Factors Limiting the Growth of Eucalyptus and the Characteristics of Growth and Water Use under Water and Fertilizer Management in the Dry Season of Leizhou Peninsula, China

Abstract
:1. Introduction
2. Materials and Methods
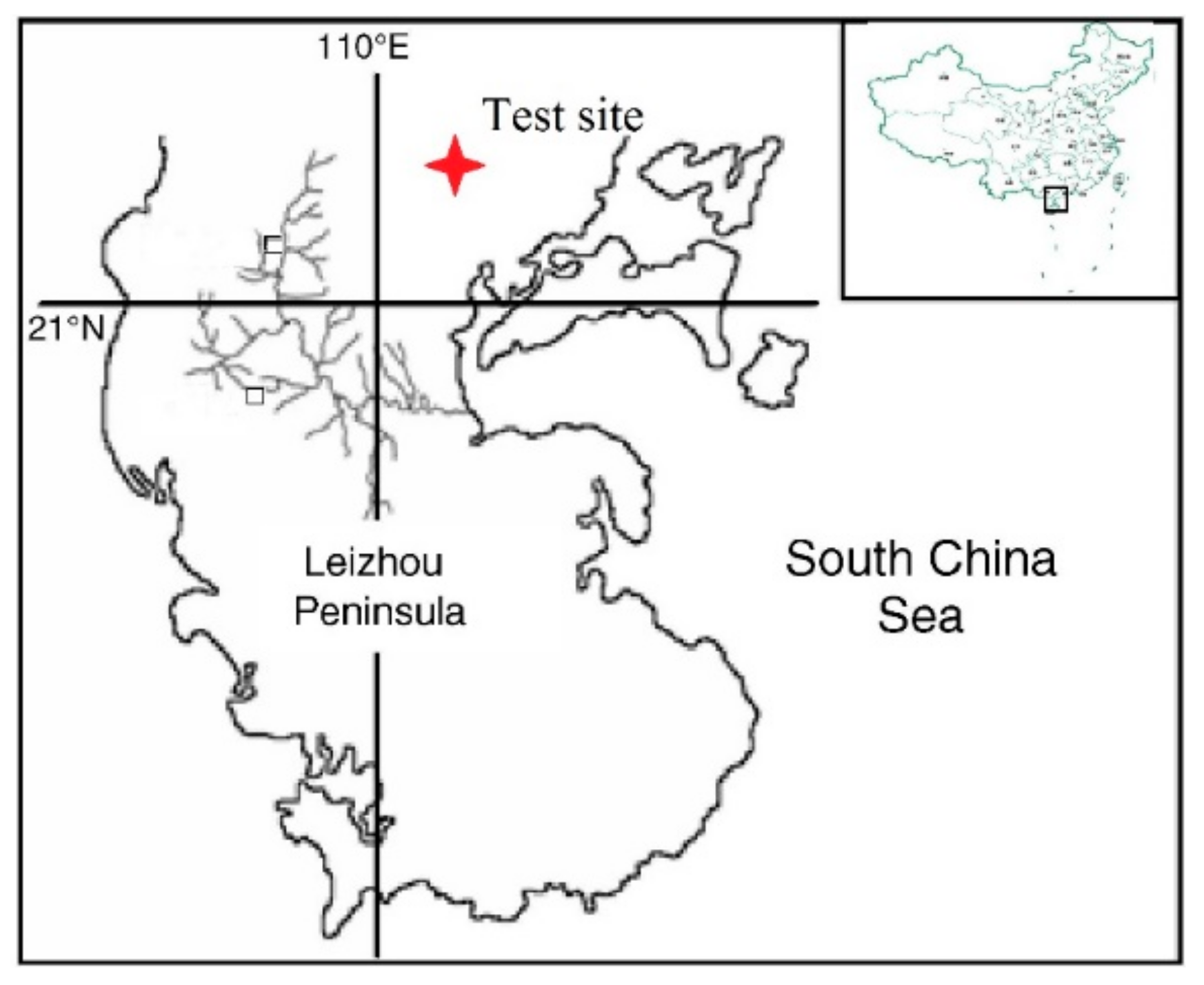
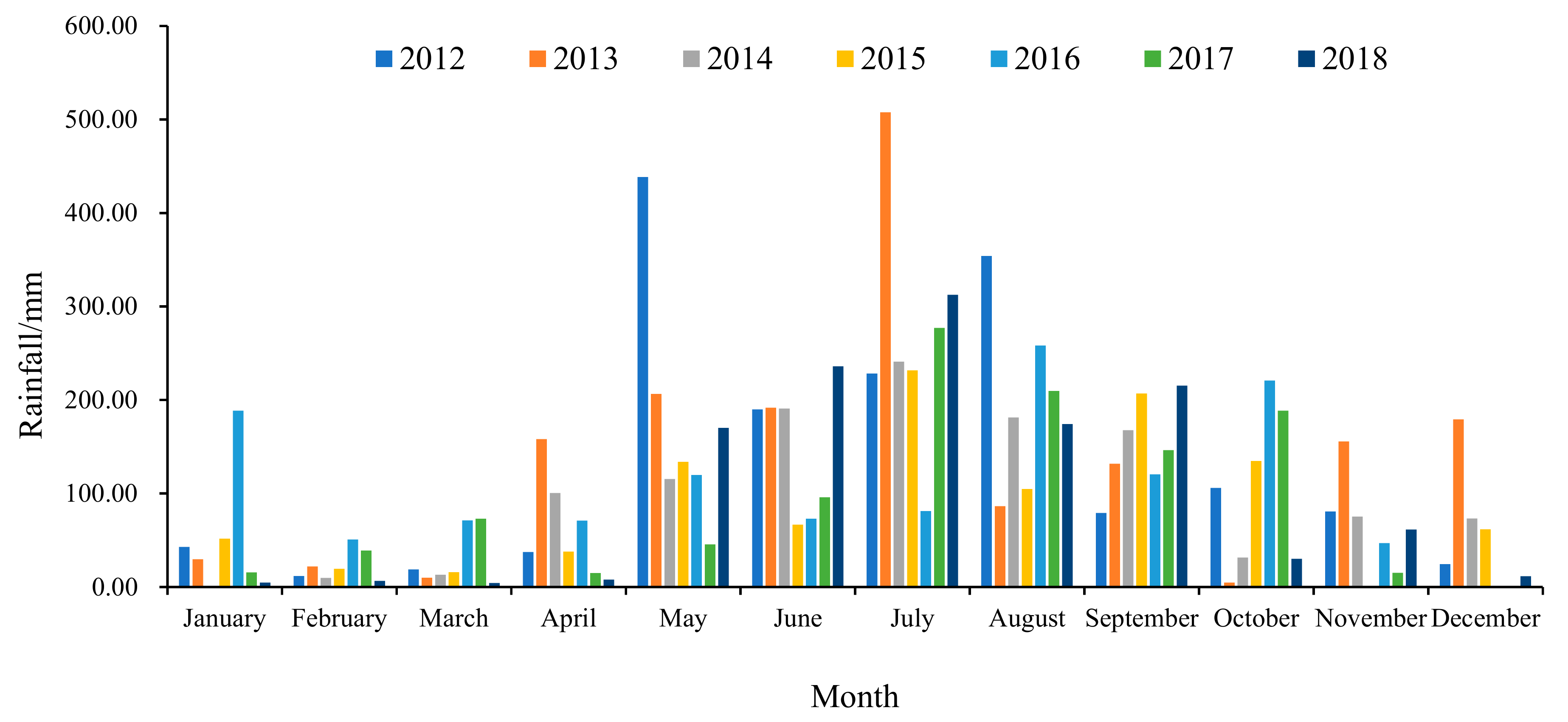
