The Effect of Barley Cover Crop Residue and Herbicide Management on the Foliar Arthropod Community in No-Till Soybeans


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site, Layout, and Treatments
2.2. Plant Biomass
2.3. Arthropod Sampling
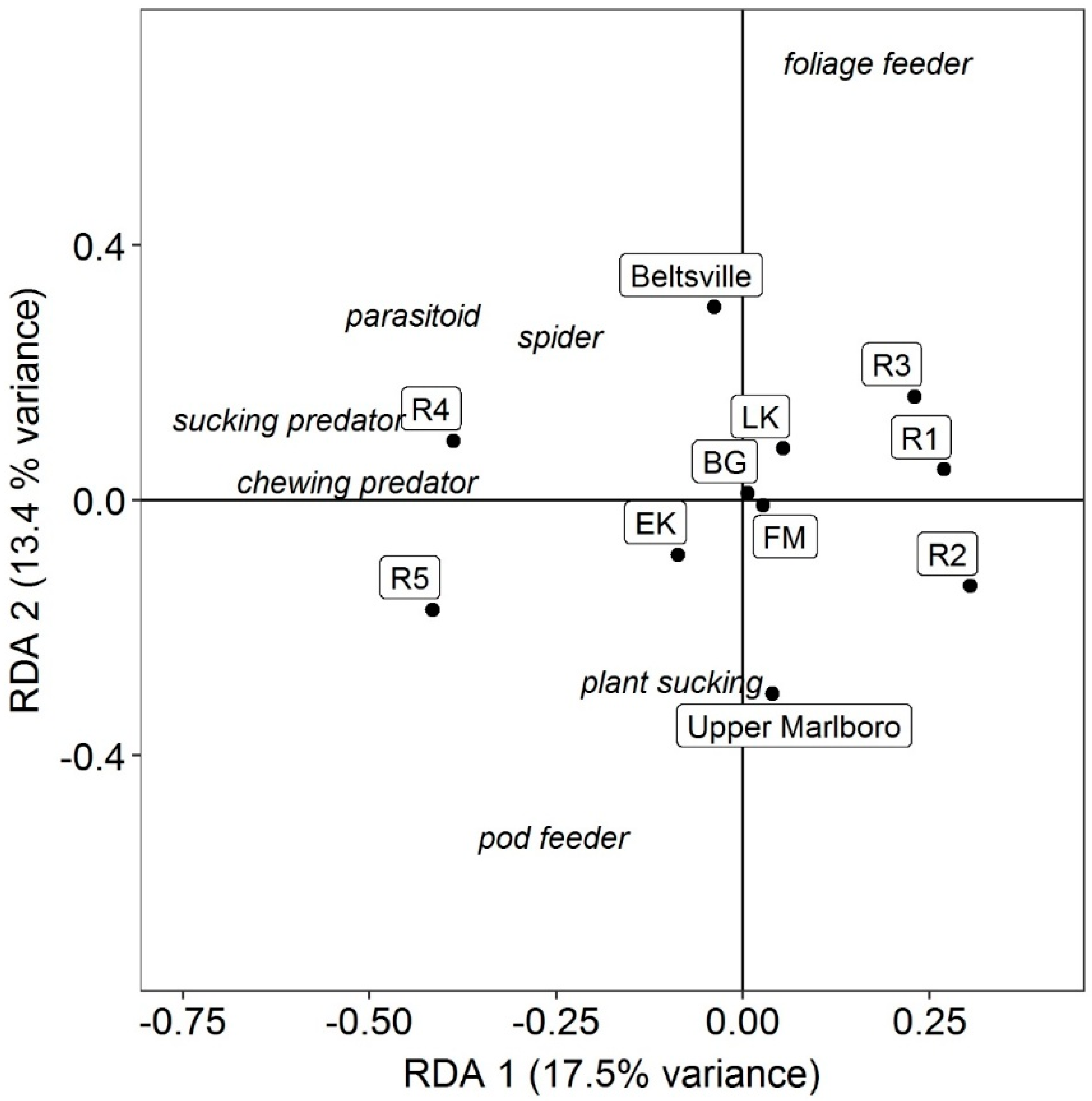
2.4. Statistical Analyses
3. Results
3.1. Cover Crop Biomass
3.2. Arthropod Counts
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Unger, P.W.; Vigil, M.F. Cover crop effects on soil water relationships. J. Soil Water Conserv. 1998, 53, 200–207. [Google Scholar]
- Price, A.J.; Arriaga, F.J.; Raper, R.L.; Balkcom, K.S.; Komecki, T.S.; Reeves, D.W. Comparison of mechanical and chemical winter cereal cover crop termination systems and cotton yield in conservation agriculture. J. Cotton Sci. 2009, 13, 238–245. [Google Scholar]
- Liebman, M.; Dyck, E. Crop-rotation and intercropping strategies for weed management. Ecol. Appl. 1993, 3, 92–122. [Google Scholar] [CrossRef] [PubMed]
- Hooks, C.R.R.; Hinds, J.; Zobel, E.; Patton, T. Impact of crimson clover dying mulch on two eggplant insect herbivores. J. Appl. Entomol. 2013, 137, 170–180. [Google Scholar] [CrossRef]
- Brainard, D.C.; Bellinder, R.R. Weed suppression in a broccoli–winter rye intercropping system. Weed Sci. 2004, 52, 281–290. [Google Scholar] [CrossRef]
- Morton, T.A.; Bergtold, J.S.; Price, A.J. The economics of cover crop biomass for corn and cotton. In Proceedings of the Southern Conservation Systems Conference, Amarillo, TX, USA, 26–28 June 2006; pp. 69–76. [Google Scholar]
- Teasdale, J.R. Interaction of light, soil moisture, and temperature with weed suppression by hairy vetch residue. Weed Sci. 1993, 41, 46–51. [Google Scholar] [CrossRef]
- Creamer, N.G.; Bennett, M.A.; Stinner, B.R.; Cardina, J.; Regnier, E.E. Mechanisms of weed suppression in cover crop-based production systems. HortScience 1996, 31, 410–413. [Google Scholar]
- Putnam, A.R.; Defrank, J.; Barnes, J.P. Exploitation of allelopathy for weed control in annual and perennial cropping systems. J. Chem. Ecol. 1983, 9, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Mirsky, S.B.; Ryan, M.R.; Teasdale, J.R.; Curran, W.S.; Reberg-Horton, C.S.; Spargo, J.T.; Wells, M.S.; Keene, C.L.; Moyer, J.W. Overcoming weed management challenges in cover crop-based organic rotational no-till soybean production in the eastern United States. Weed Technol. 2013, 27, 193–203. [Google Scholar] [CrossRef]
- Mohler, C.L.; Teasdale, J.R. Response of weed emergence to rate of Vicia villosa Roth and Secale cereale L. residue. Weed Res. 1993, 33, 487–499. [Google Scholar] [CrossRef]
- Lawton, J.H.; Strong, D.R., Jr. Community patterns and competition in folivorous insects. Am. Nat. 1981, 118, 317–338. [Google Scholar] [CrossRef]
