
Developing an Effective Push–Pull System for Managing Outbreaks of the Invasive Pest Bactrocera dorsalis (Diptera: Tephritidae) in Nephelium lappaceum Orchards

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Works Taken before All Experiments
2.3. Experiment 1: Push–Pull System with and without Low-Concentration Pesticide
2.4. Experiment 2: Drone-Based System vs. Manual-Based System
2.5. Experiment 3: Food Bait vs. ME Bait
2.6. Experiment 4: Two Types of Repellents and Attractants vs. a Single Type
2.7. Data Analysis
3. Results
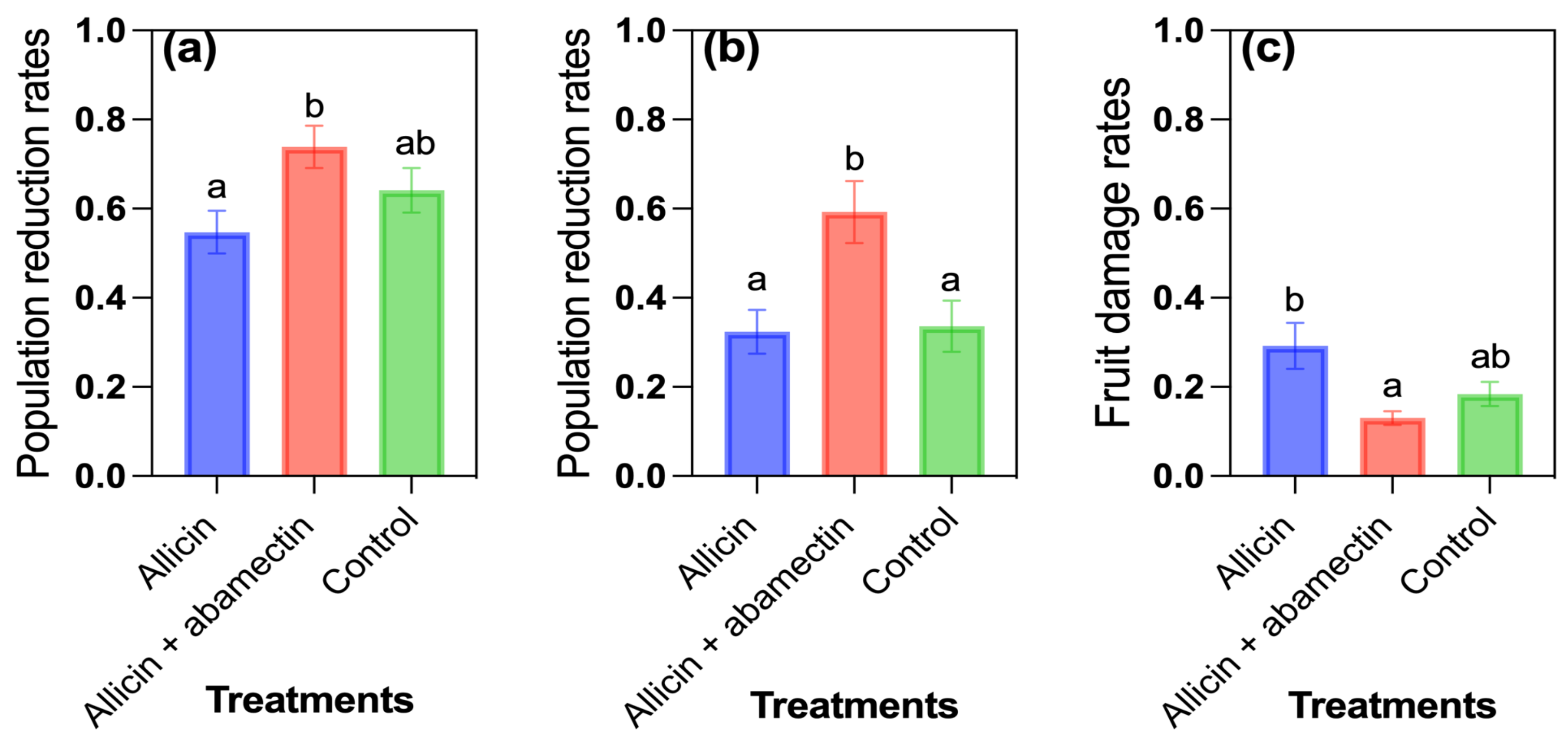
3.1. Push–Pull System with and without Low-Toxicity Pesticide
3.2. Drone-Based System vs. Manual-Based System
3.3. Food Bait vs. ME Bait
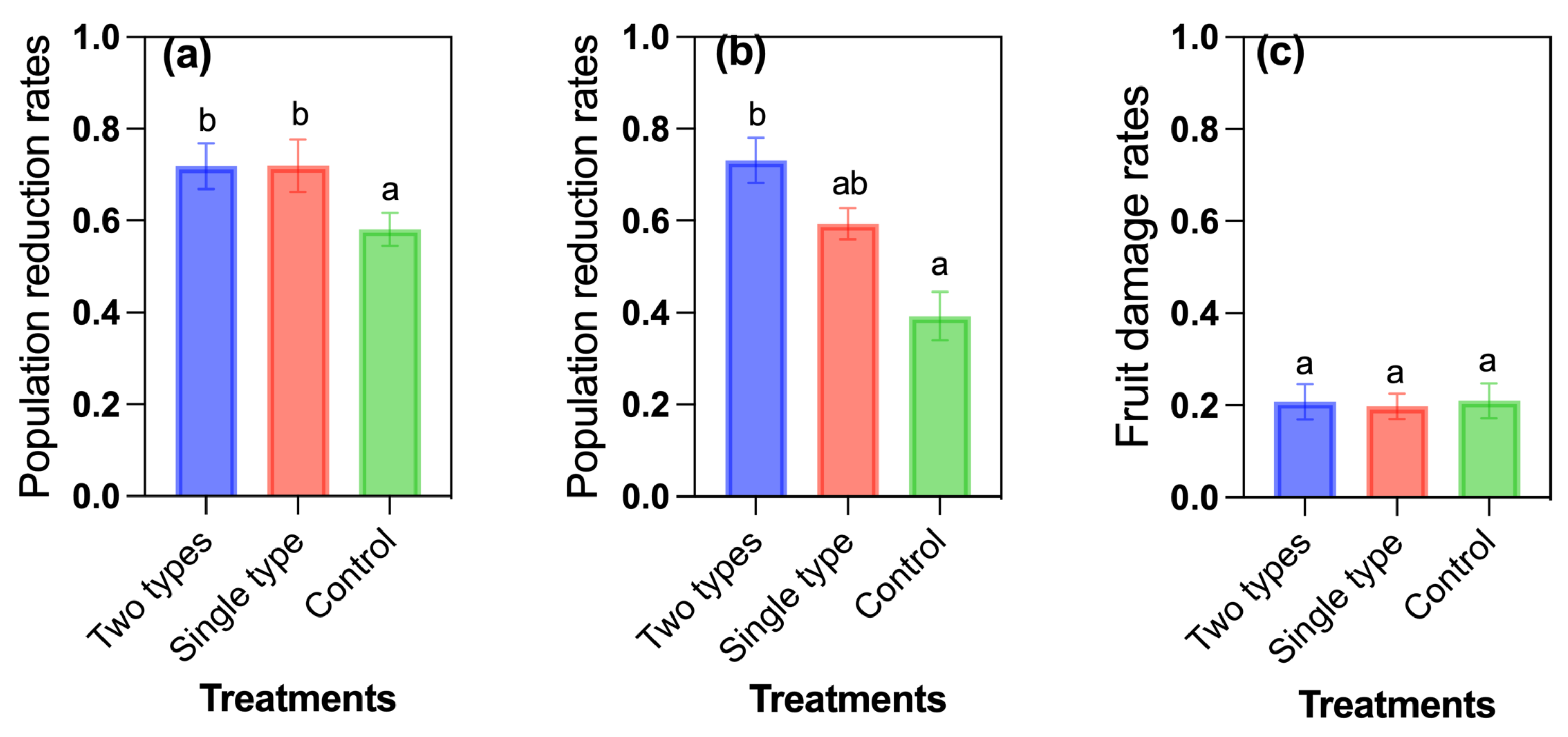
3.4. Two Types of Repellents and Attractants vs. A Single Type
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, H.; Zhang, D.; Xu, Y.; Wang, L.; Cheng, D.; Qi, Y.; Zeng, L.; Lu, Y. Invasion, expansion, and control of Bactrocera dorsalis (Hendel) in China. J. Integr. Agric. 2019, 18, 771–787. [Google Scholar] [CrossRef]
- Boinahadji, A.K.; Coly, E.V.; Diedhiou, C.A.; Sembene, P.M. Oviposition preference and offspring performance of the oriental fruit fly Bactrocera dorsalis (Diptera, Tephritidae) on eight host plants. Int. J. Adv. Res. 2020, 8, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Pangihutan, J.C.; Dono, D.; Hidayat, Y. The potency of minerals to reduce oriental fruit fly infestation in chili fruits. PeerJ 2022, 10, e13198. [Google Scholar] [CrossRef] [PubMed]
- Weems, H.V.; Heppner, J.B.; Nation, J.L.; Steck, G.J. Oriental Fruit Fly, Bactrocera dorsalis (Hendel) (Insecta: Diptera: Tephritidae). Available online: https://edis.ifas.ufl.edu/publication/IN240 (accessed on 10 September 2023).
