
In-Depth Characterization of Crown Gall Disease of Tobacco in Serbia
by
 , , ,
, , ,
Renata Iličić
1,
Aleksandra Jelušić
2,
Goran Barać
1,
Dušan Nikolić
3,
Nemanja Stošić
4,
Marco Scortichini
5 and 
Tatjana Popović Milovanović
6,* 1
Faculty of Agriculture, University of Novi Sad, Trg Dositeja Obradovića 8, 21000 Novi Sad, Serbia
2
Institute for Multidisciplinary Research, University of Belgrade, Kneza Višeslava 1, 11030 Belgrade, Serbia
3
Institute for Science Application in Agriculture, Bulevar Despota Stefana 68b, 11108 Belgrade, Serbia
4
Academy of Applied Studies, Dobropoljska 5, 15000 Šabac, Serbia
5
Research Centre for Olive, Fruit and Citrus Crops, Council for Agronomical Research and Economics (CREA), Via di Fioranello, 52, I-00134 Roma, Italy
6
Institute for Plant Protection and Environment, Teodora Drajzera 9, 11040 Belgrade, Serbia
*
Author to whom correspondence should be addressed.
Agronomy 2024, 14(4), 851; https://doi.org/10.3390/agronomy14040851
Submission received: 15 March 2024
/
Revised: 12 April 2024
/
Accepted: 15 April 2024
/
Published: 19 April 2024
(This article belongs to the Special Issue Diseases of Herbaceous Plants)
Abstract
:In August 2020, the unusual appearance of crown gall symptoms was observed on the tobacco plants (hybrid PVH2310) grown in fields in the Golubinci (Srem district, Serbia) locality. The causal agent isolated from galls located on tobacco roots formed circular, convex, and glistening light blue colonies, and then dark to olive-green-colored bacterial colonies on a semi-selective D1 medium. Molecular analysis based on multiplex PCR and multi-locus sequence analysis (MLSA) using concatenated sequences of the atpD, dnaK, glnA, and rpoB genes as well as 16S rRNA identified Serbian tobacco isolates such as Agrobacterium tumefaciens (biovar 1). Two duplex PCR methods confirmed the presence of the virD2 and virC genes in tobacco isolates. Pathogenicity tests performed on carrot discs and squash fruits resulted in tumor/gall formation after 12 to 16 days post inoculation, respectively. Pathogenicity was also confirmed on tobacco plants, where isolates caused tumor development 21−25 days after inoculation. API 50 CH generated results regarding the biochemical features of the Serbian tobacco isolates. As A. tumefaciens (biovar 1) as a cause of tobacco crown gall has previously been documented solely in Japan, there is presently no data on its wider occurrence. Therefore, this first detailed investigation of A. tumefaciens isolated from naturally infected tobacco in Serbia will contribute to a better understanding of it at the global level.
1. Introduction
Tobacco (Nicotiana tabacum L.) is considered to have originated from North and South America and has subsequently been introduced for cultivation in other parts of the globe. It is well represented in South America, Mesoamerica and the West Indies, as well as in the Pacific Islands, Asia, and Europe [1]. In 2023, 5.145 ha of land was under tobacco production in Serbia, with an average yield of 1.3 t/ha. Considered an industrial plant in this country, tobacco of the Virginia, Burley, and Oriental type is mostly grown [2].
Plant pathogenic bacteria from the genus Agrobacterium cause crown gall disease, which manifests as tumors and hairy root disease in a variety of crops, including those of high economic importance [3]. In addition to tumor formation, it may lead to symptoms such as dwarfing and the development of small and chlorotic leaves, which ultimately cause plants to wither and die [4].
Agrobacterium tumefaciens, a soil-borne, Gram-negative plant pathogenic bacterium, causes disease symptoms in a variety of plant species by integrating a transfer DNA (T-DNA) from a tumor-inducing plasmid into the host genome [5,6]. The genes required for T-DNA processing and transfer are located in a virulence region with several vir operons (virA-H) on pTi [7]. The A. tumefaciens host range is extensive, and this bacterium has been reported to cause crown gall disease in more than 600 plants belonging to 331 genera [8]. Agrobacterium strains have been divided into three biovars based on their metabolic and growth characteristics: A. tumefaciens (biovar 1-bv1), A. rhizogenes (biovar 2-bv2), and A. vitis (biovar 3-bv3) [5]. Later, these were considered different genera/species in the Rhizobiaceae family, and were named as follows: Agrobacterium species complex (bv1), Rhizobium rhizogenes (bv2), and Allorhizobium vitis (bv3) [9]. At present, the A. tumefaciens species complex has been subdivided into multiple genomospecies with different numerical identifiers [10,11,12,13,14,15,16].
In tobacco, crown gall disease was first found on the roots of crops grown in a field in Kawai-mura, Iwate Prefecture, Japan, in 1995, and A. tumefaciens (bv1) was determined as the causative agent [17]. Since then, the disease has continued to occur in tobacco grown in the same field. On the other hand, the occurrence of this bacterium has not been reported on tobacco in any other country.
In Serbia, symptoms of crown gall disease on tobacco roots were observed in 2020 for the first time. Therefore, the aim of the present study was to determine the genotypic and phenotypic features of Serbian tobacco isolates obtained from infected plants in the field, thereby expanding the current knowledge of the causal agents closely related to Agrobacterium as a tobacco pathogen.
2. Materials and Methods
2.1. Sample Collection and Bacterial Isolation
In 2020, crown gall symptoms were observed on the roots of tobacco plants (hybrid PVH2310, ProfiGen do Brasil Ltd.a., Santa Cruz do Sul—RS, Brazil) grown in the Golubinci (Srem district, Serbia) production fields. Meteorological data were taken from the nearest meteorological station, Sremska Mitrovica, distanced 35 km from the sampling location. The average values of the three-decade air temperatures during the tobacco vegetation period (May–October) were similar to the annual average temperature, with the highest deviation being 1.4 °C in May (Scheme 1a). The total precipitation was lower in July and September, and higher in August and October compared to the perennial average for the observed tobacco vegetation period. Double the perennial average of precipitation was recorded in August (Scheme 1b).