- Altieri, M.A. The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- Obermaier, E.; Heisswolf, A.; Poethke, J.; Randlkofer, B.; Meiners, T. Plant architecture and vegetation structure: Two ways for insect herbivores to escape parasitism. Eur. J. Entomol. 2008, 105, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Mulvaney, M.J.; Price, A.J.; Wood, C.W. Cover crop residue and organic mulches provide weed control during limited-input no-till collard production. J. Sustain. Agric. 2011, 35, 312–328. [Google Scholar] [CrossRef]
- Bryant, A.; Brainard, D.C.; Haramoto, E.R.; Szendrei, Z. Cover crop mulch and weed management influence arthropod communities in strip-tilled cabbage. Environ. Entomol. 2013, 42, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Bryant, A.; Coudron, T.; Brainard, D.; Szendrei, Z. Cover crop mulches influence biological control of the imported cabbageworm (Pieris rapae L., Lepidoptera: Pieridae) in cabbage. Biol. Control 2014, 73, 75–83. [Google Scholar] [CrossRef]
- Wortman, S.E.; Francis, C.A.; Bernards, M.L.; Drijber, R.A.; Lindquist, J.L. Optimizing cover crop benefits with diverse mixtures and an alternative termination method. Agron. J. 2012, 104, 1425–1435. [Google Scholar] [CrossRef]
- Koch, R.L.; Porter, P.M.; Harbur, M.M.; Abrahamson, M.D.; Wyckhuys, K.A.G.; Ragsdale, D.W.; Buckman, K.; Sezen, Z.; Heimpel, G.E. Response of soybean insects to an autumn-seeded rye cover crop. Environ. Entomol. 2012, 41, 750–760. [Google Scholar] [CrossRef]
- Olson, D.M.; Davis, R.F.; Brown, S.L.; Roberts, P.; Phatak, S.C. Cover crop, rye residue and in-furrow treatment effects on thrips. J. Appl. Entomol. 2006, 130, 302–308. [Google Scholar] [CrossRef]
- Smith, A.W.; Hammond, R.B.; Strinner, B.R. Influence of rye-cover crop management on soybean foliage arthropods. Environ. Entomol. 1988, 17, 109–114. [Google Scholar] [CrossRef]
- Lundgren, J.G.; Fergen, J.K. Enhancing predation of a subterranean insect pest: A conservation benefit of winter vegetation in agroecosystems. Appl. Soil Ecol. 2011, 51, 9–16. [Google Scholar] [CrossRef]
- Jabbour, R.; Pisani-Gareau, T.; Smith, R.G.; Mullen, C.; Barbercheck, M.E. Cover crop and tillage intensities alter ground-dwelling arthropod communities during the transition to organic production. Renew. Agric. Food Syst. 2015, 31, 361–374. [Google Scholar] [CrossRef]
- Blubaugh, C.K.; Kaplan, I. Tillage compromises weed seed predator activity across developmental stages. Biol. Control 2015, 81, 76–82. [Google Scholar] [CrossRef]
