Enhanced Antioxidant Activity and Secondary Metabolite Production in Tartary Buckwheat under Polyethylene Glycol (PEG)-Induced Drought Stress during Germination

 , ,
, ,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Effect of PEG-Induced Drought Stress on Seed Germination and Seedling Morphology
2.1.1. Germination Percentage
2.1.2. Relative Water Content
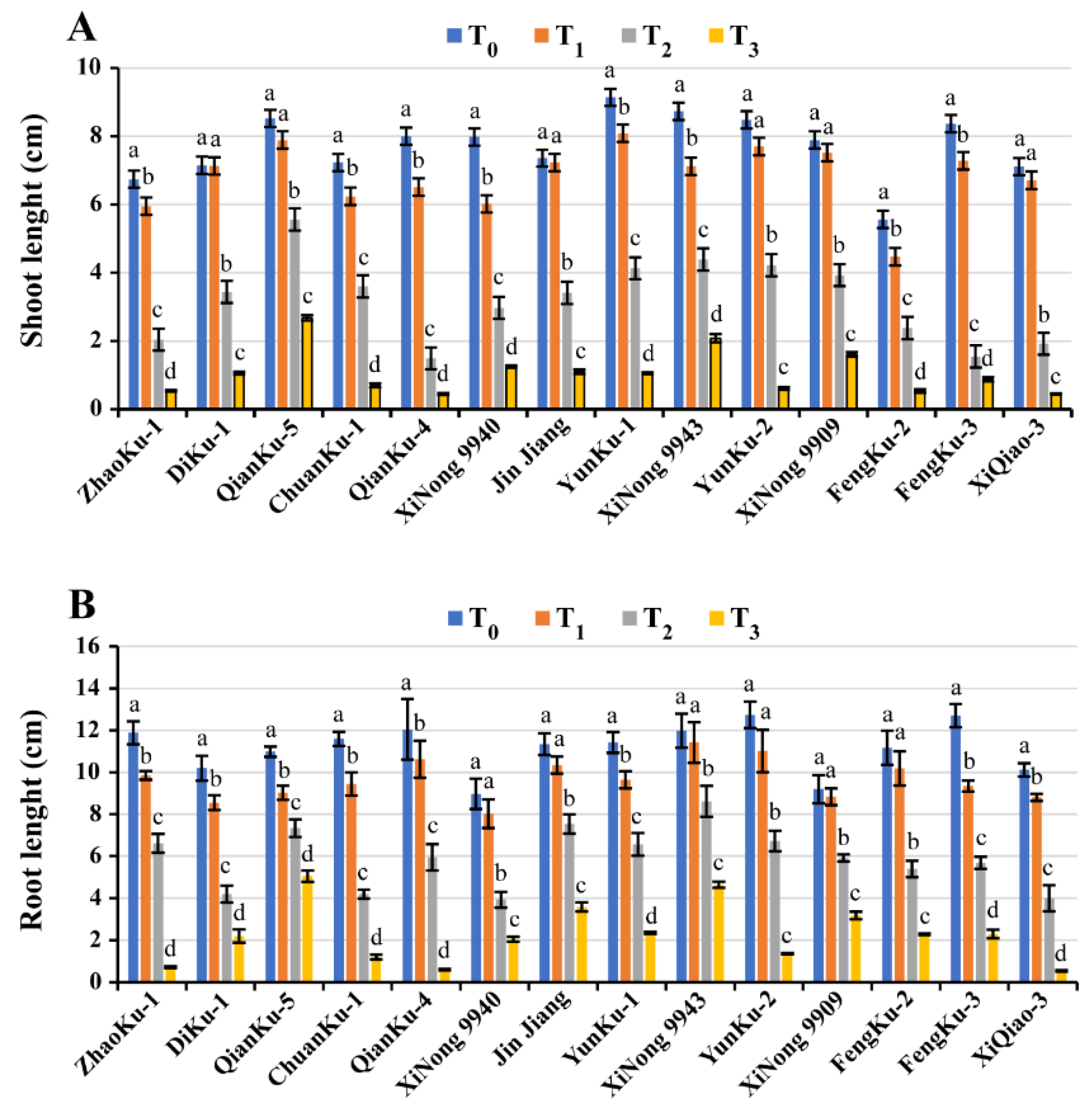
2.1.3. Shoots and Roots Length
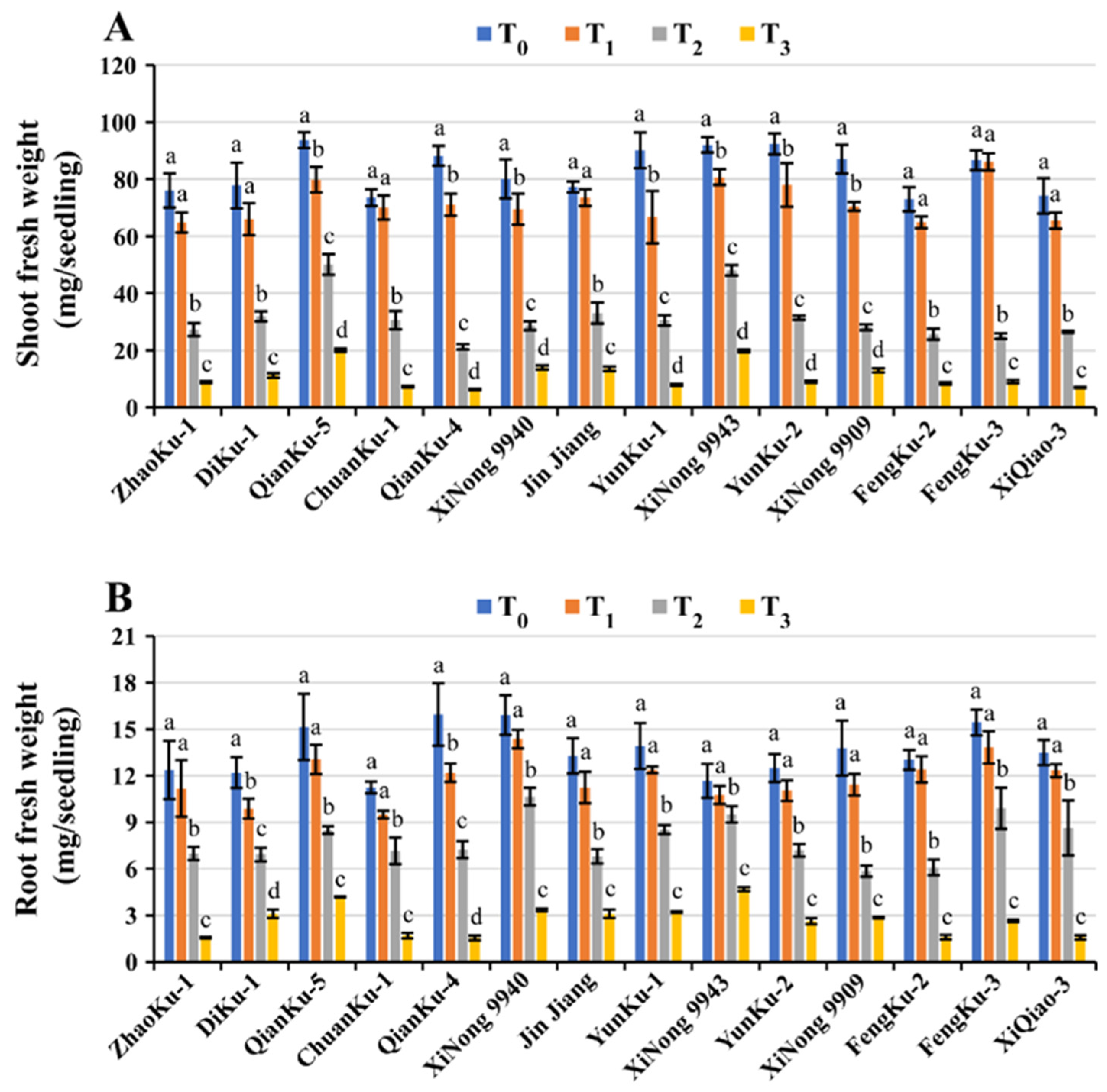
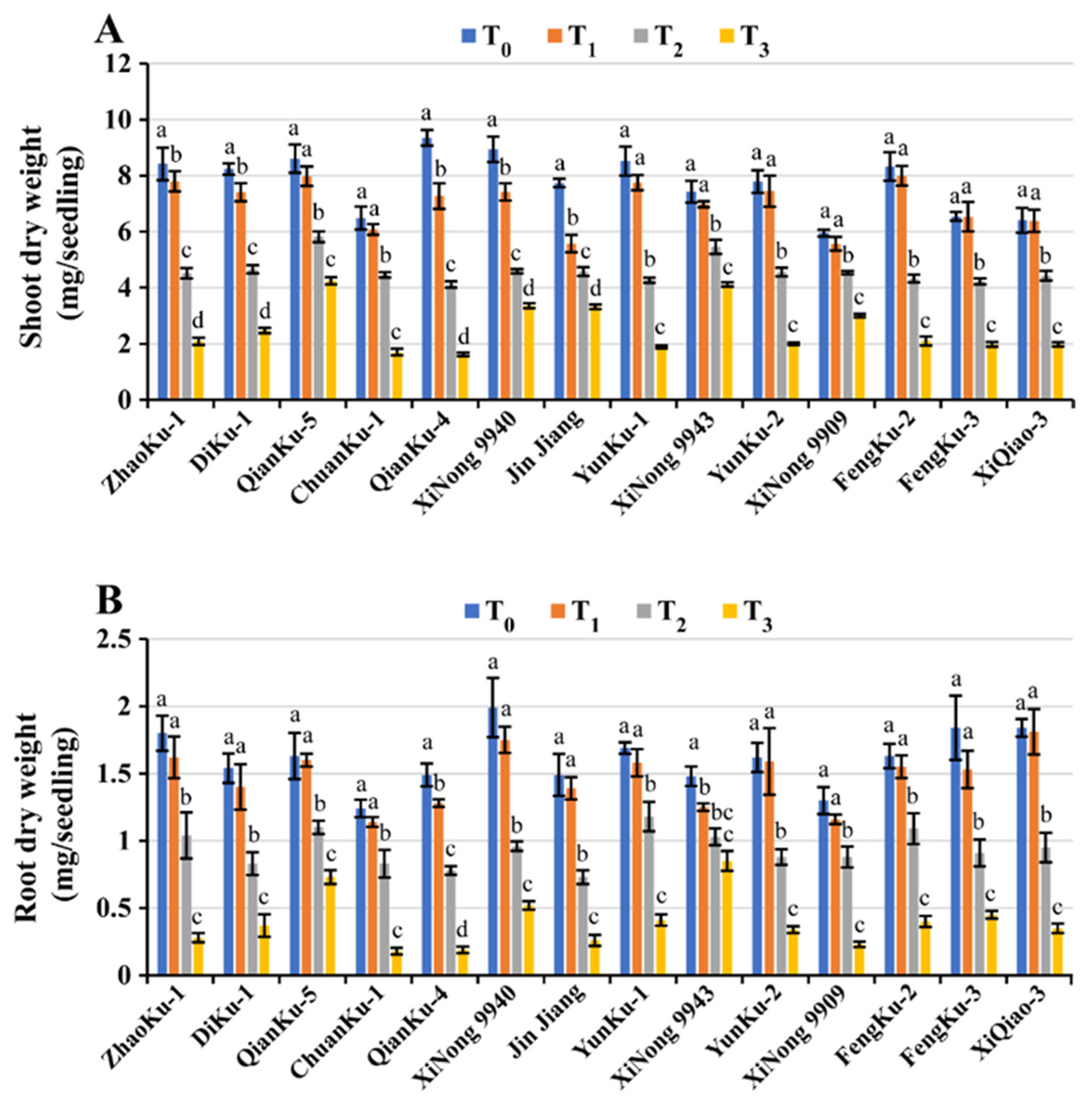
2.1.4. Fresh and Dry Weight of Shoots and Roots
2.2. Measurement of Stress Tolerance Index Based on Seed Germination and Seedling Morphology

2.3. Effect of PEG-Induced Drought Stress on Physiological and Biochemical Mechanisms


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | GM (%) | Shoot Length (mm) | Root Length (mm/Seedling) | SFW (mg/Seedling) | RFW (mg/Seedling) | SDW (mg/Seedling) | RDW (mg/Seedling) | RWC (%) | STI Rank Summation Index | Ranking |
|---|---|---|---|---|---|---|---|---|---|---|
| XiNong 9943 | 79.55 | 51.89 | 68.67 | 53.81 | 71.32 | 74.29 | 70.50 | 89.75 | 1.63 | 1 |
| QianKu-5 | 85.56 | 63.11 | 64.97 | 53.37 | 56.69 | 69.88 | 70.14 | 88.01 | 2.38 | 2 |
| XiNong 9940 | 73.33 | 42.77 | 52.03 | 46.63 | 59.42 | 57.31 | 54.10 | 83.71 | 6.88 | 3 |
| Jin Jiang | 72.41 | 53.17 | 63.08 | 51.85 | 53.11 | 58.05 | 53.24 | 82.53 | 6.88 | 4 |
| YunKu-2 | 71.86 | 49.25 | 49.92 | 42.73 | 55.63 | 59.95 | 57.82 | 83.62 | 7.00 | 5 |
| XiNong 9909 | 66.24 | 55.18 | 65.00 | 42.72 | 48.72 | 73.50 | 58.21 | 79.28 | 7.13 | 6 |
| YunKu-1 | 66.95 | 48.43 | 54.14 | 38.86 | 57.80 | 54.42 | 62.52 | 82.51 | 7.50 | 7 |
| DiKu-1 | 64.36 | 54.22 | 48.90 | 46.75 | 54.43 | 58.86 | 56.28 | 80.25 | 7.63 | 8 |
| FengKu-2 | 51.00 | 44.30 | 53.30 | 45.20 | 51.46 | 57.74 | 62.17 | 84.26 | 8.38 | 9 |
| ChuanKu-1 | 60.23 | 48.59 | 42.68 | 48.89 | 54.42 | 62.92 | 57.80 | 80.09 | 8.75 | 10 |
| XiQiao-3 | 52.19 | 42.57 | 43.98 | 44.47 | 55.70 | 66.56 | 56.34 | 82.78 | 8.88 | 11 |
| FengKu-3 | 63.79 | 38.67 | 45.46 | 46.18 | 56.95 | 64.89 | 52.36 | 81.13 | 9.13 | 12 |
| ZhaoKu-1 | 61.17 | 42.19 | 48.20 | 44.22 | 53.17 | 57.05 | 54.44 | 85.18 | 9.63 | 13 |
| QianKu-4 | 52.54 | 35.21 | 47.47 | 37.24 | 43.85 | 46.38 | 50.34 | 79.93 | 13.25 | 14 |
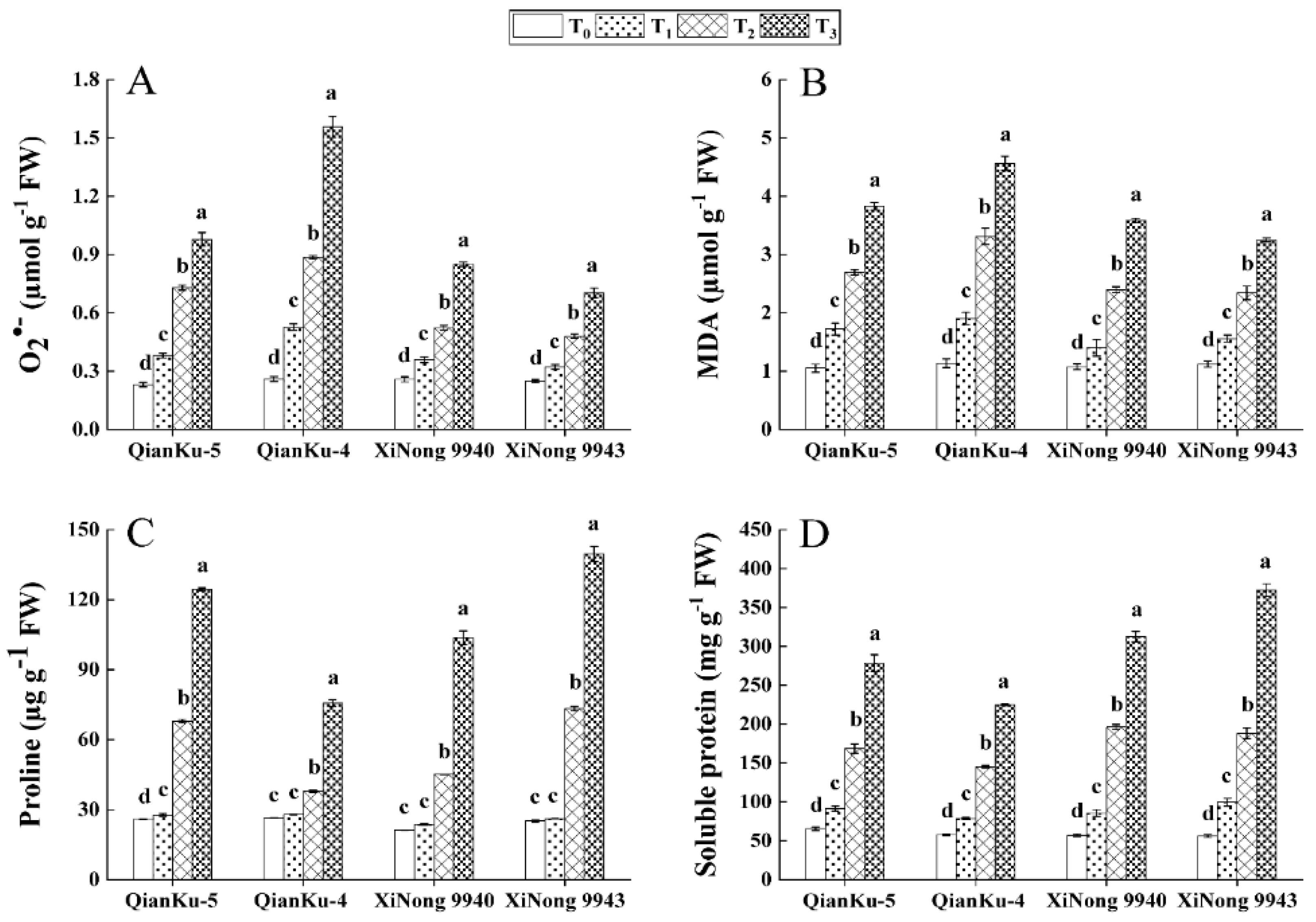
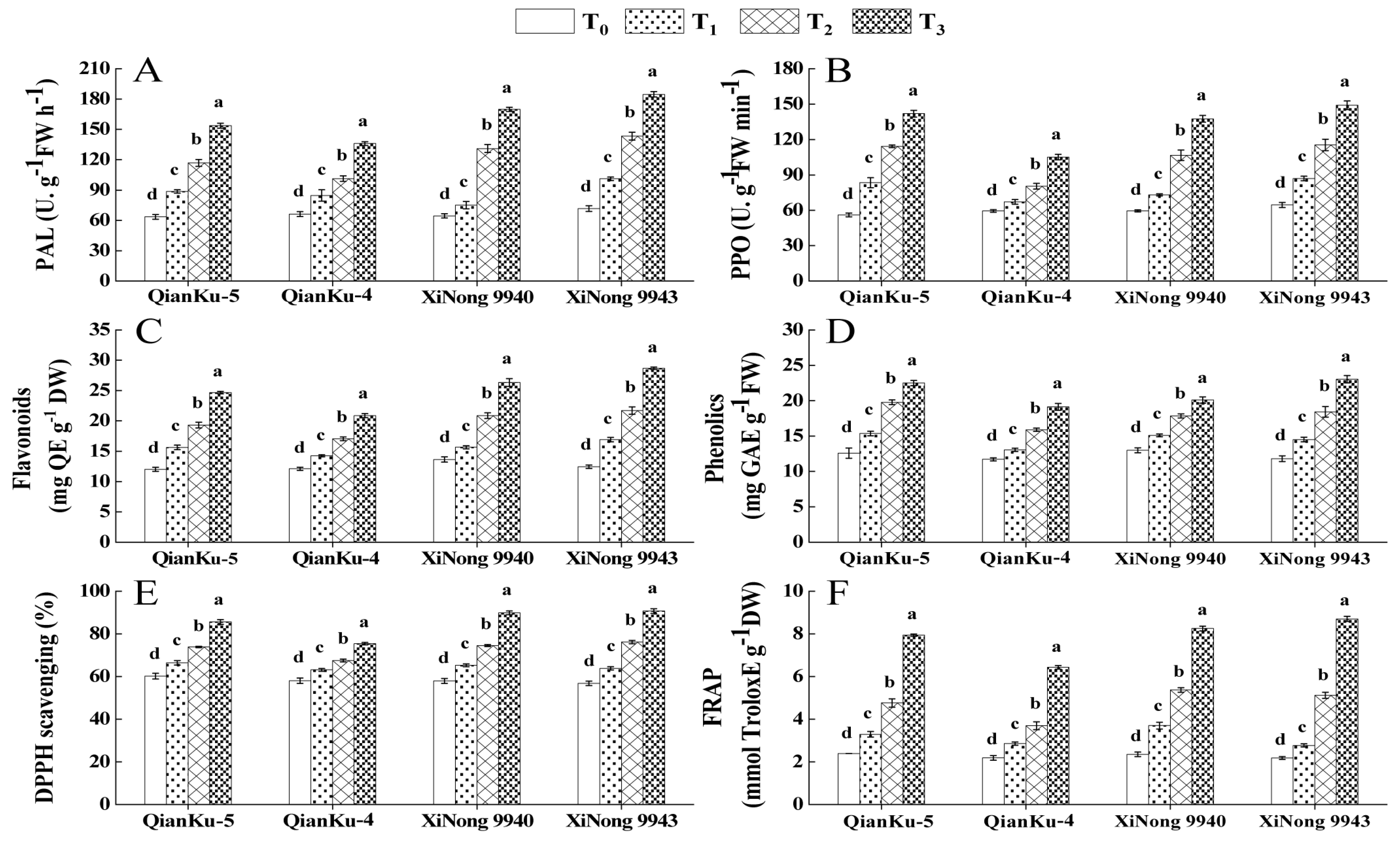
2.3.1. ROS, MDA, and Osmotic Solutes Content

2.3.2. Activities of Enzymatic Antioxidants
2.3.3. Secondary Metabolites as Antioxidants

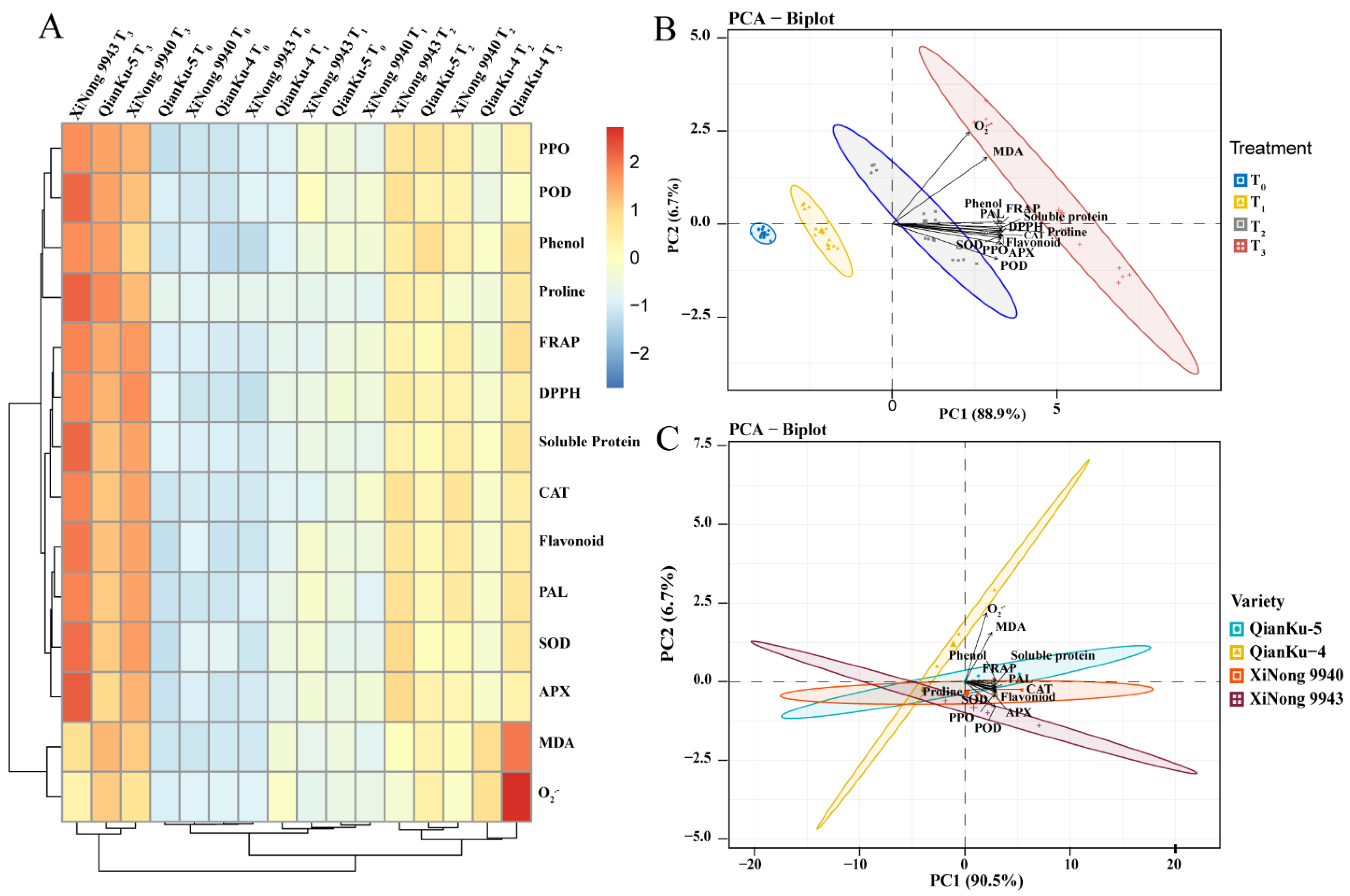
2.3.4. Pearson’s Correlation Analysis
2.3.5. Heatmap and PCA

3. Discussion
4. Materials and Methods
4.1. Experimental Materials and Locations
4.2. Germination Percentage, Morphological Parameter, and Relative Water Content
4.3. Measurement of Stress Tolerance Index
4.4. Determination of Superoxide Anion, MDA Content, Antioxidant Activity, and Osmotic Solutes
