-enriched VC has an imperative influence on okra (Abelmoschus esculentus L.) growth [121]. The growth parameters were considerably higher in the plants mixed with vermicompost. However, the growth characteristics were more responsive to VC enriched with Azospirillum. Biochemical parameters such as carotenoid, total chlorophyll, and protein contents of okra showed a positive impact by adding enriched VC. Parastesh et al. [122] studied the residual and direct impacts of phosphate solubilizing bacteria (PSB)-enriched VC on the enhancement of phosphorus uptake by wheat and tomato plant. The PSB-enriched VC significantly enhanced the phosphorus concentration in wheat shoots (20–39%) and tomatoes (26–53%) treated with vermicompost. These findings highlight the significance of vermicompost enriched with beneficial microbes as organic fertilizer for cereals and vegetable cultivation.
4. Effects of Vermicompost as Bio-Alleviator of Abiotic and Biotic Stress
Plant stress is categorized into abiotic and biotic stress. Abiotic stress on plants can be either physical or chemical, such as drought and salinity, whereas biotic stress is plant exposure to biological agents such as insects and diseases [123]. Recent findings have revealed that EWs’ cast can tolerate, decrease, and ameliorate the salinity [22]. Various EWs species, such as Eutyphoeus incommodus, demonstrated high survival in the saline environment and can be used to reclaim soil salinity [124]. This is due to the humic substances in VC, which react with plant roots and soil organic substances, thus enhancing salinity tolerance.
4.1. Soil Salinity
Salinity is the most important growth limiting factor and a threat to crop production. Saline soil contains higher concentrations of soluble salts, affecting the plant by lowering the nutrient uptake and plant water relations. Plant growth is hindered by salinity, resulting in extreme decline in its yield, which relates to a decline in photosynthetic efficiency [125]. Photosynthetic elements, involving chlorophylls and carotenoids, are also influenced by salinity. In an experiment conducted by Alamer et al. [126], total carotenoid and chlorophyll a, b were significantly reduced under saline conditions. Decreased water absorption caused by plants due to the osmotic effect of salinity harms their development. Further plant exposure to salinity leads to the absorption and accumulation of Na+ and Cl− and causes ion toxicity in plant tissues. Similarly, higher Na+ and Cl− contents in soil solution reduced the absorption of essential minerals such as K+, Ca2+, and Mg2+, causing nutritional imbalance [127]. However, the use of VC assisted the maize seedlings to cope with the toxic effects of salinity, which led to an increase in photosynthetic pigments. Several studies showed that VC reduces the adverse effects of salt stress on many crop plants [128,129]. VC improved plant tolerance by stimulating enzymatic activities, protein synthesis, and boosting the physiological functions of plants [130]. The growth of maize, barley, rice, wheat, and soybean improved by developing tolerance against NaCl stress using vermicompost-inhibited microorganisms such as Azotobacter chroococcum and Bacillus subtilis [131]. VC and its derivatives reduced the impact of salinity and improved plant performance in Table 2.
4.2. Drought Stress
Drought stress is deemed the most damaging abiotic stress to plants as it affects food security. It negatively influences plant–water relations at cell and systemic levels, affecting the farming community with economic losses. It reduces the leaves’ turgor pressure, thus resulting in a reduction in cell volume, elongation, and division [143]. Additionally, the unavailability of water lowers CO2 absorption and impedes glucose synthesis, decreasing the photosynthetic efficiency and crop yield [144]. Drought stress reduced the number of tillers, spikes, grains, and grains weight in wheat and barley [145,146]. In the case of millet (Pennisetum glaucum L.), drought stress affected silk development and increased the anthesis to the silking gap, thus reducing the millet yield [147]. Drought stress produces reactive oxygen species (ROS) and intensifies chlorophyll degradation. VC contains Fe (iron), the prosthetic group of ROS-consuming enzymes such as catalase, peroxidase, and superoxide dismutase [148]. VC also contains other macro and micronutrients such as N, P, K, Ca, Mg, and Zn. The reduction in chlorophyll contents due to drought stress may increase the nitrogen metabolism for proline synthesis during osmotic regulation [149]. VC amendment at 10 and 20% considerably increased the chlorophyll contents, intracellular CO2 concentration, net photosynthetic rate, and quantum yield of PSII photochemistry [150]. Dehmordy et al. [151] set up an experiment to study the impact of VC on growth performance and drought tolerance of Olive (Olea europaea L.). Their findings showed that drought stress was reduced, and the addition of VC boosted the vegetative growth as compared to plants grown in unamended soil. VC significantly decreased the ion leakage and increased proline and chlorophyll contents and antioxidant enzyme activity. The interaction of VC (30%) and water stress on CO2 assimilation showed a positive response under severe water stress conditions [152]. VC with higher water holding capacity and porosity reduced the destructive effects of water stress, such as stomatal closure and low CO2 assimilation [153]. Such a decrease in stomatal closure and an increase in CO2 supply is required for the RuBisCO enzyme, which ultimately increases CO2 assimilation. Moreover, humic acid is a structural part of VC and negatively charged groups are responsible for positively charged mineral elements. Some of these elements have a significant role in the stimulation of antioxidant enzymes and photosynthetic enzymes [154]. During drought stress, transpiration is also reduced to prevent water loss, but it also hinders the active transport in phloem and passive transport in xylem vessels. Reduction in transpiration resulted in the blocking of water and nutrients uptake from roots. Mycorrhizal fungi in VC improve the roots’ water uptake [155]. The combined application of mycorrhiza and VC encouraged plant growth by triggering the photosynthesis machinery and nutrient absorption, leading to improve proteins, total soluble sugars, and antioxidant enzyme activities in the leaf and root of quinoa [156]. The availability of nutrients such as phosphorus in the rhizosphere can improve protein synthesis, photosynthesis, membrane transport, cell division, and elongation, which increases plant biomass. VC application to rapeseed cultivars increased linoleic acid and palmitic acid concentrations; however, it decreased linolenic acid contents [157]. Due to the high potential in improving soil properties and nutrient availability for plants, VC would be recommended to compensate for the decrease in plant yield imposed by water stress. The performance of different plants grown in VC-amended soil under drought stress is explained in Table 3.
4.3. Insect Pests
Organic amendments have been proposed as an efficient alternative practice to improve soil and plant health by suppressing phytopathogens such as plant-parasitic nematodes [163]. Now, more data have explained VC as a superior organic amendment to compost in terms of soil properties and plant health [164]. VC could boost plant growth and tolerance against soil-borne and above-ground pests [165]. Further findings indicated that the humic substances and phytohormones in VC could control pest infestation. Rostami et al. [166] studied the effect of vermicompost extract (VE) and vermiwash (VW) on root-knot nematode in the cucumber plant. The VE and VW at 10% and 100% concentration increased the inhibition rate of egg hatching and Meloidogyne javanica larvae mortality examined after 24, 48, and 72 h. The phytopathogens-suppressing potential of vermicompost and its derivatives are elaborated in Table 4. The mechanisms by which VC and its derivatives suppress pest attacks are various but anticipated simultaneously. Based on research outcomes, four suppression pest suppression mechanisms can be found liable; these are (I) release of phenolic substances; (II) predatory nematodes; (III) release of antibiotics; and (IV) increase in nutrient availability. Phenols are biomolecules produced with phenol oxidase enzymes. VC boosts the plant’s uptake of soluble phenolic substances for pest resistance [167]. Due to their toxicity to nematodes, phenolic substances act as repellents and deterrents. The presence of phenolic substances in plants reduces the palatability of plants, thus shifting the pest [168]. The chlorogenic acid and gallic acid in Thymus vulgaris are considered to suppress Phytophthora infestans and Fusarium oxysporum [21]. The presence of flavonoids and humic acid has been described as controlling Bemisia tabaci and Xanthomonas campestris in Cyamopsis tetragonoloba [169]. VC and its derivatives’ application increased the population and diversity of predatory nematodes in soil [170]. These predators adversely affect the pest population by attacking them directly. Renčo et al. [171] described a significant decrease in eggs and juveniles of Globodera pallida and G. rostochiensis (potato cyst nematodes) with VT application. Similar findings were reported by Sivasabari et al. [172] for Rotylenchulus reniformis and Meloidogyne incognita due to the presence of beneficial microbes in VT. Huang et al. [173] and Poveda et al. [174] studied the effect of VW, liquid VC, and coelomic fluid from E. fetida to control root-knot disease in vitro and greenhouse. The results revealed that the three treatments (VW, liquid VC, and coelomic fluid) inhibited egg hatching. The coelomic fluid contains bioactive molecules that negatively influence the physiological functions of Meloidogyne javanica larva. VC is enriched with microbes (Bacillus and Pseudomonas sp.) and fungi (Trichoderma sp.) which releases cuticle degrading proteases to inhibit nematode multiplication. The microbes in the rhizosphere compete with pathogens for nutrients and stimulate the plant to produce secondary metabolites under stress conditions, thus inducing systemic resistance in plants [175]. Soil amendment with an organic fertilizer such as VC has good buffering capacity and nutrient stability, establishing an unfavorable environment for pests [176]. Macro and micronutrients present in VC increase the potential of plants to defend themselves from pest attacks.
4.4. Plant Diseases
Bacteria, viruses, and fungi are the primary causes of plant diseases, with fungi being the most common causal organism. Among the various secondary effects of VC on plants, disease suppression is the most significant. This is mainly associated with the release of coelomic fluid from EWs, which kills pathogens in waste material during the vermicomposting process. Consequently, VC possesses insecticidal and antifungal properties due to coelomic fluid and other bioactive compounds released by EWs, making it effective in suppressing insects and diseases. Bioactive compounds are produced from numerous amino acid chains to combat specific soil pathogens [187]. CF is secreted from dorsal pores of EWs and functions as a defense mechanism due to its anti-microbial properties [188].
Streptomycin is an antibiotic used to control crop diseases, while salicylic acid effectively induces phytopathogens’ resistance mechanism. In recent years, the use of VC and VT as bio-control agents has increased considerably. Hakim et al. [189] investigated the effect of compost, VC, and tea on bacterial vascular wilt in tomato seedlings. VC at concentrations of 30% and 40% by volume reduced bacterial vascular wilt at 60% and 66.67%, respectively. The treatment containing 30% VC tea by volume decreased disease severity by 53.33%. In addition to suppressing bacterial diseases, VC can also suppress fungal diseases by reducing sporulation, retarding the growth of pathogenic fungi, and thus reducing infection. Fungal diseases are commonly associated with acute injury and plant mortality. The extensive use of fungicides has led to environmental pollution and the development of resistance against antifungal chemicals [190]. EWs’ secretions (Lumbricin-PG with antifungal properties) are attached to the vermicompost surface and protect plants against fungal attack. This is also confirmed by You et al. [191], where a bioactive metabolite (ergosterol peroxide) with antifungal activity against R. solani AG1-IB was secreted by microbes during vermicomposting of bamboo. Amooaghaie et al. [192] reported that VC is an efficient biocontrol agent against Phytophthora infestans and Fusarium oxysporum. The application of VC to Brassica juncea crops inhibited the fungal mycelium of Alternaria brassicae at the highest rate of 47.43%. Mu et al. [193] isolated Bacillus subtilis M29 (an antifungal bacterial strain) from vermicompost. These bacterial strains produced several volatile compounds that inhibited Botrytis cinerea growth (grey mold). A study conducted by Tian et al. [194] investigated the effectiveness of vermicompost, biochar, and their combination on the occurrence of Fusarium root rot. The combined application of VC, biochar, and Fusarium (VBF) reduced the disease index by up to 80.96%, compared to the control, and a higher concentration of vanillin (a phenolic acid from root exudates) was observed in VBF treatment. The vanillin concentration showed a negative correlation with the disease index. Vermicompost contains several Rhizobacteria species that effectively suppress fungal growth on beans by 50–60% [195]. Various mechanisms have been identified as being responsible for disease suppression, as shown in Figure 4, including nutrient competition, antibiosis, hyperparasitism, and induced systemic resistance [196].