- Jackson, D.M.; Harrison, H.F. Effects of a killed-cover crop mulching system on sweetpotato production, soil pests, and insect predators in South Carolina. J. Econ. Entomol. 2008, 101, 1871–1880. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Ayuso, M.; Gabriel, J.L.; Quemada, M. The kill date as a management tool for cover cropping success. PLoS ONE 2014, 9, e109587. [Google Scholar] [CrossRef] [PubMed]
- Hance, E.; Harks, D.P. Agriculture in Maryland Summary for 2013; Maryland Department of Agriculture: Annapolis, MD, USA, 2014.
- Schaefer Center for Public Policy. Maryland Farmers’ Winter Cover Crop Participation: Report on a Mail Survey and Four Focus Groups of Maryland Farmers; Maryland Department of Agriculture: Annapolis, MD, USA, 2005.
- Creamer, N.G.; Dabney, S.M. Killing cover crops mechanically: Review of recent literature and assessment of new research results. Am. J. Altern. Agric. 2002, 17, 32–40. [Google Scholar]
- Buchanan, A.L.; Zobel, E.; Hinds, J.; Rosario-Lebron, A.; Hooks, C.R.R. Can row spacing influence arthropod communities in soybean? Implications for early and late planting. Environ. Entomol. 2015, 44, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Friendly, F.G.B.M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; et al. Vegan 2.4-6: Community ecology package. Available online: https://crantastic.org/packages/vegan/versions/67602 (accessed on 20 April 2018).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language Environment for Statistical Computing; R Found. Stat. Comput.: Vienna, Austria, 2016. [Google Scholar]
- Tremelling, M.J.; McSorley, R.; Gallaher, R.N. Effects of winter cover crops on the soil surface invertebrate community. Soil Crop Sci. Soc. Fla. Proc. 2002, 62, 77–82. [Google Scholar]
- Young, O.P.; Edwards, G.B. Spiders in United States field crops and their potential effect on crop pests. J. Arachnol. 1990, 18, 1–27. [Google Scholar] [CrossRef]
- Chen, L.L.; You, M.S.; Chen, S.B. Effects of cover crops on spider communities in tea plantations. Biol. Control 2011, 59, 326–335. [Google Scholar] [CrossRef]
- Sunderland, K.; Samu, F. Effects of agricultural diversification on the abundance, distribution, and pest control potential of spiders. Entomol. Exp. Appl. 2000, 95, 1–13. [Google Scholar]
- Dunbar, M.W.; Gassmann, A.J.; O’Neal, M.E. Limited impact of a fall-seeded, spring-terminated rye cover crop on beneficial arthropods. Environ. Entomol. 2017, 46, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Hooks, C.R.R.; Wang, K.-H.; Meyer, S.L.; Lekveishvili, M.; Hinds, J.; Zobel, E.; Rosario-Lebron, A.; Lee-Bullock, M. Impact of no-till cover cropping of Italian ryegrass on above and below ground faunal communities inhabiting a soybean field with emphasis on soybean cyst nematodes. J. Nematol. 2011, 43, 172–181. [Google Scholar] [PubMed]