4.5. Estimation of Total Antioxidant Capacity and Non-Enzymatic Antioxidant Activity
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hossain, M.S.; Li, J.; Sikdar, A.; Hasanuzzaman, M.; Uzizerimana, F.; Muhammad, I.; Yuan, Y.; Zhang, C.; Wang, C.; Feng, B. Exogenous melatonin modulates the physiological and biochemical mechanisms of drought tolerance in Tartary buckwheat (Fagopyrum tataricum (L.) Gaertn). Molecules 2020, 25, 2828. [Google Scholar] [CrossRef]
- Jain, C.; Saxena, R. Varietal differences against PEG induced drought stress in cowpea. Octa J. Environ. Res. 2016, 4, 58–62. [Google Scholar]
- Muscolo, A.; Sidari, M.; Anastasi, U.; Santonoceto, C.; Maggio, A. Effect of PEG-induced drought stress on seed germination of four lentil genotypes. J. Plant Interact. 2014, 9, 354–363. [Google Scholar] [CrossRef]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Wu, C.; Wang, Q.; Xie, B.; Wang, Z.; Cui, J.; Hu, T. Effects of drought and salt stress on seed germination of three leguminous species. Afr. J. Biotechnol. 2011, 10, 17954–17961. [Google Scholar]
- Li, J.; Hossain, M.S.; Ma, H.; Yang, Q.; Gong, X.; Yang, P.; Feng, B. Comparative metabolomics reveals differences in flavonoid metabolites among different coloured buckwheat flowers. J. Food Compos. Anal. 2020, 85, 103335. [Google Scholar] [CrossRef]
- Monshi, F.I.; Khan, N.; Kimura, K.; Seita, S.; Yamamoto, Y.; Katsube-Tanaka, T. Structure and diversity of 13S globulin zero-repeat subunit, the trypsin-resistant storage protein of common buckwheat (Fagopyrum esculentum M.) seeds. Breed. Sci. 2020, 70, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Monshi, F.I.; Katsube-Tanaka, T. 2S albumin g13 polypeptide, less related to Fag e 2, can be eliminated in common buckwheat (Fagopyrum esculentum Moench) seeds. Food Chem. Mol. Sci. 2022, 5, 100138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.L.; Zhou, M.L.; Tang, Y.; Li, F.L.; Tang, Y.X.; Shao, J.R.; Xue, W.T.; Wu, Y.M. Bioactive compounds in functional buckwheat food. Food Res. Int. 2012, 49, 389–395. [Google Scholar] [CrossRef]
- Katsube-Tanaka, T.; Monshi, F.I. Characterization of 2S albumin allergenic proteins for anaphylaxis in common buckwheat. Food Chem. Mol. Sci. 2022, 5, 100127. [Google Scholar] [CrossRef] [PubMed]
- Okçu, G.; Kaya, M.D.R.; Atak, M. Effects of salt and drought stresses on germination and seedling growth of pea (Pisum sativum L.). Turk. J. Agric. For. 2005, 29, 237–242. [Google Scholar]
- Kaya, M.D.; Okçu, G.; Atak, M.; Çıkılı, Y.; Kolsarıcı, Ö. Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Eur. J. Agron. 2006, 24, 291–295. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Vanzolini, S.; Araki, C.A.D.S.; Silva, A.C.T.M.D.; Nakagawa, J. Seedling length test in the evaluation of the physiological quality of soybean seeds. Rev. Bras. Sementes 2007, 29, 90–96. [Google Scholar] [CrossRef]