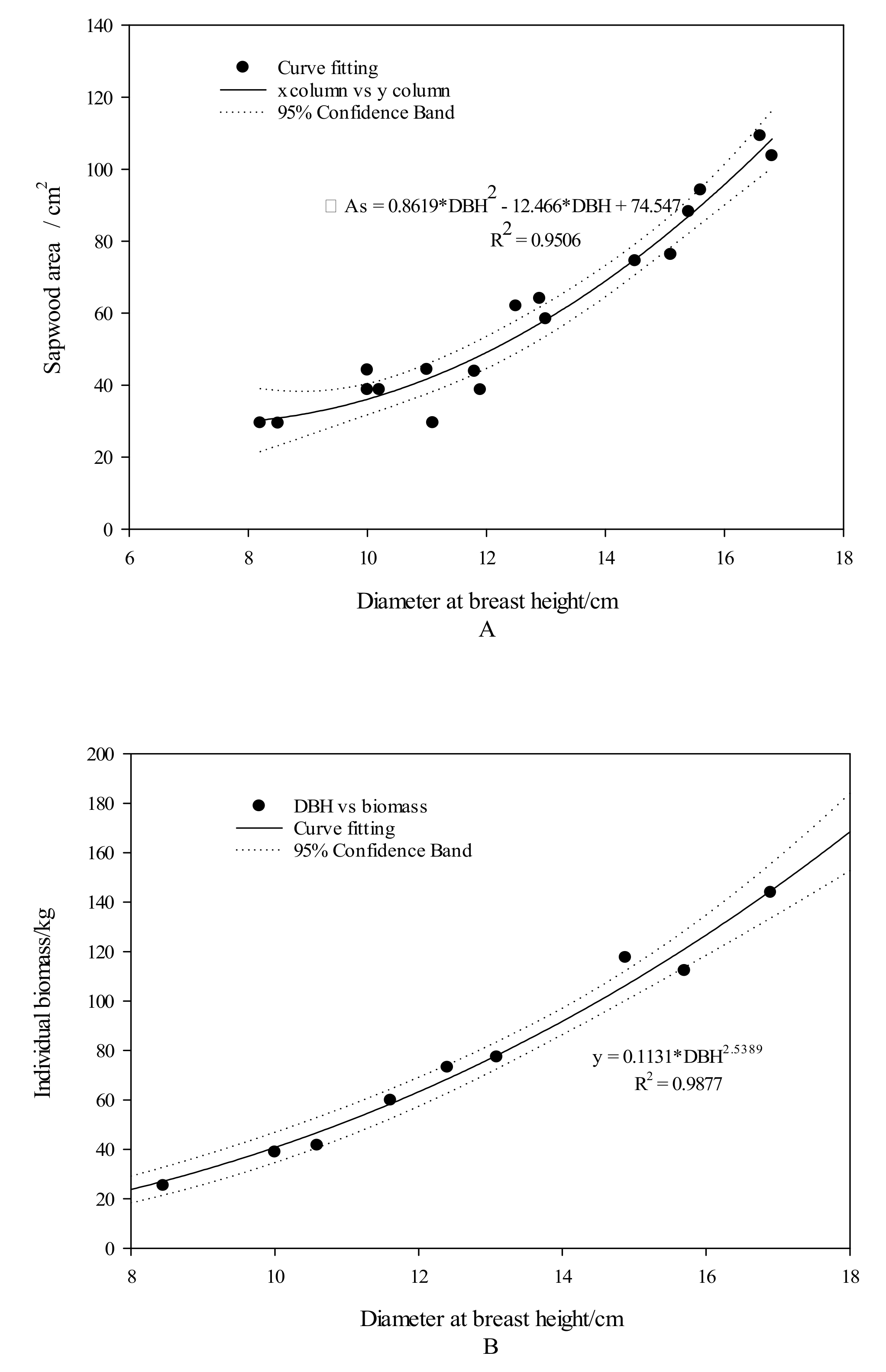
2.1. Study Site and Materials
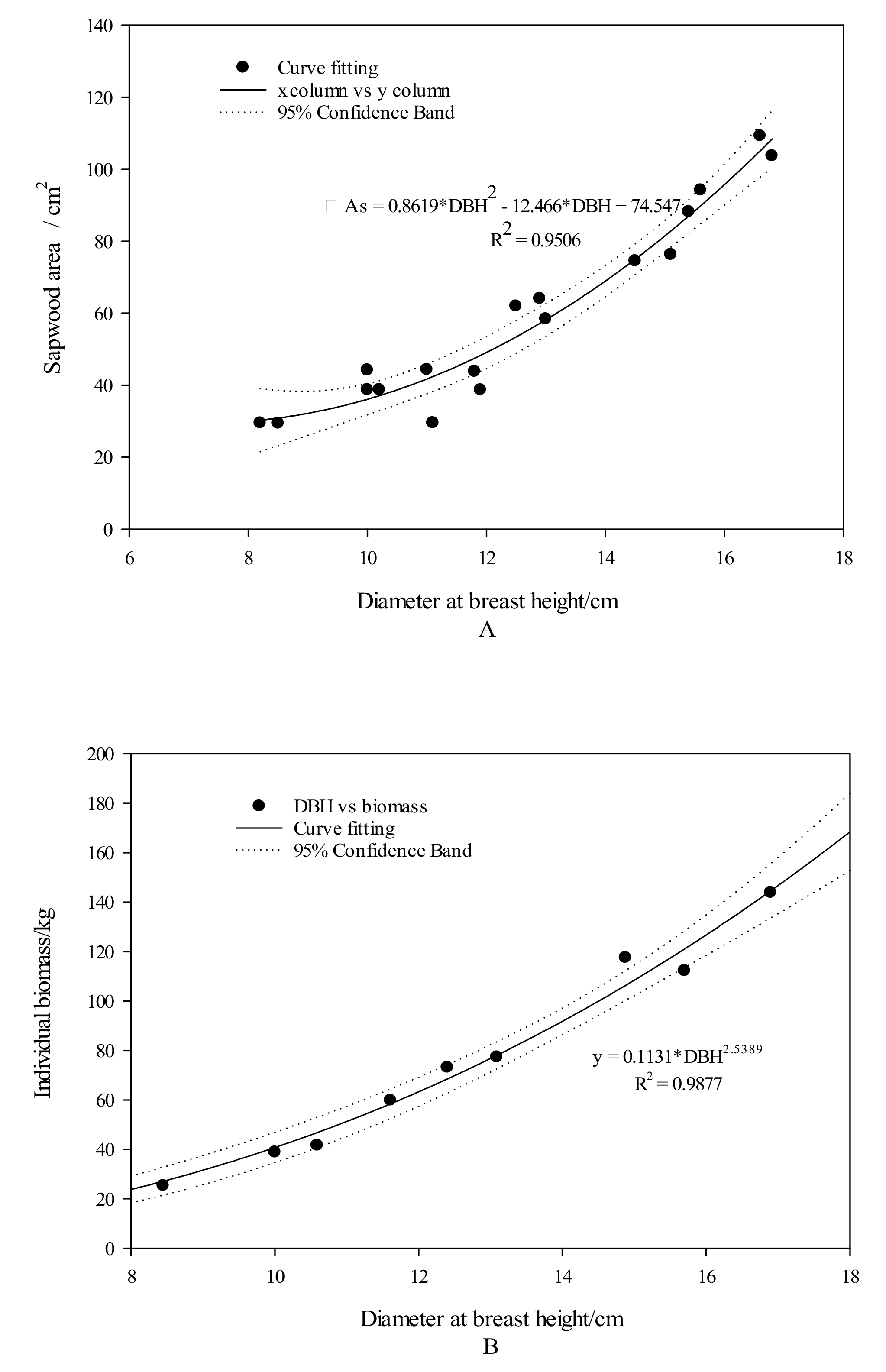
