Screening of EMS-Induced Drought-Tolerant Sugarcane Mutants Employing Physiological, Molecular and Enzymatic Approaches


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Source and In Vitro Callus Regeneration
2.2. Measurement of Optimal EMS Concentration and Mutagenic Treatment
2.3. Measurement of Optimal In Vitro PEG-6000.Stimulated Osmotic Stress
2.4. Physiological Index Detection
2.4.1. Chlorophyll a
2.4.2. Leaf Chlorophyll Content
2.4.3. Photosynthetic Rate
2.5. DNA Isolation
2.6. SSR Marker PCR Amplification and CELL-1 Enzyme Digestion
3. Results
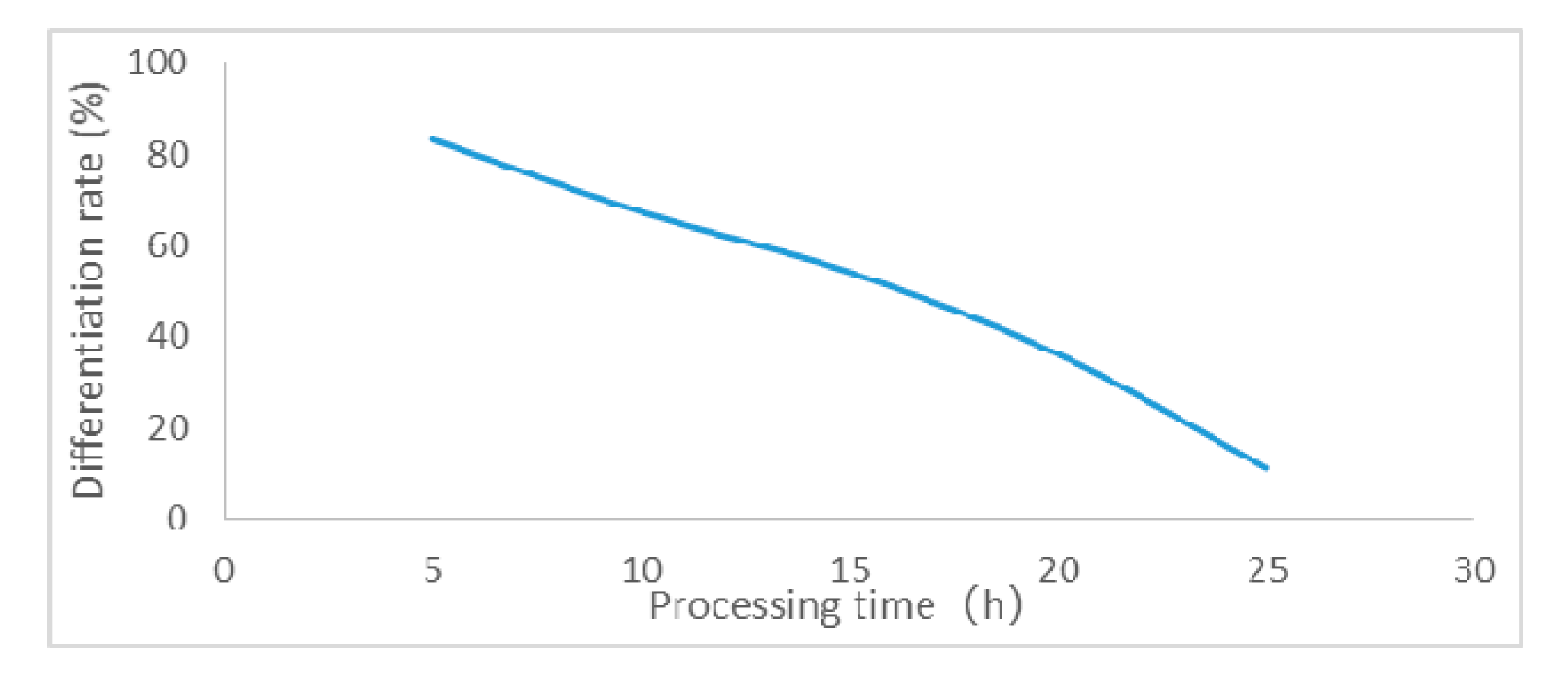
3.1. Effect of Various Concentrations of EMS on Sugarcane Calli Differentiation
3.2. Effect of Various Concentrations of PEG-6000-Stimulated Stress on Seedling Growth
3.3. Physiological Analysis
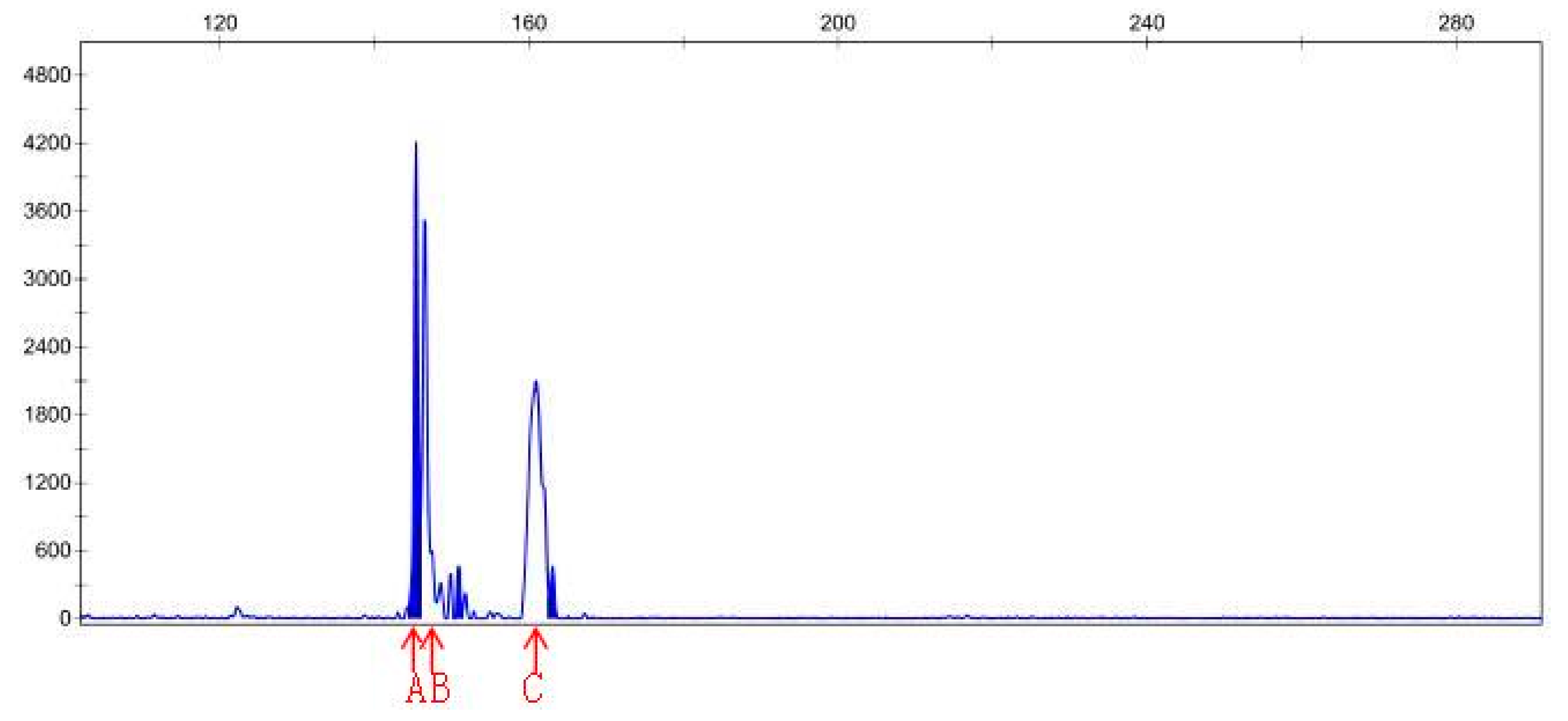
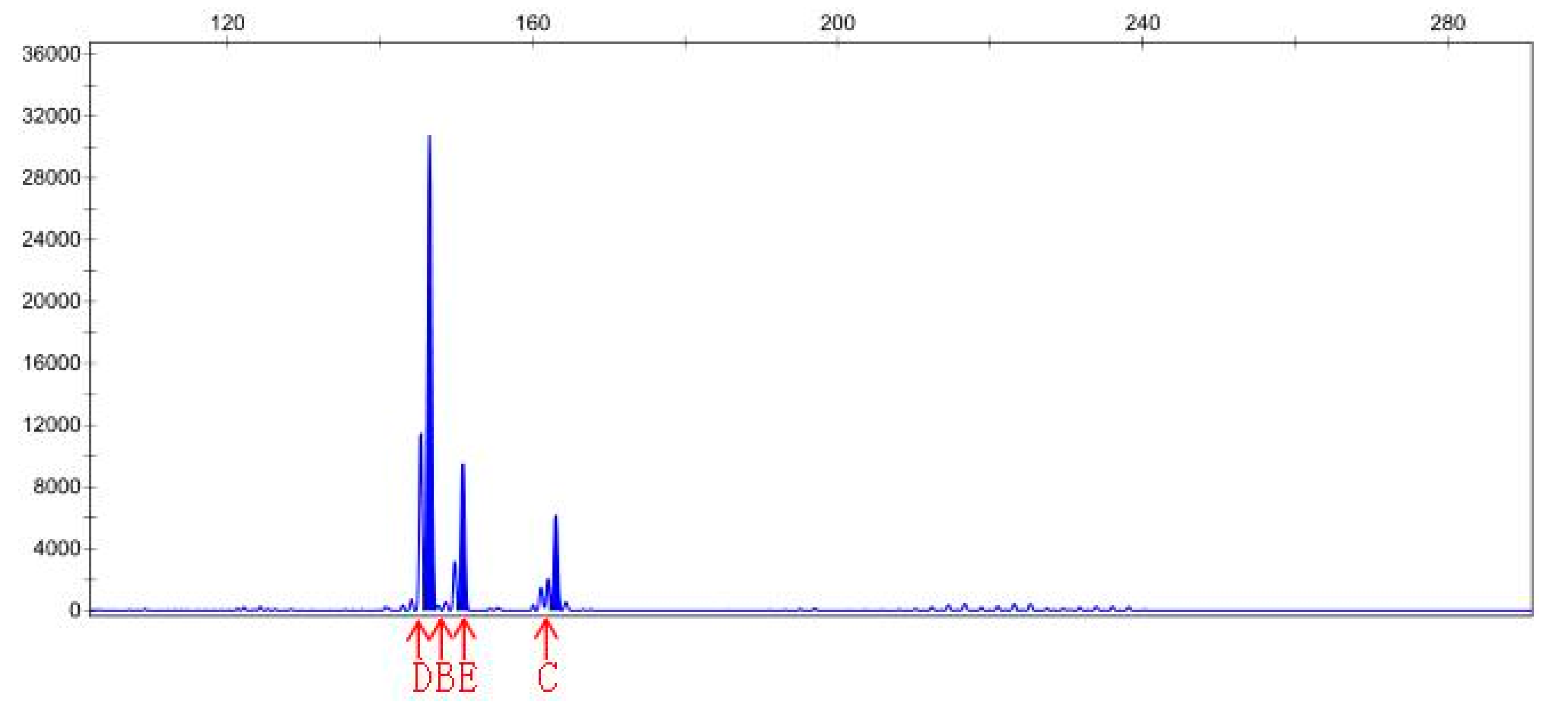
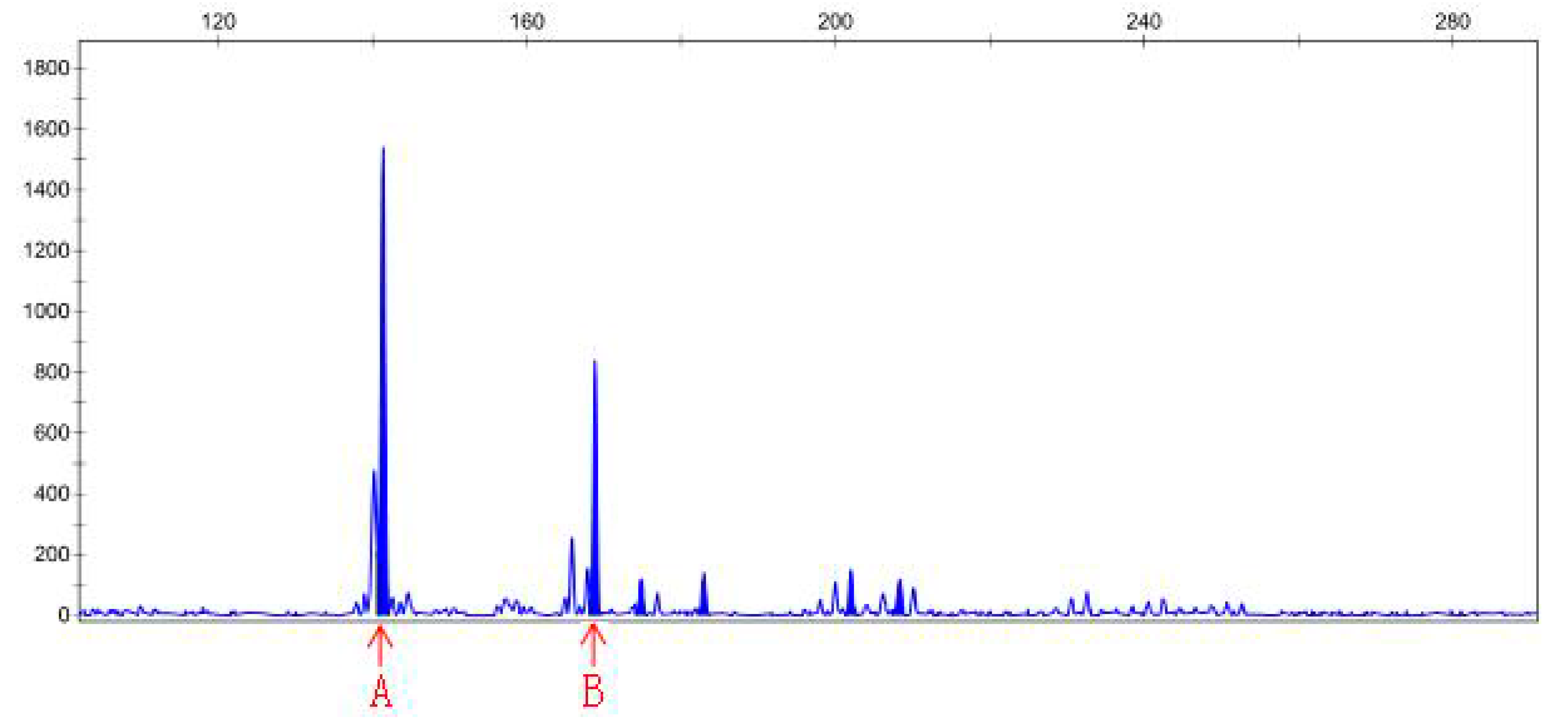
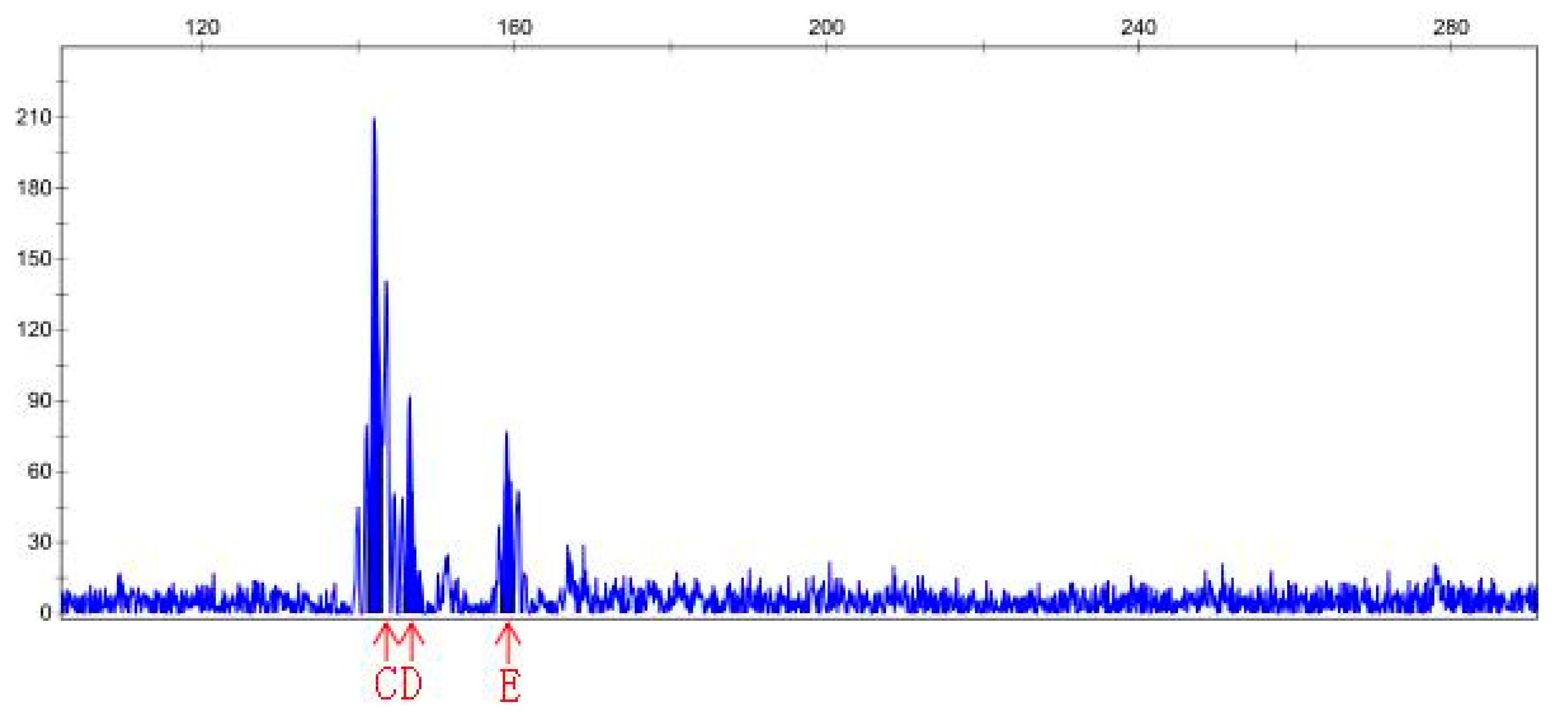
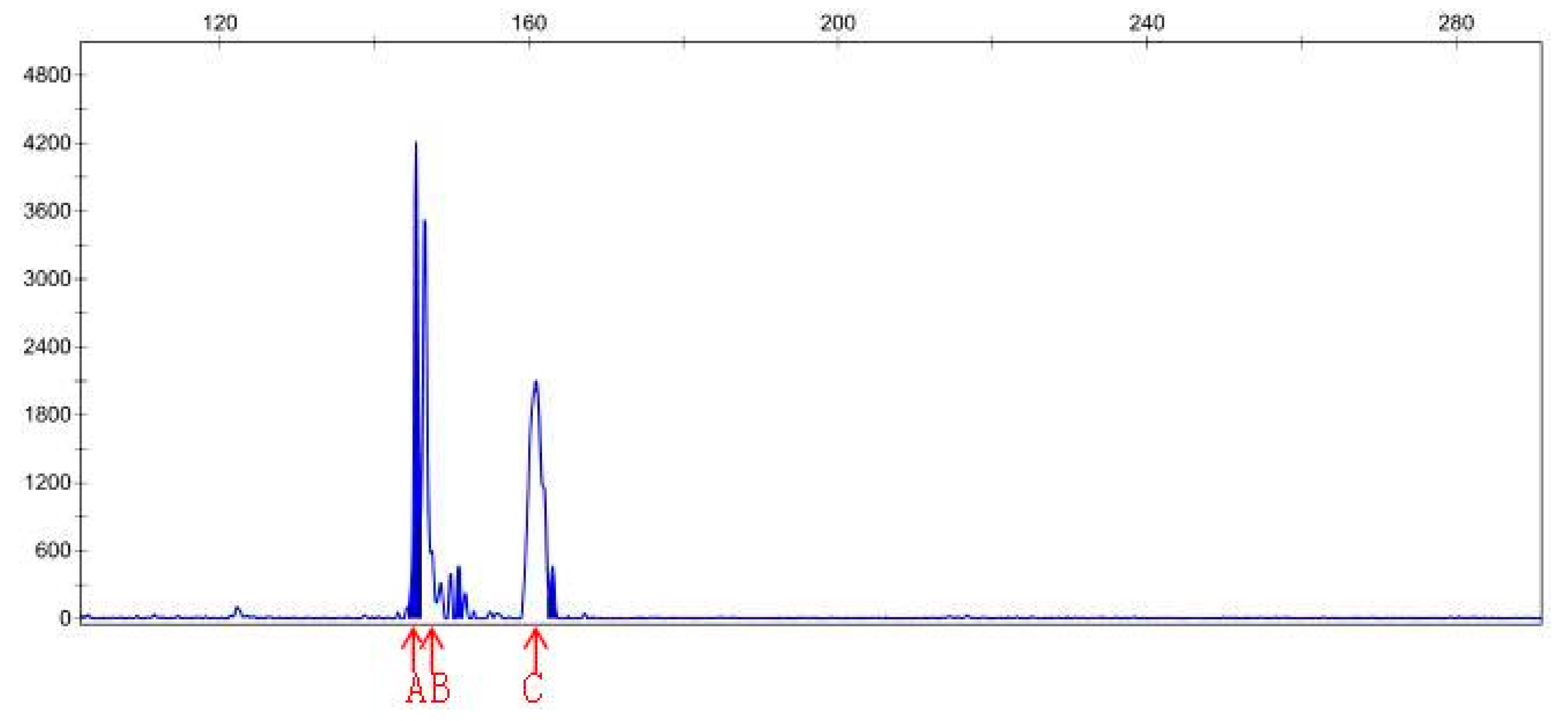
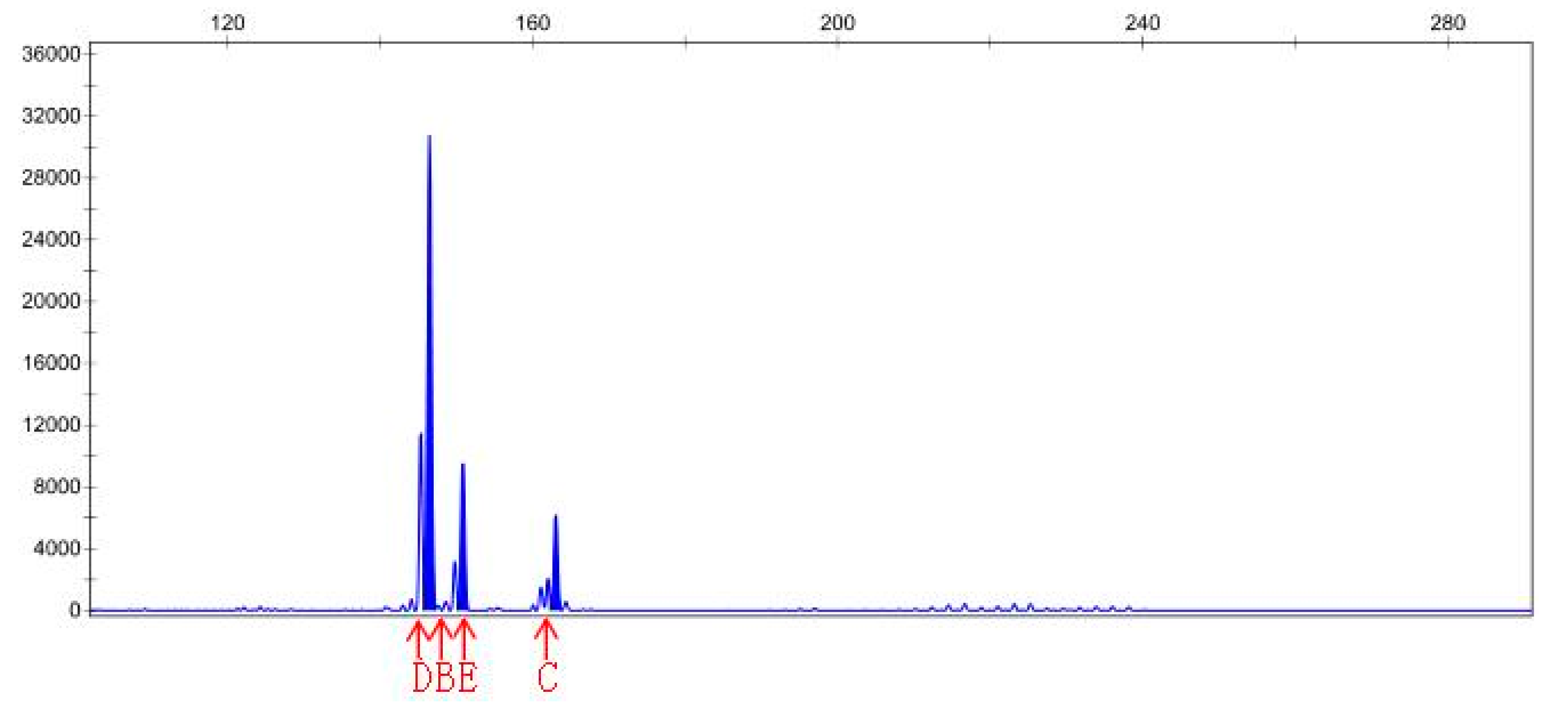
3.4. SSR Analysis
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Gentile, A.; Ferreira, T.H.; Mattos, R.S.; Dias, L.I.; Hoshino, A.A.; Carneiro, M.S.; Souza, G.M.; Calsa, T.; Nogueira, R.M.; Endres, L.; et al. Effects of drought on the microtranscriptome of field-grown sugarcane plants. Planta 2013, 237, 783–798. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, T.H.S.; Tsunada, M.S.; Bassi, D.; Araújo, P.; Mattiello, L.; Guidelli, G.V.; Righetto, G.L.; Gonçalves, V.R.; Lakshmanan, P.; Menossi, M. Sugarcane Water Stress Tolerance Mechanisms and Its Implications on Developing Biotechnology Solutions. Front. Plant Sci. 2017, 8, 1077. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-R.R.; Yang, L.-T.T. Sugarcane agriculture and sugar industry in China. Sugar Tech 2015, 17, 1–8. [Google Scholar] [CrossRef]