- Leslie, A.W.; Wang, K.-H.; Meyer, S.L.F.; Marahatta, S.; Hooks, C.R.R. Influence of cover crops on arthropods, free-living nematodes, and yield in a succeeding no-till soybean crop. Appl. Soil Ecol. 2017, 117–118, 21–31. [Google Scholar] [CrossRef]
- Fox, A.F.; Kim, T.N.; Bahlai, C.A.; Woltz, J.M.; Gratton, C.; Landis, D.A. Cover crops have neutral effects on predator communities and biological control services in annual cellulosic bioenergy cropping systems. Agric. Ecosyst. Environ. 2016, 232, 101–109. [Google Scholar] [CrossRef]
- Davis, H.N.; Currie, R.S.; Klocke, N.L.; Buschman, L.L. Winter annual cover crop has only minor effects on major corn arthropod pests. J. Econ. Entomol. 2010, 103, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Laub, C.A.; Luna, J.M. Winter cover crop suppression practices and natural enemies of armyworm (Lepidoptera: Noctuidae) in no-till corn. Environ. Entomol. 1992, 21, 41–49. [Google Scholar] [CrossRef]
- Gill, H.K.; McSorley, R.; Branham, M. Effect of organic mulches on soil surface insects and other arthropods. Fla. Entomol. 2011, 94, 226–232. [Google Scholar] [CrossRef]
- House, G.J.; Alzugaray, M.D.R. Influence of cover cropping and no-tillage practices on community composition of soil arthropods in a North Carolina agroecosystem. Environ. Entomol. 1989, 18, 302–307. [Google Scholar] [CrossRef]


{kind=link}
| Year | Location | Treatment 1 | Barley Planting 2 | Herbicide Spray 3 | Soybean Planting | Late Herbicide | Harvest |
|---|---|---|---|---|---|---|---|
| 2013 | Beltsville | EK | 21 Sep | 15 Apr | 21 May | 11 Jul | 25 Oct |
| LK | 21 Sep | 21 May | 21 May | 11 Jul | 25 Oct | ||
| FM | 21 Sep | 21 May | 21 May | 11 Jul | 25 Oct | ||
| BG | - | 21 May | 21 May | 11 Jul | 25 Oct | ||
| Upper Marlboro | EK | 21 Sep | 16 Apr | 20 May | - | 8 Oct | |
| LK | 21 Sep | 20 May | 20 May | - | 8 Oct | ||
| FM | 21 Sep | 20 May | 20 May | - | 8 Oct | ||
| BG | - | 20 May | 20 May | - | 8 Oct | ||
| 2014 | Beltsville | EK | 24 Sep | 18 Apr | 27 May | 2 Jul | 24 Oct |
| LK | 24 Sep | 27 May | 27 May | 2 Jul | 24 Oct | ||
| FM | 24 Sep | 27 May | 27 May | 2 Jul | 24 Oct | ||
| BG | - | 27 May | 27 May | 2 Jul | 24 Oct | ||
| Upper Marlboro | EK | 24 Sep | 18 Apr | 27 May | - | 25 Nov | |
| LK | 24 Sep | 27 May | 27 May | - | 25 Nov | ||
| FM | 24 Sep | 27 May | 27 May | - | 25 Nov | ||
| BG | - | 27 May | 27 May | - | 25 Nov |
| Weeks after Planting | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Site | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
| 2013 | Beltsville | - | - | R1 | R2 | R3 | R3 | R3 | R3 | R4 | R5 |
| 2013 | Upper Marlboro | - | - | R1 | R2 | R3 | R3 | R3 | R3 | R4 | R5 |
| 2014 | Beltsville | R1 | R2 | R2 | R3 | R3 | R3 | R3 | R4 | R4 | R5 |
| 2014 | Upper Marlboro | R1 | R1 | R2 | R3 | R3 | R3 | R3 | R4 | R4 | R5 |