- Henning, F.A.; Mertz, L.M.; Jacob Junior, E.A.; Machado, R.D.; Fiss, G.; Zimmer, P.D. Chemical composition and reserve mobilization in soybean seeds with high and low vigor. Bragantia 2010, 69, 727–734. [Google Scholar] [CrossRef]
- Mahpara, S.; Zainab, A.; Ullah, R.; Kausar, S.; Bilal, M.; Latif, M.I.; Arif, M.; Akhtar, I.; Al-Hashimi, A.; Elshikh, M.S.; et al. The impact of PEG-induced drought stress on seed germination and seedling growth of different bread wheat (Triticum aestivum L.) genotypes. PLoS ONE 2022, 17, e0262937. [Google Scholar] [CrossRef]
- Mustamu, N.E.; Tampubolon, K.; Alridiwirsah; Basyuni, M.; Al-Taey, D.K.A.; Jawad, K.A.J.H.; Mehdizadeh, M. Drought stress induced by polyethylene glycol (PEG) in local maize at the early seedling stage. Heliyon 2023, 9, e20209. [Google Scholar] [CrossRef]
- Pavli, O.I.; Foti, C.; Skoufogianni, G.; Karastergiou, G.; Panagou, A.; Khah, E.M. PEG-induced drought stress during germination: Effects on soybean germplasm. Agric. Res. Technol. Open Access J. 2020, 23, 556250. [Google Scholar] [CrossRef]
- Basal, O.; Szabó, A.; Veres, S. PEG-induced drought stress effects on soybean germination parameters. J. Plant Nutr. 2020, 43, 1768–1779. [Google Scholar] [CrossRef]
- Nestorović Živković, J.; Aničić, N.; Matekalo, D.; Skorić, M.; Filipović, B.; Marković, T.; Dmitrović, S. Polyethylene glycol (PEG)-induced dehydration alters enzymatic and non-enzymatic components of the antioxidant defense system in Nepeta nervosa royle ex bentham. Horticulturae 2023, 9, 1277. [Google Scholar] [CrossRef]
- Wang, W.B.; Kim, Y.H.; Lee, H.S.; Kim, K.Y.; Deng, X.P.; Kwak, S.S. Analysis of antioxidant enzyme activity during germination of alfalfa under salt and drought stresses. Plant Physiol. Biochem. 2009, 47, 570–577. [Google Scholar] [CrossRef]
- Falleh, H.; Jalleli, I.; Ksouri, R.; Boulaaba, M.; Guyot, S.; Magné, C.; Abdelly, C. Effect of salt treatment on phenolic compounds and antioxidant activity of two Mesembryanthemum edule provenances. Plant Physiol. Biochem. 2012, 52, 1–8. [Google Scholar] [CrossRef]
- Lian, H.; Qin, C.; Shen, J.; Ahanger, M.A. Alleviation of adverse effects of drought stress on growth and nitrogen metabolism in mungbean (Vigna radiata) by sulphur and nitric oxide involves up-regulation of antioxidant and osmolyte metabolism and gene expression. Plants 2023, 12, 3082. [Google Scholar] [CrossRef]
- Jothimani, K.; Arulbalachandran, D. Physiological and biochemical studies of black gram (Vigna mungo (L.) Hepper) under polyethylene glycol induced drought stress. Biocatal. Agric. Biotechnol. 2020, 29, 101777. [Google Scholar] [CrossRef]
- Tovar, M.J.; Romero, M.P.; Girona, J.; Motilva, M.J. L-Phenylalanine ammonia-lyase activity and concentration of phenolics in developing olive (Olea europaea L. cv Arbequina) fruit grown under different irrigation regimes. J. Sci. Food Agric. 2002, 82, 892–898. [Google Scholar] [CrossRef]
