
Molecular Regulation of Flowering Time in Grasses

1
Graduate School of Environmental Science, Hokkaido University, Kita 10 Nishi 5, Sapporo, Hokkaido 060-0810, Japan
2
Field Science Center for Northern Biosphere, Hokkaido University, Sapporo, Hokkaido 060-0810, Japan
*
Author to whom correspondence should be addressed.
Agronomy 2017, 7(1), 17; https://doi.org/10.3390/agronomy7010017
Submission received: 29 November 2016
/
Revised: 11 February 2017
/
Accepted: 13 February 2017
/
Published: 20 February 2017
(This article belongs to the Special Issue Genetics and Breeding for Productivity Traits in Forage and Bioenergy Grasses)
{kind=link}
Abstract
:Flowering time is a key target trait for extending the vegetative phase to increase biomass in bioenergy crops such as perennial C4 grasses. Molecular genetic studies allow the identification of genes involved in the control of flowering in different species. Some regulatory factors of the Arabidopsis pathway are conserved in other plant species such as grasses. However, differences in the function of particular genes confer specific responses to flowering. One of the major pathways is photoperiod regulation, based on the interaction of the circadian clock and environmental light signals. Depending on their requirements for day-length plants can be classified as long-day (LD), short-day (SD), and day-neutral. The CONSTANS (CO) and Heading Date 1 (Hd1), orthologos genes, are central regulators in the flowering of Arabidopsis and rice, LD and SD plants, respectively. Additionally, Early heading date 1 (Ehd1) induces the expression of Heading date 3a (Hd3a), conferring SD promotion and controls Rice Flowering Locus T 1 (RFT1) in LD conditions, independently of Hd1. Nevertheless, the mechanisms promoting flowering in perennial bioenergy crops are poorly understood. Recent progress on the regulatory network of important gramineous crops and components involved in flowering control will be discussed.
1. Introduction
Global climate change and energy security issues have promoted interest in the production and increased availability of alternative energy sources. Lignocellulosic biomass is a promising feedstock source for biorefineries producing biofuel, which can mitigate greenhouse gas emissions [1,2,3] and reduce dependency on fossil oil [4,5]. Perennial C4 bioenergy crops such as switchgrass (Panicum virgatum L.) and Miscanthus spp. provide good targets as non-edible plant species [6,7,8] having advantages with regard to land utilization and the avoidance of conflict with food security [9,10,11] to provide efficient production systems at low cost.
One of the most important traits in the plant life cycle is the timing of flowering, the floral transition between the vegetative and reproductive phases of plant development [12,13]. Consideration of flowering time is an important strategy in the cultivation of grain crops in northern latitudes. Early flowering is useful in regions where growing seasons are short to enhance grain yield stability by avoiding drought or adverse temperatures [14,15,16]. Flowering time is also a major determinant of biomass yield in perennial C4 bioenergy crops, because delayed flowering time allows an extended period of vegetative growth and produces more biomass. Thus, earlier flowering will produce lower yields than late flowering in terms of feedstock production [17]. However, biomass potential also depends on environmental conditions. In switchgrass, lowland ecotypes that originate from southern areas flower later in high latitude areas, but the yield advantages of these southern switchgrasses are often not realized at northern latitudes due to high winter mortality [18].
Natural variation in flowering time is related to latitude in several plant species. Migration of plants into different latitudes often require the adoption of different signals to induce flowering and promote adaptive responses to diverse growing seasons [19]. Factors such as photoperiod and temperature that vary over large geographical scales are involved [20]. Plants possess an internal biological clock providing circadian rhythms that respond to fluctuations in day-length and thus anticipate upcoming seasonal changes [21] to regulate flowering. Depending on their requirements for day-length (light period in a 24-h cycle) to promote flowering, plants can be classified as long-day (LD) plants when photoperiod exceeds a critical day-length, short-day (SD) plants when photoperiod is shorter than a critical day-length and day-neutral plants when flowering occurs irrespective of day-length [22,23,24]. Winter annuals (e.g., wheat (Triticum aestivum L.)), biennials (e.g., sugar beet (Beta vulgaris L.)) and numerous perennials (e.g., orchardgrass (Dactylis glomerata L.)) are obligatory LD plants. These plants, however, flower only after vernalization during a cold period [23]. The molecular basis of flowering time regulation has been extensively studied using classical Quantitative Trait Loci (QTL) approaches in model plant species such as Arabidopsis thaliana [25,26,27], a LD plant, and rice (Oryza sativa L.), a SD plant [28]. Grasses have multiple pathways to control flowering time but only some of them are conserved in Arabidopsis thaliana (L.) Heynh. [29]. These studies have been crucial in establishing the multiple pathways that control flowering, of which the photoperiod pathway is of major importance [30]. The use of model species has played a major role in understanding the molecular mechanisms involved in flowering time to help in the genetic improvement of crop development. Nevertheless, little is known about the mechanisms promoting flowering in perennial C4 bioenergy crops. In this review, we discuss recent progress concerning the regulatory network and components involved with flowering control in different species including C4 grasses such as sorghum (Sorghum bicolor (L.) Moench), switchgrass and Miscanthus spp. To understand how plants initiate flowering is a crucial step in developing selection criteria in breeding programs of grasses used as bioenergy crops.