- Guo, J.; Ling, H.; Wu, Q.; Xu, L.; Que, Y. The choice of reference genes for assessing gene expression in sugarcane under salinity and drought stresses. Sci. Rep. 2014, 4, 7042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, R.R.; Andrade Dias Brito da Cunha, B.; Martins, P.K.; Martins, M.T.B.; Alekcevetch, J.C.; Chalfun-Júnior, A.Ô.; Andrade, A.C.; Ribeiro, A.P.; Qin, F.; Mizoi, J.; et al. Induced over-expression of AtDREB2A CA improves drought tolerance in sugarcane. Plant Sci. 2014, 221–222, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, P. Effect of different levels of drought during the formative phase on growth parameters and its relationship with dry matter accumulation in sugarcane. J. Agron. Crop Sci. 2000, 185, 83–89. [Google Scholar] [CrossRef]
- Ribeiro, R.V.; MacHado, R.S.; MacHado, E.C.; MacHado, D.F.S.P.; Magalhães Filho, J.R.; Landell, M.G.A. Revealing drought-resistance and productive patterns in sugarcane genotypes by evaluating both physiological responses and stalk yield. Exp. Agric. 2013, 49, 212–224. [Google Scholar] [CrossRef]
- Sales, C.R.G.; Ribeiro, R.V.; Silveira, J.A.G.; Machado, E.C.; Martins, M.O.; Lagôa, A.M.M.A. Superoxide dismutase and ascorbate peroxidase improve the recovery of photosynthesis in sugarcane plants subjected to water deficit and low substrate temperature. Plant Physiol. Biochem. 2013, 73, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.J.; Zhang, K.K.; Du, C.Z.; Li, J.; Xing, Y.X.; Yang, L.T.; Li, Y.R. Effect of Drought Stress on Anatomical Structure and Chloroplast Ultrastructure in Leaves of Sugarcane. Sugar Tech 2015, 17, 41–48. [Google Scholar] [CrossRef]
- Lopes, M.S.; Araus, J.L.; van Heerden, P.D.R.; Foyer, C.H. Enhancing drought tolerance in C4 crops. J. Exp. Bot. 2011, 62, 3135–3153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tayyab, M.; Islam, W.; Khalil, F.; Ziqin, P.; Caifang, Z.; Arafat, Y.; Hui, L.; Rizwan, M.; Ahmad, K.; Waheed, S.; et al. Biochar: An efficient way to manage low water availability in plants. Appl. Ecol. Environ. Res. 2018, 16, 2565–2583. [Google Scholar] [CrossRef]