Seven samples, each consisting of one tobacco plant with crown gall symptoms, were collected from the field for analysis. Prior to the bacterial isolation, tumors were rinsed under tap water, dried on filter paper, and treated with a commercial 10% bleach solution for 3−5 min; they were subsequently washed with sterilized distilled water (SDW). After repeating this entire procedure, small fragments taken from the interior of the tumor tissues were macerated in SDW, plated onto semi-selective D1 medium [18] 60 min later and incubated at 26 °C for 10 days. All selected isolates were stored in Luria Bertani (LB) broth containing 20% (v/v) glycerol at a temperature of −20 °C.
2.2. Molecular Characterization
2.2.1. DNA Isolation
Using a genomic DNA isolation kit (DNeasy Plant Mini Kit, Qiagen GmbH, Hilden, Germany), the genomic DNA of the tested Serbian isolates was extracted from cultures grown on Nutrient Agar (NA) for 48 h, following the manufacturer’s instructions. The DNA was resuspended in 100 µL of TE buffer and long-term stored at −20 °C.
2.2.2. Multiplex PCR Detection of Agrobacterium Species/Biovars
As shown in Table 1, the DNA of each bacterial isolate was tested by conducting multiplex PCR, with UF + B1R + B2R + AvR + ArR serving as primers targeting 23S rRNA, in line with the approach described by [19]. PCR was performed using 25 μL of DreamTaq Green master mix (Thermo Scientific, Vilnius, Lithuania), 1 µL of bacterial DNA as a template, and 100 pM of appropriate primer. The amplified PCR products were visualized by gel electrophoresis on 1.5% agarose gel stained with Midori Green Advance under UV light. The fragment sizes were estimated in relation to the GeneRuler 100 bp Plus DNA Ladder (Thermo Scientific™).
2.2.3. Detection of Virulence Genes
Two duplex PCR analyses were carried out using the A/C (virD2) and CYT/CYT (ipt) primers [20] for the first reaction, and PGF/PGR (pehA) [21] and VCF3/VCR3 (virC) [22] primers for the second reaction (Table 1), adopting the amplification conditions outlined by Kuzmanović et al. [23]. The PCR products were separated by electrophoresis using 2% agarose gel, stained with Midori Green Advance. The fragment sizes were assessed by comparing them to the GeneRuler 100 bp DNA Ladder (Thermo Scientific™).
2.2.4. Multi-Locus Sequence Analysis (MLSA)
As shown in Table 1, multi-locus sequence analysis (MLSA) was performed based on the sequences of four housekeeping genes—atpD, dnaK, glnA, and rpoB [24]—as well as with partial sequences of 16S rRNA [25].
The PCR mix (25 µL) consisted of 12.5 µL of Color OptiTaq PCR Master Mix (2×), 9.5 µL of ultrapure DNases/RNase-free water, 1 µL of each primer (10 µM), and 1 µL of DNA. The following protocol was adopted for PCR amplifications: initial denaturation at 95 °C for 10 min (16S rRNA) or 3 min (atpD, dnaK, glnA, and rpoB), followed by 30 (16S rRNA) or 35 (atpD, dnaK, glnA, and rpoB) cycles of denaturation at 94 °C for 1 min, annealing at 55 °C for 90 s (16S rRNA) or at 60 °C (atpD), 65 °C (dnaK and glnA) and 68 °C (rpoB) for 1 min, extension at 72 °C for 150 s (16S rRNA, dnaK, glnA, and rpoB) or 90s (atpD), and a final extension step at 72 °C for 10 min.
The obtained PCR products were sent for sequencing to Eurofins Genomics, located in Hamburg (Germany). Upon receipt, all sequences were manually checked for quality using BioEdit software v.7.2.
The Basic Local Alignment Search Tool (BLASTn) available in the National Center for Biotechnology Information (NCBI) database was used for the preliminary identification of Serbian tobacco isolates based on the sequenced genes. Further, we performed phylogenetic analysis to finally identify and compare Serbian tobacco isolates with 15 different Agrobacterium spp. from the NCBI (Table 2). Concatenated sequences (3452 nt) of each of the five sequenced genes were used to construct a neighbor-joining (NJ) phylogenetic tree. The ClustalW multiple alignment tool from BioEdit v.7.2 was used to align sequences. The phylogenetic tree was generated in MegaX software version 10.0.5 using the Kimura two-parameter model [26]. The Mesorhizobium huakuii strain NZP2235 served as an out-group (Table 2) for rooting the tree. The sequences of the tobacco isolates were deposited in the NCBI database and the accession numbers were obtained.
2.3. Phenotypic Characterization
2.3.1. Pathogenicity
The pathogenicity of the isolates was evaluated on carrot (Daucas carota L.) disc and squash (Cucurbita pepo var. italica L.) fruits. The tested isolates grown on NA at 28 °C for 48 h were suspended in SDW to a final concentration of 107 CFU mL−1. SDW served as a negative control treatment. The carrot and squash fruits were washed, dried, and then disinfected with 70% ethanol before being artificially inoculated in line with the methods described by Ryder et al. [27] and Tolba and Soliman [28], respectively. The inoculated samples were stored in plastic boxes on filter paper at 25 °C for three weeks. Tumor formation was considered a positive reaction.
Selected Serbian tobacco isolates were also tested for pathogenicity to N. tabacum hybrid PVH2310. A bacterial suspension (at a final concentration of 108 CFU mL−1) was prepared from cultures grown on NA at 28 °C for 48 h. The stems of 1-month-old tobacco plants grown in an air-conditioned greenhouse at a constant temperature of 28 °C were inoculated by the needle prick method. Three plants were inoculated with each strain, and the experiment was repeated twice. Upon symptom development, pathogenicity was evaluated based on tumor formation at the inoculation site [17]. Reisolations were performed on D1 medium as soon as symptoms were observed, and the congruence between the reisolates and the original isolates was assessed by multiplex PCR, according to the protocol described by Puławska et al. [6].
2.3.2. Biochemical Tests
The biochemical characterization of the Serbian tobacco isolates was performed according to conventional bacteriological methods described by Schaad et al. [29]. Additional tests were performed using the API 50 CH (bioMérieux, France), under the conditions indicated by the manufacturer, and the results were read after 72 to 96 h of incubation at 26 °C. The tested bacterial isolates were suspended in API 50 CHB/E medium (bioMérieux, Craponne, France), following the manufacturer’s instructions.