- Elahi, N.N.; Farrukh, F.; Jalaluddin, S.; Ahmed, H.M.; Saima, S.; Mustafa, S.; Alahmadi, T.A.; Ansari, M.J.; Battaglia, M.L.; Danish, S. Comparing the positive impacts and stress induction by polyethylene glycol (PEG 6000) variable levels on canola (Brassica napus L.) growth, yield, and oil contents. ACS Omega 2023, 8, 29046–29059. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Wei, F.; Qin, S.; Khan, A.; Kashif, M.H.; Zhou, R. Polyethylene glycol induced drought stress strongly influences seed germination, root morphology and cytoplasm of different kenaf genotypes. Ind. Crops Prod. 2019, 137, 180–186. [Google Scholar] [CrossRef]
- Vendruscolo, E.C.G.; Schuster, I.; Pileggi, M.; Scapim, C.A.; Molinari, H.B.C.; Marur, C.J.; Vieira, L.G.E. Stress-induced synthesis of proline confers tolerance to water deficit in transgenic wheat. J. Plant Physiol. 2007, 164, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Partheeban, C.; Chandrasekhar, C.N.; Jeyakumar, P.; Ravikesavan, R.; Gnanam, R. Effect of PEG-induced drought stress on seed germination and seedling characters of maize (Zea mays L.) genotypes. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1095–1104. [Google Scholar] [CrossRef]
- Souza, R.P.; Machado, E.C.; Silva, J.A.B.; Lagôa, A.M.M.A.; Silveira, J.A.G. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Exp. Bot. 2004, 51, 45–56. [Google Scholar] [CrossRef]
- Peršić, V.; Ament, A.; Antunović, D.J.; Drezner, G.; Cesar, V. PEG-induced physiological drought for screening winter wheat genotypes sensitivity-integrated biochemical and chlorophyll a fluorescence analysis. Front. Plant Sci. 2022, 13, 987702. [Google Scholar] [CrossRef] [PubMed]
- Bilgili, D.; Atak, M.; Mavi, K. Effects of PEG-induced drought stress on germination and seedling performance of bread wheat genotypes. YYU J. Agric. Sci. 2017, 29, 765–771. [Google Scholar] [CrossRef]
- Hou, M.; Ma, M. Effect of PEG-simulated Drought stress on seed germination of three medicinal liquorice (Glycyrrhiza) species. Legume Res. 2022, 45, 1388–1393. [Google Scholar] [CrossRef]
- Darkwa, K.; Ambachew, D.; Mohammed, H.; Asfaw, A.; Blair, M.W. Evaluation of common bean (Phaseolus vulgaris L.) genotypes for drought stress adaptation in Ethiopia. Crop J. 2016, 4, 367–376. [Google Scholar] [CrossRef]
- Steiner, F.; Zuffo, A.M.; Zoz, T.; Zoz, A.; Zoz, J. Drought tolerance of wheat and black oat crops at early stages of seedling growth. Amaz. J. Agric. Environ. Sci. 2017, 40, 576–585. [Google Scholar] [CrossRef]
- Arjenaki, F.G.; Amini Dehaghi, M.; Jabbari, A.R. Effects of priming on seed germination of marigold (Calendula officinalis). Adv. Environ. Biol. 2011, 5, 276–280. [Google Scholar]