- Khalil, F.; Rauf, S.; Monneveux, P.; Anwar, S.; Iqbal, Z. Genetic analysis of proline concentration under osmotic stress in sunflower (Helianthus annuus L.). Breed. Sci. 2016, 66, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Rauf, S.; Al-Khayri, J.M.; Zaharieva, M.; Monneveux, P.; Khalil, F. Breeding strategies to enhance drought tolerance in crops. Adv. Plant Breed. Strateg. Agron. Abiot. Biot. Stress Traits 2016, 2, 397–445. [Google Scholar] [CrossRef]
- Masoabi, M.; Lloyd, J.; Kossmann, J.; van der Vyver, C. Ethyl Methanesulfonate Mutagenesis and In Vitro Polyethylene Glycol Selection for Drought Tolerance in Sugarcane (Saccharum spp.). Sugar Tech 2018, 20, 50–59. [Google Scholar] [CrossRef]
- Jankowicz-Cieslak, J.; Till, B.J. Chemical Mutagenesis of Seed and Vegetatively Propagated Plants Using EMS. Curr. Protoc. Plant Biol. 2016, 1, 617–635. [Google Scholar]
- WEI, C.L.; Chang-lian, W.E.I. Analysis and countermeasures of the degradation status of sugarcane variety ROC22 in Guangxi. J. South. Agric. 2012, 43, 2113–2117. [Google Scholar]
- Agarwal, P.; Jaiswal, V.; Kumar, S.; Balyan, H.S.; Gupta, P.K. Chromosome mapping of four novel mutants in bread wheat (Triticum aestivum L.). Acta Physiol. Plant. 2015, 37, 66. [Google Scholar] [CrossRef]
- Rastogi, J.; Siddhant, P.B.; Sharma, B.L. Somaclonal Variation: A new dimension for sugarcane improvement. GERF Bull. Biosci. 2015, 6, 5–10. [Google Scholar]
- Mallick, M.; Bharadwaj, C.; Srivastav, M.; Sharma, N.; Awasthi, O.P. Molecular characterization of Kinnow mandarin clones and mutants using cross genera SSR markers. Indian J. Biotechnol. 2017, 16, 244–249. [Google Scholar]