2. Conservation and Divergence in Flowering Pathways
2.1. Arabidopsis and Rice
In the last decades, studies on the model plant Arabidopsis have revealed that molecular mechanisms discovered in that species are evolutionally conserved in other species [15]. Genetic approaches in Arabidopsis have identified three genes that control flowering: GIGANTEA (GI)—CONSTANS (CO)—FLOWERING LOCUS (FT) [15,28]. Loss of function mutations in each gene for flowering control delay flowering under LD conditions but no effect is produced under SD [31]. GI is a key regulator of the photoperiodic pathway and in the evening promotes CO transcription under LD conditions [32]. The most extensively gene studied in Arabidopsis flowering is CO that confers LD responses. CO encodes a B-box zinc finger transcription factor and CCT domain genes that promote flowering under LD conditions and activate the expression of FT [33,34,35], a major component of the florigen that induces flower differentiation [29]. Its inactivation causes flowering delay, while its over-expression induces early flowering. CO and FT are expressed in the phloem and act there to promote flowering [31]. This signalling pathway is conserved in rice: OsGI-Hd1-Hd3a [28], mediated by LD responses. OsGIGANTEA (OsGI) acts an activator of Heading date1 (Hd1), an ortholog of CO, and controls flowering time by modulating rhythmic flowering under SD [36]. Hd1 encodes a zinc finger type transcriptional activator with the conserved CCT (CO, CO-like, TIMING OF CAB EXPRESSION1 (TOC1)) domain [15,37]. In contrast to Arabidopsis CO, Hd1 promotes Heading date 3a (Hd3a) expression in SD but expression is modified in LD conditions [38,39] where Hd1 function is converted into a repressor. Rice involves at least two flowering pathways that control the expression of the florigens: Hd1 that is conserved in rice and Arabidopsis, and Early heading date 1(Ehd1), without an ortholog in Arabidopsis [39,40] (Figure 1). Moreover, Grain Number, Plant Height, and Heading Date7 (Ghd7) is unique in grasses [16]. Ehd1 is a B-type response regulator that induces the expression of Hd3a in rice, conferring SD promotion of flowering in the absence of a functional allele of Hd1. It also controls Rice Flowering Locus T (RFT) gene in LD conditions independently of Hd1 [40]. In LD conditions Hd1 acts as a flowering repressor inhibiting Hd3a expression but promotes its expression and subsequent flowering in SD [41]. Ghd7 is a small protein with a CCT-domain that represses Ehd1 expression and downstream Hd3a/RFT1 expression in LD conditions to delay flowering [42,43,44,45]. Recent studies demonstrated that the interaction between Ghd7 and Hd1 can play a critical role in repressing Ehd1. Under SD conditions Hd1 activates the expression of Ehd1 at night but not in the day while under LD conditions Hd1 represses its expression in the morning. Indeed, Hd1 repressor activity requires a proper Ghd7 function under LD conditions to repress Ehd1 in the morning. In contrast, Ghd7 can repress the expression of Ehd1, Hd3a and RFT1 by itself under all photoperiod conditions [15].
2.2. C4 Grasses
Previous studies identified two floral activators in sorghum, a SD plant: SbEhd1 and SbCO. SbCO is a homolog of the floral activator CO in Arabidopsis and an ortholog of Hd1 in rice. It promotes early flowering in both LD and SD conditions, and increases the expression of SbEhd1, SbCN8, SbCN12 and SbCN15 [16]. Genetic analyses and expression studies in sorghum reveal that SbCO shares a conserved CCT-domain with TOC1, PSEUDORESPONSE REGULATOR PROTEIN 37 (PRR37), Ghd7 and HEME ACTIVATOR PROTEINS (HAP). SbCO also increases expression of SbEhd1, a promoter of Hd3a in rice. In comparison with rice, Ehd1 regulates positively the expression of RFT1 to promote flowering; however, no ortholog of RFT1 is present in the sorghum genome. SbPRR37 (Ma1) and Ghd7 (Ma6) inhibit flowering, reducing the expression of SbEhd1 and SbCN8/12 (florigens) under LD conditions, but not in SD (Figure 1). The ability of SbPRR37 to inhibit their expression could be due to inhibition of SbEhd1 or SbCO, activators of SbCN8 and SbCN12 expression [14,16]. The PhyB regulation of SbCN15 expression may modify flowering time in a photoperiod-insensitive manner [46] (Figure 1). In switchgrass, the flowering time regulatory network is similarly to maize (Zea mays L.) and is regulated by both photoperiod-dependent and autonomous pathways. Some conserved flowering genes such as FT-like gene (ZCN8 in maize) and INDETERMINATE 1 (ID1) have also been identified in the maize genome. The study of genes involved in flowering of switchgrass is relatively new. Hence, the functions of FT-like gene in switchgrass germplasm have not been clarified yet but may contribute to delayed flowering time as in maize [18]. Thus, the switchgrass FT homolog may have similar functions to the maize FT gene and is down-regulated by the expression of LONG VEGETATIVE PHASE ONE (AtLOV1) in switchgrass [18]. Overexpression of AtLOV1 causes delayed flowering time in switchgrass but does not enhance cold tolerance as in Arabidopsis [18]. Sorghum is closely related to Miscanthus spp., a promising candidate C4 bioenergy crop in temperate climates. The CO/Hd1 sequence in Miscanthus sinensis Andersson was identified as MsiHd1 with two types of diverged loci, MsiHd1a and MsiHd1b. The MsiHd1 gene encodes two conserved B-box zinc finger domains and a CCT domain. Two to five different alleles of MsiHd1 were found in Miscanthus accessions from mainland Asia and from Japan, suggesting that MsiHd1 consists of at least three loci in the Miscanthus genome with small differences in the number of functional alleles [38]. From preliminary data we identified at least three alleles suggesting that MsiEhd1 has two loci in the Miscanthus genome, MsiEhd1a and MsiEhd1b in comparison to sorghum, rice and maize, which have only one gene. We also detected two loci in MsiGhd7. The current diploid M. sinensis evolved from genome duplication of its progenitor that was very close to a sorghum ancestor [46]. Gene duplication is a key mechanism in evolution because it can provide genes with new functions.
