3.5. Structural Equation Modeling of Yield and Agronomic Traits
The relationship between yield and main agronomic traits was analyzed using the structural equation model (
Figure 5). The results showed that the reduction in total root length had a significant positive effect on plant height. The reduction in green leaf area per plant was positively affected by plant height, total root length reduction, and root diameter reduction. The reduction in green leaf area per plant had a significant positive effect on the number of effective branches. The 1000-grain weight and yield were negatively affected by the reduction in total root length, the reduction in root diameter, the reduction in green leaf area per plant, and the number of effective branches.
The results of the cluster analysis on quinoa yield and subsequent analysis of agronomic traits across different yield categories provide valuable insights into the relationship between yield and various plant characteristics. The cluster analysis identified three distinct yield categories based on grain yield: low-yielding (LY), medium-yielding (MY), and high-yielding (HY) varieties. These categories provide a basis for understanding the performance of different quinoa cultivars in terms of yield potential.
Yield components such as effective branch number and 1000-grain weight varied significantly among yield categories. High-yielding varieties demonstrated fewer effective branches but higher 1000-grain weight, suggesting efficient resource allocation towards grain production. This study also found a positive correlation between yield per plant and 1000-grain weight. Previous research has mentioned a positive correlation of yield with seed size, harvest index, and 1000-grain weight [
20,
31,
32]. On the other hand, the number of effective branches at maturity decreased with increasing yield, consistent with the results of Woo et al. [
19].
The harvest index (HI) reflects the ability of crop photosynthetic assimilation products to be converted into economic products. A high harvest index indicates that plants have a mechanism for supplying a higher biomass to seeds. In this study, the harvest index of quinoa was low (8.77–13.15%), but the HI increased with the yield, and the HY varieties depicted a higher HI. This suggests that increasing the HI is an important way to increase the yield of quinoa [
33]. Therefore, increasing the 1000-grain weight, reducing the branch number, and improving the biomass allocation mechanism can be considered important research directions for increasing quinoa yield.
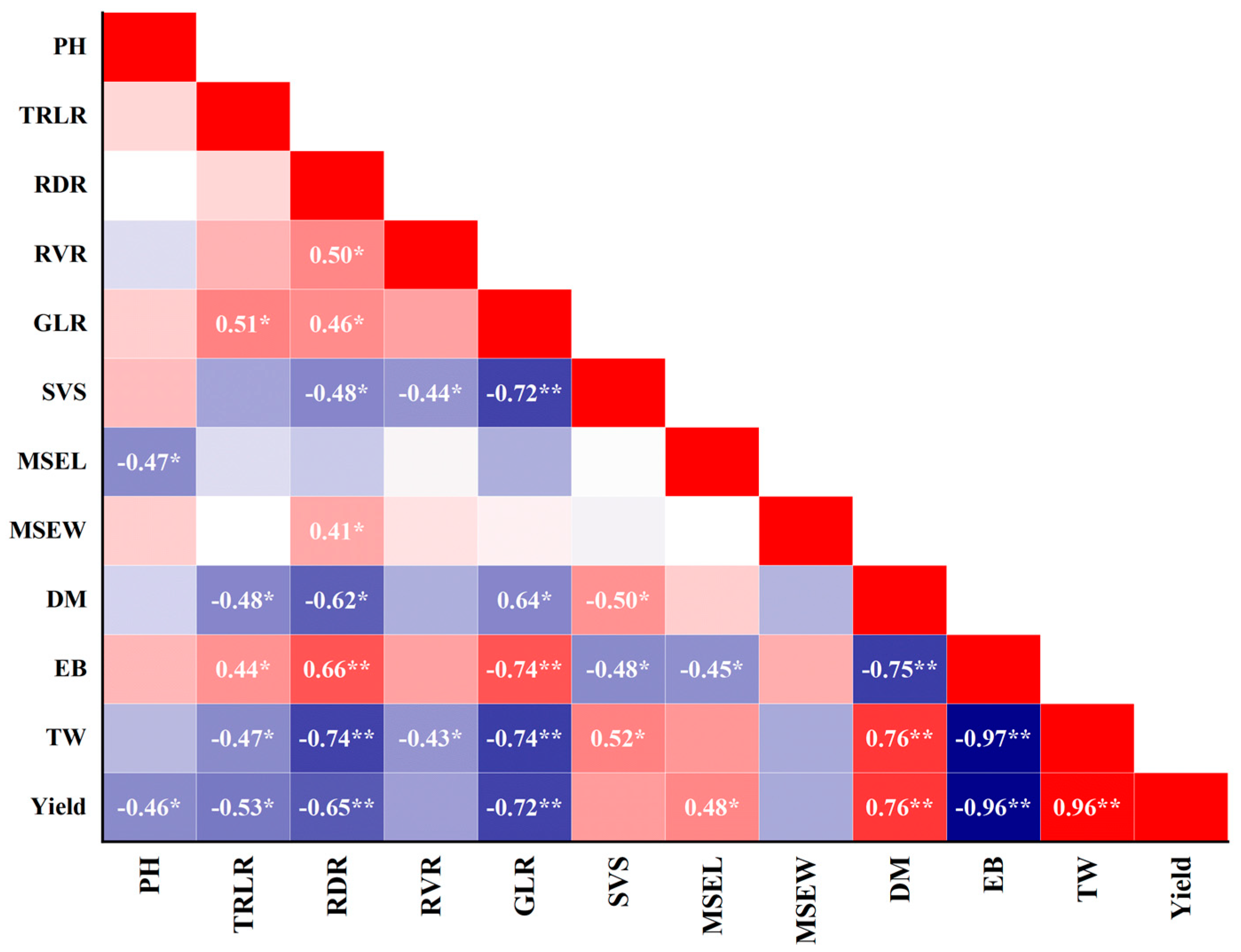
Correlation analysis revealed significant relationships between yield and various agronomic traits. Positive correlations were observed between yield and traits such as dry matter accumulation, stem-breaking force, and 1000-grain weight, while negative correlations were observed with traits like effective branch number. These correlations highlight the importance of specific traits in determining yield potential and suggest potential breeding targets for improving quinoa productivity.