- Koskosidis, A.; Ebrahim, K.H.A.H.; Mavromatis, A.; Pavli, O.; Vlachostergios, D.N. Effect of PEG-induced drought stress on germination of ten chickpea (Cicer arietinum L.) genotypes. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 294–304. [Google Scholar] [CrossRef]
- Carvalho, M.; Matos, M.; Castro, I.; Monteiro, E.; Rosa, E.; Lino-Neto, T.; Carnide, V. Screening of worldwide cowpea collection to drought tolerant at a germination stage. Sci. Hortic. 2019, 247, 107–115. [Google Scholar] [CrossRef]
- Lotfi, N.; Soleimani, A.; Vahdati, K.; Çakmakçı, R. Comprehensive biochemical insights into the seed germination of walnut under drought stress. Sci. Hortic. 2019, 250, 329–343. [Google Scholar] [CrossRef]
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. Int. Agrophysics 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Han, B.; Duan, X.; Wang, Y.; Zhu, K.; Zhang, J.; Wang, R.; Hu, H.; Qi, F.; Pan, J.; Yan, Y.; et al. Methane protects against polyethylene glycol-induced osmotic stress in maize by improving sugar and ascorbic acid metabolism. Sci. Rep. 2017, 7, 46185. [Google Scholar] [CrossRef]
- Li, Z.; Su, X.; Chen, Y.; Fan, X.; He, L.; Guo, J.; Wang, Y.; Yang, Q. Melatonin improves drought resistance in maize seedlings by enhancing the antioxidant system and regulating abscisic acid metabolism to maintain stomatal opening under PEG-induced drought. J. Plant Biol. 2021, 64, 299–312. [Google Scholar] [CrossRef]
- Silva, P.; Matos, M. Assessment of the impact of aluminum on germination, early growth and free proline content in Lactuca sativa L. Ecotoxicol. Environ. Saf. 2016, 131, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Othmani, A.; Ayed, S.; Chamekh, Z.; Slama-Ayed, O.; Teixeira Da Silva, A.J.; Rezgui, M.; Slim-Amara, H.; Younes, M.B. Screening of seedlings of durum wheat (Triticum durum Desf.) cultivars for tolerance to PEG-induced drought stress. Pak. J. Bot. 2021, 53, 823–832. [Google Scholar] [CrossRef]
- Altaf, A.; Gull, S.; Zhu, X.; Zhu, M.; Rasool, G.; Ibrahim, M.E.H.; Aleem, M.; Uddin, S.; Saeed, A.; Shah, A.Z.; et al. Study of the effect of peg-6000 imposed drought stress on wheat (Triticum aestivum L.) cultivars using relative water content (RWC) and proline content analysis. Pak. J. Agric. Sci. 2021, 58, 357–367. [Google Scholar]
- Miranda, R.D.S.; Fonseca, B.S.F.D.; Pinho, D.S.; Batista, J.Y.N.; Brito, R.R.D.; Silva, E.M.D.; Ferreira, W.S.; Costa, J.H.; Lopes, M.D.; Sousa, R.H.B.D.; et al. Selection of soybean and cowpea cultivars with superior performance under drought using growth and biochemical aspects. Plants 2023, 12, 3134. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shi, S. Physiological and proteomic responses of contrasting alfalfa (Medicago sativa L.) varieties to PEG-induced osmotic stress. Front. Plant Sci. 2018, 9, 242. [Google Scholar] [CrossRef]