- USDA. A Review of Recorded Host Plants of Oriental Fruit Fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae), Version 3.1. 2016. Available online: https://www.ars.usda.gov/research/publications/publication/?seqNo115=347371 (accessed on 7 October 2023).
- Choi, K.S.; Samayoa, A.C.; Hwang, S.Y.; Huang, Y.B.; Ahn, J.J. Thermal effect on the fecundity and longevity of Bactrocera dorsalis adults and their improved oviposition model. PLoS ONE 2020, 15, e0235910. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Zhao, R.; Zhou, W.; Pan, Y.; Tian, H.; Yin, Z.; Chen, W. Fruit fly in a challenging environment: Impact of short-term temperature stress on the survival, development, reproduction, and trehalose metabolism of Bactrocera dorsalis (Diptera: Tephritidae). Insects 2022, 13, 753. [Google Scholar] [CrossRef] [PubMed]
- Stephens, A.E.A.; Kriticos, D.J.; Leriche, A. The current and future potential geographical distribution of the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Bull. Entomol. Res. 2007, 97, 369–378. [Google Scholar] [CrossRef]
- Jaffar, S.; Lu, Y. Toxicity of some essential oils constituents against oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Insects 2022, 13, 954. [Google Scholar] [CrossRef]
- Sim, S.B.; Curbelo, K.M.; Manoukis, N.C.; Cha, D.H. Evaluating Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) response to methyl eugenol: Comparison of three common bioassay methods. J. Econ. Entomol. 2022, 115, 556–564. [Google Scholar] [CrossRef]
- Stibick, J.N.L. Natural Enemies of True Fruit Flies (Tephritidae). 2004. Available online: https://www.ippc.int/static/media/uploads/resources/natural_enemies_of_true_fruit_flies.pdf (accessed on 7 October 2023).
- Garcia, F.R.M.; Ovruski, S.M.; Suárez, L.; Cancino, J.; Liburd, O.E. Biological control of Tephritid fruit flies in the Americas and Hawaii: A review of the use of parasitoids and predators. Insects 2020, 11, 662. [Google Scholar] [CrossRef]
- Manoukis, N.C.; Cha, D.H.; Collignon, R.M.; Shelly, T.E. Terminalia larval host fruit reduces the response of Bactrocera dorsalis (Diptera: Tephritidae) adults to the male lure methyl eugenol. J. Econ. Entomol. 2018, 111, 1644–1649. [Google Scholar] [CrossRef]
- Orankanok, W.; Chinvinijkul, S.; Thanaphum, S.; Sitilob, P.; Enkerlin, W.R. Area-wide integrated control of oriental fruit fly Bactrocera dorsalis and guava fruit fly Bactrocera correcta in Thailand. In Area-Wide Control of Insect Pests; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 517–526. [Google Scholar] [CrossRef]
- Shelly, T.; McInnis, D. Sterile insect technique and control of tephritid fruit flies: Do species with complex courtship require higher overflooding ratios? Ann. Entomol. Soc. Am. 2016, 109, 1–11. [Google Scholar] [CrossRef]
- Vargas, R.I.; Piñero, J.C.; Leblanc, L. An overview of pest species of Bactrocera fruit flies (Diptera: Tephritidae) and the integration of biopesticides with other biological approaches for their management with a focus on the Pacific Region. Insects 2015, 6, 297–318. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Liang, G.; Zeng, L.; Lu, Y. Detoxification enzymes activities in different Bactrocera dorsalis (Hendel) populations and their relationship with the resistant levels. J. Environ. Entomol. 2014, 36, 58–67. [Google Scholar]