| Site | Treatment 1 | Mass ± SEM (kg ha−1) | |||
|---|---|---|---|---|---|
| Beltsville | EK | 160.1 | ± | 60.5 | cd 2 |
| LK | 2211.9 | ± | 83.2 | a | |
| FM | 2123.4 | ± | 112.9 | a | |
| BG | 896.0 | ± | 254.3 | bc | |
| Upper Marlboro | EK | 85.8 | ± | 21.2 | d |
| LK | 753.4 | ± | 100.9 | b | |
| FM | 851.8 | ± | 62.7 | b | |
| BG | 120.4 | ± | 27.2 | d | |
| Beltsville | Upper Marlboro | ||||
|---|---|---|---|---|---|
| Feeding Guild | Family | 2013 1 | 2014 | 2013 | 2014 |
| Spider | Salticidae | 25 | 47 | 24 | 48 |
| Araneidae | 5 | 69 | 0 | 37 | |
| Oxyopidae | 149 | 101 | 42 | 94 | |
| Thomisidae | 18 | 16 | 35 | 11 | |
| Lycosidae | 0 | 12 | 0 | 26 | |
| Clubionidae | 0 | 3 | 0 | 0 | |
| Ctenidae | 0 | 1 | 0 | 0 | |
| Tetragnathidae | 0 | 8 | 0 | 9 | |
| Linyphiidae | 0 | 4 | 0 | 2 | |
| Pholcidae | 0 | 1 | 0 | 0 | |
| Parasitoid | Platygastridae | 159 | 11 | 57 | 0 |
| unspecified 2 | 0 | 407 | 0 | 104 | |
| Sceleonidae | 10 | 23 | 1 | 15 | |
| Chalcididae | 0 | 2 | 0 | 3 | |
| Proctotrupidae | 0 | 1 | 0 | 2 | |
| Braconidae | 0 | 76 | 0 | 29 | |
| Eulophidae | 0 | 18 | 0 | 5 | |
| Ichneumonidae | 0 | 8 | 0 | 3 | |
| Tiphiidae | 0 | 178 | 0 | 25 | |
| Aphelinidae | 0 | 1 | 0 | 1 | |
| Encyrtidae | 0 | 1 | 0 | 0 | |
| Mymaridae | 0 | 0 | 0 | 1 | |
| Eurytomidae | 0 | 0 | 0 | 1 | |
| Trichogrammatidae | 0 | 0 | 0 | 2 | |
| Chewing predator | Asilidae | 5 | 6 | 0 | 0 |
| Mantidae | 1 | 1 | 1 | 0 | |
| Coccinellidae | 21 | 262 | 36 | 193 | |
| Carabidae | 0 | 5 | 0 | 3 | |
| Syrphidae | 0 | 101 | 0 | 4 | |
| Cantharidae | 0 | 0 | 0 | 1 | |
| Sucking predator | Geocoridae | 543 | 346 | 223 | 326 |
| Pentatomidae | 3 | 8 | 1 | 1 | |
| Chrysopidae | 5 | 1 | 3 | 13 | |
| Anthocoridae | 48 | 166 | 225 | 161 | |
| Nabidae | 100 | 340 | 37 | 93 | |
| Hemerobiidae | 0 | 10 | 0 | 3 | |
| Reduviidae | 0 | 0 | 0 | 4 | |
| Foliage feeder | Coccinellidae | 287 | 92 | 43 | 1 |
| Erebidae | 346 | 756 | 274 | 455 | |
| Meloidae | 0 | 1 | 0 | 0 | |
| Scarabaeidae | 428 | 284 | 90 | 96 | |
| Chrysomelidae | 2 | 345 | 0 | 254 | |
| Noctuidae | 0 | 1 | 0 | 0 | |
| Hesperiidae | 0 | 3 | 0 | 0 | |
| Plant sucking | Cicadellidae | 32 | 732 | 109 | 689 |
| Membracidae | 22 | 0 | 40 | 18 | |
| unspecified | 404 | 0 | 896 | 0 | |
| Aphididae | 0 | 0 | 0 | 60 | |
| Pod feeder | Pentatomidae | 33 | 164 | 69 | 115 |
| Miridae | 112 | 102 | 108 | 229 | |
| Unassigned | unspecified | 0 | 2 | 67 | 30 |
| Chrysomelidae | 0 | 0 | 0 | 2 | |
| Curculionidae | 2 | 5 | 0 | 30 | |
| Lampyridae | 4 | 21 | 17 | 5 | |
| Lygaeidae | 0 | 0 | 0 | 0 | |
| Elateridae | 18 | 5 | 0 | 12 | |
| Noctuidae | 0 | 0 | 0 | 1 | |
| Apidae | 0 | 0 | 1 | 0 | |
| Cynipidae | 0 | 0 | 18 | 3 | |