3. Results
3.1. Symptoms and Bacterial Isolation
In August 2020, the appearance of crown gall symptoms was observed on the tobacco plants grown at the Golubinci production site. The diseased plants exhibited a lack of vigor, stunted growth, and a reduction in foliage. On the root system, the crown gall symptoms included round, white and soft tumors of different sizes (Figure 1a). In all cases, typical Agrobacterium circular, convex, glistening colonies with a characteristic light blue, and then dark to olive green color, were formed on D1 medium after 10 days of incubation (Figure 1b). Seven pure isolates (coded as DA5, DA17, DA21, DA34, DA40, DA52, and DA65) were selected for further testing.
3.2. Molecular Characterization
3.2.1. Multiplex PCR Detection of Agrobacterium Species/Biovars
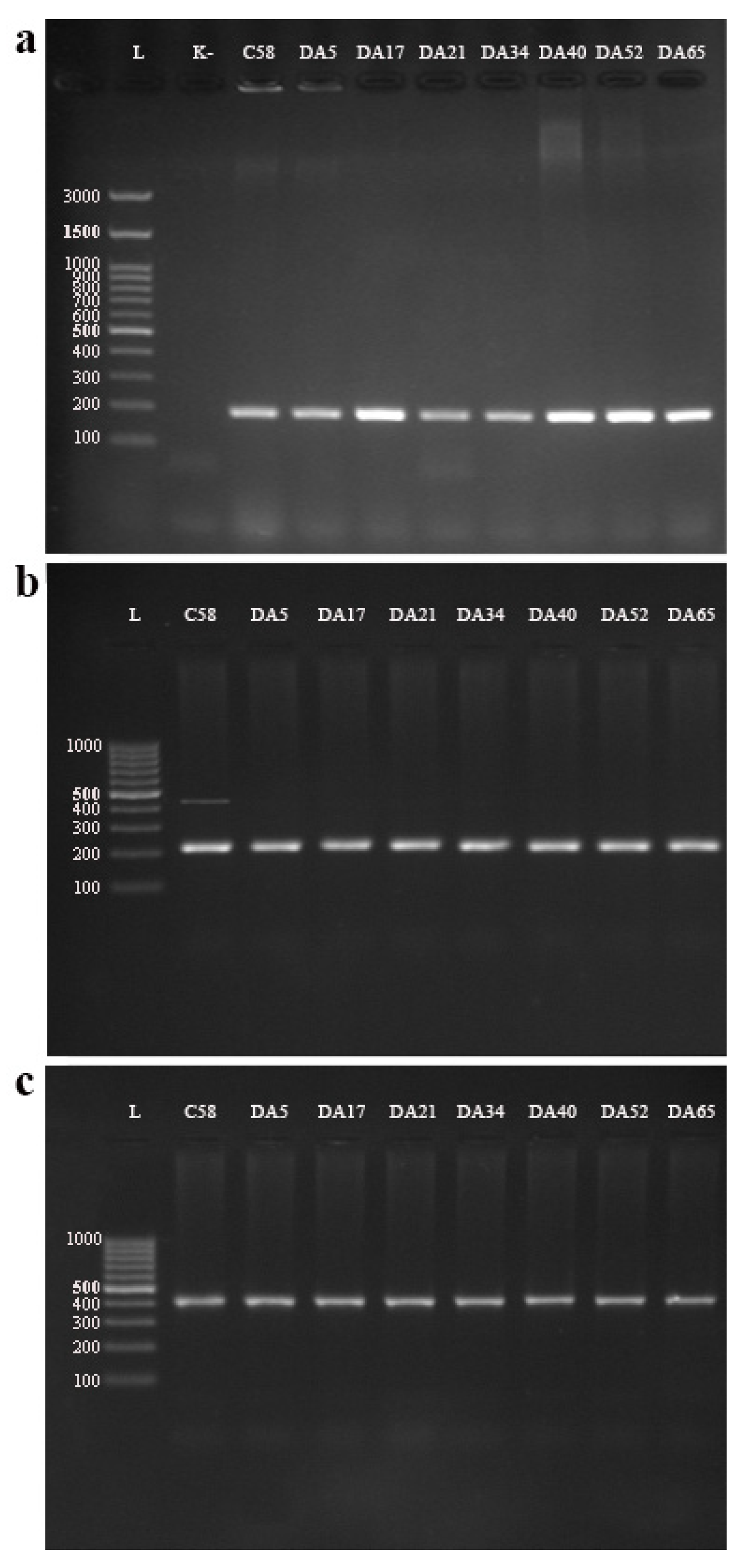
In a multiplex PCR (using primers UF + B1R + B2R + AvR + ArR) assay targeting 23S rRNA gene sequences, a 184 bp fragment specific for A. tumefaciens (bv1) was amplified in all seven studied tobacco isolates. The C58 control strain yielded amplification products at the expected position of 184 bp (Figure 2a).
3.2.2. Detection of Virulence Genes
Among the four primer pairs (A/C and CYT/CYT; PGF/PGR and VCF3/VCR3) used in the pathogenicity evaluation, A/C’ and VCF3/VCR3 successfully amplified the fragment of the expected length (224 bp and 414 bp, respectively) in the tobacco isolates (Figure 2b,c). At the same time, a positive PCR reaction in the reference strain (C58) was observed for the primer pairs A/C’, CYT/CYT’, and VCF3/VCR3. Using the PGF/PGR primer pair, the gene for polygalacturonase (pehA) was not detected in any of the tobacco isolates, along with reference strain C58, since it is characteristic of A. vitis.
3.2.3. Multi-locus Sequence Analysis (MLSA)
According to the BLASTn analysis, seven Serbian tobacco isolates were preliminarily identified as A. tumefaciens, exhibiting the highest homology (99.84–100%) with A. tumefaciens strains originating from different hosts/countries (CFBP 5621, HAMBI 105, Gle002, Yol001, Yol002) for the atpD, dnaK, glnA and rpoB genes. According to the BLASTn analysis of 16S rRNA sequences, the tobacco isolates showed the highest homology (100%) with the A. tumefaciens strains obtained from the NCBI database (CFBP6623, CFBP6624, Yol001, Yol002, Gle002, HAMBI 105, 12D1, AR2, 186, 0176, 1D1460, 183, 175-HR29, etc.).