3. QTLs Analysis
To understand the complex genetic network of flowering in perennial ryegrass (Lolium perenne L.), a C3 forage grass, a number of genes have been identified through QTL mapping, using different plant material and genetic maps and by sequence homology with Arabidopsis, rice and maize [47]. The genomic and phenotypic variations associated with perennial ryegrass LpFT3, an ortholog of FT, were assessed in a diverse collection of nine European germplasm populations, identifying a total of 7 haplotypes. The results indicated a significant association between allelic variation in the LpFT3 gene and flowering time. Haplotype C was associated with early flowering and the A and B haplotypes with late flowering. The variations were identified in the predicted sequence and in non-coding regions, mainly within the 5′ region of the coding sequence which is strongly conserved [48]. Comparative analysis established close proximity between genetic markers related to the DGL1, Ph1 and OsPIPK1 ortholoci and the corresponding perennial ryegrass QTLs. This suggests that DGL1 and Ph1 ortholoci may provide candidate genes for the herbage yield-related QTLs on linkage group 3 (LG3). The physical location of the OsPIPK1 gene (a heading date locus) was located at the 28.2 Mb position of rice chromosome 3, close to the predicted CDO795 ortholocus (23.1 Mb). Further studies have suggested that the CDO795-linked heading date QTL was equivalent to a rice heading date QTL, dth3.3 (Gramene QTL Acc. ID AQFE011). As a consequence, the perennial ryegrass OsPIPK1 ortholocus may be related to the heading date QTLs on LG4 [49]. In addition, the major QTL in the F2/WSC and ILGI perennial ryegrass populations was identified on LG7, which is associated with the position of the genes Hd3a and Hd1, two heading date genes of rice on chromosome 6. However, analysis of the ILGI population grown in Japan identified a QTL on LG4, but not the QTL reported on LG7 [50,51]. This result emphasizes the importance of adaptation in plants to the broad range of agro-environmental conditions in which they grow. In sorghum, three significant QTL associated with flowering time, PHYB (Ma3), PHYC (Ma5) [52] and SbGHD7 (Ma6) [14,52] were identified, through analysis of flowering variation in LD using an F2 population, which explained ~50% of the phenotypic variance for flowering time [52]. Recessive ma3R alleles from 58 M populations associated with Ma3 QTL produced early flowering time phenotypes; however, dominant alleles of SbGhd7 (Ma6) and SbPRR37 act in an additive manner to delay floral initiation for ~175 days until day-lengths decrease below 12.3 h [14,52]. Sorghum accessions exhibit significant variations in flowering time in response to day-length. One QTL controlling photoperiod sensitivity was detected on chromosome 1 under SD, and one QTL controlling photoperiod insensitivity expression was detected on chromosome 4 under 12 h and natural photoperiod conditions, from the SSR markers Xtxp61 and Xtxp51 respectively [53]. Under LD, a cross between tropical and temperate sorghums (Sorghum propinquum (Kunth) Hitchc.×S. bicolor (L.) Moench), revealed one QTL FlrAvgD1 located in chromosome 6 in a 10 kb interval, which accounted for 85.7% of the variation in flowering time. This interval contains a single annotated gene, Sb06g012260, which is a member of the FT family of transcription factors. Sb06g012260 is unique to panicoids and suppresses flowering, although it is quite distant evolutionarily from other FT family members that are floral suppressors [54]. In M. sinensis five putative flowering QTLs were detected using the Multiple QTL model (MQM) approach for plants grown in the years 2000 and 2001 [55]. Only QTL F12 was detected in both years on LG1, F11 and F13 were only detected in the first year while F14 and F15 were detected in the second year. So these QTLs may depend on interactions between genotype and environment. A genome-wide association study may be identified association with gene network in flowering time.
4. Circadian Clock and Photoperiod Response
The circadian clock plays an important role in seasonal flowering time regulation of angiosperms; photoperiodic time measurement is based on the interaction between the endogenous circadian clock and environmental light signals in Arabidopsis [56,57]. The plant circadian system consists of biochemical timing mechanisms that temporarily modulate the function of several signalling pathways to measures changes in day-length and promote suitable timing of flowering to maximize reproductive success [2,3,6,7,9,10,13,14,15,21,24,26,28,29,31,36,38,41,42,46,48,49,56,57,58,59,60,61,62,63,64,65,66]. The photoperiod response on flowering time varies among grasses. Barley (Hordeum vulgare L.) and wheat are LD plants, while rice and sorghum are SD plants [52]. Flowering is regulated through the CO and FT genes [66]. GI plays an important role in regulating the circadian clock and flowering, promoting CO gene expression and light response. The rice ortholog of GI, OsGI, is a positive regulator of Hd1 expression under both SDs and LDs [24]. Mutation in OsGI reduced photoperiod sensitivity in rice [36] and affected the expression of LATE ELONGATED HYPOCOTYL (LHY) and several PSEUDO RESPONSE REGULATOR (PRR) genes. However, PRR37 expression was not affected in the osgi mutant, suggesting independent control of heading date by these factors [66]. Hd1 is predominantly regulated by the circadian clock through OsGIGANTEA (OsGI) [64] and possesses two contrasting functions in the regulation of the rice ortholog of Arabidopsis FT gene, Hd3a. The bi-functionally mechanism of Hd1 involves the action of the red-light photoreceptor phytochrome B (phyB), a primary cause of long-day suppression of flowering in rice [39,60]. Over-expression of Hd1 causes a delay in flowering under SD conditions and a single extension of day-length decreases Hd3a expression consistently with the duration of daylight [44,59]. The repression of flowering by Hd1 under LD conditions is enhanced by the kinase activity of Heading date 6 (Hd6), a gene encoding the α subunit of protein kinase CK2 (CK2α) [58]. Hd6 is a QTL involved in photoperiod response in rice. To induce delayed flowering under LD conditions, Hd6 requires the presence of functional Hd1 alleles and plays a critical role in Hd1 activity. Despite this, Hd6 regulation is not mediated by changes in the circadian clock [37,65]. The rice genome contains two important genes for photoperiodic regulation: Ehd1 and Ghd7, specific to grass species such as rice, maize and sorghum but absent in the Arabidopsis genome [44]. The expression of Hd3a is also regulated by Ehd1 conferring SD promotion of flowering and controlling FT-like gene expression independently of the Hd1/CO photoperiodic flowering pathway [30,40] (Figure 1). Hd1 and Ehd1 expression are controlled by the circadian clock, although Ehd1 is also regulated by both blue and red light. In sorghum, SbCO expression is not altered significantly in response to day-length. However, Ghd7, a floral repressor regulated by the circadian clock and light, represses the expression of SbEhd1 and SbCN8 [14]. Ehd1 expression in rice is strongly repressed by Ghd7 in LD conditions but in SD conditions Ghd7 rarely affects flowering time [36].