- Zhuang, W.; Li, J.; Cao, M.; Feng, W.; Li, Y. Effects of salt-drought intercross stress on physiological and biochemical characteristics of Ammodendron argenteum (Pall.) Kuntze seedlings. J. Wuhan Bot. Res. 2010, 28, 730–736. [Google Scholar]
- Tian, Z.; Wang, F.; Zhang, W.; Liu, C.; Zhao, X. Antioxidant mechanism and lipid peroxidation patterns in leaves and petals of marigold in response to drought stress. Hortic. Environ. Biotechnol. 2012, 53, 183–192. [Google Scholar] [CrossRef]
- Mahmud, S.; Kamruzzaman, M.; Bhattacharyya, S.; Alharbi, K.; Abd El Moneim, D.; Mostofa, M.G. Acetic acid positively modulates proline metabolism for mitigating PEG-mediated drought stress in maize and arabidopsis. Front. Plant Sci. 2023, 14, 1167238. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef]
- Umezawa, T.; Fujita, M.; Fujita, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Engineering drought tolerance in plants: Discovering and tailoring genes to unlock the future. Curr. Opin. Biotechnol. 2006, 17, 113–122. [Google Scholar] [CrossRef]
- Duhan, S.; Kumari, A.; Lal, M.; Sheokand, S. Oxidative stress and antioxidant defense under combined waterlogging and salinity stresses. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Wiley: Hoboken, NJ, USA, 2019; pp. 113–142. [Google Scholar]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Chen, K.; Arora, R. Dynamics of the antioxidant system during seed osmopriming, post-priming germination, and seedling establishment in spinach (Spinacia oleracea). Plant Sci. 2011, 180, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.A.; Javed, R.; Adeel, M.; Rizwan, M.; Yang, Y. PEG 6000-stimulated drought stress improves the attributes of in vitro growth, steviol glycosides production, and the activity of enzyme in Stevia rebaudiana Bertoni. Plants 2020, 9, 1552. [Google Scholar] [CrossRef] [PubMed]
- Kakar, H.A.; Ullah, S.; Shah, W.; Ali, B.; Satti, S.Z.; Ullah, R.; Muhammad, Z.; Eldin, S.M.; Ali, I.; Alwahibi, M.S.; et al. Seed priming modulates physiological and agronomic attributes of maize (Zea mays L.) under induced polyethylene glycol osmotic stress. ACS Omega 2023, 8, 22788–22808. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Shen, Y.; Ni, Z.; Wang, Q.; Lei, Z.; Xu, N.; Deng, Q.; Lin, L.; Wang, J.; Lv, X.; et al. Exogenous melatonin application delays senescence of kiwifruit leaves by regulating the antioxidant capacity and biosynthesis of flavonoids. Front. Plant Sci. 2018, 9, 426. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.; Qamar, M.A.; Bokhari, T.H.; Abbas, M.; Hussain, F.; Masood, N.; Keshavarzi, A.; Qureshi, N.; Nazir, A. Total phenolic, chromium contents and antioxidant activity of raw and processed sugars. Inf. Process. Agric. 2017, 4, 83–89. [Google Scholar] [CrossRef]
- Huang, Y.C.; Chang, Y.H.; Shao, Y.Y. Effects of genotype and treatment on the antioxidant activity of sweet potato in Taiwan. Food Chem. 2006, 98, 529–538. [Google Scholar] [CrossRef]