According to the constructed neighbor-joining phylogenetic tree shown in Figure 3, the seven Serbian tobacco isolates (DA5, DA17, DA21, DA34, DA40, DA52, and DA65) formed a homogeneous group and clustered with the HAMBI 105, Gle002, Yol001, and Yol002 reference A. tumefaciens strains. The remaining four comparative A. tumefaciens strains (BIM B-1315G, 12D1, 183, and O54/95) formed a separate subcluster within the same cluster. Other comparative strains from the NCBI database (Table 2) were placed separately within different clusters, each corresponding to different Agrobacterium species (i.e., A. pusense, A. leguminum, A. fabrum, A. vaccinii, and A. larrymoorei). An outgroup strain, R. rhizogenes LBA9402, was clearly separated on a monophyletic tree branch. As all tested tobacco isolates were shown to be genetically homogenous, only one (DA5) was randomly selected and deposited into the NCBI database under the accession numbers PP471554 (atpD), PP471555 (dnaK), PP471556 (glnA), PP471557 (rpoB), and PP464228 (16S rRNA).
3.3. Phenotypic Characterization of Tobacco Isolates
3.3.1. Pathogenicity
In the tumorigenicity tests, all tobacco isolates, along with reference strain C58, on the carrot discs and squash fruits were positive 12 to 16 days after inoculation. Positive responses were indicated by the formation of differently sized and shaped tumors/galls on the inoculated carrot discs (Figure 4a), as well as on the squash fruit (Figure 4b). Negative controls were symptomless.
In the pathogenicity assay involving tobacco plants, symptoms associated with crown gall disease emerged 21 to 25 days after inoculation, as indicated by the formation of galls with different sizes at the inoculation site (Figure 4c). No symptoms were observed on the plants that were inoculated with SDW. To fully adhere to Koch’s postulates, the pathogen was reisolated from young tumor tissue on D1 medium and was identified by the amplification of a 184 bp fragment via multiplex PCR.
3.3.2. Biochemical Tests
The biochemical test results showed that the isolates were Gram negative, catalase positive, and oxidase negative, and did not produce fluorescent pigment on King’s B medium. In the API test (API 50 CH), the Agrobacterium tobacco isolates were positive for the fermentation of glycerol, d-arabinose, l-arabinose, d-ribose, d-xylose, d-adonitol, methyl-β-d-xylopyranoside, d-galactose, d-glucose, d-fructose, d-mannose, l-rhamnose, dulcitol, inositol, d-mannitol, d-sorbitol, aesculin ferric citrate, salicin, d-cellobiose, d-maltose, d-lactose, d-melibiose, d-sucrose, d-trehalose, d-raffinose, d-turanose, d-lyxose, d-tagatose, d-fucose, l-fucose, d-arabitol, and l-arabitol, while being negative for erythritol, l-xylose, l-sorbose, methyl α-d-mannopyranoside, methyl α-d-glucopyranoside, N-acetylglucosamine, amygdalin, arbutin, inulin, d-melezitose, amidon (starch), glycogen, xylitol, gentiobiose, potassium gluconate, potassium 2-ketogluconate, and potassium 5-ketogluconate. The API test results for strain C58 differed only in glycerol (−), methyl-β-d-xylopyranoside (−), d-galactose (−), l-sorbose (+), d-fucose (−), l-fucose (−) and d-arabitol (−) fermentation (Table 3).
4. Discussion
The Gram-negative soil bacterium A. tumefaciens is widely distributed globally and is known to induce crown gall disease on various plants, including fruit trees as well as ornamental and herbaceous plants across different regions [3,13,30,31]. The present study marks the first attempt to examine A. tumefaciens as the causative agent of tobacco crown gall in Serbia. Thus far, this bacterial pathogen in tobacco fields has only been observed in Japan, with the first report dating back to 1995 [17]. Thus, by offering a more comprehensive understanding of tobacco isolates through genotyping, the present study fills an important gap in the extant knowledge regarding this pathogen’s interaction with tobacco. Although the relationship between tobacco plants and the bacterium A. tumefaciens is widely recognized and utilized as a model system for studying pathogenicity and virulence under controlled conditions, natural infections of tobacco by this bacterium have not garnered sufficient scholarly attention.
Based on the phylogenetic analysis performed as a part of the present study, the Serbian tobacco isolates exhibited the closest association with A. tumefaciens strains originating from soil and walnut trees in the USA. This finding may indicate a common evolutionary history, a pathway for dissemination, or adaptation across different geographical regions. Genetic diversity was not observed among the A. tumefaciens tobacco isolates from Serbia based on the genes used for sequencing. In the research conducted by Furuya et al. [17], tobacco isolates were solely identified through PCR, utilizing the universal primers VCF/VCR for detecting pathogenic Agrobacterium spp. It is noteworthy that there are no publicly accessible genomic data on isolates from tobacco when studied as an A. tumefaciens host; therefore, this study provides pioneering genomic information regarding A. tumefaciens strains originating from tobacco.
According to Aujoulat et al. [24], the MLSA scheme, using seven housekeeping genes encoding proteins involved in transcription (rpoB), the stress response (dnaK, groEL), amino acid biosynthesis (glnA, trpE), and energy metabolism (atpD, zwf), is suitable for analyzing strains belonging to the three biovars in the Agrobacterium genus—bv1, bv2, and A. vitis (bv3). In this study, we provided sequences from 16S rRNA with gene sets used in MLSA (rpoB, dnaK, glnA, atpD). Considering that the phylogenetic analysis clearly clustered A. tumefaciens in a clade separated from other Agrobacterium spp., the MLSA scheme adopted in our work displayed good discriminatory power for Agrobacterium phylogeny. On the other hand, taxon-specific sets of genes for the MLSA of taxa with gall-causing bacteria were identified by consulting the pertinent literature [11,32,33,34,35].
Furthermore, using a set of specific primers [19], the tobacco isolates were identified at the species/biovar level (A. tumefaciens, bv1), indicating that these tobacco isolates were members of this species. Two duplex PCR methods conducted with four different primer pairs confirmed the presence of the plasmid virD2 and virC pathogenicity gene, indicating the presence of Ti or Ri plasmids in the tested isolates [16,22,23]. Kuzmanović et al. [36] stated that VCF3/VCR3 primers (targeting virC1-virC2 gene) are applicable in a wide variety of tumorigenic agrobacteria and are therefore considered a reliable tool for detecting crown gall pathogens. The VirD2 gene plays an important role in T-DNA processing and transfer, which is important for virulence [37].