5. Photoreceptors Involved in Flowering Time
Plants use the phytochrome system to regulate time of flowering and adjust growth based on the duration of dark and light periods (photoperiodism), while the spectrum of the light also affects flowering. Plants use many photoreceptors to detect the intensity and quality of light, including PHYTOCHROMES (PHY), which absorb the red and far-red region of the visible spectrum, and the CRYPTOCHROMES (CRY) [63]. Arabidopsis contains five PHYs (A-E), where accumulation of CO in LD is due to stabilization mediated by phytochrome A (PHYA), cryptochromes (CRY1/2) and SUPPRESSOR OF PHYA-105 (SPA1) [61]. However, it has been shown that PHYB signals delay flowering by destabilizing CO protein during the morning and have an inhibitory effect on FT expression [60]. The phytochromes PHYA, PHYB, CRY1 and CRY2 are directly clock regulated under specific light conditions. For example, over-expression of the photoreceptor PHYA under SD conditions promotes flowering, but phyA mutants delay flowering in LD conditions. In contrast to Arabidopsis, rice and sorghum encodes three phytochromes (PHYA, PHYB and PHYC) [12,52], where phyA mutants of rice do not produce significant alterations in flowering time [52,63]. This is despite the high similarity between the PHYA locus in Arabidopsis rice, sorghum and maize [62] suggesting that a similar response would be expected. However, PHYA mutations in combination with PHYB or PHYC cause early flowering in rice [61]. In addition, studies have shown that PHYC plays an essential role in the acceleration of wheat flowering under LD photoperiods. Moreover, it is stable and functionally active even in the absences of other phytochromes, compared with rice and Arabidopsis [67]. Blue and far-red lights promote flowering in Arabidopsis and rice, acting through the action of PHYA, CRY1 and CRY2 photoreceptors in Arabidopsis, while red light delays flowering [12,33,43,67]. On the other hand, PHYB modulates the expression of genes in response to red light and is the main component of the shade-avoidance mechanism in Arabidopsis. PhyB mutants revealed that PHYB inhibits flowering under both LD and SD photoperiods, but an over-expression of PHYB in LDs results in early flowering [22]. PHYB is responsible for delayed flowering and Hd3a suppression in the presence of a night-break (NB) treatment and activates the Hd1 expression in rice. The NB treatment is a short exposure to light in the middle of night and was widely used to understand the role of the circadian clock and light on flowering. In recent studies, photoperiodic sensitivity 5 (SE5) and PHYB also suppress Ehd1 expression, by suppressing Oryza sativa CO-like4 (OsCOL4) [39]. Rice mutants deficient in PHYB have reduced sensitivity to red light and are early flowering [43,60]. In contrast, phyB null mutations in wheat are connected with delayed flowering [24]. In sorghum, under LD, PHYB (Ma3) is required for elevated expression of SbPRR37 and SbGHD7 during the evening to inhibit flowering (Figure 1). This response results in repression of SbEHD1, SbCN12, SbCN8 and floral initiation. Ghd7 represses Ehd1 expression in response to the red light signal in the morning mediated by phytochromes. In SD conditions, PHYB may have a limited effect on the expression of these genes as peak SbPRR37 and SbGHD7 expression is highest in the morning and lowest during the evening compared with expression in LD. The inactivation of PHYB results in early flowering in LD [43,52].
6. Conclusions and Perspectives
Genetic analysis in Arabidopsis has allowed the identification of different pathways that promote flowering in response to environmental conditions and developmental regulation. The primary mechanism of the photoperiod pathway in plants is evolutionary conserved for flowering signalling. CO is the central regulator in promoting flowering and exhibits complex regulation. In addition day-length and the circadian clock control critical aspects of flowering. The effect of GI on flowering is associated with promoting expression of genes related to the circadian pathway. Arabidopsis is considered as a model plant to understand flower development while some grasses have defined their own responses and adaptation strategies. For example, Ehd1 and Ghd7 genes are unique in grasses in relation to the promotion and repression of flowering time respectively. Due to the lack of nucleotide information in Miscanthus, few genetic resources have been developed to clarify the relationship of the Miscanthus genome to its close relatives, sorghum and sugarcane (Saccharum spp.). Comparisons between the Sorghum genome and the genus Miscanthus reveal that whole genome duplication occurred in Miscanthus after its divergence from a common ancestor shared with sorghum. The base chromosome number of Miscanthus is approximately twice that of sorghum with nominally diploid and tetraploid species [46,68]. Analysis of natural variation in flowering in different ecotypes of grasses with economic value, such as Miscanthus spp. and switchgrass, is necessary to clarify the molecular network of flowering time control in these species. Through breeding programs, favorable alleles of QTLs then can then be efficiently introduced into elite cultivars to generate new varieties with high biomass productivity and beneficial adaptations to environmental changes.
Acknowledgments
We gratefully acknowledge Mervyn O. Humphreys, for critical reading of the manuscript. F.D.B.N. acknowledgments the Ministry of Education, Culture, Sports, Science, and Technology (MEXT) scholarship for Master Course study at the Hokkaido University, Japan.