- Krol, A.; Amarowicz, R.; Weidner, S. Changes in the composition of phenolic compounds and antioxidant properties of grapevine roots and leaves (Vitis vinifera L.) under continuous long-term drought stress. Acta Physiol. Plant. 2014, 36, 1491–1499. [Google Scholar] [CrossRef]
- Gabr, A.M.M.; Sytar, O.; Ahmed, A.R.; Smetanska, I. Production of phenolic acid and antioxidant activity in transformed hairy root cultures of common buckwheat (Fagopyrum esculentum M.). Aust. J. Basic Appl. Sci. 2012, 6, 577–586. [Google Scholar]
- Golkar, P.; Bakhtiari, M.A.; Bazarganipour, M. The effects of nanographene oxide on the morpho-biochemical traits and antioxidant activity of Lepidium sativum L. under in vitro salinity stress. Sci. Hortic. 2021, 288, 110301. [Google Scholar] [CrossRef]
- Sharma, J.; Kumar, S.; Kumar, V.; Singh, P.; Khyalia, P.; Verma, S.; Saini, S.; Sharma, A. Foliar application of glycine betaine to ameliorate lead toxicity in barley plants by modulating antioxidant enzyme activity and biochemical parameters. Environ. Res. Commun. 2023, 5, 075002. [Google Scholar] [CrossRef]
- Yao, X.; Zhou, M.; Ruan, J.; Peng, Y.; Ma, C.; Wu, W.; Gao, A.; Weng, W.; Cheng, J. Physiological and biochemical regulation mechanism of exogenous hydrogen peroxide in alleviating NaCl stress toxicity in Tartary buckwheat (Fagopyrum tataricum (L.) Gaertn). Int. J. Mol. Sci. 2022, 23, 10698. [Google Scholar] [CrossRef] [PubMed]
- Meng, H.L.; Sun, P.Y.; Wang, J.R.; Sun, X.Q.; Zheng, C.Z.; Fan, T.; Chen, Q.F.; Li, H.Y. Comparative physiological, transcriptomic, and WGCNA analyses reveal the key genes and regulatory pathways associated with drought tolerance in Tartary buckwheat. Front. Plant Sci. 2022, 13, 985088. [Google Scholar] [CrossRef] [PubMed]
- Oksana, S.; Marek, K.; Marian, B.; Marek, Z. Cultivar-dependent and drought-induced modulation of secondary metabolites, adaptative defense in Fagopyrum esculentum L. Physiol. Mol. Biol. Plants 2022, 29, 1605–1618. [Google Scholar] [CrossRef] [PubMed]
- Bowman, W.D. The relationship between leaf water status, gas exchange, and spectral reflectance in cotton leaves. Remote Sens. Environ. 1989, 30, 249–255. [Google Scholar] [CrossRef]
- Subhani, G.M.; Ahmad, J.; Anwar, J.; Hussain, M.; Mahmood, A. Identification of drought tolerant genotypes of barley (Hordeum vulgare L.) through stress tolerance indices. JAPS J. Anim. Plant Sci. 2015, 25, 686–692. [Google Scholar]
- Škerget, M.; Kotnik, P.; Hadolin, M.; Hraš, A.R.; Simonič, M.; Knez, Ž. Phenols, proanthocyanidins, flavones and flavonols in some plant materials and their antioxidant activities. Food Chem. 2005, 89, 191–198. [Google Scholar] [CrossRef]
- Ordonez, A.; Gomez, J.; Vattuone, M.; Lsla, M. Antioxidant activities of Sechium edule (Jacq.) Swartz extracts. Food Chem. 2006, 97, 452–458. [Google Scholar] [CrossRef]