Serbian tobacco isolates displayed phenotypic characteristics consistent with those of A. tumefaciens [17]. All isolates induced tumors on carrot discs, squash fruits, and young tobacco plants following artificial inoculation. Likewise, in the study conducted by Furuya et al. [17], all tested Nicotiana species were susceptible to the A. tumefaciens isolated from naturally infected tobacco plants. Based on their biochemical characteristics, the Serbian tobacco isolates most closely resembled A. tumefaciens, but the biovar phenotype features differed in some cases [29]. Strains with intermediate/atypical biochemical characters have been frequently reported, suggesting that more accurate additional tests are needed for precise phenotypic identification [38,39]. Regarding the results yielded by pathogenicity and biochemical tests, the molecular phylogenies of housekeeping genes and the determined presence of pathogenicity genes, the Serbian tobacco isolates were identified as A. tumefaciens.
The ease of Agrobacterium spread through tobacco transplants presents a latent threat not only to tobacco crops, but also to overall agricultural production. Due to the difficulty of controlling crown gall disease, this study highlights the importance of early and accurate detection for mitigating crop damage.
5. Conclusions
In conclusion, the results obtained in the present study provide novel insight into the taxonomic position of Agrobacterium isolates associated with tobacco, enhancing the current understanding of the Agrobacterium population in Serbia. To reduce the risk of significant crop losses resulting from crown gall infection, rigorous surveys are advised, especially in regions experiencing frequent outbreaks of economic significance (in this particular case, crucial tobacco plantations grown in monoculture).
Author Contributions
Conceptualization, R.I., M.S. and T.P.M.; methodology, R.I., A.J., G.B. and T.P.M.; software, A.J. and N.S.; validation, R.I., A.J., G.B. and T.P.M.; formal analysis, R.I., A.J., G.B. and T.P.M.; investigation, R.I., A.J., G.B. and T.P.M.; resources, R.I., D.N. and T.P.M.; data curation, R.I., A.J., G.B. and T.P.M.; writing—original draft preparation, R.I., A.J., G.B., D.N., N.S., M.S. and T.P.M.; writing—review and editing, R.I., A.J., G.B., M.S. and T.P.M.; visualization, R.I., A.J., G.B., D.N., N.S., M.S. and T.P.M.; supervision, R.I., A.J., M.S. and T.P.M.; funding acquisition, R.I., D.N. and T.P.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Ministry of Science, Technological Development and Innovation of the Republic of Serbia [contract numbers 451-03-66/2024-03/200117, 451-03-66/2024-03/200053, 451-03-66/2024-03/200010].
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- CABI. Nicotiana tabacum (tobacco). 2014. Available online: https://www.cabidigitallibrary.org/doi/10.1079/cabicompendium.36326 (accessed on 10 December 2023).
- SYRS. Statistical Yearbook of the Republic of Serbia; Statistical Office of the Republic of Serbia: Belgrade, Serbia, 2023.
- Hooykaas, M.J.G.; Hooykaas, P.J.J. Complete genomic sequence and phylogenomics analysis of Agrobacterium strain AB2/73: A new Rhizobium species with a unique mega-Ti plasmid. BMC Microbiol. 2021, 21, 295. [Google Scholar] [CrossRef] [PubMed]
- Tekiner, N.; Kotan, R. Pathogenicity of Different Rhizobium radiobacter (Agrobacterium tumefaciens) Isolates and Their Diagnosis with Classical Methods. Kahramanmaraş Sütçü İmam Üniversitesi Tarım ve Doğa Derg. 2022, 25, 149–157. [Google Scholar] [CrossRef]
- Portier, P.; Fischer-Le Saux, M.; Mougel, C.; Lerondelle, C.; Chapulliot, D.; Thioulouse, J.; Nesme, X. Identification of genomic species in Agrobacterium biovar 1 by AFLP genomic markers. Appl. Environ. Microbiol. 2006, 72, 7123–7131. [Google Scholar] [CrossRef]
- Tiwari, M.; Mishra, A.K.; Chakrabarty, D. Agrobacterium-mediated gene transfer: Recent advancements and layered immunity in plants. Planta 2022, 256, 37. [Google Scholar] [CrossRef]
- Nabi, N.; Hafsa, A.B.; Zellama, M.S.; Chaouachi, M. Pathogenicity, Phylogenetic relationship and NGS based identifcation and assembly of tumorigenic Agrobacterium radiabacter plasmidic and chromosomic reads isolated from Prunus duclcis. Genomics 2019, 111, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Vijayanand, K.G.; Reddy, M.P.; Singh, A.S.; Naraynan, S. Agrobacterium tumefaciens-Mediated Genetic Transformation: Mechanism and Factors. J. For. Environ. Sci. 2009, 25, 195–204. [Google Scholar]
- Hooykaas, P.J. The Ti plasmid, driver of Agrobacterium pathogenesis. Phytopathology 2023, 113, 594–604. [Google Scholar] [CrossRef]
- Mougel, C.; Thioulouse, J.; Perrie‘re, G.; Nesme, X. A mathematical method for determining genome divergence and species delineation using AFLP. Int. J. Syst. Evol. Microbiol. 2002, 52, 573–586. [Google Scholar] [CrossRef]
- Costechareyre, D.; Bertolla, F.; Nesme, X. Homologous recombination in Agrobacterium: Potential implications for the genomic species concept in bacteria. Mol. Biol. Evol. 2009, 26, 167–176. [Google Scholar] [CrossRef]
- Lassalle, F.; Campillo, T.; Vial, L.; Baude, J.; Costechareyre, D.; Chapulliot, D.; Shams, M.; Abrouk, D.; Lavirre, C.; Oger-Desfeux, C.; et al. Genomic species are ecological species as revealed by comparative genomics in Agrobacterium tumefaciens. Genome Biol. Evol. 2011, 3, 762–781. [Google Scholar] [CrossRef]
- Mafakheri, H.; Taghavi, S.M.; Puławska, J.; De Lajudie, P.; Lassalle, F.; Osdaghi, E. Two novel genomospecies in the Agrobacterium tumefaciens species complex associated with rose crown gall. Phytopathology 2019, 109, 1859–1868. [Google Scholar] [CrossRef] [PubMed]