Author Contribution
F.D.B.N., written paper; T.Y., edited paper, mentored MS student (F.D.B.N.)
Conflicts of Interest
The authors declare no conflict of interest.
References
- Clifton-Brown, J.C.; Stampfl, P.F.; Jones, M.B. Miscanthus biomass production for energy in Europe and its potential contribution to decreasing fossil fuel carbon emission. Glob. Chang. Biol. 2004, 10, 509–518. [Google Scholar] [CrossRef]
- Lewandowski, I.; Kicherer, A.; Vonier, P. CO2-balance for the cultivation and combustion of Miscanthus. Biomass Bioenerg. 1995, 8, 81–90. [Google Scholar] [CrossRef]
- Lewandowski, I.; Kicherer, A. Combustion quality of biomass: Practical relevance and experiments to modify the biomass quality of Miscanthus × giganteus. Eur. J. Agr. 1997, 6, 163–177. [Google Scholar] [CrossRef]
- Fargione, J.; Hill, J.; Tilman, D.; Polasky, S.; Hawthorne, P. Land clearing and the biofuel carbon debt. Science 2008, 319, 1235–1238. [Google Scholar] [CrossRef] [PubMed]
- Timothy, S.; Ralph, H.; Houghton, R.A.; Fengxia, D.; Amani, E.; Jacinto, F.; Simla, T.; Dermot, H.; Tun-Hsiang, Y. Use of US croplands for biofuels increases greenhouse gases through emissions from land-use change. Science 2008, 319, 1238–1240. [Google Scholar]
- Oliver, R.J.; Finch, J.W.; Taylor, G. Second generation bioenergy crops and climate change: A review of the effects of elevated atmospheric CO2 and drought on water use and the applications for yield. GCB Bioenerg. 2009, 1, 97–114. [Google Scholar] [CrossRef]
- Somerville, C. Biofuels. Curr. Biol. 2007, 17, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.S.; Tiller, K.H.; Al-Ahmad, H.; Stewart, N.R.; Stewart, C.N. Plants to power: Bioenergy to fuel the future. Trends Plant Sci. 2008, 13, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Heaton, E.A.; Frank, G.; Dohleman, F.G.; Long, S.P. Meeting biofuel goals with less land: The potential of Miscanthus. Glob. Chang. Biol. 2008, 14, 2000–2014. [Google Scholar] [CrossRef]
- Henry, R.J. Evaluation of plant biomass resources available for replacement of fossil oil. Plant Biotechnol. J. 2010, 8, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Socolow, R.; Foley, J.A.; Hill, J.; Larson, E.; Lynd, L.; Pacala, S.; Reilly, J.; Searchinger, T.; Somerville, C.; et al. Beneficial biofuels-the food, energy, and environment trilemma. Science 2009, 325, 270–271. [Google Scholar] [CrossRef] [PubMed]
- Galvão, V.C.; Schmid, M. Regulation of flowering by endogenous signals. In Advance in Botanical Research; Fornara, F., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 72, pp. 63–102. [Google Scholar]
- Joshi, C.P.; DiFazio, S.P.; Chittaranjan, K. Genetics, Genomics and Breeding of Poplar; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Murphy, R.L.; Morishige, D.T.; Brady, J.A.; Rooney, W.L.; Yang, S.; Klein, P.E.; Mullet, J.E. Ghd7 (Ma6) represses sorghum flowering in long days: Ghd7 alleles enhance biomass accumulation and grain production. Plant Genome 2014, 7, 1–10. [Google Scholar] [CrossRef]
- Nemoto, Y.; Nonoue, Y.; Yano, M.; Izawa, T. Hd1, a CONSTANS ortholog in rice, functions as an Ehd1 repressor through interaction with monocot-specific CCT-domain protein Ghd7. Plant J. 2016, 86, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Weers, B.D.; Morishige, D.T.; Mullet, J.E. CONSTANS is a photoperiod regulated activator of flowering in sorghum. BMC Plant Biol. 2014, 14, 148. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, P.P.; Evans, J.; Daum, C.; Deshpande, S.; Barry, K.W.; Kennedy, M.; Casler, M.D. Genome-wide associations with flowering time in switchgrass using exome-capture sequencing data. New Phytol. 2017, 213, 154–169. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Sathitsuksanoh, N.; Tang, Y.; Udvardi, M.K.; Zhang, J.; Shen, Z.; Balota, M.; Harich, K.; Zhang, P.; Zhao, B. Overexpression of AtLOV1 in switchgrass alters plant architecture, lignin content, and flowering time. PLoS ONE 2012, 7, e47399. [Google Scholar] [CrossRef] [PubMed]
- Colasanti, J.; Coneva, V. Mechanisms of floral induction in grasses: Something borrowed, something new. Plant Physiol. 2009, 149, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Brachi, B.; Faure, N.; Horton, M.; Flahauw, E.; Vazquez, A.; Nordborg, M.; Roux, F. Linkage and association mapping of Arabidopsis thaliana flowering time in nature. PLoS Genet. 2010, 6, e1000940. [Google Scholar] [CrossRef] [PubMed]