- Zayed, M.S.; Taha, E.K.A.; Hassan, M.M.; Elnabawy, E.S.M. Enhance systemic resistance significantly reduces the silverleaf whitefly population and increases the yield of wweet pepper, Capsicum annuum L. var. annuum. Sustainability 2022, 14, 6583. [Google Scholar] [CrossRef]
- Nalyanya, G.; Moore, C.B.; Schal, C. Integration of repellents, attractants, and insecticides in a “push-pull” strategy for managing German cockroach (Dictyoptera: Blattellidae) populations. J. Med. Entomol. 2000, 37, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Jaworski, C.C.; Ullah, F.; Jamil, M.; Badshah, H.; Ullah, F.; Luo, Y. Efficacy of lure mixtures in baited traps to attract different fruit fly species in guava and vegetable fields. Front. Insect Sci. 2023, 2, 984348. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.R.; Midega, C.A.O.; Amudavi, D.M.; Hassanali, A.; Pickett, J.A. On-farm evaluation of the ‘push-pull’ technology for the control of stemborers and striga weed on maize in western Kenya. Field Crops Res. 2008, 106, 224–233. [Google Scholar] [CrossRef]
- Lang, J.; Chidawanyika, F.; Khan, Z.R.; Schuman, M.C. Ecological chemistry of pest control in push-pull intercropping systems: What we know, and where to go? Chimia 2022, 76, 906. [Google Scholar] [CrossRef]
- Mala, M.; Baishnab, M.; Mollah, M.M.I. Push-pull strategy: An integrated approach to manage insect-pest and weed infestation in cereal cropping systems. J. Biosci. Agric. Res. 2020, 25, 2122–2127. [Google Scholar] [CrossRef]
- Guea, O.G.M.; Castrejón-Ayala, F.; Robledo, N.; Jiménez-Pérez, A.; Sánchez-Rivera, G. Plant selection for the establishment of push-pull strategies for Zea mays-Spodoptera frugiperda pathosystem in Morelos, Mexico. Insects 2020, 11, 349. [Google Scholar] [CrossRef] [PubMed]
- Byers, J.A.; Levi-Zada, A. Modelling push-pull management of pest insects using repellents and attractive traps in fruit tree orchards. Pest Manag. Sci. 2022, 78, 3630–3637. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.R.; Pittchar, J.O.; Midega, C.A.O.; Pickett, J.A. Push-pull farming system controls fall armyworm: Lessons from Africa. Outlooks Pest Manag. 2018, 29, 220–224. [Google Scholar] [CrossRef]
- Pyke, B.; Rice, M.; Sabine, B.; Zalucki, M.P. The push-pull strategy-behavioral control of Heliothis. Aust. Cotton Grow. 1987, 9, 7–9. [Google Scholar]
- Martel, J.W.; Alford, A.R.; Dickens, J.C. Synthetic host volatiles increase efficacy of trap cropping for management of Colorado potato beetle, Leptinotarsa decemlineata (Say). Agric. For. Entomol. 2005, 7, 79–86. [Google Scholar] [CrossRef]
- Zhang, R.L.; Chen, D.; Liu, J.; Zhuang, G.F.; Zhang, Z. The use of push-pull strategy in medical pests integrated management. Chin. J. Vector Biol. Control 2016, 6, 624–628. [Google Scholar]
- Verghese, A.; Mouly, R.; Shivananda, T.; Soumya, C.; Rashmi, M. A push-pull strategy for the management of the Oriental fruit fly, Bactrocera dorsalis (Hendel) in mango. Pest Manag. Hortic. Ecosyst. 2021, 26, 269–271. [Google Scholar]
- Adefegha, S.A.; Ogunsuyi, O.B.; Oboh, G. Purple onion in combination with garlic exerts better ameliorative effects on selected biomarkers in high-sucrose diet-fed fruit fly (Drosophila melanogaster). Comp. Clin. Pathol. 2020, 3, 713–720. [Google Scholar] [CrossRef]
- Kayode, O.T.; Rotimi, D.; Emmanuel, F.; Iyobhebhe, M.; Kayode, A.A.A.; Ojo, A.O. Contraceptive and biochemical effect of juice extract of Allium cepa, Allium sativum, and their combination in Canton fruit flies. J. Food Biochem. 2021, 18, e13821. [Google Scholar] [CrossRef]