Competition is the primary factor in disease suppression mechanisms, and its effectiveness is typically associated with microbial population and activity in the soil. A higher population of microbes reduces the availability of nutrients, space, and energy for pathogens. Under Fe deficiency conditions, some microbes, such as Pythium species, produce siderophores, which reduce disease occurrence by decreasing Fe availability to phytopathogens [197]. Antibiosis is an interaction where one organism affects others by secreting specific antibiotics or metabolites [198]. Pseudomonas in vermicompost produces several metabolites such as phenazines and C-acelyphloroglucinols with broad-spectrum action against fungal pathogens, [199]. Hyperparasitism is a phenomenon in which phylogenetically unrelated microorganisms colonize pathogens, leading to their death. During the composting process, available cellulose and glucose activate Trichoderma sp., which secretes chitinase enzyme to parasitize Rhizoctonia solani [200]. Induced systemic resistance is a state of increased defensive capacity that plants develop when stimulated by any phytopathogen. The application of compost reduces disease severity and induces systemic resistance against pathogens [201]. Sang et al. [202] reported that compost tea protects cucumber and pepper plants from anthracnose disease due to the induced systemic resistance mechanism. Reactive oxygen species (ROS), callose deposition, and Ca2+ play significant roles in activating this mechanism. Callose is a β-1, 3 glucan polymer that accumulates in vulnerable parts of plant cell walls during pathogen attack. Pseudomonas fluorescens strain 63–28 enhances tomato plant resistance to Fusarium oxysporum by immediately accumulatingcallose and chitinases [203]. Pyocyanin produced by Pseudomonas aeruginosa 7NSK2 increases H2O2 (a reactive oxygen species) in leaves and induces resistance to blast disease (Magnaporthe grisea) [204]. The beneficial root-fungus Mortierella hyalina stimulates the Ca2+-dependent signaling pathway to develop resistance against Alternaria brassicae [205].
5. Conclusions
This review elaborates various mechanisms involved in plant growth promotion and the mitigation of biotic and abiotic stresses using vermicompost and its derivatives. The application of vermicompost enhances soil health and crop productivity due to improved nutrient uptake, the presence of humic substances, phytohormones, and enhanced microbial activities in VC. The current review also addresses questions related to the mitigation of drought, salinity, insect pests, and plant diseases using vermicompost. With higher water-holding capacity and porosity, VC reduces the detrimental effects of water stress. It aids in improving plant tolerance against salinity stress by stimulating enzymatic activities. VC suppresses plant pests and diseases by various mechanisms, such as competition, predation, releasing metabolites, and inducing systemic resistance against foliar pathogens. However, the possibility of using VC for the reduction of toxic elements needs to be addressed. The role of EWs in improving organic matter decomposition, soil structure, soil fertility, microbial population, and diversity is also discussed. VC and its derivatives are environmentally safe products and should be used to mitigate phytopathogens. This review gives an updated perspective on the possible use of vermicompost to mitigate biotic and abiotic stresses such as drought, salinity, insect pests, and diseases on plants.
Author Contributions
Conceptualization: S.u.R. and M.B.; Data curation: S.u.R., F.D.C. and M.B.; Writing—original draft preparation: F.D.C.; Writing—reviewing and editing: S.u.R., A.A., M.B. and F.P.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the project “National Biodiversity Future Center—NBFC”(project code: CN00000033).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
VC: Vermicompost; VT: Vermi-tea; HS: Humic substances; VW: Vermiwash; OM: Organic matter; HA: Humic acid; EWs: Earthworms; RCF: Relative cast fertility; GAM: Gut-associated microbes; VCL: Vermicompost leachates; PGPR: Plant growth-promoting rhizobacteria; ZSB: Zinc-solubilizing bacteria; ROS: Reactive oxygen species; VE: Vermicompost extract; VBF: VC, biochar, and Fusarium.
References
- Erisman, J.W.; Sutton, M.A.; Galloway, J.; Klimont, Z.; Winiwarter, W. How a century of ammonia synthesis changed the world. Nat. Geosci. 2008, 1, 636–639. [Google Scholar] [CrossRef]
- Kumar, A.; Bohra, B. Green technology in relation to sustainable agriculture. In Green Technologies for Sustainable Agriculture; Daya Publishing House: Delhi, India, 2006. [Google Scholar]
- De Castro, F.; Vergaro, V.; Benedetti, M.; Baldassarre, F.; Del Coco, L.; Dell’Anna, M.M.; Mastrorilli, P.; Fanizzi, F.P.; Ciccarella, G. Visible light-activated water-soluble platicur nanocolloids: Photocytotoxicity and metabolomics studies in cancer cells. ACS Appl. 2020, 3, 6836–6851. [Google Scholar] [CrossRef] [PubMed]
- De Castro, F.; Stefàno, E.; Migoni, D.; Iaconisi, G.N.; Muscella, A.; Marsigliante, S.; Benedetti, M.; Fanizzi, F.P. Synthesis and Evaluation of the Cytotoxic Activity of Water-Soluble Cationic Organometallic Complexes of the Type [Pt (η1−C2H4OMe)(L)(Phen)]+(L = NH3, DMSO; Phen = 1,10-Phenanthroline). Pharmaceutics 2021, 13, 642. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, M.; Antonucci, D.; De Castro, F.; Girelli, C.R.; Lelli, M.; Roveri, N.; Fanizzi, F.P. Metalated nucleotide chemisorption on hydroxyapatite. J. Inorg. Biochem. 2015, 153, 279–283. [Google Scholar] [CrossRef]
- Ravindran, B.; Wong, J.W.; Selvam, A.; Sekaran, G. Influence of microbial diversity and plant growth hormones in compost and vermicompost from fermented tannery waste. Bioresour. Technol. 2016, 217, 200–204. [Google Scholar] [CrossRef]
- Datta, S.; Singh, J.; Singh, S.; Singh, J. Earthworms, pesticides and sustainable agriculture: A review. Environ. Sci. Pollut. Res. 2016, 23, 8227–8243. [Google Scholar] [CrossRef]
- Tripathi, Y.; Hazaria, P.; Kaushik, P.; Kumar, A. Vermitechnology and waste management. In Verms and Vermitechnology, SB Nangia; APH Publishing Corp: New Delhi, India, 2005; pp. 9–21. [Google Scholar]
- Coria-Cayupaán, Y.S.; de Pinto, M.A.I.S.; Nazareno, M.A. Variations in bioactive substance contents and crop yields of lettuce (Lactuca sativa L.) cultivated in soils with different fertilization treatments. J. Agric. Food Chem. 2009, 57, 10122–10129. [Google Scholar] [CrossRef] [PubMed]
- Goutam, K.C.; Goutam, B.; Susanta, K.C. The effect of vermicompost and other fertilizers on cultivation of tomato plants. J. Hortic. For. 2011, 3, 42–45. [Google Scholar]
- Ruz-Jerez, B.; Ball, P.R.; Tillman, R. Laboratory assessment of nutrient release from a pasture soil receiving grass or clover residues, in the presence or absence of Lumbricus rubellus or Eisenia fetida. Soil Biol. Biochem. 1992, 24, 1529–1534. [Google Scholar] [CrossRef]
- Parkin, T.B.; Berry, E.C. Nitrogen transformations associated with earthworm casts. Soil Biol. Biochem. 1994, 26, 1233–1238. [Google Scholar] [CrossRef]
- Adhikary, S. Vermicompost, the story of organic gold: A review. Agric. Sci. 2012, 3, 905–917. [Google Scholar] [CrossRef]
- Atiyeh, R.; Arancon, N.; Edwards, C.; Metzger, J. Influence of earthworm-processed pig manure on the growth and yield of greenhouse tomatoes. Bioresour. Technol. 2000, 75, 175–180. [Google Scholar] [CrossRef]
- Yatoo, A.M.; Rasool, S.; Ali, S.; Majid, S.; Rehman, M.U.; Ali, M.; Eachkoti, R.; Rasool, S.; Rashid, S.M.; Farooq, S. Vermicomposting: An eco-friendly approach for recycling/management of organic wastes. In Bioremediation and Biotechnology; Springer: Cham, Switzerland, 2020; pp. 167–187. [Google Scholar]
- Ayyobi, H.; Hassanpour, E.; Alaqemand, S.; Fathi, S.; Olfati, J.; Peyvast, G. Vermicompost leachate and vermiwash enhance French dwarf bean yield. Int. J. Veg. Sci. 2014, 20, 21–27. [Google Scholar] [CrossRef]
- Bidabadi, S.S. Waste management using vermicompost derived liquids in sustainable horticulture. Trends Hortic. 2018, 1, 175. [Google Scholar]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. 2017, 4, 5. [Google Scholar] [CrossRef]
- Puglisi, E.; Pascazio, S.; Suciu, N.; Cattani, I.; Fait, G.; Spaccini, R.; Crecchio, C.; Piccolo, A.; Trevisan, M. Rhizosphere microbial diversity as influenced by humic substance amendments and chemical composition of rhizodeposits. J. Geochem. Explor. 2013, 129, 82–94. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Vajranabhaiah, S. Biological activity of earthworm casts: An assessment of plant growth promotor levels in the casts. Proc. Anim. Sci. 1986, 95, 341–351. [Google Scholar] [CrossRef]
- Amooaghaie, R.; Golmohammadi, S. Effect of vermicompost on growth, essential oil, and health of Thymus Vulgaris. Compos. Sci. Util. 2017, 25, 166–177. [Google Scholar] [CrossRef]
- Sahab, S.; Suhani, I.; Srivastava, V.; Chauhan, P.S.; Singh, R.P.; Prasad, V. Potential risk assessment of soil salinity to agroecosystem sustainability: Current status and management strategies. Sci. Total Environ. 2021, 764, 144164. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Hussain, D.; Bellitürk, K.; Javed, T.; Hussain, S.; Bashir, S.; Raza, A.; Alotaibi, S.; Kalaji, H.M. Rice straw vermicompost enriched with cellulolytic microbes ameliorate the negative effect of drought in wheat through modulating the morpho-physiological attributes. Front. Environ. Sci. 2022, 10, 497. [Google Scholar] [CrossRef]
- Basco, M.; Bisen, K.; Keswani, C.; Singh, H. Biological management of Fusarium wilt of tomato using biofortified vermicompost. Mycosphere 2017, 8, 467–483. [Google Scholar] [CrossRef]
- Manandhar, T.; Yami, K. Biological control of foot rot disease of rice using fermented products of compost and vermicompost. Sci. World J. 2008, 6, 52–57. [Google Scholar] [CrossRef]
- Pathma, J.; Sakthivel, N. Microbial diversity of vermicompost bacteria that exhibit useful agricultural traits and waste management potential. SpringerPlus 2012, 1, 26. [Google Scholar] [CrossRef]
- Pattnaik, S.; Reddy, M.V. Nutrient status of vermicompost of urban green waste processed by three earthworm species—Eisenia fetida, Eudrilus eugeniae, and Perionyx excavatus. Appl. Environ. Soil Sci. 2010, 2010, 967526. [Google Scholar] [CrossRef]
- Ahmed, N.; Al-Mutairi, K.A. Earthworms Effect on Microbial Population and Soil Fertility as Well as Their Interaction with Agriculture Practices. Sustainability 2022, 14, 7803. [Google Scholar] [CrossRef]