- Hayama, R.; Coupland, G. Shedding light on the circadian clock and the photoperiodic control of flowering. Curr. Opin. Plant Biol. 2003, 6, 13–19. [Google Scholar] [CrossRef]
- Blázquez, M.A.; Piñeiro, M.; Valverde, F. Bases moleculares de la floración. Prensa científica. Temas 61. Investig. Cienc. 2011, 416, 29–36. [Google Scholar]
- Gardner, F.P.; Pearce, B.; Mitchell, R. Physiology of Crop Plants, 1st ed.; Iowa State University Press: Ames, IA, USA, 1985. [Google Scholar]
- Pearce, S.; Kippes, N.; Chen, A.; Debernardi, J.M.; Dubcovsky, J. RNA-seq studies using wheat PHYTOCHROME B and PHYTOCHROME C mutants reveal shared and specific functions in the regulation of flowering and shade-avoidance pathways. BMC Plant Biol. 2016, 16, 141. [Google Scholar] [CrossRef] [PubMed]
- Coupland, G. Regulation of flowering time: Arabidopsis as a model system to study genes that promote or delay flowering. Philos. Trans. R Soc. Lond. B Biol. Sci. 1995, 350, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.G.; Dean, C. Arabidopsis, the Rosetta stone of flowering time? Science 2002, 296, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Valentin, F.L.; Van Mourik, S.; Posé, D.; Kim, M.C.; Schmid, M.; Van Ham, R.; Busscher, M.; Sanchez-Perez, G.F.; Molenaar, J.; Angenent, G.C.; et al. A quantitative and dynamic model of the Arabidopsis flowering time gene regulatory network. PLoS ONE 2015, 10, e0116973. [Google Scholar]
- Peña, N. Caracterización de la Variación Natural de Hd3a y RFT1 en Cultivares de Tipo Japónica de Oryza sativa. Máster’s Thesis in Molecular and Cellular Biotechnology of Plants, The Technical University of Valencia, Valencia, Spain, 2013. [Google Scholar]
- Mauro-Herrera, M.; Wang, X.; Barbier, H.; Brutnell, T.P.; Devos, K.M.; Doust, A.N. Genetic control and comparative genomic analysis of flowering time in Setaria (Poaceae). G3 (Bethesda) 2013, 3, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Wada, M.; Shimazaki, K.; Lino, M. Light Sensing in Plants; Springer Science & Business Media: New York, NY, USA, 2005; pp. 333–346. [Google Scholar]
- Mizoguchi, T.; Wright, L.; Fujiwara, S.; Cremer, F.; Lee, K.; Onouchi, H.; Mouradov, A.; Fowler, S.; Kamada, H.; Putterill, J.; et al. Distinct roles of GIGANTEA in promoting flowering and regulating circadian rhythms in Arabidopsis. Plant Cell 2005, 17, 2255–2270. [Google Scholar] [CrossRef] [PubMed]
- David, K.M.; Armbruster, U.; Tama, N.; Putterill, J. Arabidopsis GIGANTEA protein is post-transcriptionally regulated by light and dark. FEBS Lett. 2006, 580, 1193–1197. [Google Scholar] [CrossRef] [PubMed]
- Max Planck Institute for Plant Breeding Research. Control of Flowering Time. Available online: http://www.mpipz.mpg.de/coupland (accessed on 8 September 2016).
- Valverde, F. CONSTANS and the evolutionary origin of photoperiodic timing of flowering. J. Exp. Bot. 2011, 62, 2453–2463. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Chang, P.; Ng, K.; Chang, C.; Sheu, P.; Tsai, J. A model comparison study of the flowering time regulatory network in Arabidopsis. BMC Syst. Biol. 2014, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; An, G. OsGI controls flowering time by modulating rhythmic flowering time regulators preferentially under short day in rice. J. Plant Biol. 2015, 58, 137–145. [Google Scholar] [CrossRef]
- Takahashi, Y.; Shomura, A.; Sasaki, T.; Yano, M. Hd6, a rice quantitative trait locus involved in photoperiod sensitivity, encodes the subunit of protein kinase CK2. Proc. Natl. Acad. Sci. USA 2001, 98, 7922–7927. [Google Scholar] [CrossRef]