- Nybom, H.; Weising, K.; Rotter, B. DNA fingerprinting in botany: Past, present, future. Investig. Genet. 2014, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Ul Haq, S.; Kumar, P.; Singh, R.K.; Verma, K.S.; Bhatt, R.; Sharma, M.; Kachhwaha, S.; Kothari, S.L. Assessment of Functional EST-SSR Markers (Sugarcane) in Cross-Species Transferability, Genetic Diversity among Poaceae Plants, and Bulk Segregation Analysis. Genet. Res. Int. 2016, 2016, 7052323. [Google Scholar] [CrossRef] [PubMed]
- Khanal, S.; Schwartz, B.M.; Kim, C.; Adhikari, J.; Rainville, L.K.; Auckland, S.A.; Paterson, A.H. Cross-taxon application of sugarcane EST-SSR to genetic diversity analysis of bermudagrass (Cynodon spp.). Genet. Resour. Crop Evol. 2017, 64, 2059–2070. [Google Scholar] [CrossRef]
- Szurman-Zubrzycka, M.; Chmielewska, B.; Gajewska, P.; Szarejko, I. Mutation detection by analysis of DNA heteroduplexes in TILLING populations of diploid species. In Biotechnologies for Plant Mutation Breeding: Protocols; Jankowicz-Cieslak, J., Tai, T., Kumlehn, J., Till, B., Eds.; Springer: Cham, Switzerland, 2016; pp. 281–303. ISBN 9783319450216. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Van der Weele, C.M.; Spollen, W.G.; Sharp, R.E.; Baskin, T.I. Growth of Arabidopsis thaliana seedlings under water deficit studied by control of water potential in nutrient-agar media. J. Exp. Bot. 2000, 51, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.D.A.; Jifon, J.L.; Da Silva, J.A.G.; Sharma, V. Use of physiological parameters as fast tools to screen for drought tolerance in sugarcane. Braz. J. Plant Physiol. 2007, 19, 193–201. [Google Scholar] [CrossRef]
- Jamil, S.; Shahzad, R.; Talha, G.M.; Sakhawat, G.; Sajid-Ur-Rahman; Sultana, R.; Iqbal, M.Z. Optimization of Protocols for in Vitro Regeneration of Sugarcane (Saccharum officinarum). Int. J. Agron. 2017, 2017, 2089381. [Google Scholar] [CrossRef]