- Fragoso, C.; Kanyonyo, J.; Moreno, A.; Senapati, B.K.; Blanchart, E.; Rodriguez, C. A survey of tropical earthworms: Taxonomy, biogeography and environmental plasticity. Earthworm Manag. Trop. Agroecosyst. 1999, 1–26. [Google Scholar]
- Eggleton, P.; Inward, K.; Smith, J.; Jones, D.T.; Sherlock, E. A six year study of earthworm (Lumbricidae) populations in pasture woodland in southern England shows their responses to soil temperature and soil moisture. Soil Biol. Biochem. 2009, 41, 1857–1865. [Google Scholar] [CrossRef]
- Vivas, A.; Moreno, B.; Garcia-Rodriguez, S.; Benitez, E. Assessing the impact of composting and vermicomposting on bacterial community size and structure, and microbial functional diversity of an olive-mill waste. Bioresour. Technol. 2009, 100, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Gopal, M.; Bhute, S.S.; Gupta, A.; Prabhu, S.; Thomas, G.V.; Whitman, W.B.; Jangid, K. Changes in structure and function of bacterial communities during coconut leaf vermicomposting. Antonie Van Leeuwenhoek 2017, 110, 1339–1355. [Google Scholar] [CrossRef]
- Edwards, C.A.; Bohlen, P.J. Biology and Ecology of Earthworms; Springer Science & Business Media: London, UK, 1996; Volume 3. [Google Scholar]
- Vickers, N.J. Animal communication: When i’m calling you, will you answer too? Curr. Biol. 2017, 27, R713–R715. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, D.; Hodson, M.E.; Eggleton, P.; Kirk, C. Earthworm induced mineral weathering: Preliminary results. Eur. J. Soil Biol. 2007, 43, S176–S183. [Google Scholar] [CrossRef]
- Al-Maliki, S.; Scullion, J. Interactions between earthworms and residues of differing quality affecting aggregate stability and microbial dynamics. Appl. Soil Ecol. 2013, 64, 56–62. [Google Scholar] [CrossRef]
- Ritsema, C.J.; Dekker, L. Preferential flow in water repellent sandy soils: Principles and modeling implications. J. Hydrol. 2000, 231, 308–319. [Google Scholar] [CrossRef]
- Atiyeh, R.M.; Domínguez, J.; Subler, S.; Edwards, C.A. Changes in biochemical properties of cow manure during processing by earthworms (Eisenia andrei, Bouché) and the effects on seedling growth. Pedobiologia 2000, 44, 709–724. [Google Scholar] [CrossRef]
- Lavelle, P.; Charpentier, F.; Villenave, C.; Rossi, J.-P.; Derouard, L.; Pashanasi, B.; André, J.; Ponge, J.-F.; Bernier, N. Effects of earthworms on soil organic matter and nutrient dynamics at a landscape scale over decades. Earthworm Ecol. 2004, 2, 145–160. [Google Scholar]
- Alegre, J.; Pashanasi, B.; Lavelle, P. Dynamics of soil physical properties in Amazonian agroecosystems inoculated with earthworms. Soil Sci. Soc. Am. J. 1996, 60, 1522–1529. [Google Scholar] [CrossRef]
- Ernst, G.; Henseler, I.; Felten, D.; Emmerling, C. Decomposition and mineralization of energy crop residues governed by earthworms. Soil Biol. Biochem. 2009, 41, 1548–1554. [Google Scholar] [CrossRef]
- Lubbers, I.M.; Pulleman, M.M.; Van Groenigen, J.W. Can earthworms simultaneously enhance decomposition and stabilization of plant residue carbon? Soil Biol. Biochem. 2017, 105, 12–24. [Google Scholar] [CrossRef]
- Aira, M.; Domínguez, J. Earthworm effects without earthworms: Inoculation of raw organic matter with worm-worked substrates alters microbial community functioning. PLoS ONE 2011, 6, e16354. [Google Scholar] [CrossRef] [PubMed]
- Aira, M.; Monroy, F.; Domínguez, J. Changes in bacterial numbers and microbial activity of pig slurry during gut transit of epigeic and anecic earthworms. J. Hazard. Mater. 2009, 162, 1404–1407. [Google Scholar] [CrossRef] [PubMed]
- Chapuis-Lardy, L.; Bayon, R.-C.L.; Brossard, M.; López-Hernández, D.; Blanchart, E. Role of soil macrofauna in phosphorus cycling. In Phosphorus in Action; Springer: Berlin/Heidelberg, Germany, 2011; pp. 199–213. [Google Scholar]
- Blouin, M.; Hodson, M.E.; Delgado, E.A.; Baker, G.; Brussaard, L.; Butt, K.R.; Dai, J.; Dendooven, L.; Pérès, G.; Tondoh, J. A review of earthworm impact on soil function and ecosystem services. Eur. J. Soil Sci. 2013, 64, 161–182. [Google Scholar] [CrossRef]
- Seeber, J.; Seeber, G.; Langel, R.; Scheu, S.; Meyer, E. The effect of macro-invertebrates and plant litter of different quality on the release of N from litter to plant on alpine pastureland. Biol. Fertil. Soils 2008, 44, 783–790. [Google Scholar] [CrossRef]
- Sampedro, L.; Domínguez, J. Stable isotope natural abundances (δ13C and δ15N) of the earthworm Eisenia fetida and other soil fauna living in two different vermicomposting environments. Appl. Soil Ecol. 2008, 38, 91–99. [Google Scholar] [CrossRef]
- Thakuria, D.; Schmidt, O.; Finan, D.; Egan, D.; Doohan, F.M. Gut wall bacteria of earthworms: A natural selection process. ISME J. 2010, 4, 357–366. [Google Scholar] [CrossRef]
- De Menezes, A.B.; Prendergast-Miller, M.T.; Macdonald, L.M.; Toscas, P.; Baker, G.; Farrell, M.; Wark, T.; Richardson, A.E.; Thrall, P.H. Earthworm-induced shifts in microbial diversity in soils with rare versus established invasive earthworm populations. FEMS Microbiol. Ecol. 2018, 94, fiy051. [Google Scholar] [CrossRef]
- Fujii, K.; Ikeda, K.; Yoshida, S. Isolation and characterization of aerobic microorganisms with cellulolytic activity in the gut of endogeic earthworms. Int. Microbiol. 2012, 15, 121–130. [Google Scholar] [PubMed]
- Johnsen, A.R.; Wick, L.Y.; Harms, H. Principles of microbial PAH-degradation in soil. Environ. Pollut. 2005, 133, 71–84. [Google Scholar] [CrossRef]
- Chan, K.; Baker, G.; Conyers, M.; Scott, B.; Munro, K. Complementary ability of three European earthworms (Lumbricidae) to bury lime and increase pasture production in acidic soils of south-eastern Australia. Appl. Soil Ecol. 2004, 26, 257–271. [Google Scholar] [CrossRef]
- Van Groenigen, J.W.; Van Groenigen, K.J.; Koopmans, G.F.; Stokkermans, L.; Vos, H.M.; Lubbers, I.M. How fertile are earthworm casts? A meta-analysis. Geoderma 2019, 338, 525–535. [Google Scholar] [CrossRef]
- Le Bayon, R.-C.; Milleret, R. Effects of earthworms on phosphorus dynamics–a review. Dyn. Soil Dyn. Plant 2009, 3, 21–27. [Google Scholar]
- Nuutinen, V.; Pitkänen, J.; Kuusela, E.; Widbom, T.; Lohilahti, H. Spatial variation of an earthworm community related to soil properties and yield in a grass–clover field. Appl. Soil Ecol. 1998, 8, 85–94. [Google Scholar] [CrossRef]
- Curry, J.P.; Schmidt, O. The feeding ecology of earthworms–a review. Pedobiologia 2007, 50, 463–477. [Google Scholar] [CrossRef]
- Neilson, R.; Boag, B. Feeding preferences of some earthworm species common to upland pastures in Scotland. Pedobiologia 2003, 47, 1–8. [Google Scholar] [CrossRef]
- Lowe, C.N.; Butt, K.R. Influence of food particle size on inter-and intra-specific interactions of Allolobophora chlorotica (Savigny) and Lumbricus terrestris: The 7th international symposium on earthworm ecology Cardiff Wales·2002. Pedobiologia 2003, 47, 574–577. [Google Scholar]
- Satchell, J.E. Lumbricidae. In Soil Biology; Burges, A., Raw, F., Eds.; Academic Press: London, UK, 1967; pp. 259–322. [Google Scholar]
- Moody, S.; Briones, M.; Piearce, T.; Dighton, J. Selective consumption of decomposing wheat straw by earthworms. Soil Biol. Biochem. 1995, 27, 1209–1213. [Google Scholar] [CrossRef]
- Drake, H.L.; Horn, M.A. As the worm turns: The earthworm gut as a transient habitat for soil microbial biomes. Annu. Rev. Microbiol. 2007, 61, 169–189. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Tiwari, B.; Mishra, R. Microbial populations, enzyme activities and nitrogen-phosphorus-potassium enrichment in earthworm casts and in the surrounding soil of a pineapple plantation. Biol. Fertil. Soils 1989, 8, 178–182. [Google Scholar] [CrossRef]
- Decaëns, T.; Rangel, A.; Asakawa, N.; Thomas, R.J. Carbon and nitrogen dynamics in ageing earthworm casts in grasslands of the eastern plains of Colombia. Biol. Fertil. Soils 1999, 30, 20–28. [Google Scholar] [CrossRef]
- Brossard, M.; Lavelle, P.; Laurent, J.-Y. Digestion of a vertisol by the endogeic earthworm Polypheretima elongata, Megascolecidae, increases soil phosphate extractibility. Eur. J. Soil Biol. 1996, 32, 107. [Google Scholar]
- Edwards, C.; Lofty, J. Biology of Earthworms; Chapman and Hall: London, UK, 1977. [Google Scholar]
- Hong, S.W.; Lee, J.S.; Chung, K.S. Effect of enzyme producing microorganisms on the biomass of epigeic earthworms (Eisenia fetida) in vermicompost. Bioresour. Technol. 2011, 102, 6344–6347. [Google Scholar] [CrossRef]
- Schönholzer, F.; Hahn, D.; Zeyer, J. Origins and fate of fungi and bacteria in the gut of Lumbricus terrestris L. studied by image analysis. FEMS Microbiol. Ecol. 1999, 28, 235–248. [Google Scholar] [CrossRef]
- Pedersen, J.C.; Hendriksen, N.B. Effect of passage through the intestinal tract of detritivore earthworms (Lumbricus spp.) on the number of selected Gram-negative and total bacteria. Biol. Fertil. Soils 1993, 16, 227–232. [Google Scholar] [CrossRef]
- Monroy, F.; Aira, M.; Domínguez, J. Reduction of total coliform numbers during vermicomposting is caused by short-term direct effects of earthworms on microorganisms and depends on the dose of application of pig slurry. Sci. Total Environ. 2009, 407, 5411–5416. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.G.; Barois, I.; Lavelle, P. Regulation of soil organic matter dynamics and microbial activityin the drilosphere and the role of interactionswith other edaphic functional domains. Eur. J. Soil Biol. 2000, 36, 177–198. [Google Scholar] [CrossRef]
- Hoeffner, K.; Monard, C.; Santonja, M.; Cluzeau, D. Feeding behaviour of epi-anecic earthworm species and their impacts on soil microbial communities. Soil Biol. Biochem. 2018, 125, 1–9. [Google Scholar] [CrossRef]
- Loera-Muro, A.; Troyo-Diéguez, E.; Murillo-Amador, B.; Barraza, A.; Caamal-Chan, G.; Lucero-Vega, G.; Nieto-Garibay, A. Effects of vermicompost leachate versus inorganic fertilizer on morphology and microbial traits in the early development growth stage in mint (Mentha spicata L.) and rosemary (Rosmarinus officinalis L.) plants under closed hydroponic system. Horticulturae 2021, 7, 100. [Google Scholar] [CrossRef]
- Bidabadi, S.S.; Dehghanipoodeh, S.; Wright, G.C. Vermicompost leachate reduces some negative effects of salt stress in pomegranate. Int. J. Recycl. Org. Waste Agric. 2017, 6, 255–263. [Google Scholar] [CrossRef]
- Suthar, S. Evidence of plant hormone like substances in vermiwash: An ecologically safe option of synthetic chemicals for sustainable farming. Ecol. Eng. 2010, 36, 1089–1092. [Google Scholar] [CrossRef]