- Nagano, H.; Clark, L.V.; Zhao, H.; Peng, J.; Yoo, J.H.; Heo, K.; Yu, C.Y.; Anzoua, K.G.; Matsuo, T.; Sacks, E.; et al. Contrasting allelic distribution of CO/Hd1 homologues in Miscanthus sinensis from the East Asian mainland and the Japanese archipelago. J. Exp. Bot. 2015, 66, 4227–4237. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Chen, D.; Fang, J.; Wang, P.; Deng, X.; Chu, C. Understanding the genetic and epigenetic architecture in complex network of rice flowering pathways. Protein Cell 2014, 5, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Izawa, T.; Fuse, T.; Yamanouchi, U.; Kubo, T.; Shimatani, Z.; Yano, M.; Yoshimura, A. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes Dev. 2004, 18, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.H.; Ito, S.; Imaizumi, T. Similarities in the circadian clock and photoperiodism in plants. Curr. Opin. Plant Biol. 2010, 13, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Nonoue, Y.; Yano, M.; Izawa, T. A pair of floral regulators sets critical day length for Hd3a florigen expression in rice. Nat. Genet. 2010, 42, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, H.; Taoka, K.; Shimamoto, K. Regulation of flowering in rice: Two florigen genes, a complex gene network, and natural variation. Curr. Opin. Plant Biol. 2011, 14, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, H.; Taoka, K. Florigen signaling. In Signaling Pathways in Plants; The Enzymes, Machida, Y., Lin, C., Tamanoi, F., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; Volume 35, pp. 130–135. [Google Scholar]
- Xue, W.; Xing, Y.; Weng, X.; Zhao, Y.; Tang, W.; Wang, L.; Zhou, H.; Yu, S.; Xu, C.; Li, X.; et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet. 2008, 40, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.F.; Jensen, E.; Alexandrov, N.; Troukhan, M.; Zhang, L.; Thomas-Jones, S.; Farrar, K.; Clifton-Brown, J.; Donnison, I.; Swaller, T.; et al. High resolution genetic mapping by genome sequencing reveals genome duplication and tetraploid genetic structure of the diploid Miscanthus sinensis. PLoS ONE 2012, 7, e33821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fe, D.; Cericola, F.; Byrne, S.; Lenk, I.; Asraf, B.H.; Pedersen, M.G.; Roulund, N.; Asp, T.; Janss, L.; Jensen, C.S.; et al. Genomic dissection and prediction of heading date in perennial ryegrass. BMC Genom. 2015, 16, 921. [Google Scholar] [CrossRef] [PubMed]
- Skøt, L.; Sanderson, R.; Thomas, A.; Skøt, K.; Thorogood, D.; Latypova, G.; Asp, T.; Armstead, I. Allelic variation in the perennial ryegrass FLOWERING LOCUS T gene is associated with changes in flowering time across a range of populations. Plant Physiol. 2011, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Shinozuka, H.; Cogan, N.O.; Spangenberg, G.C.; Forster, J.W. Quantitative Trait Locus (QTL) meta-analysis and comparative genomics for candidate gene prediction in perennial ryegrass (Lolium perenne L.). BMC Genet. 2012, 8, 101. [Google Scholar] [CrossRef] [PubMed]
- Armstead, I.P.; Turner, L.B.; Marshall, A.H.; Humphreys, M.O.; King, I.P.; Thorogood, D. Identifying genetic components controlling fertility in the outcrossing grass species perennial ryegrass (Lolium perenne) by quantitative trait loci analysis and comparative genetics. New Phytol. 2008, 178, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Jones, E.S.; Cogan, N.O.I.; Vecchies, A.C.; Nomura, T.; Hisano, H.; Shimamoto, Y.; Smith, K.F.; Hayward, M.D.; Forster, J.W. QTL analysis of morphological, developmental, and winter hardiness associated traits in perennial ryegrass. Crop Sci. 2004, 44, 925–935. [Google Scholar] [CrossRef]
- Yang, S.; Murphy, R.L.; Morishige, D.T.; Klein, P.E.; Rooney, W.L.; Mullet, J.E. Sorghum phytochrome B inhibits flowering in long days by activating expression of SbPRR37 and SbGHD7, repressors of SbEHD1, SbCN8 and SbCN12. PLoS ONE 2014, 9, e105352. [Google Scholar] [CrossRef] [PubMed]
- El Mannai, Y.; Shehzad, T.; Okuno, K. Variation in flowering time in sorghum core collection and mapping of QTLs controlling flowering time by association analysis. Genet. Resour. Crop Evol. 2011, 58, 983–989. [Google Scholar]
- Cuevas, H.E.; Zhou, C.; Tang, H.; Khadke, P.P.; Das, S.; Lin, Y.; Ge, Z.; Clemente, T.; Upadhyaya, H.D.; Hash, T.C.; et al. The evolution of photoperiod-insensitive flowering in sorghum, a genomic model for panicoid grasses. Mol. Biol. Evol. 2016. [Google Scholar] [CrossRef]
- Atienza, S.G.; Ramirez, M.C.; Martin, A. Mapping QTLs controlling flowering date in Miscanthus sinensis Anderss. Cereal Res. Commun. 2003, 31, 265–271. [Google Scholar]