- Serrat, X.; Esteban, R.; Guibourt, N.; Moysset, L.; Nogués, S.; Lalanne, E. EMS mutagenesis in mature seed-derived rice calli as a new method for rapidly obtaining TILLING mutant populations. Plant Methods 2014, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Naz, S.; Alam, S.S.; Iqbal, J. In vitro induced mutation for screening of red rot (Colletotrichum falcatum) resistance in sugarcane (Saccharum officinarum). Pak. J. Bot. 2007, 39, 1979–1994. [Google Scholar]
- Kenganal, M.; Hanchinal, R.R.; Nadaf, H.L. Ethyl methanesulfonate (EMS) induced mutation and selection for salt tolerance in sugarcane in vitro. Indian J. Plant Physiol. 2008, 13, 405–410. [Google Scholar]
- Nikam, A.A.; Devarumath, R.M.; Shitole, M.G.; Ghole, V.S.; Tawar, P.N.; Suprasanna, P. Gamma radiation, in vitro selection for salt (NaCl) tolerance, and characterization of mutants in sugarcane (Saccharum officinarum L.). In Vitro Cell. Dev. Biol. Plant 2014, 50, 766–776. [Google Scholar] [CrossRef]
- Oloriz, M.I.; Gil, V.; Rojas, L.; Veitía, N.; Höfte, M.; Jiménez, E. Selection and characterisation of sugarcane mutants with improved resistance to brown rust obtained by induced mutation. Crop Pasture Sci. 2012, 62, 1037–1044. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.C.; Ramgareeb, S.; Rutherford, R.S.; Snyman, S.J.; Watt, M.P. An in vitro mutagenesis protocol for the production of sugarcane tolerant to the herbicide imazapyr. In Vitro Cell. Dev. Biol. Plant 2012, 48, 417–427. [Google Scholar] [CrossRef]
- De Almeida Silva, M.; Jifon, J.L.; Sharma, V.; da Silva, J.A.G.; Caputo, M.M.; Damaj, M.B.; Guimarães, E.R.; Ferro, M.I.T. Use of physiological parameters in screening drought tolerance in sugarcane genotypes. Sugar Tech 2011, 13, 191. [Google Scholar] [CrossRef]
- Da Graça, J.P.; Rodrigues, F.A.; Farias, J.R.B.; de Oliveira, M.C.N.; Hoffmann-Campo, C.B.; Zingaretti, S.M. Physiological parameters in sugarcane cultivars submitted to water deficit. Braz. J. Plant Physiol. 2010, 22, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Mathobo, R.; Marais, D.; Steyn, J.M. The effect of drought stress on yield, leaf gaseous exchange and chlorophyll fluorescence of dry beans (Phaseolus vulgaris L.). Agric. Water Manag. 2017, 180, 118–125. [Google Scholar] [CrossRef]