- Kaur, A.; Pati, P.K.; Ohri, P.; Kaur, A. Effects of Vermicompost and Vermicompost Leachate on the Biochemical and Physiological Response of Withania somnifera (L.) Dunal. J. Soil Sci. Plant Nutr. 2022, 22, 3228–3242. [Google Scholar] [CrossRef]
- Aslam, Z.; Ahmad, A.; Bellitürk, K.; Iqbal, N.; Idrees, M.; Rehman, W.U.; Akbar, G.; Tariq, M.; Raza, M.; Riasat, S. 26. Effects of vermicompost, vermi-tea and chemical fertilizer on morpho-physiological characteristics of tomato (Solanum lycopersicum) in Suleymanpasa District, Tekirdag of Turkey. Pure Appl. Biol. 2020, 9, 1920–1931. [Google Scholar] [CrossRef]
- Gong, X.; Li, S.; Sun, X.; Wang, L.; Cai, L.; Zhang, J.; Wei, L. Green waste compost and vermicompost as peat substitutes in growing media for geranium (Pelargonium zonale L.) and calendula (Calendula officinalis L.). Sci. Hortic. 2018, 236, 186–191. [Google Scholar] [CrossRef]
- Yadav, A.; Garg, V. Influence of vermi-fortification on chickpea (Cicer arietinum L.) growth and photosynthetic pigments. Int. J. Recycl. Org. Waste Agric. 2015, 4, 299–305. [Google Scholar] [CrossRef]
- Rekha, G.S.; Kaleena, P.K.; Elumalai, D.; Srikumaran, M.P.; Maheswari, V.N. Effects of vermicompost and plant growth enhancers on the exo-morphological features of Capsicum annum (Linn.) Hepper. Int. J. Recycl. Org. Waste Agric. 2018, 7, 83–88. [Google Scholar] [CrossRef]
- Arancon, N.Q.; Owens, J.D.; Converse, C. The effects of vermicompost tea on the growth and yield of lettuce and tomato in a non-circulating hydroponics system. J. Plant Nutr. 2019, 42, 2447–2458. [Google Scholar] [CrossRef]
- Purwanto, A. Development of technology vermicompost production for the coffee plant Industry. J. Phys. Conf. Ser. 2021, 1876, 012020. [Google Scholar]
- Sundararasu, K.; Jeyasankar, A. Effect of vermiwash on growth and yield of brinjal, Solanum melongena (eggplant or aubergine). Asian J. Sci. Technol. 2014, 5, 171–173. [Google Scholar]
- Ravindran, B.; Lee, S.R.; Chang, S.W.; Nguyen, D.D.; Chung, W.J.; Balasubramanian, B.; Mupambwa, H.A.; Arasu, M.V.; Al-Dhabi, N.A.; Sekaran, G. Positive effects of compost and vermicompost produced from tannery waste-animal fleshing on the growth and yield of commercial crop-tomato (Lycopersicon esculentum L.) plant. J. Environ. Manag. 2019, 234, 154–158. [Google Scholar] [CrossRef]
- Mahmud, M.; Abdullah, R.; Yaacob, J.S. Effect of vermicompost on growth, plant nutrient uptake and bioactivity of ex vitro pineapple (Ananas comosus var. MD2). Agronomy 2020, 10, 1333. [Google Scholar] [CrossRef]
- Ose, A.; Andersone-Ozola, U.; Ievinsh, G. Substrate-Dependent Effect of Vermicompost on Yield and Physiological Indices of Container-Grown Dracocephalum moldavica Plants. Agriculture 2021, 11, 1231. [Google Scholar] [CrossRef]
- Aslam, Z.; Aljuaid, B.S.; Abbas, R.N.; Bashir, S.; Almas, M.H.; Awan, T.H.; Belliturk, K.; Al-Taisan, W.a.A.; Mahmoud, S.F.; Bashir, S. Reduction in the Allelopathic Potential of Conocarpus erectus L. through Vermicomposting. Sustainability 2022, 14, 12840. [Google Scholar]
- Bziouech, S.A.; Dhen, N.; Helaoui, S.; Ammar, I.B.; Dridi, B.A.M. Effect of vermicompost soil additive on growth performance, physiological and biochemical responses of tomato plants (Solanum lycopersicum L. var. Firenze) to salt stress. Emir. J. Food Agric. 2022, 34, 316–328. [Google Scholar]
- Olivares, F.L.; Busato, J.G.; de Paula, A.M.; da Silva Lima, L.; Aguiar, N.O.; Canellas, L.P. Plant growth promoting bacteria and humic substances: Crop promotion and mechanisms of action. Chem. Biol 2017, 4, 30. [Google Scholar] [CrossRef]
- Canellas, L.P.; Zandonadi, D.B.; Busato, J.G.; Baldotto, M.A.; Simões, M.L.; Martin-Neto, L.; Façanha, A.R.; Spaccini, R.; Piccolo, A. Bioactivity and chemical characteristics of humic acids from tropical soils sequence. Soil Sci. 2008, 173, 624–637. [Google Scholar] [CrossRef]
- Nardi, S.; Ertani, A.; Francioso, O. Soil–root cross-talking: The role of humic substances. J. Plant Nutr. Soil Sci. 2017, 180, 5–13. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, R.; Kumar, S.; Gupta, R.; Patil, R. Vermicompost substitution influences growth, physiological disorders, fruit yield and quality of strawberry (Fragaria x ananassa Duch.). Bioresour. Technol. 2008, 99, 8507–8511. [Google Scholar] [CrossRef]
- Garcia-Mina, J.; Antolin, M.; Sanchez-Diaz, M. Metal-humic complexes and plant micronutrient uptake: A study based on different plant species cultivated in diverse soil types. Plant Soil 2004, 258, 57–68. [Google Scholar] [CrossRef]
- Zanin, L.; Tomasi, N.; Cesco, S.; Varanini, Z.; Pinton, R. Humic substances contribute to plant iron nutrition acting as chelators and biostimulants. Front. Plant Sci. 2019, 10, 675. [Google Scholar] [CrossRef]
- Aguiar, N.O.; Olivares, F.L.; Novotny, E.H.; Dobbss, L.B.; Balmori, D.M.; Santos-Júnior, L.G.; Chagas, J.G.; Façanha, A.R.; Canellas, L.P. Bioactivity of humic acids isolated from vermicomposts at different maturation stages. Plant Soil 2013, 362, 161–174. [Google Scholar] [CrossRef]
- Dobbss, L.B.; Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Peres, L.E.P.; Azevedo, M.; Spaccini, R.; Piccolo, A.; Façanha, A.R. Bioactivity of chemically transformed humic matter from vermicompost on plant root growth. J. Agric. Food Chem. 2010, 58, 3681–3688. [Google Scholar] [CrossRef] [PubMed]
- Ievinsh, G.; Vikmane, M.; Ķirse, A.; Karlsons, A. Effect of vermicompost extract and vermicompost-derived humic acids on seed germination and seedling growth of hemp. Proc. Latv. Acad. Sci. 2017, 71, 286–292. [Google Scholar] [CrossRef]
- Gomes, G.A.; Pereira, R.A.; Sodré, G.A.; Gross, E. Humic acids from vermicompost positively influence the nutrient uptake in mangosteen seedlings. Pesqui. Agropecu. Trop. 2019, 49. [Google Scholar]
- El-Hameid, A.; Adel, S. Improving productivity and quality of mango using humic acid and vermicomposting leachate in North Sinai. Egypt. J. Desert Res. 2018, 68, 37–59. [Google Scholar] [CrossRef]
- Rosa, M.; Olimpo, A.; Fastuca, R.; Caprioglio, A. Perceptions of dental professionals and laypeople to altered dental esthetics in cases with congenitally missing maxillary lateral incisors. Prog. Orthod. 2013, 14, 34. [Google Scholar] [CrossRef] [PubMed]
- Balmori, D.; Domínguez, C.; Carreras, C.; Rebatos, S.; Farías, L.; Izquierdo, F.; Berbara, R.; García, A.C. Foliar application of humic liquid extract from vermicompost improves garlic (Allium sativum L.) production and fruit quality. Int. J. Recycl. Org. Waste Agric. 2019, 8, 103–112. [Google Scholar] [CrossRef]
- Baldotto, L.E.B.; Baldotto, M.A.; Giro, V.B.; Canellas, L.P.; Olivares, F.L.; Bressan-Smith, R. Desempenho do abacaxizeiro’Vitória’em resposta à aplicação de ácidos húmicos durante a aclimatação. Rev. Bras. Ciência Solo 2009, 33, 979–990. [Google Scholar] [CrossRef]
- Nikbakht, A.; Kafi, M.; Babalar, M.; Xia, Y.P.; Luo, A.; Etemadi, N.-a. Effect of humic acid on plant growth, nutrient uptake, and postharvest life of gerbera. J. Plant Nutr. 2008, 31, 2155–2167. [Google Scholar] [CrossRef]
- Hernández, A.; Castillo, H.; Ojeda, D.; Arras, A.; López, J.; Sánchez, E. Effect of vermicompost and compost on lettuce production. Chil. J. Agric. Res. 2010, 70, 583–589. [Google Scholar] [CrossRef]
- Subler, S.; Edwards, C.; Metzger, J. Comparing vermicomposts and composts. Biocycle 1998, 39, 63–66. [Google Scholar]
- Sinha, R.K.; Hahn, G.; Singh, P.K.; Suhane, R.K.; Allam, A. Organic farming by vermiculture: Producing safe, nutritive and protective foods by earthworms (Charles Darwin’s friends of farmers). Am. J. Exp. Agric. 2011, 1, 363–399. [Google Scholar] [CrossRef]
- Zhang, H.; Tan, S.; Wong, W.; Ng, C.; Teo, C.; Ge, L.; Chen, X.; Yong, J. Mass spectrometric evidence for the occurrence of plant growth promoting cytokinins in vermicompost tea. Biol. Fertil. Soils 2014, 50, 401–403. [Google Scholar] [CrossRef]
- Aremu, A.O.; Stirk, W.A.; Kulkarni, M.G.; Tarkowská, D.; Turečková, V.; Gruz, J.; Šubrtová, M.; Pěnčík, A.; Novák, O.; Doležal, K. Evidence of phytohormones and phenolic acids variability in garden-waste-derived vermicompost leachate, a well-known plant growth stimulant. J. Plant Growth Regul. 2015, 75, 483–492. [Google Scholar] [CrossRef]
- Li, Z.; He, Y. Roles of brassinosteroids in plant reproduction. Int. J. Mol. Sci. 2020, 21, 872. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.; Tan, S.; Ge, L.; Chen, X.; Letham, D.; Yong, J. The importance of phytohormones and microbes in biostimulants: Mass spectrometric evidence and their positive effects on plant growth. In Proceedings of the II World Congress on the Use of Biostimulants in Agriculture 1148, Florence, Italy, 16–19 November 2015; pp. 49–60. [Google Scholar]
- Steffen, G.P.K.; Maldaner, J.; de Morais, R.M.; Saldanha, C.W.; Missio, E.L.; Steffen, R.B.; Osorio Filho, B.D. The vermicompost anticipates flowering and increases tomato productivity. Agrocienc. Urug. 2019, 23, e54. [Google Scholar] [CrossRef]
- Sardoei, A. Vermicompost effects on the growth and flowering of marigold (Calendula officinalis). Eur. J. Exp. Biol. 2014, 4, 651–655. [Google Scholar]
- Arora, V.; Singh, C.; Sidhu, A.; Thind, S. Irrigation, tillage and mulching effects on soybean yield and water productivity in relation to soil texture. Agric. Water Manag. 2011, 98, 563–568. [Google Scholar] [CrossRef]
- Lazcano, C.; Domínguez, J. The use of vermicompost in sustainable agriculture: Impact on plant growth and soil fertility. Soil Nutr. 2011, 10, 187. [Google Scholar]
- Ansari, A.; Ismail, S. Role of Earthworms in vermitechnology. J. Agric. Technol. 2012, 8, 405–415. [Google Scholar]
- Song, X.; Liu, M.; Wu, D.; Griffiths, B.S.; Jiao, J.; Li, H.; Hu, F. Interaction matters: Synergy between vermicompost and PGPR agents improves soil quality, crop quality and crop yield in the field. Appl. Soil Ecol. 2015, 89, 25–34. [Google Scholar] [CrossRef]
- Karnwal, A. Pseudomonas spp., a zinc-solubilizing vermicompost bacteria with plant growth-promoting activity moderates zinc biofortification in tomato. Int. J. Veg. Sci. 2021, 27, 398–412. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Bellitürk, K.; Hussain, S.; Bibi, I. Soil Application of Cellulolytic Microbe–Enriched Vermicompost Modulated the Morpho-physiological and Biochemical Responses of Wheat Cultivars under Different Moisture Regimes. J. Soil Sci. Plant Nutr 2022, 22, 4153–4167. [Google Scholar] [CrossRef]
- Shrimal, P.; Khan, T. Studies on the effects of vermicompost on growth parameters and chlorophyll content of bengal gram (Cicer arietinum L.) var. RSG-896. IOSR J. Environ. Sci. Toxicol. Food Technol. 2017, 11, 12–16. [Google Scholar] [CrossRef]