- Keita, N.D.; Sarr, A.G.R.J.; Diome, T.; Cissokho, B. Study of the effects of biological extracts of “heals everything” Gymnanthemum amygdalinum and garlic Allium sativum on fruit flies Bactrocera dorsalis and their stage L3 larvae. J. Entomol. Zool. Stud. 2024, 12, 38–43. [Google Scholar] [CrossRef]
- Wen, T.; Sang, M.; Wang, M.; Han, L.; Gong, Z.; Tang, X.; Long, X.; Xiong, H.; Peng, H. Rapid detection of d-limonene emanating from citrus infestation by Bactrocera dorsalis (Hendel) using a developed gas-sensing system based on QCM sensors coated with ethyl cellulose. Sens. Actuators B Chem. 2021, 328, 129048. [Google Scholar] [CrossRef]
- De Souza, R.B.; Guimarães, J.R. Effects of avermectins on the environment based on its toxicity to plants and soil invertebrates-a review. Water Air Soil Pollut. 2022, 233, 259. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Hong, Y.; Huang, Z.; Zhang, J.; Huang, Q. Avermectin induces the oxidative stress, genotoxicity, and immunological responses in the Chinese mitten crab, Eriocheir sinensis. PLoS ONE 2019, 14, e0225171. [Google Scholar] [CrossRef] [PubMed]
- Pirasath, S.; Nageswaran, B.; Karunasena, R.P.V.; Gevakaran, M. Acute abamectin toxicity: A case report. Toxicol. Commun. 2021, 5, 66–68. [Google Scholar] [CrossRef]
- Qiu, D.; Xu, N.; Zhang, Q.; Zhou, W.; Wang, Y.; Zhang, Z.; Yu, Y.; Lu, T.; Sun, L.; Zhou, N.Y.; et al. Negative effects of abamectin on soil microbial communities in the short term. Front. Microbiol. 2022, 5, 1053153. [Google Scholar] [CrossRef] [PubMed]
- Biasazin, T.D.; Wondimu, T.W.; Herrera, S.L.; Larsson, M.; Mafra-Neto, A.; Gessese, Y.W.; Dekker, T. Dispersal and competitive release affect the management of native and invasive Tephritid fruit flies in large and smallholder farms in Ethiopia. Sci. Rep. 2021, 11, 2690. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.A.; Bezerra-Silva, G.C.D.; Mastrangelo, T. The host marking pheromone application on the management of fruit flies-A review. Braz. Arch. Biol. Technol. 2012, 55, 835–842. [Google Scholar] [CrossRef]
- Alkema, J.T.; Dicke, M.; Wertheim, B. Context-dependence and the development of push-pull approaches for integrated management of Drosophila suzukii. Insects 2019, 10, 454. [Google Scholar] [CrossRef]
- Bharathi, T.E.; Sathiyanandam, V.K.R.; David, P.M.M. Attractiveness of some food baits to the melon fruit fly, Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). Int. J. Trop Insect Sci. 2004, 24, 125–134. [Google Scholar] [CrossRef]
- Ekesi, S.; Mohamed, S.; Tanga, C.M. Comparison of food-based attractants for Bactrocera invadens (Diptera: Tephritidae) and evaluation of Mazoferm-Spinosad bait spray for field suppression in mango. J. Econ. Entomol. 2014, 107, 299–309. [Google Scholar] [CrossRef]
- Hasnain, M.; Saeed, S.; Naeem-Ullah, U.; Ullah, S. Development of synthetic food baits for mass trapping of Bactrocera zonata S. (Diptera: Tephritidae). J. King Saud Univ. Sci. 2022, 34, 101667. [Google Scholar] [CrossRef]
- Ono, H.; Hee, A.K.; Jiang, H. Recent advancements in studies on chemosensory mechanisms underlying detection of semiochemicals in Dacini fruit flies of economic importance (Diptera: Tephritidae). Insects 2021, 12, 106. [Google Scholar] [CrossRef] [PubMed]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. JASA 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Abbas, A.; Zhang, Z.; Zheng, H.; Alami, M.M.; Alrefaei, A.F.; Abbas, Q.; Naqvi, S.A.H.; Rao, M.J.; Mosa, W.F.A.; Hussainn, A.; et al. Drones in plant disease assessment, efficient monitoring, and detection: A way forward to smart agriculture. Agronomy 2023, 13, 1524. [Google Scholar] [CrossRef]