- Flores-Félix, J.D.; Menéndez, E.; Peix, A.; García-Fraile, P.; Velázquez, E. History and current taxonomic status of genus Agrobacterium. Syst. Appl. Microbiol. 2020, 43, 126046. [Google Scholar] [CrossRef]
- Singh, N.K.; Lavire, C.; Nesme, J.; Vial, L.; Nesme, X.; Mason, C.E.; Lassalle, F.; Venkateswaran, K. Comparative Genomics of Novel Agrobacterium G3 Strains Isolated From the International Space Station and Description of Agrobacterium tomkonis sp. nov. Front. Microbiol. 2021, 12, 765943. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, A.J.; Wu, Y.; Chang, J.H.; Lai, E.M.; Kuo, C.H. Virulence and Ecology of Agrobacteria in the Context of Evolutionary Genomics. Annu. Rev. Phytopathol. 2023, 61, 1–23. [Google Scholar] [CrossRef]
- Furuya, N.; Shimokusuzono, F.; Nakamura, Y.; Nishimura, K.; Takeshita, M.; Matsuyama, N.; Manabe, K.; Takanami, Y. Crown gall of tobacco caused by Agrobacterium tumefaciens biovar 1 in tobacco fields. J. Gen. Pant Pathol. 2004, 70, 39–44. [Google Scholar] [CrossRef]
- Kado, C.I.; Heskett, M.G. Selective media for isolation of Agrobacterium, Corynebacterium, Erwinia, Pseudomonas and Xanthomonas. Phytopathology 1970, 60, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Puławska, J.; Willems, A.; Sobiczewski, P. Rapid and specific identification of four Agrobacterium species and biovars using multiplex PCR. Syst. Appl. Microbiol. 2006, 29, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Haas, J.H.; Moore, L.W.; Ream, W. Universal PCR primers for detection of phytopathogenic Agrobacterium strains. Appl. Environ. Microbiol. 1995, 61, 2879–2884. [Google Scholar] [CrossRef] [PubMed]
- Szegedi, E.; Bottka, S. Detection of Agrobacterium vitis by polymerase chain reaction in grapevine bleeding sap after isolation on a semiselective medium. Vitis 2002, 41, 37–42. [Google Scholar]
- Suzaki, K.; Yoshida, K.; Sawada, H. Detection of tumorigenic Agrobacterium strains from infected apple saplings by colony PCR with improved PCR primers. J. Gen. Plant Pathol. 2004, 70, 342–347. [Google Scholar] [CrossRef]
- Kuzmanović, N.; Ivanović, M.; Ćalić, A.; Gašić, K.; Obradović, A. Differentiation of Phytopathogenic agrobacterium spp. Pestic. Phytomed. 2011, 26, 245–253. [Google Scholar] [CrossRef]
- Aujoulat, F.; Jumas-Bilak, E.; Masnou, A.; Sallé, F.; Faure, D.; Segonds, C.; Marchandin, H.; Teyssier, C. Multilocus sequence-based analysis delineates a clonal population of Agrobacterium (Rhizobium) radiobacter (Agrobacterium tumefaciens) of human origin. J. Bacteriol. 2011, 193, 2608–2618. [Google Scholar] [CrossRef] [PubMed]
- Caccamo, D.; Di Cello, F.; Fani, R.; Gugliandolo, C.; Maugeri, T.L. Polyphasic approach to the characterisation of marine luminous bacteria. Res. Microbiol. 1999, 150, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Ryder, M.H.; Tate, M.E.; Kerr, A. Virulence Properties of Strains of Agrobacterium on the Apical and Basal Surfaces of Carrot Root Discs. Plant Physiol. 1985, 77, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Tolba, I.H.; Soliman, M.A. Efficacy of Native Antagonistic Bacterial Isolates in Biological Control of Crown Gall Disease in Egypt. Ann. Agric. Sci. 2013, 58, 43–49. [Google Scholar] [CrossRef]
- Schaad, N.W.; Jones, J.B.; Chun, W. Laboratory Guide for Identification of Plant Pathogenic Bacteria; The American Phytopathological Society: St. Paul, MN, USA, 2001. [Google Scholar]
- Puławska, J. Crown gall of stone fruits and nuts, economic significance and diversity of its causal agents: Tumorigenic Agrobacterium spp. J. Plant Pathol. 2010, 92, S87–S98. [Google Scholar]
- Holmes, J.E.; Sanghera, H.; Punja, Z.K. Crown gall development on cannabis (Cannabis sativa L., marijuana) plants caused by Agrobacterium tumefaciens species-complex. Can. J. Plant Pathol. 2023, 45, 433–445. [Google Scholar] [CrossRef]
- Alexandre, A.; Laranjo, M.; Young, J.P.; Oliveira, S. dnaJ is a useful phylogenetic marker for Alphaproteobacteria. Int. J. Syst. Evol. Microbiol. 2008, 58, 2839–2849. [Google Scholar] [CrossRef] [PubMed]
- Puławska, J.; Kałuzna, M. Phylogenetic relationship and genetic diversity of Agrobacterium spp. isolated in Poland based on gyrB gene sequence analysis and RAPD. Eur. J. Plant Pathol. 2012, 133, 379–390. [Google Scholar] [CrossRef]
- Pérez-Yépez, J.; Armas-Capote, N.; Velázquez, E.; Pérez-Galdona, R.; Rivas, R.; León-Barrios, M. Evaluation of seven housekeeping genes for multilocus sequence analysis of the genus Mesorhizobium: Resolving the taxonomic affiliation of the Cicer canariense rhizobia. Syst. Appl. Microbiol. 2014, 37, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Kuzmanović, N.; Puławska, J.; Smalla, K.; Nesme, X. Agrobacterium rosae sp. nov., isolated from galls on different agricultural crops. Syst. Appl. Microbiol. 2018, 41, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Kuzmanović, N.; Behrens, P.; Idczak, E.; Wagner, S.; Götz, M.; Spröer, C.; Smalla, K. A novel group of Rhizobium tumorigenes-like agrobacteria associated with crown gall disease of rhododendron and blueberry. Phytopathology 2019, 109, 1840–1848. [Google Scholar] [CrossRef] [PubMed]
- Mysore, K.S.; Bassuner, B.; Deng, X.B.; Darbinian, N.S.; Motchoulski, A.; Ream, W.; Gelvin, S.B. Role of the Agrobacterium tumefaciens VirD2 protein in T-DNA transfer and integration. Mol. Plant Microbe Interact. 1998, 11, 668–683. [Google Scholar] [CrossRef] [PubMed]
- Zoina, A.; Raio, A.; Peluso, R.; Spasiano, A. Characterisation of agrobacteria from weeping fig (Ficus benjamina). Plant Pathol. 2001, 50, 620–627. [Google Scholar] [CrossRef]
- Peluso, R.; Raio, A.; Morra, F.; Zoina, A. Physiological, biochemical and molecular analyses of an Italian collection of Agrobacterium tumefaciens strains. Eur. J. Plant Pathol. 2003, 109, 291–300. [Google Scholar] [CrossRef]
Scheme 1.