- Fikru, T.K.; Fikreyohannes, G. Response of garlic (Allium sativum L.) to vermicompost and mineral N fertilizer application at Haramaya, Eastern Ethiopia. Afr. J. Food Agric. Res. 2018, 13, 27–35. [Google Scholar] [CrossRef]
- Baliah, T.; Muthulakshmi, P. Effect of microbially enriched vermicompost on the growth and biochemical characteristics of Okra (Abelmoschus esculentus (L.) Moench). Adv. Plants Agric. Res 2017, 6, 00228. [Google Scholar]
- Parastesh, F.; Alikhani, H.A.; Etesami, H. Vermicompost enriched with phosphate–solubilizing bacteria provides plant with enough phosphorus in a sequential cropping under calcareous soil conditions. J. Clean. Prod. 2019, 221, 27–37. [Google Scholar] [CrossRef]
- Verma, S.; Nizam, S.; Verma, P.K. Biotic and abiotic stress signaling in plants. In Stress Signaling in Plants: Genomics and Proteomics Perspective; Springer: New York, NY, USA, 2013; Volume 1, pp. 25–49. [Google Scholar]
- Sharif, F.; Danish, M.U.; Ali, A.S.; Khan, A.U.; Shahzad, L.; Ali, H.; Ghafoor, A. Salinity tolerance of earthworms and effects of salinity and vermi amendments on growth of Sorghum bicolor. Arch. Agron. Soil Sci. 2016, 62, 1169–1181. [Google Scholar]
- Dikobe, T.B.; Mashile, B.; Sinthumule, R.R.; Ruzvidzo, O. Distinct Morpho-Physiological Responses of Maize to Salinity Stress. Am. J. Plant Sci. 2021, 12, 946–959. [Google Scholar] [CrossRef]
- Akoto-Danso, E.K.; Manka’abusi, D.; Steiner, C.; Werner, S.; Häring, V.; Nyarko, G.; Marschner, B.; Drechsel, P.; Buerkert, A. Agronomic effects of biochar and wastewater irrigation in urban crop production of Tamale, northern Ghana. Nutr. Cycl. Agroecosyst. 2019, 115, 231–247. [Google Scholar] [CrossRef]
- Alsaeedi, A.; El-Ramady, H.; Alshaal, T.; El-Garawani, M.; Elhawat, N.; Al-Otaibi, A. Exogenous nanosilica improves germination and growth of cucumber by maintaining K+/Na+ ratio under elevated Na+ stress. Plant Physiol. Biochem. 2018, 125, 164–171. [Google Scholar] [CrossRef]
- Zhang, Q.; Gu, X.; Singh, V.P.; Kong, D.; Chen, X. Spatiotemporal behavior of floods and droughts and their impacts on agriculture in China. Global Planet. Chang. 2015, 131, 63–72. [Google Scholar] [CrossRef]
- Abbott, L.; Macdonald, L.; Wong, M.; Webb, M.; Jenkins, S.; Farrell, M. Potential roles of biological amendments for profitable grain production–A review. Agric. Ecosyst. Environ. 2018, 256, 34–50. [Google Scholar] [CrossRef]
- García, A.C.; Santos, L.A.; Izquierdo, F.G.; Sperandio, M.V.L.; Castro, R.N.; Berbara, R.L.L. Vermicompost humic acids as an ecological pathway to protect rice plant against oxidative stress. Ecol. Engin. 2012, 47, 203–208. [Google Scholar] [CrossRef]
- Shilev, S. Plant-growth-promoting bacteria mitigating soil salinity stress in plants. Appl. Sci. 2020, 10, 7326. [Google Scholar] [CrossRef]
- Cicek, N.; Erdogan, M.; Yucedag, C.; Cetin, M. Improving the Detrimental Aspects of Salinity in Salinized Soils of Arid and Semi-arid Areas for Effects of Vermicompost Leachate on Salt Stress in Seedlings. Water Air Soil Pollut. 2022, 233, 197. [Google Scholar] [CrossRef]
- Hafez, E.M.; Omara, A.E.D.; Alhumaydhi, F.A.; El-Esawi, M.A. Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Physiol. Plant. 2021, 172, 587–602. [Google Scholar] [CrossRef] [PubMed]
- Sorkhi, F. Effect of vermicompost fertilizer on antioxidant enzymes and chlorophyll contents in Borago officinalis under salinity stress. Iran. J. Plant Physiol. 2021, 11, 3589–3598. [Google Scholar]
- Reyes-Perez, J.J.; Amador, B.M.; Hernández-Montiel, L.G.; Rangel, P.P.; Rueda-Puente, E.O.; Ruiz-Espinoza, F.H. Vermicompost humates as a salinity mitigator in the germination of basil. Cienc. Rural. 2021, 52. [Google Scholar] [CrossRef]
- Benazzouk, S.; Dobrev, P.I.; Djazouli, Z.-E.; Motyka, V.; Lutts, S. Positive impact of vermicompost leachate on salt stress resistance in tomato (Solanum lycopersicum L.) at the seedling stage: A phytohormonal approach. Plant Soil. 2020, 446, 145–162. [Google Scholar] [CrossRef]
- Oo, A.; Iwai, C.; Saenjan, P. Soil properties and maize growth in saline and nonsaline soils using cassava-industrial waste compost and vermicompost with or without earthworms. Land Degrad. Dev. 2015, 26, 300–310. [Google Scholar] [CrossRef]
- Demir, Z. Alleviation of adverse effects of sodium on soil physicochemical properties by application of vermicompost. Compos. Sci. Util. 2020, 28, 100–116. [Google Scholar] [CrossRef]
- Beyk-Khormizi, A.; Sarghein, S.H.; Sarafraz-Ardakani, M.R.; Moshtaghioun, S.M.; Mousavi-Kouhi, S.M.; Ganjeali, A. Ameliorating effect of vermicompost on Foeniculum vulgare under saline condition. J. Plant Nutr. 2022, 46, 1601–1615. [Google Scholar] [CrossRef]
- Alamer, K.H.; Perveen, S.; Khaliq, A.; Zia Ul Haq, M.; Ibrahim, M.U.; Ijaz, B. Mitigation of salinity stress in maize seedlings by the application of vermicompost and sorghum water extracts. Plants 2022, 11, 2548. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Yu, Z.; Xu, L.; Zhao, Y.; Yi, S.; Shen, C.; Wang, Y.; Li, Y.; Zuo, W.; Gu, C. Effects of vermicompost application on growth and heavy metal uptake of barley grown in mudflat salt-affected soils. Agronomy 2022, 12, 1007. [Google Scholar] [CrossRef]
- Ibrahim, H.A.; Heba, S.; Hassan, N.M. Impact on growth, yield and nutritional status of tomato plants grown in saline soil by vermicompost and ascorbic acid. Int. J. Health Sci. 2022, 6, 10134–10143. [Google Scholar] [CrossRef]
- Xiao, X.; Wu, Z.-C.; Chou, K.-C. A multi-label classifier for predicting the subcellular localization of gram-negative bacterial proteins with both single and multiple sites. PLoS ONE 2011, 6, e20592. [Google Scholar] [CrossRef]
- Kaur, H.; Manna, M.; Thakur, T.; Gautam, V.; Salvi, P. Imperative role of sugar signaling and transport during drought stress responses in plants. Physiol. Plant. 2021, 171, 833–848. [Google Scholar] [CrossRef]
- Istanbuli, T.; Baum, M.; Touchan, H.; Hamwieh, A. Evaluation of morpho-physiological traits under drought stress conditions in barley (Hordeum vulgare L.). Photosynthetica 2020, 58, 1059–1067. [Google Scholar] [CrossRef]
- Nofouzi, F. Evaluation of seed yield of durum wheat (Triticum durum) under drought stress and determining correlation among some yield components using path coefficient analysis. Cuad. Investig. UNED 2018, 10, 179–183. [Google Scholar] [CrossRef]
- Kalagare, V.S.; Ganesan, N.M.; Iyanar, K.; Chitdeshwari, T.; Chandrasekhar, C. Strategy of multiple selection indices for discrimination of potential genotypes and associated traits for yield improvement in pearl millet [Pennisetum glaucum (L.) R. Br.]. Electron. J. Plant Breed. 2021, 12, 895–906. [Google Scholar]
- Roy, S.; Arunachalam, K.; Dutta, B.K.; Arunachalam, A. Effect of organic amendments of soil on growth and productivity of three common crops viz. Zea mays, Phaseolus vulgaris and Abelmoschus esculentus. Appl. Soil Ecol. 2010, 45, 78–84. [Google Scholar] [CrossRef]
- Chen, J.; Xu, W.; Velten, J.; Xin, Z.; Stout, J. Characterization of maize inbred lines for drought and heat tolerance. J. Soil Water Conserv. 2012, 67, 354–364. [Google Scholar] [CrossRef]
- Hosseinzadeh, S.; Amiri, H.; Ismaili, A. Effect of vermicompost fertilizer on photosynthetic characteristics of chickpea (Cicer arietinum L.) under drought stress. Photosynthetica 2016, 54, 87–92. [Google Scholar] [CrossRef]
- Dehmordy, M.; Gholami, M.; Baninasab, B. Effect of vermicompost fertilizer on growth and drought tolerance of Olive (Olea europaea L. cv. Zard). J. Plant Physiol. 2018, 7, 1–18. [Google Scholar]
- Hosseinzadeh, S.R.; Amiri, H.; Ismaili, A. Evaluation of photosynthesis, physiological, and biochemical responses of chickpea (Cicer arietinum L. cv. Pirouz) under water deficit stress and use of vermicompost fertilizer. J. Integr. Agric. 2018, 17, 2426–2437. [Google Scholar] [CrossRef]
- Hosseinzadeh, S.R.; Amiri, H.; Ismaili, A. Nutrition and biochemical responses of chickpea (Cicer arietinum L.) to vermicompost fertilizer and water deficit stress. J. Plant Nutr. 2017, 40, 2259–2268. [Google Scholar] [CrossRef]
- Özenç, D.B. Growth and transpiration of tomato seedlings grown in Hazelnut Husk compost under water-deficit stress. Compos. Sci. Util. 2008, 16, 125–131. [Google Scholar] [CrossRef]
- Atik, A. Effects of planting density and treatment with vermicompost on the morphological characteristics of oriental beech (Fagus orientalis Lipsky.). Compos. Sci. Util. 2013, 21, 87–98. [Google Scholar] [CrossRef]
- Benaffari, W.; Boutasknit, A.; Anli, M.; Ait-El-Mokhtar, M.; Ait-Rahou, Y.; Ben-Laouane, R.; Ben Ahmed, H.; Mitsui, T.; Baslam, M.; Meddich, A. The native arbuscular mycorrhizal fungi and vermicompost-based organic amendments enhance soil fertility, growth performance, and the drought stress tolerance of quinoa. Plants 2022, 11, 393. [Google Scholar] [CrossRef]
- Feizabadi, A.; Noormohammadi, G.; Fatehi, F. Changes in growth, physiology, and fatty acid profile of rapeseed cultivars treated with vermicompost under drought stress. J. Soil Sci. Plant Nutr. 2021, 21, 200–208. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Hussain, S.; Javed, T.; Hussain, S.; Bashir, S.; Hussain, I.; Belliturk, K.; Adamski, R.; Siuta, D. Soil application of wheat straw vermicompost enhances morpho-physiological attributes and antioxidant defense in wheat under drought stress. Front. Environ. Sci. 2022, 10, 387. [Google Scholar] [CrossRef]
- Kiran, S. Effects of vermicompost on some morphological, physiological and biochemical parameters of lettuce (Lactuca sativa var. crispa) under drought stress. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 352–358. [Google Scholar] [CrossRef]
- Paymaneh, Z.; Sarcheshmehpour, M.; Mohammadi, H.; Hesni, M.A. Vermicompost and/or compost and arbuscular mycorrhizal fungi are conducive to improving the growth of pistachio seedlings to drought stress. App. Soil Ecol. 2023, 182, 104717. [Google Scholar] [CrossRef]
- Alhverdizadeh, S.; Danaee, E. Effect of Humic Acid and Vermicompost on Some Vegetative Indices and Proline Content of Catharanthus roseous under Low Water Stress. Environ. Eng. Res. 2023, 9, 141–152. [Google Scholar]
- Khosravi Shakib, A.; Rezaei Nejad, A.; Khandan Mirkohi, A.; Kalate Jari, S. Vermicompost and manure compost reduce water-deficit stress in pot marigold (Calendula officinalis L. cv. Candyman Orange). Compos. Sci. Util. 2019, 27, 61–68. [Google Scholar] [CrossRef]
- Collange, B.; Navarrete, M.; Peyre, G.; Mateille, T.; Tchamitchian, M. Root-knot nematode (Meloidogyne) management in vegetable crop production: The challenge of an agronomic system analysis. Crop Prot. 2011, 30, 1251–1262. [Google Scholar] [CrossRef]