- Hajdu, A.; Ádám, É.; Sheerin, D.J.; Dobos, O.; Bernula, P.; Hiltbrunner, A.; Kozma-Bognar, L.; Nagy, F. High-level expression and phosphorylation of phytochrome B modulates flowering time in Arabidopsis. Plant J. 2015, 83, 794–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izawa, T.; Mihara, M.; Suzuki, Y.; Gupta, M.; Itoh, H.; Nagano, A.J.; Motoyama, R.; Sawada, Y.; Yano, M.; Hirai, M.Y.; et al. Os-GIGANTEA confers robust diurnal rhythms on the global transcriptome of rice in the field. Plant Cell 2011, 23, 1741–1755. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Nonoue, Y.; Ono, N.; Shibaya, T.; Ebana, K.; Matsubara, K.; Ogiso-Tanaka, E.; Tanabata, T.; Sugimoto, K.; Taguchi-Shiobara, F.; et al. Genetic architecture of variation in heading date among Asian rice accessions. BMC Plant Biol. 2015, 15, 115. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, R.; Aoki, M.; Kurotani, K.; Yokoi, S.; Shinomura, T.; Takano, M.; Shimamoto, K. Phytochrome B regulates Heading date 1 (Hd1)-mediated expression of rice florigen Hd3a and critical day length in rice. Mol. Genet. Genom. 2011, 285, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, R.; Tamaki, S.; Yokoi, S.; Inagaki, N.; Shinomura, T.; Takano, M.; Shimamoto, K. Suppression of the floral activator Hd3a is the principal cause of the night break effect in rice. Plant Cell 2015, 17, 3326–3336. [Google Scholar] [CrossRef] [PubMed]
- Jarillo, J.A.; Del Olmo, I.; Gómez-Zambrano, A.; Lázaro, A.; López-González, L.; Miguel, E.; Narro-Diego, L.; Sáez, D.; Piñeiro, M. Photoperiodic control of flowering time. Span. J. Agric. Res. 2008, 6, 221–244. [Google Scholar] [CrossRef]
- Morishige, D.T.; Childs, K.L.; Moore, L.D.; Mullet, J.E. Targeted analysis of orthologous phytochrome A regions of the sorghum, maize, and rice genomes using gene-island sequencing. Plant Physiol. 2002, 130, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- Mouradov, A.; Cremer, F.; Coupland, G. Control of flowering time: Interacting pathways as a basis for diversity. Plant Cell 2002, S111–S130. [Google Scholar]
- Naranjo, L.; Talon, M.; Domingo, C. Diversity of floral regulatory genes of japonica rice. BMC Genom. 2014, 15, 101. [Google Scholar] [CrossRef] [PubMed]
- Ogiso, E.; Takahashi, Y.; Sasaki, T.; Yano, M.; Izawa, T. The role of casein kinase II in flowering time regulation has diversified during evolution. Plant Physiol. 2010, 152, 808–820. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, R.; Gómez-Ariza, J.; Brambilla, V.; Fornara, F. Molecular control of seasonal flowering in rice, arabidopsis and temperate cereals. Ann. Bot. 2014, 114, 1445–1458. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Li, C.; Hu, W.; Lau, M.Y.; Lin, H.; Rockwell, N.C.; Martin, S.S.; Jernstedt, J.A.; Lagarias, J.C.; Dubcovsky, J. PHYTOCHROME C plays a major role in the acceleration of wheat flowering under long-day photoperiod. Proc. Natl. Acad. Sci. USA 2014, 111, 10037–10044. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, K.; Chae, W.B.; Mitros, T.; Varala, K.; Xie, L.; Barling, A.; Glowacka, K.; Hall, M.; Jezowski, S.; Ming, R.; et al. A framework genetic map for Miscanthus sinensis from RNAseq-based markers shows recent tetraploidy. BMC Genom. 2012, 13, 142. [Google Scholar] [CrossRef] [PubMed]
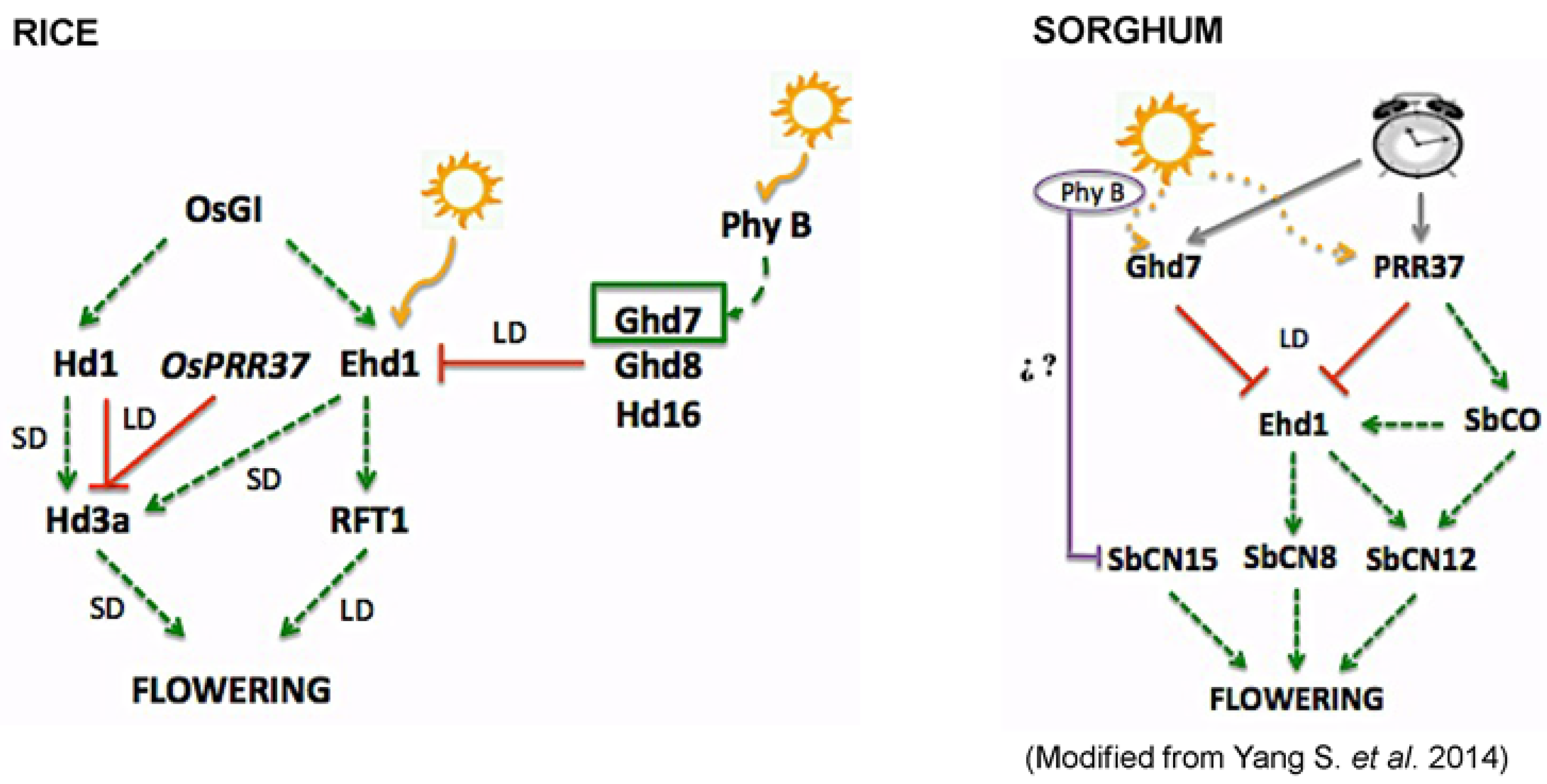
Figure 1.
A simplified model of flowering time under short-day (SD) and long-day (LD) conditions in rice and sorghum. A dashed green arrow indicates transcriptional activation and a solid red line indicates transcriptional repression.
Figure 1.
A simplified model of flowering time under short-day (SD) and long-day (LD) conditions in rice and sorghum. A dashed green arrow indicates transcriptional activation and a solid red line indicates transcriptional repression.

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nuñez, F.D.B.; Yamada, T. Molecular Regulation of Flowering Time in Grasses. Agronomy 2017, 7, 17. https://doi.org/10.3390/agronomy7010017
AMA Style
Nuñez FDB, Yamada T. Molecular Regulation of Flowering Time in Grasses. Agronomy. 2017; 7(1):17. https://doi.org/10.3390/agronomy7010017
Chicago/Turabian StyleNuñez, Fiorella D. B., and Toshihiko Yamada. 2017. "Molecular Regulation of Flowering Time in Grasses" Agronomy 7, no. 1: 17. https://doi.org/10.3390/agronomy7010017
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.