- Dinh, T.H.; Watanabe, K.; Takaragawa, H.; Nakabaru, M.; Kawamitsu, Y. Photosynthetic response and nitrogen use efficiency of sugarcane under drought stress conditions with different nitrogen application levels. Plant Prod. Sci. 2017, 20, 412–422. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | SSR Name | Repeat Motif | Tm (°C) | F-Primer | R-Primer | Result |
|---|---|---|---|---|---|---|
| 1 | SMC336BS | (TG)23(AG)19 | 62 | ATTCTAGTGCCAATCCATCTCA | CATGCCAACTTCCAAACAGAC | 5 |
| 2 | SMC569CS | (TG)37 | 62 | GCGATGGTTCCTATGCAACTT | TTCGTGGCTGAGATTCACACTA | 0 |
| 3 | SMC31CUQ | (TC)10(AC)22 | 62 | CATGCCAACTTCCAATACAGACT | AGTGCCAATCCATCTCAGAGA | 5 |
| 4 | SMC334BS | (TG)36 | 60 | CAATTCTGACCGTGCAAAGAT | CGATGAGCTTGATTGCGAATG | 5 |
| 5 | Sep-17 | (CCT)9 | 60 | ACCCTGCTGGTCTCCTCC | ACGTTCGACGTCGTGTAGTG | 5 |
| 6 | Sep-23 | (AGA)9 | 60 | GTGTTCAGGCAGATGGTCCT | GTCGATGGCACCGATTTATT | 5 |
| 7 | Sep-59 | (CGG)9 | 60 | TTCCATTTACTCCTCCGTGC | CTCCCCCTCCTCGTACTTGT | 5 |
| 8 | Sep-6 | (GCA)5(ACA)13 | 60 | CAGCCCATTAACCAAGCAAT | GAAGCAGCTGTTGCTCACTG | 5 |
| 9 | Sep-70 | (GCG)8 | 60 | AACTCACCCAACAAAGCGAC | AGACGAAGAGCTCGTGGATG | |
| 10 | Sep-8 | (CT)17 | 60 | CTTGCTTCCCCTTTACTCCC | GAGGCGCCTTACTGTTCTTG | 5 |
| 11 | Sep-84 | (GGT)9 | 60 | AGAGACCGTAATGGTGACCG | ACCACCACCACCACCATACT | 5 |
| 12 | Sep-89 | (CGT)10 | 60 | AGCTCTGATTTTTGGGGGTT | GGAAGACAGTGGACGAGGTC | 5 |
| 13 | SMC286CS | (TG)43 | 58 | TCAAATGGGACCTTATTGGAG | TCCCTCGATCTCCGTTGTT | 5 |
| 14 | SMC119CG | (TTG)12 | 58 | TTCATCTCTAGCCTACCCCAA | AGCAGCCATTTACCCAGGA | 5 |
| 15 | mSSCIR43 | (GT)3(AT)2(GT)29 | 52 | ATTCAACGATTTTCACGAG | AACCTAGCAATTTACAAGAG | 0 |
| ROC22 | FN39 | ||||||
|---|---|---|---|---|---|---|---|
| Treatment Time (h) | Treatment Concentration (%) | Number of Germinated Calli | Relative Differentiation Rate (%) | Treatment Time (h) | Treatment Concentration (%) | Number of Germinated Calli | Relative Differentiation Rate (%) |