- Shahrooz, M.; Talaeizadeh, A.; Alasty, A. Agricultural spraying drones: Advantages and disadvantages. In Proceedings of the 2020 Virtual Symposium in Plant Omics Sciences (OMICAS), Bogotá, Colombia, 23–27 November 2020; pp. 1–5. [Google Scholar]
- Chen, P.; Douzals, J.P.; Lan, Y.; Cotteux, E.; Delpuech, X.; Pouxviel, G.; Zhan, Y. Characteristics of unmanned aerial spraying systems and related spray drift: A review. Front. Plant Sci. 2022, 13, 870956. [Google Scholar] [CrossRef]
- Ugwu, J.A. Evaluation of food lures for oriental fruit fly Bactrocera dorsalis (Diptera: Tephritidae) trapping on Chrysophyllum albidum in Ibadan, Nigeria. J. For. Res. Manag. 2020, 17, 72–81. [Google Scholar]
- Toukem, N.K.; Yusuf, A.A.; Dubois, T.; Abdel-Rahman, E.M.; Adan, M.S.; Mohamed, S.A. Landscape vegetation productivity influences population dynamics of key pests in small avocado farms in Kenya. Insects 2020, 11, 424. [Google Scholar] [CrossRef]
- Showler, A.T.; Harlien, J.L.; de Léon, A.A.P. Effects of laboratory grade. limonene and a commercial limonene-based insecticide on Haematobia irritans (Muscidae: Diptera): Deterrence, mortality, and reproduction. J. Med. Entomol. 2019, 56, 1064–1070. [Google Scholar] [CrossRef]
- Hassanali, A.; Herren, H.; Khan, Z.R.; Pickett, J.A.; Woodcock, C.M. Integrated pest management: The push-pull approach for controlling insect pests and weeds of cereals, and its potential for other agricultural systems including animal husbandry. Philos. Trans. R. Soc. B 2008, 363, 611–621. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Push (Repellent Used) | Pull (Attractant Used) |
|---|---|
| Experiment 1: Push–Pull System with and without Low-Concentration Pesticide | |
| Allicin | Methyl eugenol |
| Allicin + abamectin | Methyl eugenol |
| Experiment 2: Drone-Based System vs. Manual-Based System | |
| Drone-applied allicin + abamectin | Methyl eugenol |
| Manually applied allicin + abamectin | Methyl eugenol |
| Experiment 3: Food Bait vs. ME Bait | |
| Manually applied allicin + abamectin | Food bait |
| Manually applied allicin + abamectin | Methyl eugenol |
| Experiment 4: Two Types of Repellents and Attractants vs. a Single Type | |
| Allicin + d-limonene + abamectin | Food bait + methyl eugenol |
| Allicin | Methyl eugenol |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, J.; Shan, Z.; Zou, Y.; Lin, X.; Cui, Z.; Yan, R.; Cao, F. Developing an Effective Push–Pull System for Managing Outbreaks of the Invasive Pest Bactrocera dorsalis (Diptera: Tephritidae) in Nephelium lappaceum Orchards. Agronomy 2024, 14, 890. https://doi.org/10.3390/agronomy14050890
Wen J, Shan Z, Zou Y, Lin X, Cui Z, Yan R, Cao F. Developing an Effective Push–Pull System for Managing Outbreaks of the Invasive Pest Bactrocera dorsalis (Diptera: Tephritidae) in Nephelium lappaceum Orchards. Agronomy. 2024; 14(5):890. https://doi.org/10.3390/agronomy14050890
Chicago/Turabian StyleWen, Jian, Zhe Shan, Yan Zou, Xianwu Lin, Zhifu Cui, Rihui Yan, and Fengqin Cao. 2024. "Developing an Effective Push–Pull System for Managing Outbreaks of the Invasive Pest Bactrocera dorsalis (Diptera: Tephritidae) in Nephelium lappaceum Orchards" Agronomy 14, no. 5: 890. https://doi.org/10.3390/agronomy14050890