- Doan, T.T.; Henry-des-Tureaux, T.; Rumpel, C.; Janeau, J.-L.; Jouquet, P. Impact of compost, vermicompost and biochar on soil fertility, maize yield and soil erosion in Northern Vietnam: A three year mesocosm experiment. Sci. Total Environ. 2015, 514, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Cardoza, Y.J.; Buhler, W.G. Soil organic amendment impacts on corn resistance to Helicoverpa zea: Constitutive or induced? Pedobiologia 2012, 55, 343–347. [Google Scholar] [CrossRef]
- Rostami, M.; Olia, M.; Arabi, M. Evaluation of the effects of earthworm Eisenia fetida-based products on the pathogenicity of root-knot nematode (Meloidogyne javanica) infecting cucumber. Int. J. Recycl. Org. Waste Agric. 2014, 3, 58. [Google Scholar] [CrossRef]
- Koul, O. Phytochemicals and insect control: An antifeedant approach. Crit. Rev. Plant Sci. 2008, 27, 1–24. [Google Scholar] [CrossRef]
- Mohamadi, P.; Razmjou, J.; Naseri, B.; Hassanpour, M. Humic fertilizer and vermicompost applied to the soil can positively affect population growth parameters of Trichogramma brassicae (Hymenoptera: Trichogrammatidae) on eggs of Tuta absoluta (Lepidoptera: Gelechiidae). Neotrop. Entomol. 2017, 46, 678–684. [Google Scholar] [CrossRef]
- Karthikeyan, M.; Hussain, N.; Gajalakshmi, S.; Abbasi, S. Effect of vermicast generated from an allelopathic weed lantana (Lantana camara) on seed germination, plant growth, and yield of cluster bean (Cyamopsis tetragonoloba). Environ. Sci. Pol. Res. 2014, 21, 12539–12548. [Google Scholar] [CrossRef]
- You, X.; Tojo, M.; Ching, S.; Wang, K.-H. Effects of vermicompost water extract prepared from bamboo and kudzu against and. J. Nematol. 2018, 50, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Renčo, M.; Kováčik, P. Assessment of the nematicidal potential of vermicompost, vermicompost tea, and urea application on the potato-cyst nematodes Globodera rostochiensis and Globodera pallida. J. Plant Prot. Res. 2015, 55, 187–192. [Google Scholar] [CrossRef]
- Sivasabari, K.; Parthasarathy, S.; Chandran, D.; Sankaralingam, S.; Ajaykumar, R. Vermicompost and Vermi-leachate in Pest and Disease Management. In Vermicomposting for Sustainable Food Systems in Africa; Springer: Singapore, 2023; pp. 279–304. [Google Scholar]
- Huang, K.; Li, F.; Wei, Y.; Fu, X.; Chen, X. Effects of earthworms on physicochemical properties and microbial profiles during vermicomposting of fresh fruit and vegetable wastes. Bioresour. Technol. 2014, 170, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar] [CrossRef]
- Molinari, S.; Leonetti, P. Bio-control agents activate plant immune response and prime susceptible tomato against root-knot nematodes. PLoS ONE 2019, 14, e0213230. [Google Scholar] [CrossRef]
- Singh, A.; Jain, A.; Sarma, B.K.; Abhilash, P.; Singh, H.B. Solid waste management of temple floral offerings by vermicomposting using Eisenia fetida. Waste Manag. 2013, 33, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Liu, M.; Jiang, L.; Chen, X.; Griffiths, B.S.; Li, H.; Hu, F. Vermicompost increases defense against root-knot nematode (Meloidogyne incognita) in tomato plants. Appl. Soil Ecol. 2016, 105, 177–186. [Google Scholar] [CrossRef]
- Machfudz, M.; Basit, A.; Handoko, R.N.S. Effectiveness of vermicompost with additives of various botanical pesticides in controlling Plutella xylostella and their effects on the yield of cabbage (Brassica oleracea L. var. Capitata). Asian J. Agric. Biol. 2020, 8, 223–232. [Google Scholar]
- Kaur, A.; Kaur, A.; Ohri, P. Combined effects of vermicompost and vermicompost leachate on the early growth of Meloidogyne incognita stressed Withania somnifera (L.) Dunal. Environ.Sci. Pol. Res. 2022, 29, 51686–51702. [Google Scholar] [CrossRef]
- Zuhair, R.; Moustafa, Y.T.A.; Mustafa, N.S.; El-Dahshouri, M.F.; Zhang, L.; Ageba, M.F. Efficacy of amended vermicompost for bio-control of root knot nematode (RKN) Meloidogyne incognita infesting tomato in Egypt. Environ. Technol. Innov. 2022, 27, 102397. [Google Scholar] [CrossRef]
- Edwards, C.A.; Arancon, N.Q.; Vasko-Bennett, M.; Askar, A.; Keeney, G.; Little, B. Suppression of green peach aphid (Myzus persicae) (Sulz.), citrus mealybug (Planococcus citri)(Risso), and two spotted spider mite (Tetranychus urticae) (Koch.) attacks on tomatoes and cucumbers by aqueous extracts from vermicomposts. Crop Prot. 2010, 29, 80–93. [Google Scholar] [CrossRef]
- Hussain, N.; Abbasi, T.; Abbasi, S. Evaluating the fertilizer and pesticidal value of vermicompost generated from a toxic and allelopathic weed ipomoea. J. Saudi Soc. Agric. 2020, 19, 43–50. [Google Scholar] [CrossRef]
- dos Santos Pereira, T.; Macêdo, A.G.; da Silva, J.; Pinheiro, J.B.; de Paula, A.M.; Biscaia, D.; Busato, J.G. Water-extractable fraction of vermicomposts enriched with Trichoderma enhances the growth of bell pepper and tomato as well as their tolerance against Meloidogyne incognita. Sci. Horti. 2020, 272, 109536. [Google Scholar] [CrossRef]
- Razmjou, J.; Mohammadi, M.; Hassanpour, M. Effect of vermicompost and cucumber cultivar on population growth attributes of the melon aphid (Hemiptera: Aphididae). J. Econ. Entomol. 2011, 104, 1379–1383. [Google Scholar] [CrossRef]
- Ullah, M.I.; Riaz, M.; Arshad, M.; Khan, A.H.; Afzal, M.; Khalid, S.; Mehmood, N.; Ali, S.; Khan, A.M.; Zahid, S.M.A. Application of Organic Fertilizers Affect the Citrus Leafminer, Phyllocnistis citrella (Lepidoptera: Gracillariidae) Infestation and Citrus Canker Disease in Nursery Plantations. Int. J. Insect Sci. 2019, 11, 1179543319858634. [Google Scholar] [CrossRef] [PubMed]
- Arancon, N.Q.; Galvis, P.A.; Edwards, C.A. Suppression of insect pest populations and damage to plants by vermicomposts. Bioresour. Technol. 2005, 96, 1137–1142. [Google Scholar] [CrossRef]
- Gudeta, K.; Julka, J.; Kumar, A.; Bhagat, A.; Kumari, A. Vermiwash: An agent of disease and pest control in soil, a review. Heliyon 2021, 7, e06434. [Google Scholar] [CrossRef]
- Nadana, G.R.V.; Rajesh, C.; Kavitha, A.; Sivakumar, P.; Sridevi, G.; Palanichelvam, K. Induction of growth and defense mechanism in rice plants towards fungal pathogen by eco-friendly coelomic fluid of earthworm. Environ. Technol. Innov. 2020, 19, 101011. [Google Scholar] [CrossRef]
- Hakim Rabet, S.; Ketabchi, S. The effect of compost fertilizers, vermicompost and their tea on bacterial vascular wilt and growth indices in tomato seedlings. J. Appl. Res. Plant Prot. 2021, 9, 61–74. [Google Scholar]
- Kinay, P.; Mansour, M.F.; Gabler, F.M.; Margosan, D.A.; Smilanick, J.L. Characterization of fungicide-resistant isolates of Penicillium digitatum collected in California. Crop Prot. 2007, 26, 647–656. [Google Scholar] [CrossRef]
- You, X.; Wakana, D.; Ishikawa, K.; Hosoe, T.; Tojo, M. Antifungal activity of compounds isolated from bamboo vermicompost against Rhizoctonia solani AG1-IB. Adv. Microbiol. 2019, 9, 957. [Google Scholar] [CrossRef]
- Jackson, K.S.; Kumar, A.; Kumar, V. Evaluation of Organic Amendments Against Alternaria Brassicae Causing Alternaria Leaf Spot on Mustard under In Vitro and Poly House Conditions. Plant Cell Biotechnol. Mol. Biol. 2019, 20, 285–293. [Google Scholar]
- Mu, J.; Li, X.; Jiao, J.; Ji, G.; Wu, J.; Hu, F.; Li, H. Biocontrol potential of vermicompost through antifungal volatiles produced by indigenous bacteria. Biol. Control 2017, 112, 49–54. [Google Scholar] [CrossRef]
- Tian, G.-L.; Bi, Y.-M.; Jiao, X.-L.; Zhang, X.-M.; Li, J.-F.; Niu, F.-B.; Gao, W.-W. Application of vermicompost and biochar suppresses Fusarium root rot of replanted American ginseng. Appl. Microbiol. Biotechnol. 2021, 105, 6977–6991. [Google Scholar] [CrossRef] [PubMed]
- Kalantari, S.; Marefat, A.; Naseri, B.; Hemmati, R. Improvement of bean yield and Fusarium root rot biocontrol using mixtures of Bacillus, Pseudomonas and Rhizobium. Trop. Plant Pathol. 2018, 43, 499–505. [Google Scholar] [CrossRef]
- Baum, C.; Eichler-Löbermann, B.; Hrynkiewicz, K. Impact of organic amendments on the suppression of Fusarium wilt. In Organic Amendments and Soil Suppressiveness in Plant Disease Management; Springer: Cham, Switzerland, 2015; pp. 353–362. [Google Scholar]
- Srivastava, R.; Khalid, A.; Singh, U.; Sharma, A. Evaluation of arbuscular mycorrhizal fungus, fluorescent Pseudomonas and Trichoderma harzianum formulation against Fusarium oxysporum f. sp. lycopersici for the management of tomato wilt. Biol. Cont. 2010, 53, 24–31. [Google Scholar] [CrossRef]
- Litterick, A.; Wood, M. The use of composts and compost extracts in plant disease control. In Disease Control in Crops; Wiley-Blackwell: Hoboken, NJ, USA, 2009. [Google Scholar]
- Dowling, D.N.; O’Gara, F. Metabolites of Pseudomonas involved in the biocontrol of plant disease. Trends Biotechnol. 1994, 12, 133–141. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codon, A.C. Biocontrol mechanisms of Trichoderma strains. Intern. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Pharand, B.; Carisse, O.; Benhamou, N. Cytological aspects of compost-mediated induced resistance against Fusarium crown and root rot in tomato. Phytopathology 2002, 92, 424–438. [Google Scholar] [CrossRef]
- Sang, M.K.; Kim, K.D. Biocontrol activity and primed systemic resistance by compost water extracts against anthracnoses of pepper and cucumber. Phytopathology 2011, 101, 732–740. [Google Scholar] [CrossRef]
- M’piga, P.; Belanger, R.; Paulitz, T.; Benhamou, N. Increased resistance to Fusarium oxysporumf. sp. radicis-lycopersiciin tomato plants treated with the endophytic bacterium Pseudomonas fluorescens strain 63-28. Physiol. Mol. Plant Pathol. 1997, 50, 301–320. [Google Scholar] [CrossRef]
- De Vleesschauwer, D.; Cornelis, P.; Höfte, M. Redox-active pyocyanin secreted by Pseudomonas aeruginosa 7NSK2 triggers systemic resistance to Magnaporthe grisea but enhances Rhizoctonia solani susceptibility in rice. Mol. Plant-Microbe Interact. 2006, 19, 1406–1419. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.M.; Ludwig, A.; Furch, A.C.; Mithöfer, A.; Scholz, S.; Reichelt, M.; Oelmüller, R. The beneficial root-colonizing fungus Mortierella hyalina promotes the aerial growth of Arabidopsis and activates calcium-dependent responses that restrict Alternaria brassicae–induced disease development in roots. Mol. Plant-Microbe Interact. 2019, 32, 351–363. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Vermicompost serves as a plant growth regulator and biocontrol agent, enhancing the physical, biochemical, and fertility properties of soil while suppressing pest attacks.