| 5 | 0 | 84 | 100 | 5 | 0 | 85 | 100 |
| 0.05 | 80 | 95.2 | 0.05 | 81 | 95.2 | ||
| 0.1 | 70 | 83.3 | 0.1 | 70 | 84.7 | ||
| 0.2 | 48 | 57.1 | 0.2 | 49 | 57.6 | ||
| 0.5 | 30 | 35.7 | 0.5 | 31 | 36.4 | ||
| 10 | 0 | 83 | 100 | 10 | 0 | 84 | 100 |
| 0.05 | 78 | 93.9 | 0.05 | 77 | 91.6 | ||
| 0.1 | 56 | 67.4 | 0.1 | 53 | 63.0 | ||
| 0.2 | 34 | 40.9 | 0.2 | 28 | 33.3 | ||
| 0.5 | 9 | 10.8 | 0.5 | 5 | 5.9 | ||
| 15 | 0 | 83 | 100 | 15 | 0 | 82 | 100 |
| 0.05 | 76 | 91.5 | 0.05 | 70 | 85.3 | ||
| 0.1 | 45 | 54.2 | 0.1 | 39 | 47.5 | ||
| 0.2 | 21 | 25.3 | 0.2 | 4 | 4.8 | ||
| 0.5 | 4 | 4.8 | 0.5 | 0 | 0 | ||
| 20 | 0 | 80 | 100 | 20 | 0 | 83 | 100 |
| 0.05 | 69 | 86.2 | 0.05 | 66 | 79.5 | ||
| 0.1 | 29 | 36.2 | 0.1 | 24 | 28.9 | ||
| 0.2 | 3 | 3.7 | 0.2 | 0 | 0 | ||
| 0.5 | 0 | 0 | 0.5 | 0 | 0 | ||
| 25 | 0 | 81 | 100 | 25 | 0 | 79 | 100 |
| 0.05 | 66 | 81.4 | 0.05 | 60 | 75.9 | ||
| 0.1 | 9 | 11.1 | 0.1 | 6 | 7.5 | ||
| 0.2 | 0 | 0 | 0.2 | 0 | 0 | ||
| 0.5 | 0 | 0 | 0.5 | 0 | 0 | ||
| Genotypes | Fv/Fm | Diff b/w Chl Content Before and After Trt | Photosynthetic Rate |
|---|---|---|---|
| ROC22 | 0.77 | 0.22 | 4.89 |
| MR22-1 | 0.75 | 0.15 | 3.88 |
| MR22-2 | 0.75 | 0.43 | 7.84 |
| MR22-3 | 0.72 | 0.16 | 5.05 |
| MR22-4 | 0.76 | −0.08 | 4.55 |
| MR22-5 | 0.74 | −0.22 | 2.45 |
| MR22-6 | 0.76 | 0.28 | 7.36 |
| MR22-7 | 0.78 | 0.27 | 0.95 |
| MR22-8 | 0.76 | 0.43 | 4.31 |
| MR22-9 | 0.78 | 0.13 | 2.16 |
| MR22-10 | 0.76 | −0.07 | 7.13 |
| MR22-11 | 0.77 | 0.26 | 0.52 |
| MR22-12 | 0.75 | 0.01 | 2.98 |
| MR22-13 | 0.76 | 0.61 | 4.05 |
| MR22-15 | 0.69 | −0.04 | 10.65 |
| MR22-17 | 0.75 | 0.19 | 2.39 |
| MR22-20 | 0.73 | −0.14 | 12.13 |
| MR22-25 | 0.74 | 0.04 | 7.70 |
| FN 39 | 0.77 | 0.39 | 3.38 |
| FN39-1 | 0.76 | 0.55 | 3.26 |
| FN39-2 | 0.74 | 0.52 | 2.85 |
| Genotypes | Average | 5% Sig Level | 1% Sig Level |
|---|---|---|---|