The ear (panicle) length of the main stem was found to be significantly higher in the HY varieties, and this increase was positively correlated with yield. On the other hand, the ear width of the main stem at the maturity stage was significantly lower in the HY varieties, and it gradually decreased with increasing yield. The yield was negatively correlated with the ear width and positively correlated with the ear length of the main stem. These findings align with a previous study by Dumschott et al. [
7]. However, Manjarres-Hernández et al. [
25] reported that quinoa accessions with longer panicle lengths (>57.94 cm) had a lower yield. This discrepancy might be because the maximum average panicle length (43.04 cm) in the present study was lower than the limit referred to by Manjarres-Hernández et al. [
25].
Additionally, the dry matter weight at maturity increased with increasing yield. Considering the research by Qi et al. [
34], it is evident that quinoa with longer main inflorescence and more branches requires more biomass. In this study, it is speculated that as yield increased, the growth resources were continuously transferred to the panicle, causing a delay in stem growth after flowering. Therefore, breeding high-yield quinoa with dwarf, thick stems, longer main spike length, and strong plants can be considered a promising direction.
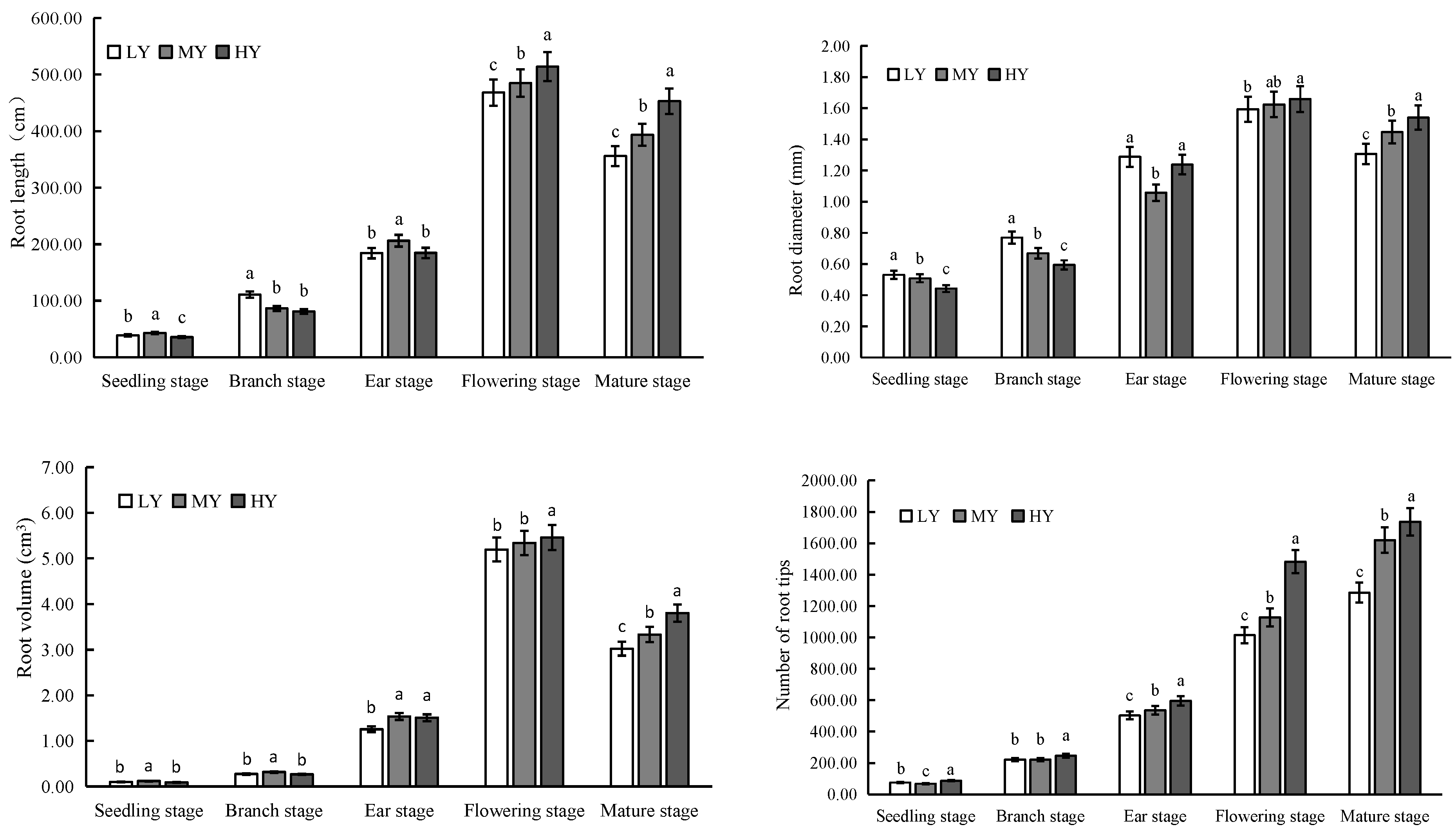
Roots serve as the primary interface between plants and the soil [
35]. The strength of roots can be determined by various factors such as root number, morphology, and physiological indices [
36,
37]. This study found that high-yield varieties exhibited the highest values for total root length, root diameter, root volume, and root tip number during the flowering and maturity stages, which aligns with the findings of Sarropoulou et al. [
38].
In the present study, the root length and diameter increased consistently with the growth stage until the flowering stage and then growth was slowed or stopped. Previous research on rice roots has demonstrated that slowing down the root growth and delaying the physiological activities of roots after flowering enhances leaf photosynthetic efficiency, promotes grain growth, and increases yield [
39,
40]. The decrease in total root length, root diameter, and root volume after flowering was found to be associated with increased yield. Specifically, yield was significantly negatively correlated with the decrease in total diameter, total root length, and root volume after anthesis. These findings are consistent with the study by Ma et al. [
41]. The quinoa varieties that attained higher root length and diameter at the flowering stage and maintained this until the maturity stage showed enhanced grain weight and yield.
Dry matter accumulation per plant increased with the growth period, with high-yielding varieties accumulating the highest dry matter at maturity. This suggests that high-yielding varieties allocate more resources to biomass production, contributing to increased yield potential. Photosynthesis plays a crucial role in the accumulation of dry matter, which directly impacts crop yield. After flowering, enhancing physiological activity and improving photosynthetic performance can delay the senescence of flag leaves, promote the assimilation of synthesis, and increase grain weight [