| Vespidae | 0 | 0 | 5 | 8 | |
| Chrysididae | 0 | 0 | 3 | 3 | |
| Pompilidae | 0 | 0 | 1 | 0 | |
| Scoliidae | 0 | 0 | 1 | 0 | |
| Thyreocoridae | 0 | 0 | 0 | 14 | |
| Berytidae | 0 | 0 | 0 | 41 | |
| Alydidae | 0 | 0 | 0 | 2 | |
| Dependent Variable | Independent Variable | Wald-χ2 | df | p |
|---|---|---|---|---|
| spider | treatment | 6.06 | 3 | 0.109 |
| stage | 24.37 | 4 | <0.001 | |
| site | 5.63 | 1 | 0.018 | |
| treatment × stage | 12.51 | 12 | 0.406 | |
| treatment × site | 4.61 | 3 | 0.203 | |
| stage × site | 4.81 | 4 | 0.308 | |
| treatment × stage × site | 16.62 | 12 | 0.165 | |
| chewing predator | treatment | 4.46 | 3 | 0.216 |
| stage | 134.41 | 4 | <0.001 | |
| site | 0.68 | 1 | 0.411 | |
| treatment × stage | 16.21 | 12 | 0.182 | |
| treatment × site | 3.99 | 3 | 0.263 | |
| stage × site | 4.98 | 4 | 0.289 | |
| treatment × stage × site | 10.21 | 12 | 0.597 | |
| sucking predator | treatment | 3.62 | 3 | 0.306 |
| stage | 113.88 | 4 | <0.001 | |
| site | 10.63 | 1 | 0.001 | |
| treatment × stage | 7.93 | 12 | 0.791 | |
| treatment × site | 0.89 | 3 | 0.827 | |
| stage × site | 3.54 | 4 | 0.472 | |
| treatment × stage × site | 3.88 | 12 | 0.985 | |
| parasitoid | treatment | 0.23 | 3 | 0.972 |
| stage | 57.56 | 4 | <0.001 | |
| site | 60.10 | 1 | <0.001 | |
| treatment × stage | 13.30 | 12 | 0.348 | |
| treatment × site | 3.50 | 3 | 0.321 | |
| stage × site | 12.90 | 4 | 0.012 | |
| treatment × stage × site | 5.66 | 12 | 0.932 | |
| pod feeder | treatment | 4.62 | 3 | 0.202 |
| stage | 25.11 | 4 | <0.001 | |
| site | 31.81 | 1 | <0.001 | |
| treatment × stage | 10.94 | 12 | 0.534 | |
| treatment × site | 1.76 | 3 | 0.624 | |
| stage × site | 18.26 | 4 | 0.001 | |
| treatment × stage × site | 6.71 | 12 | 0.876 | |
| plant sucking | treatment | 4.19 | 3 | 0.242 |
| stage | 29.22 | 4 | <0.001 | |
| site | 6.97 | 1 | 0.008 | |
| treatment × stage | 10.58 | 12 | 0.566 | |
| treatment × site | 4.64 | 3 | 0.200 | |
| stage × site | 20.44 | 4 | <0.001 | |
| treatment × stage × site | 4.19 | 12 | 0.980 | |
| foliage feeder | treatment | 2.21 | 3 | 0.531 |
| stage | 66.47 | 4 | <0.001 | |
| site | 121.46 | 1 | <0.001 | |
| treatment × stage | 17.73 | 12 | 0.124 | |
| treatment × site | 5.03 | 3 | 0.170 | |
| stage × site | 23.59 | 4 | <0.001 | |
| treatment × stage × site | 13.52 | 12 | 0.332 |
| Abundance per 10 Sweeps | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Feeding Guild | Site | R1 | R2 | R3 | R4 | R5 | |||||||||||||||
| Spider | Beltsville | 1.22 | ± | 0.26 | a 1 | 0.91 | ± | 0.20 | a | 1.27 | ± | 0.13 | a | 2.02 | ± | 0.25 | a | 1.28 | ± | 0.22 | a |
| Upper Marlboro | 1.19 | ± | 0.33 | a | 0.58 | ± | 0.11 | a | 0.96 | ± | 0.10 | a | 1.19 | ± | 0.18 | a | 1.25 | ± | 0.19 | a | |