Figure 1.
Vermicompost serves as a plant growth regulator and biocontrol agent, enhancing the physical, biochemical, and fertility properties of soil while suppressing pest attacks.

Figure 2.
A theoretical representation of the processes by which the fertility of EWs casts is increased through (i) the concentration of existing fertility pools in the soil and (ii) the transformation processes during gut transit, leading to new pools of fertility. Reprinted from ref. [54], with permission from Elsevier.
Figure 2.
A theoretical representation of the processes by which the fertility of EWs casts is increased through (i) the concentration of existing fertility pools in the soil and (ii) the transformation processes during gut transit, leading to new pools of fertility. Reprinted from ref. [54], with permission from Elsevier.

Figure 3.
The physiological roles of phytohormones in plant growth and development.

Figure 4.
Mechanisms of disease suppression in plants using vermicompost and its derivative (vermi-tea). Vermicompost boosts microbial diversity and activity in the rhizosphere. These microbes protect plants by inducing systemic resistance against phytopathogens, increasing nutrient competition, releasing specific metabolites against pathogens, and directly parasitizing phytopathogens through various mechanisms.
Figure 4.
Mechanisms of disease suppression in plants using vermicompost and its derivative (vermi-tea). Vermicompost boosts microbial diversity and activity in the rhizosphere. These microbes protect plants by inducing systemic resistance against phytopathogens, increasing nutrient competition, releasing specific metabolites against pathogens, and directly parasitizing phytopathogens through various mechanisms.


{kind=link}
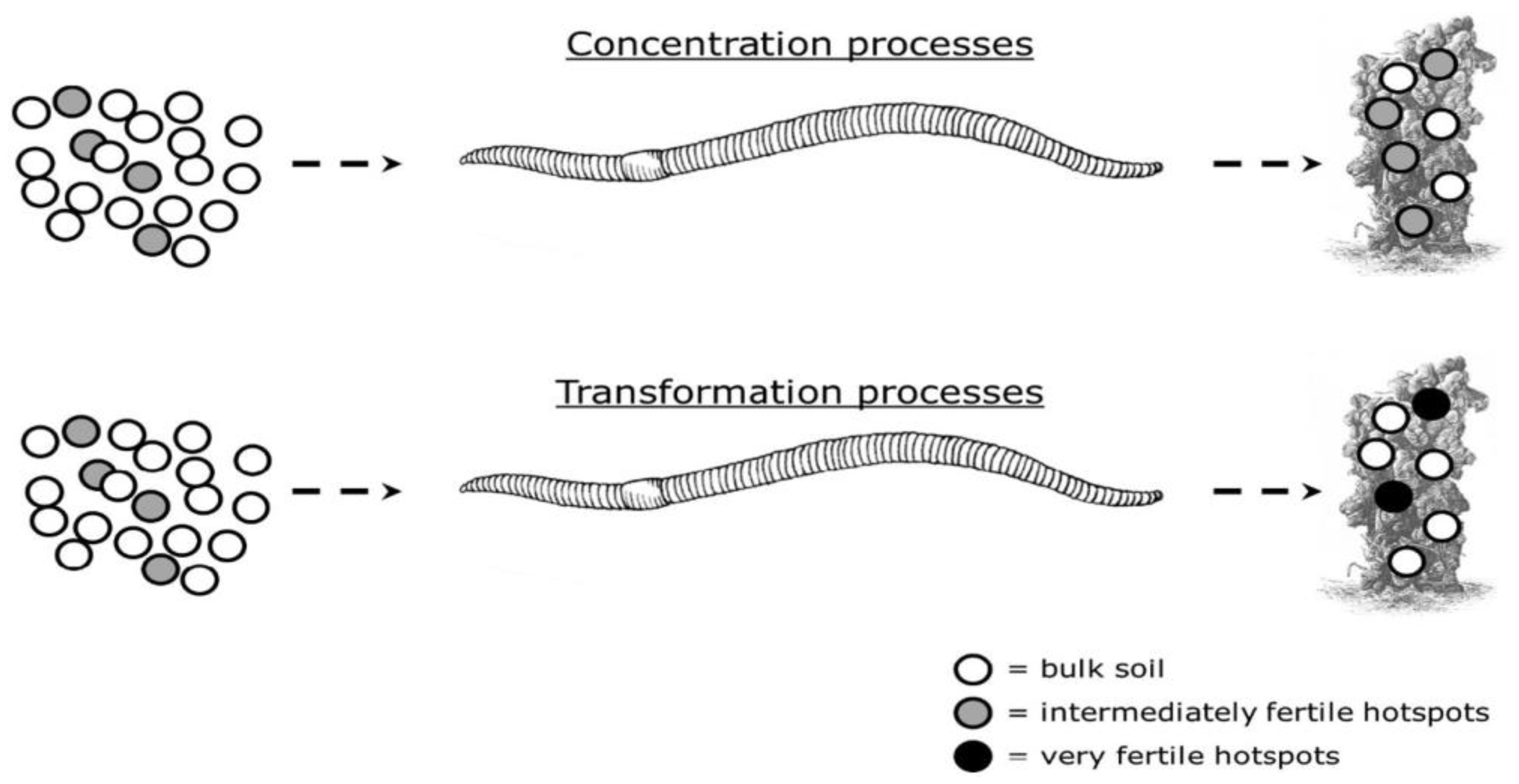{kind=link}
{kind=link}
{kind=link}
Table 1.
Impact of vermicompost and its derivatives on plant growth and yield.
| Type of Fertilizer | Source | Biochemical and Nutritional Characteristics | Test Plant | Effect on Plant Growth and Yield | References |
|---|---|---|---|---|---|
| VC | Green wastes | pH: 7.43, EC: 1.77 dsm−1, CEC:117.44 c mol kg−1, N: 21.47 g/kg, P: 4.82 g/kg, K: 7.87 g/kg, Ca: 179.15 g/kg, Mg: 12.58 g/kg, Zn: 189.12 mg/kg | Pelargonium zonale L. and Calendula officinalis L. | Growth and flowering of both plants were higher in all vermicompost-based media. | [78] |
| VC | Cow dung, food industry sludge | EC: 1.8 dS/m, pH: 6.6, C/N ratio:11.9, TKN: 26 g/kg, TAP: 9.75 g/kg, TK: 7.6 g/kg, | Cicer arietinum L. | Total chlorophyll contents and yield increased. | [79] |
| VC | Cattle dung, leaf litter, straw | - | Capsicum annum (L.) | Vermicompost treatment (50%) revealed excellent increase in chilly growth and soil quality improvement. | [80] |
| VT | Food wastes | EC: 6.36 mS/cm, pH: 6.5 and HA: 485.12 mg/L, N: 1.3, P: 204, K: 45.4, Ca: 449 and Mg: 40.1 mg/L | Lactuca sativa, Solanum lycopersicum | Increased the yield of both test plants even at reduced concentration (25%, 50%), respectively. | [81] |
| VC | skin coffee + green waste + biochar | pH: 5.60 N: 2.76%, K: 3.84%, | Coffea | Physiological and morphological parameters of coffee plant increased. | [82] |
| VW | Cow dung | - | Solanum melongena | Plant height and number of fruits increased significantly. | [83] |
| VC | Animal fleshing, Tannery waste | pH: 6.56, EC: 1.08 dS/m, TKN: 20 g/kg, Total P: 39 g/kg, Total K: 4.4 g/kg | Lycopersicom esculentum L. | Vermicompost resulted in higher plant height (10%), stem girth (8.9%), and leaf numbers (14%). | [84] |
| VC | - | Total N: 1.54%, Total P: 0.64%, Total K: 6.31%, total Mg: 0.58%, total Ca: 1.39%, total Zn: 0.01%, total S: 0.34% | Ananas comosus (L.) Merr. | Fruits contained higher total soluble solids, titratable acidity, ascorbic acid, and total chlorophyll content. | [85] |
| VC | Cow manure and grass biomass | pH: 8.03, N: 20 mg/L, P: 1997 mg/L, K: 8300 mg/L, Mg: 3600 mg/L, Ca: 7850 mg/L | Dracocephalum moldavica L. | Increased the plant’s biomass (148%) in peat-amended vermicompost and (68%) in soil amended with vermicompost. | [86] |
| VC | C. erectus leaves + FYM | pH: 6.69, nitrogen: 1.86%, P: 0.15%, K: 0.41%, Zn: 39.4 ppm, Ca: 4085.80 ppm, Fe: 1572 ppm | Vigna radiata L. | The economical and biological yield of mung beans were boosted up to 59% and 34%, respectively. | [87] |
| VC | - | EC: 2.15 mS/cm, pH: 8.74, OM: 74%, TOC: 43.02%, Porosity: 62.5% | Solanum lycopersicum | Plants cultivated on VC substrate indicated enhanced growth and better resistance to salinity stress. | [88] |
VC: Vermicompost; VW: Vermiwash; VT: Vermi-tea; EC: Electrical conductivity; CEC: Cation exchange capacity; TKN: Total Kjeldahl nitrogen; TAP: Total available phosphorus; N: Nitrogen; P: Phosphorus; K: Potassium; Ca: Calcium; Mg: Magnesium; Zn: Zinc; S: Sulfur; Fe: Iron; OM: Organic matter; TOC: Total organic carbon; HA: Humic acid.
Table 2.
Impact of vermicompost and its derivatives on soil properties and plant performance under saline conditions.