Meteorological data for sampling site: (a) Average monthly temperatures recorded for the period 1991–2020 and average monthly temperature in 2020; (b) Average monthly sum of precipitation recorded for the period 1991–2020 and the average sum of monthly precipitation in 2020.
Scheme 1.
Meteorological data for sampling site: (a) Average monthly temperatures recorded for the period 1991–2020 and average monthly temperature in 2020; (b) Average monthly sum of precipitation recorded for the period 1991–2020 and the average sum of monthly precipitation in 2020.

Figure 1.
A. tumefaciens. (a) Symptoms of naturally infected tobacco plants; (b) colonies on D1 medium 10 days after incubation.
Figure 1.
A. tumefaciens. (a) Symptoms of naturally infected tobacco plants; (b) colonies on D1 medium 10 days after incubation.

Figure 2.
(a) Multiplex-PCR detection of Agrobacterium species/biovars using primers UF + B1R + B2R + AvR + ArR; (b) PCR detection of virulence genes virD2 and ipt, using primers A/C and CYT/CYT, respectively; (c) PCR detection of virulence genes virC and pehA, using primers VCF3/VCR3 and PGF/PGR, respectively. Letter “L” represents DNA ladder—(a) GeneRuler 100 bp Plus DNA Ladder (Thermo Scientific™) and (b,c) GeneRuler 100 bp DNA Ladder (Thermo Scientific™).
Figure 2.
(a) Multiplex-PCR detection of Agrobacterium species/biovars using primers UF + B1R + B2R + AvR + ArR; (b) PCR detection of virulence genes virD2 and ipt, using primers A/C and CYT/CYT, respectively; (c) PCR detection of virulence genes virC and pehA, using primers VCF3/VCR3 and PGF/PGR, respectively. Letter “L” represents DNA ladder—(a) GeneRuler 100 bp Plus DNA Ladder (Thermo Scientific™) and (b,c) GeneRuler 100 bp DNA Ladder (Thermo Scientific™).

Figure 3.
Neighbor-joining phylogenetic tree based on the concatenated sequences of 16S rRNA and the housekeeping genes atpD, dnaK, glnA, and rpoB for the seven Serbian tobacco isolates obtained in this study (marked with red triangles) and the 15 strains of different Agrobacterium spp. sourced from the NCBI database. The tree was rooted with M. huakuii NZP2235 from the NCBI.
Figure 3.
Neighbor-joining phylogenetic tree based on the concatenated sequences of 16S rRNA and the housekeeping genes atpD, dnaK, glnA, and rpoB for the seven Serbian tobacco isolates obtained in this study (marked with red triangles) and the 15 strains of different Agrobacterium spp. sourced from the NCBI database. The tree was rooted with M. huakuii NZP2235 from the NCBI.

Figure 4.
A. tumefaciens. Pathogenicity of isolate DA21 on: (a) carrot discs; (b) squash fruit; (c) tobacco plant.
Figure 4.
A. tumefaciens. Pathogenicity of isolate DA21 on: (a) carrot discs; (b) squash fruit; (c) tobacco plant.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used in this study.
| Primer | Primer Sequence | Region | Fragment Length (bp) |
|---|---|---|---|
| UF/ B1R | 5′-GTAAGAAGCGAACGCAGGGAACT-3′ 5′-GACAATGACTGTTCTACGCGTAA-3′ | Chromosomal (23S rRNA) gene, A. tumefaciens/biovar 1 | 184 |
| UF/ B2R | 5′-TCCGATACCTCCAGGGCCCCTCACA-3′ | Chromosomal (23S rRNA) gene, A. rhizogenes/biovar 2 | 1066 |
| UF/ AvR | 5′-AACTAACTCAATCGCGCTATTAAC-3′ | Chromosomal (23S rRNA) gene, A. vitis | 478 |
| UF/ ArR | 5′-AAAACAGCCACTACGACTGTCTT-3′ | Chromosomal (23S rRNA) gene, A. rubi | 1006 |
| A/ C | 5′-ATGCCCGATCGAGCTCAAGT-3′ 5′-TCGTCTGGCTGACTTTCGTCATAA-3′ | Ti and Ri plasmid virD2 gene | 224 |
| CYT/ CYT | 5′-GATCG(G/C)GTCCAATG(C/T)TGT-3′ 5′-GATATCCATCGATC(T/C)CTT-3′ | Ti plasmid ipt gene | 427 |
| VCF3/ VCR3 | 5′-GGCGGGCGYGCYGAAAGRAARACYT-3′ 5′-AAGAACGYGGNATGTTGCATCTYAC-3′ | Ti and Ri plasmid virC gene | 414 |
| PGF/ PGR | 5′-GGGGCAGGATGCGTTTTTGAG-3′ 5′-GACGGCACTGGGGCTAAGGAT-3′ | Chromosomal pehA gene, A. vitis | 466 |
| atpD (800F/ 1350R) | 5′-GGCCAGGACGTTCTGTTCTT-3′ 5′-CTTGAAGCCCTTGATCGTGT-3′ | F0-F1 ATP synthase subunit beta | 465 |
| dnaK (720F/ 1400R) | 5′-GAAGACTTCGACATGCGTCT-3′ 5′-GCCGAGCAGCTTGTTGTC-3′ | Heat shock protein, 70 kDa | 480 |
| glnA (144F/ 900R) | 5′-GTCATGTTCGACGGCTCCT-3′ 5′-CCTTGGCATGCTTGATGAT-3′ | Glutamine synthetase | 474 |
| rpoB (2040F/ 2718R) | 5′-GAAAACGACGACGCCAAC-3′ 5′-GCGCAGAAGCTTTTCTTCC-3′ | Beta subunit RNA polymerase | 534 |
| 27F 1495R | 5′-GAGAGTTTGATCCTGGCTCAG-3′ 5′-CTACGGCTACCTTGTTACGA-3′ | 16S rRNA | 1550 |
Table 2.