| ROC 22 | 0.77 | a | a |
| MR22-15 | 0.69 | bc | bc |
| MR22-20 | 0.73 | c | c |
| Genotypes | Average | 5% Sig Level | 1% Sig Level |
|---|---|---|---|
| ROC22 | 4.89 | a | a |
| MR22-2 | 7.8367 | b | a |
| MR22-7 | 0.9489 | b | b |
| MR22-9 | 2.1615 | b | a |
| MR22-11 | 0.5202 | b | b |
| MR22-15 | 12.1331 | b | bc |
| MR22-20 | 10.6471 | b | bc |
| Sample File Name | Size | Height | Sample File Name | Size | Height |
|---|---|---|---|---|---|
| Sep-23-15P | 141.38 | 1542 | Sep-23-20P | 145.41 | 4215 |
| Sep-23-15P | 168.83 | 841 | Sep-23-20P | 150.9 | 466 |
| Sep-23-15P | 174.83 | 123 | Sep-23-20P | 163.05 | 465 |
| Sep-23-15P | 182.96 | 142 | Sep-23-20N | 146.48 | 30783 |
| Sep-23-15P | 201.94 | 150 | Sep-23-20N | 150.9 | 9531 |
| Sep-23-15P | 208.33 | 119 | Sep-23-20N | 163.06 | 6201 |
| Sep-23-15N | 142.03 | 210 | |||
| Sep-23-15N | 146.62 | 92 | |||
| Sep-23-15N | 159.06 | 77 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalil, F.; Naiyan, X.; Tayyab, M.; Pinghua, C. Screening of EMS-Induced Drought-Tolerant Sugarcane Mutants Employing Physiological, Molecular and Enzymatic Approaches. Agronomy 2018, 8, 226. https://doi.org/10.3390/agronomy8100226
Khalil F, Naiyan X, Tayyab M, Pinghua C. Screening of EMS-Induced Drought-Tolerant Sugarcane Mutants Employing Physiological, Molecular and Enzymatic Approaches. Agronomy. 2018; 8(10):226. https://doi.org/10.3390/agronomy8100226
Chicago/Turabian StyleKhalil, Farghama, Xiao Naiyan, Muhammad Tayyab, and Chen Pinghua. 2018. "Screening of EMS-Induced Drought-Tolerant Sugarcane Mutants Employing Physiological, Molecular and Enzymatic Approaches" Agronomy 8, no. 10: 226. https://doi.org/10.3390/agronomy8100226
APA StyleKhalil, F., Naiyan, X., Tayyab, M., & Pinghua, C. (2018). Screening of EMS-Induced Drought-Tolerant Sugarcane Mutants Employing Physiological, Molecular and Enzymatic Approaches. Agronomy, 8(10), 226. https://doi.org/10.3390/agronomy8100226