Table 2.
Impact of vermicompost and its derivatives on soil properties and plant performance under saline conditions.
| Amendment | Physiochemical and Nutritional Characteristics | Plant | Impact | References |
|---|---|---|---|---|
| VCL | pH: 8.32, total organic matter: 7.5%, total P: 1.2%, K: 1.5%, and N: 2.2%, and total humic + fulvic acid: 6% | Sedum album | VCL reduced effects of salinity and improved growth and quality traits of S. album. | [132] |
| VC + BC | VC had an OM content: 42%, pH: 7.4, EC: 3.8 dS/m, total N: 2.1%, total P: 7.8%, total K: 0.5%, BC had pH: 7.90, EC: 2.05 dS/m, total N: 1.9%, total P: 2.2%, total K: 2.9%, | Triticum aestivum L. | VC + BC reduced oxidative stress, proline and leaf Na+ content with increase in nutrient uptake and grain yield. Soil pH, EC, and ESP decreased. | [133] |
| VC | Organic carbon: 20.7%, EC: 5 dS/m, P: 0.7 mg/kg, K: 0.9 mg/kg, Na: 473 mg/kg | Borago officinalis | VC increased antioxidant enzymes and photosynthetic pigments activities and reduced the salinity. | [134] |
| VC | OM content: 74%, pH: 8.74, EC: 2.15 mS/cm, Salinity: 1.5 g/L, Porosity: 62.5% | Solanum lycopersicum L. | VC improved growth and better resistance to salinity stress. | [88] |
| VCH | C: 53.4%, pH: 8.7, N: 3.05%, S: 0.72%, humic acid: 4.82%, fulvic acid: 7.17% | Ocimum basilicum L. | VCH biostimulated and improved the growth and salinity tolerance of the sensitive variety. | [135] |
| VCL | - | Solanum lycopersicum L. | VCL lowers the impact of salinity on leaf senescence. | [136] |
| VW + VC | Vermicompost: pH: 7.2, EC: 2.69 mS/cm, N: 0.98%, P: 121 ppm, K: 46 ppm. Vermiwash: pH: 7.5, EC: 2.82 mS/cm, N: 0.45%, P: 41 ppm, K: 21 pmm | Sorghum bicolor L. | Vermi-amendments increased the nutritional status of soil, reduced the salt damage to plants, and improved growth. | [124] |
| VC | pH: 7.17, EC: 1.05 dS/m, TN: 4.13 g/kg, TP: 1.24 g/kg, TK: 1.70 g/kg, C:N = 15.04 | Zea mays L. | VC decreased exchangeable Na+ and improved soil microbial activities and maize growth. | [137] |
| VC | pH: 7.1, EC: 6.5 dS/m, OM: 65.5%, TN: 2.2%, TP: 7.3 mg/kg, TK: 12.8 mg/kg, Ca: 25.1 mg/kg, Zn: 216 mg/kg | VC decreased EC, bulk density, and exchangeable Na, and increased aggregate stability and total porosity, | [138] | |
| VC | - | Foeniculum vulgare | VC can limit the adverse effects of salinity stress on fennel by affecting photosynthetic pigments, membrane integrity and antioxidants activity. | [139] |
| VC + SWE | pH: 7.98, EC: 2.98 mS/cm, N: 0.07%, AP: 6.34 mg/kg, AK: 127 mg/kg, Zn: 0.97 mg/kg | Zea mays L. | VC + SWE decreased Na+ concentration, H2O2, and malondialdehyde contents at higher salinity. | [140] |
| VC | pH: 6.34, EC: 8.92 mS/cm, TN: 24.4 g/kg, TP: 15.9 g/kg, Alkaline N: 2467 mg/kg, AP: 869 mg/kg, Zn: 805 mg/kg | Hordeum vulgare L. | VC application decreased bulk density, EC, and pH of saline soil, whereas the yield of grain increased up to 512% as quantity of VC increased. | [141] |
| VC + AA | pH: 6.90, EC: 2.0 dS/m, OM: 50.3%, N: 1.20%, P: 0.50%, K: 0.80% | Lycopersicum esculentum L. cv Kasel rock | VC and ascorbic acid application decreased Na contents in leaves and increased tomato yield and quality. | [142] |
ESP: Exchangeable sodium percentage; SWE: Sorghum water extracts; VC: Vermicompost; BC: biochar; VCL: Vermicompost leachate; VCH: Vermicompost humates; VW: Vermiwash; AA: Ascorbic acid; EC: Electrical conductivity; OM: Organic matter; TN: Total nitrogen; TP: Total phosphorus; TK: Total potassium; AP: Available phosphorus; AK: Available potassium.
Table 3.
Effects of vermicompost alone and in combination with other soil amendments on plant growth performance under drought stress.
Table 3.
Effects of vermicompost alone and in combination with other soil amendments on plant growth performance under drought stress.
| Type of Fertilizer | Characteristics of Fertilizer | Application Rate | Drought Conditions | Test Plant | Impact | References |
|---|---|---|---|---|---|---|
| Wheat Straw VC | pH: 7.55, EC: 2.26 dS/m, N: 0.85%, P: 0.45%, K: 0.83%, Ca: 2.91%, S: 0.23% | 0, 4, 6, 8 t ha−1 | 30, 45, 70% FC | Triticum aestivum L. | VC application at 8 t ha−1 increased SOD activity (14.28%), POD (27.28%), and CAT (50%). Photosynthesis and transpiration rate increased by 27.65 and 49.25%, respectively. | [158] |
| VC | pH: 8.1, EC: 6.5 dS/m, OM: 65.5%, N: 2.2%, P: 1.7%, K: 1.5% | 0, 2.5 and 5% | No stress, moderate drought, and severe drought at 100, 50 and 25% of FC | Lactuca sativa var. crispa | VC increased SOD and CAT enzyme activities and lowered MDA content under drought stress. Chlorophyll a, chlorophyll b, and carotenoid contents increased in lettuce. | [159] |
| VC | pH: 7.1, EC: 1.5 dS/m, C/N: 15.5, N: 0.9–3%, P: 0.9–2.5%, K: 0.6–2.5%, Ca: 4.85–8%, | VC to soil ratio i.e., 0:100, 10:90, 20:80, and 30:70 | No stress, moderate drought (MS), and severe drought (SS) at 100, 75, and 25% of FC, respectively | Cicer arietinum L. | VC addition of 10 and 20% significantly enhanced the chlorophyll contents and Fv/Fm under MS and Fv/Fm, Ci, and PN under SS at the flowering stage. | [150] |
| C + VC + AMF | VC: pH: 7.4, EC: 2.2 dS/m, C/N: 25.8, TN: 1.32%, TP: 0.4%, TK: 0.4%, Fe: 1100 ppm, Zn: 50 ppm C: pH: 7.5, EC: 2 dS/m, C/N: 21.4, TN: 1.9%, TP: 0.8%, TK: 0.8%, Fe: 1000 ppm, Zn: 46 ppm | VC (4% w/w) and inoculation with and without AMF | Drought stress at three levels, i.e., well-watered: D0 (no drought), moderate stress: D1 and severe stress: D2 | Pistachio seedlings | VC and C increased plant growth (232% and 29%) and nutrient uptake (30% and 52% P uptake and 35% and 1% Zn uptake) under severe drought stress conditions. | [160] |
| VC + humic acid | - | VC (0, 25, and 50%) and humic acid (0, 50, and 100 mg/L) | 50 and 100% FC | Catharanthus roseous | VC and humic acid reduced the drought stress, improved vegetative indices, cell membrane stability index, and plant pigment content, and decreased proline content. | [161] |
| VC | pH: 7.1, EC: 1.5 dS/m, C/N: 17.5, N: 3.0%, P: 0.9%, K: 1.2%, Ca: 4.5%, Mg: 0.50% | 0, 10, 20, 30% vermicompost to soil ratio | 75, 50, and 25% FC | Cicer arietinum L. cv. Pirouz | VC at 30% increased the photosynthetic pigments, CO2 assimilation rate, transpiration, Fv/Fm, Ca, and K in root and leaf tissues, and proline and soluble protein contents in root tissues. | [152] |
| C + CV | - | C: 20 and 30% VC: 20 and 30% | 80, 60 and 40% AWC | Calendula officinalis L. cv. Candyman Orange | Total dry mass and water use efficiency was about 3-fold higher in plants grown in 30% VC or 30% manure compost/ | [162] |
VC: Vermicompost; C: compost; FC: field capacity; SOD: superoxide dismutase; POD: peroxidase; CAT: catalase; MDA: malondialdehyde; Fv/Fm: maximal quantum yield of PSII photochemistry; Ci: intercellular CO2 concentration; PN: net photosynthetic rate; AWC: available water content.
Table 4.
Phytopathogen-suppressing potential of vermicompost and its derivatives produced from various waste materials.
Table 4.
Phytopathogen-suppressing potential of vermicompost and its derivatives produced from various waste materials.
| Type of Fertilizer | Parental Wastes | Earthworm | Phytopathogens | Plants | Pest Control | References |
|---|---|---|---|---|---|---|
| VC | CD | E. fetida | Meloidogyne incognita | Solanum lycopersicum L. | Root-knot nematode | [177] |
| VC, N, P, M, S | CD, VR | Lumbricus rubellus | Plutella xylostella | Brassica oleracea L. | Diamondback moth | [178] |
| VC, VL | CD | E. fetida | Meloidogyne incognita | Withania somnifera | Root parasitic nematode | [179] |
| VC | CD, TL, SD | E. fetida, Lumbricus rubellus, Perionyx ecavatus | Meloidogyne incognita | Solanum lycopersicum L. | Root parasitic nematode | [180] |
| VE | FW | - | Myzus persicae, Planococcus citri, etranychus urticae | Solanum lycopersicum L. and Cucumis sativa | Green peach aphid, citrus mealybug, two spotted spider mites | [181] |
| VC | Ipomoea leaves | E. fetida | Earias vittella | Abelmoschus esculentus | Fruit borer | [182] |
| VE | CD | E. fetida | Meloidogyne incognita | Capsicum annuum L. and Solanum lycopersicum L. | Root-knot nematode | [183] |
| VC, VE | GW | - | Globodera rostochiensis and Globodera pallida | Solanum tuberosum | Potato-cyst nematodes | [171] |
| VC | CD | - | Aphis gossypii Glover | Cucumis sativus L. | Melon aphid | [184] |
| VC | BL, CD | Pheretima posthuma | Phyllocnistis citrella | C. reticulata | Citrus leaf miner | [185] |
| VC | FW | - | Myzus persicae, Pseudococcus sp. and Pieris brassicae L. | Capsicum annuum L., Lycopersicon esculentum and Brassica oleracea L. | Aphids, mealy bugs, and cabbage white caterpillars | [186] |
CD: cow dung; VR: vegetable residue; N: neem leaves; P: papaya leaves; M: marigold leaves; S: soursop leaves; VL: vermicompost leachates; TL: Taro leaves; SD: sawdust; VE: vermicompost extracts; FW: food wastes; GW: green wastes; BL: banana leaves.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rehman, S.u.; De Castro, F.; Aprile, A.; Benedetti, M.; Fanizzi, F.P. Vermicompost: Enhancing Plant Growth and Combating Abiotic and Biotic Stress. Agronomy 2023, 13, 1134. https://doi.org/10.3390/agronomy13041134
AMA Style
Rehman Su, De Castro F, Aprile A, Benedetti M, Fanizzi FP. Vermicompost: Enhancing Plant Growth and Combating Abiotic and Biotic Stress. Agronomy. 2023; 13(4):1134. https://doi.org/10.3390/agronomy13041134
Chicago/Turabian StyleRehman, Sami ur, Federica De Castro, Alessio Aprile, Michele Benedetti, and Francesco Paolo Fanizzi. 2023. "Vermicompost: Enhancing Plant Growth and Combating Abiotic and Biotic Stress" Agronomy 13, no. 4: 1134. https://doi.org/10.3390/agronomy13041134
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.