List of strains from the NCBI database used for phylogenetic analysis.
| Species | Strain | Isolation Source | Country | Acc. No. |
|---|---|---|---|---|
| A. tumefaciens | HAMBI 105 | soil | USA | CP139997 |
| Gle002 | walnut | USA: California | CP048564 | |
| Yol001 | walnut | USA: California | CP048477 | |
| Yol002 | walnut | USA: California | CP048473 | |
| 12D1 | - | - | CP033031 | |
| 183 | almond | Tunisia | CP029044 | |
| O54/95 | cherry | - | CP124967 | |
| BIM B-1315G | root endosphere of soybean | Belarus: Minsk | CP061003 | |
| A. larrymoorei | AF3.44 | Ficus benjamina | USA: Florida | CP072167 |
| CFBP5477 | - | Italy | CP124733 | |
| A. leguminum | CFBP4996 | - | - | CP120211 |
| A. fabrum | C58 | USA | AE007869 | |
| 1D132 | - | CP033022 | ||
| A. vaccinii | B7.6 | blueberry | Poland | CP054150 |
| A. pusense | 76 | hyphae Fusarium oxysporum f. sp. cucumerinum | China: Beijing | CP053856 |
| M. huakuii a | NZP2235 | Lotus japonicus | New Zealand | CP139858 |
a Outgroup strain.
Table 3.
Biochemical characteristics of the Serbian tobacco isolates.
| Isolate | ||||||||
|---|---|---|---|---|---|---|---|---|
| Test | DA5 | DA17 | DA21 | DA34 | DA40 | DA52 | DA65 | C58 |
| Gram reaction | − | − | − | − | − | − | − | − |
| Catalase | + | + | + | + | + | + | + | + |
| Oxidase | − | − | − | − | − | − | − | + |
| Production of fluorescent pigment | − | − | − | − | − | − | − | − |
| Fermentation of: | ||||||||
| glycerol | + | + | + | + | + | + | + | − |
| d-arabinose | + | + | + | + | + | + | + | + |
| l-arabinose | + | + | + | + | + | + | + | + |
| d-ribose | + | + | + | + | + | + | + | + |
| d-xylose | + | + | + | + | + | + | + | + |
| d-adonitol | + | + | + | + | + | + | + | + |
| methyl-β-d-xylopyranoside | + | + | + | + | + | + | + | − |
| d-galactose | + | + | + | + | + | + | + | − |
| d-glucose | + | + | + | + | + | + | + | + |
| d-fructose | + | + | + | + | + | + | + | + |
| d-mannose | + | + | + | + | + | + | + | + |
| l-rhamnose | + | + | + | + | + | + | + | + |
| dulcitol | + | + | + | + | + | + | + | + |
| inositol | + | + | + | + | + | + | + | + |
| d-mannitol | + | + | + | + | + | + | + | + |
| d-sorbitol | + | + | + | + | + | + | + | + |
| aesculin ferric citrate | + | + | + | + | + | + | + | + |
| salicin | + | + | + | + | + | + | + | + |
| d-cellobiose | + | + | + | + | + | + | + | + |
| d-maltose | + | + | + | + | + | + | + | + |
| d-lactose | + | + | + | + | + | + | + | + |
| d-melibiose | + | + | + | + | + | + | + | + |
| d-sucrose | + | + | + | + | + | + | + | + |
| d-trehalose | + | + | + | + | + | + | + | + |
| d-raffinose | + | + | + | + | + | + | + | + |
| d-turanose | + | + | + | + | + | + | + | + |
| d-lyxose | + | + | + | + | + | + | + | + |
| d-tagatose | + | + | + | + | + | + | + | + |
| d-fucose | + | + | + | + | + | + | + | − |
| l-fucose | + | + | + | + | + | + | + | − |
| d-arabitol | + | + | + | + | + | + | + | − |
| l-arabitol | + | + | + | + | + | + | + | + |
| erythritol | − | − | − | − | − | − | − | − |
| l-xylose | − | − | − | − | − | − | − | − |
| l-sorbose | − | − | − | − | − | − | − | + |
| methyl α-d-mannopyranoside | − | − | − | − | − | − | − | − |
| methyl α-d-glucopyranoside | − | − | − | − | − | − | − | − |
| N-acetylglucosamine | − | − | − | − | − | − | − | − |
| amygdalin | − | − | − | − | − | − | − | − |
| arbutin | − | − | − | − | − | − | − | − |
| inulin | − | − | − | − | − | − | − | − |
| d-melezitose | − | − | − | − | − | − | − | − |
| amidon (starch) | − | − | − | − | − | − | − | − |
| glycogen | − | − | − | − | − | − | − | − |
| xylitol | − | − | − | − | − | − | − | − |
| gentiobiose | − | − | − | − | − | − | − | − |
| potassium gluconate | − | − | − | − | − | − | − | − |
| potassium 2-ketogluconate | − | − | − | − | − | − | − | − |
| potassium 5-ketogluconate | − | − | − | − | − | − | − | − |
Legend: +: positive; −: negative.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Iličić, R.; Jelušić, A.; Barać, G.; Nikolić, D.; Stošić, N.; Scortichini, M.; Milovanović, T.P. In-Depth Characterization of Crown Gall Disease of Tobacco in Serbia. Agronomy 2024, 14, 851. https://doi.org/10.3390/agronomy14040851
AMA Style
Iličić R, Jelušić A, Barać G, Nikolić D, Stošić N, Scortichini M, Milovanović TP. In-Depth Characterization of Crown Gall Disease of Tobacco in Serbia. Agronomy. 2024; 14(4):851. https://doi.org/10.3390/agronomy14040851
Chicago/Turabian StyleIličić, Renata, Aleksandra Jelušić, Goran Barać, Dušan Nikolić, Nemanja Stošić, Marco Scortichini, and Tatjana Popović Milovanović. 2024. "In-Depth Characterization of Crown Gall Disease of Tobacco in Serbia" Agronomy 14, no. 4: 851. https://doi.org/10.3390/agronomy14040851
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.