- Chen, C.W.; Ho, C.T. Antioxidant properties of polyphenols extracted from green and black teas. J. Food Lipids 1995, 2, 35–46. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (frap) as a measure of “antioxidant power”: The frap assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]



| Parameter | O2•− | MDA | Pro | SP | SOD | POD | CAT | APX | PAL | PPO | Flav | Phenol | DPPH | FRAP |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| O2 | 1 | |||||||||||||
| MDA | 0.94 ** | 1 | ||||||||||||
| PRO | 0.611 ** | 0.766 ** | 1 | |||||||||||
| SP | 0.651 ** | 0.815 ** | 0.952 ** | 1 | ||||||||||
| SOD | 0.597 ** | 0.756 ** | 0.932 ** | 0.956 ** | 1 | |||||||||
| POD | 0.45 ** | 0.645 ** | 0.913 ** | 0.915 ** | 0.91 ** | 1 | ||||||||
| CAT | 0.602 ** | 0.773 ** | 0.911 ** | 0.963 ** | 0.941 ** | 0.919 ** | 1 | |||||||
| APX | 0.556 ** | 0.74 ** | 0.928 ** | 0.96 ** | 0.962 ** | 0.946 ** | 0.963 ** | 1 | ||||||
| PAL | 0.627 ** | 0.801 ** | 0.916 ** | 0.963 ** | 0.956 ** | 0.92 ** | 0.949 ** | 0.967 ** | 1 | |||||
| PPO | 0.58 ** | 0.763 ** | 0.929 ** | 0.942 ** | 0.93 ** | 0.94 ** | 0.939 ** | 0.951 ** | 0.953 ** | 1 | ||||
| Flav | 0.604 ** | 0.78 ** | 0.924 ** | 0.972 ** | 0.96 ** | 0.942 ** | 0.964 ** | 0.974 ** | 0.974 ** | 0.964 ** | 1 | |||
| Phenol | 0.661 ** | 0.808 ** | 0.908 ** | 0.919 ** | 0.886 ** | 0.901 ** | 0.935 ** | 0.924 ** | 0.907 ** | 0.938 ** | 0.923 ** | 1 | ||
| DPPH | 0.62 ** | 0.792 ** | 0.93 ** | 0.967 ** | 0.937 ** | 0.921 ** | 0.967 ** | 0.955 ** | 0.958 ** | 0.951 ** | 0.963 ** | 0.93 ** | 1 | |
| FRAP | 0.681 ** | 0.825 ** | 0.949 ** | 0.975 ** | 0.938 ** | 0.907 ** | 0.965 ** | 0.939 ** | 0.943 ** | 0.938 ** | 0.961 ** | 0.928 ** | 0.971 ** | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.S.; Li, J.; Wang, C.; Monshi, F.I.; Tabassum, R.; Islam, M.A.; Faruquee, M.; Muktadir, M.A.; Mia, M.S.; Islam, A.K.M.M.; et al. Enhanced Antioxidant Activity and Secondary Metabolite Production in Tartary Buckwheat under Polyethylene Glycol (PEG)-Induced Drought Stress during Germination. Agronomy 2024, 14, 619. https://doi.org/10.3390/agronomy14030619
Hossain MS, Li J, Wang C, Monshi FI, Tabassum R, Islam MA, Faruquee M, Muktadir MA, Mia MS, Islam AKMM, et al. Enhanced Antioxidant Activity and Secondary Metabolite Production in Tartary Buckwheat under Polyethylene Glycol (PEG)-Induced Drought Stress during Germination. Agronomy. 2024; 14(3):619. https://doi.org/10.3390/agronomy14030619
Chicago/Turabian StyleHossain, Md. Shakhawat, Jing Li, Chenyang Wang, Fakhrul Islam Monshi, Rehenuma Tabassum, Md Ashraful Islam, Muhiuddin Faruquee, Md. Abdul Muktadir, Md Sultan Mia, A. K. M. Mominul Islam, and et al. 2024. "Enhanced Antioxidant Activity and Secondary Metabolite Production in Tartary Buckwheat under Polyethylene Glycol (PEG)-Induced Drought Stress during Germination" Agronomy 14, no. 3: 619. https://doi.org/10.3390/agronomy14030619