| Parasitoid | Beltsville | 0.97 | ± | 0.20 | b | 1.28 | ± | 0.28 | ab | 1.23 | ± | 0.15 | ab | 3.33 | ± | 0.52 | a | 4.81 | ± | 0.92 | a |
| Upper Marlboro | 0.22 | ± | 0.11 | b | 0.56 | ± | 0.12 | ab | 0.72 | ± | 0.14 | ab | 0.97 | ± | 0.16 | ab | 1.28 | ± | 0.38 | a | |
| Chewing predator | Beltsville | 0.63 | ± | 0.33 | b | 0.13 | ± | 0.05 | ab | 0.82 | ± | 0.15 | ab | 1.25 | ± | 0.24 | ab | 1.56 | ± | 0.30 | a |
| Upper Marlboro | 0.28 | ± | 0.10 | b | 0.02 | ± | 0.02 | ab | 0.48 | ± | 0.10 | ab | 1.59 | ± | 0.31 | ab | 1.50 | ± | 0.31 | a | |
| Sucking predator | Beltsville | 3.75 | ± | 0.69 | b | 3.41 | ± | 0.51 | b | 4.03 | ± | 0.37 | ab | 8.27 | ± | 0.65 | a | 6.41 | ± | 0.73 | ab |
| Upper Marlboro | 2.13 | ± | 0.34 | b | 2.59 | ± | 0.35 | b | 2.03 | ± | 0.18 | ab | 7.84 | ± | 1.05 | a | 5.91 | ± | 0.64 | ab | |
| Foliage feeder | Beltsville | 8.78 | ± | 0.92 | b | 7.50 | ± | 0.86 | ab | 9.62 | ± | 0.70 | a | 6.25 | ± | 0.85 | b | 4.06 | ± | 0.86 | b |
| Upper Marlboro | 2.81 | ± | 0.95 | b | 1.84 | ± | 0.26 | b | 5.17 | ± | 0.41 | a | 2.89 | ± | 0.36 | ab | 2.41 | ± | 0.52 | b | |
| Plant sucking | Beltsville | 6.81 | ± | 1.16 | ab | 5.66 | ± | 0.61 | a | 2.73 | ± | 0.21 | ab | 3.20 | ± | 0.43 | b | 2.22 | ± | 0.32 | b |
| Upper Marlboro | 3.81 | ± | 0.72 | a | 8.61 | ± | 1.01 | a | 2.97 | ± | 0.19 | a | 15.00 | ± | 3.66 | a | 8.38 | ± | 1.77 | a | |
| Pod feeder | Beltsville | 0.81 | ± | 0.24 | a | 0.64 | ± | 0.15 | a | 1.23 | ± | 0.16 | a | 0.50 | ± | 0.14 | a | 1.34 | ± | 0.36 | a |
| Upper Marlboro | 1.00 | ± | 0.21 | a | 1.73 | ± | 0.28 | a | 1.09 | ± | 0.15 | a | 1.69 | ± | 0.29 | a | 3.50 | ± | 0.66 | a | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosario-Lebron, A.; Leslie, A.W.; Chen, G.; Hooks, C.R.R. The Effect of Barley Cover Crop Residue and Herbicide Management on the Foliar Arthropod Community in No-Till Soybeans. Agronomy 2018, 8, 87. https://doi.org/10.3390/agronomy8060087
Rosario-Lebron A, Leslie AW, Chen G, Hooks CRR. The Effect of Barley Cover Crop Residue and Herbicide Management on the Foliar Arthropod Community in No-Till Soybeans. Agronomy. 2018; 8(6):87. https://doi.org/10.3390/agronomy8060087
Chicago/Turabian StyleRosario-Lebron, Armando, Alan W. Leslie, Guihua Chen, and Cerruti R. R. Hooks. 2018. "The Effect of Barley Cover Crop Residue and Herbicide Management on the Foliar Arthropod Community in No-Till Soybeans" Agronomy 8, no. 6: 87. https://doi.org/10.3390/agronomy8060087
APA StyleRosario-Lebron, A., Leslie, A. W., Chen, G., & Hooks, C. R. R. (2018). The Effect of Barley Cover Crop Residue and Herbicide Management on the Foliar Arthropod Community in No-Till Soybeans. Agronomy, 8(6), 87. https://doi.org/10.3390/agronomy8060087