2.2. Research Method
2.3. Statistical Analysis
3. Results
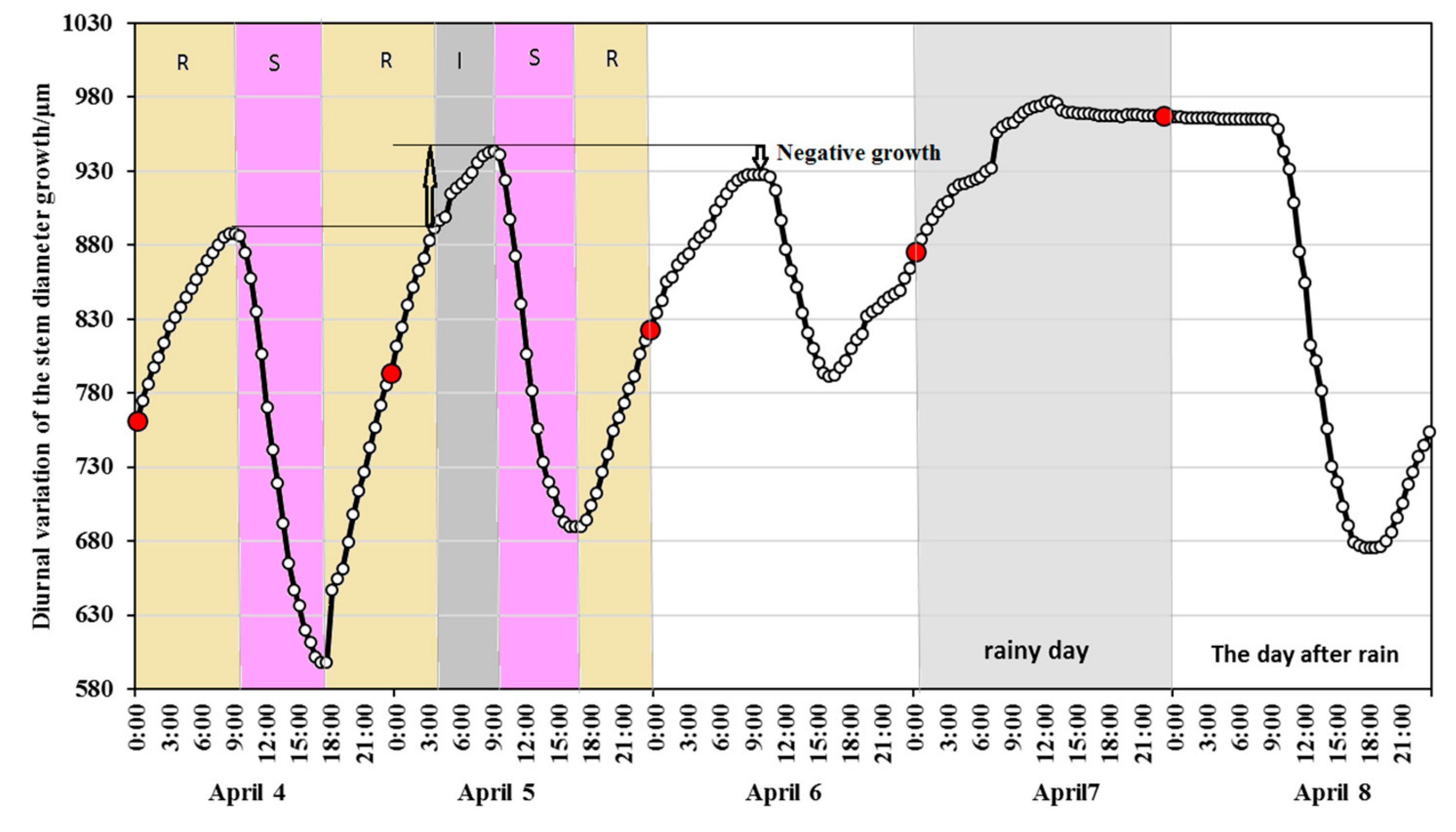