42]. The findings of this study indicate that high-yield varieties exhibit the highest green leaf area per plant and superior dry matter accumulation during the maturity stage. High-yielding varieties tended to have lower reductions in green leaf area at maturity compared to low- and medium-yielding varieties. Notably, there is a significant negative correlation between yield and the decrease in green leaf area per plant after anthesis. The reduction in green leaf area decrement after anthesis, coupled with an increase in net photosynthetic rate, led to enhanced dry matter accumulation and increased grain yield, aligning with previous research [
43,
44].
The root distribution of quinoa is shallow and the stem is fragile. During the flowering to maturity period, extreme weather conditions can lead to lodging and stem breakage. Variations in plant height were observed across different yield categories and growth stages. Interestingly, the high-yielding varieties exhibited a shorter plant height compared to the low- and medium-yielding varieties at maturity, suggesting potential trade-offs between plant height and yield [
45]. Differences in stem diameter, hardness, and panicle traits were observed among yield categories. High-yielding varieties tended to have thicker stems, higher stem hardness, longer main stem ear length, and more compact panicles, indicating structural characteristics that may contribute to higher yield stability and resistance to lodging [
46]. Additionally, the stem diameter, hardness of the middle part of the stem, and stress value of the stem at maturity all increased with increasing yield, which aligns with the results of previous studies [
47]. The yield of quinoa is positively correlated with the stress value of the stem. The low and thick stem of quinoa enhances lodging resistance, promoting the accumulation of dry matter and increasing grain weight and yield.
The structural equation model (SEM) analysis provided insights into the complex relationships among different agronomic traits and their effects on yield. The 1000-grain weight and yield were negatively affected by the reduction in total root length, the reduction in root diameter, the reduction in green leaf area per plant, and the number of effective branches. These results highlighted the importance of root characteristics, plant height, and green leaf area in influencing yield, underscoring the multifaceted nature of yield determination in quinoa.
Overall, these results contribute to a better understanding of the factors influencing quinoa yield and provide valuable information for breeding programs aimed at developing high-yielding varieties with improved agronomic traits and yield stability.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}