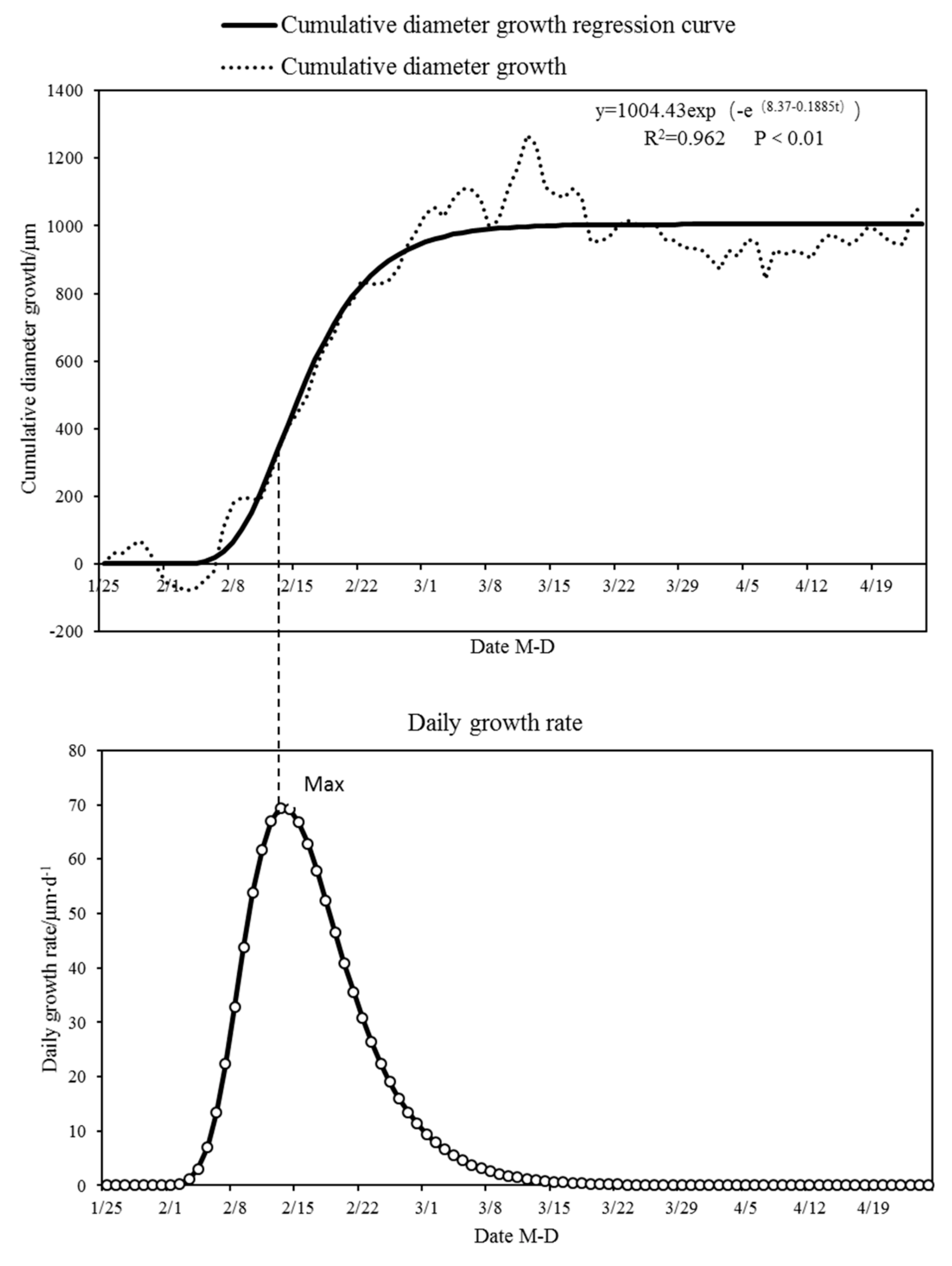
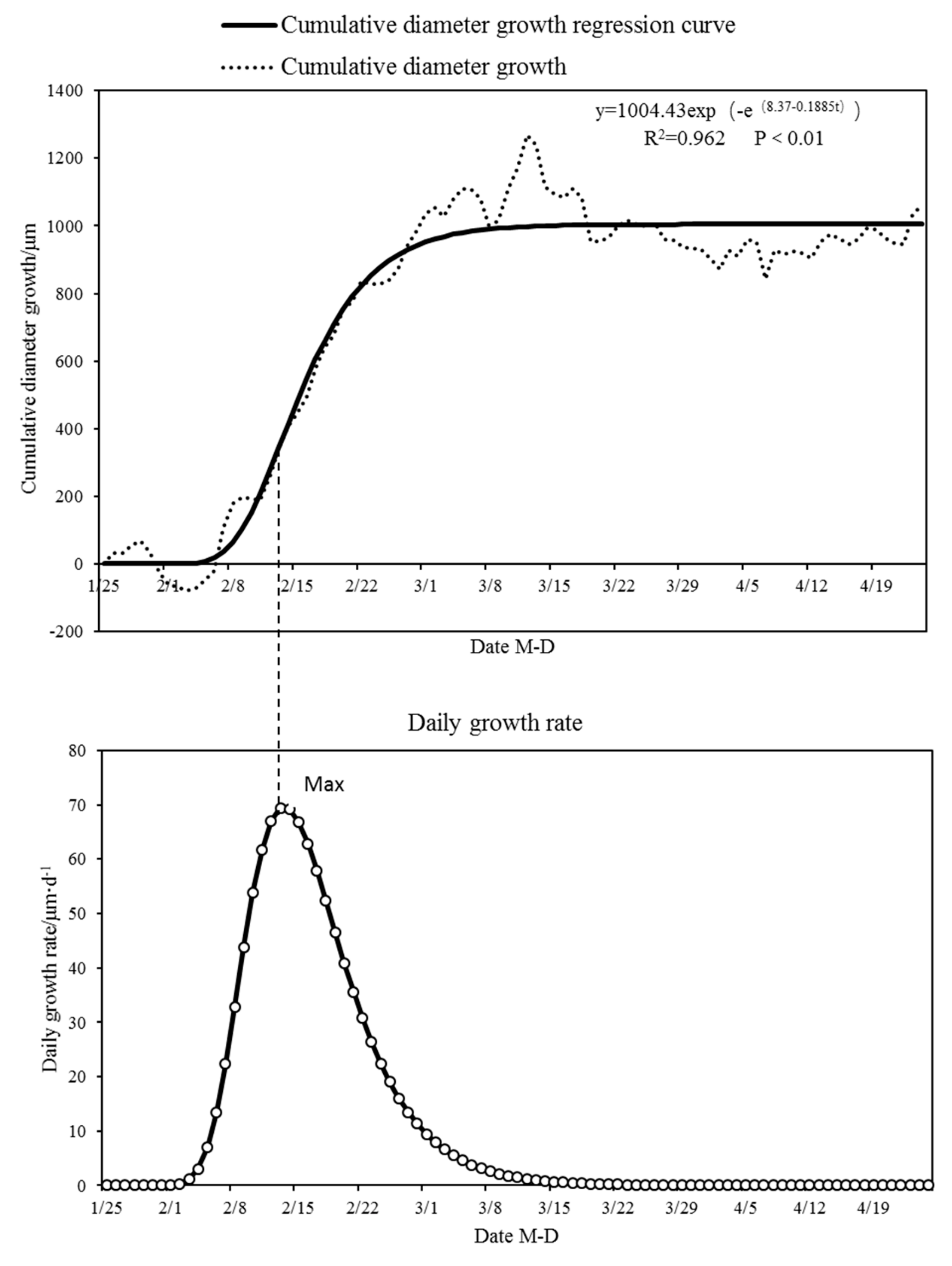
3.1. Diameter Diurnal Variation and Cumulative Change Dynamics During Dry Season
3.2. Analysis of Limiting Factors for Diameter Growth in Dry Season
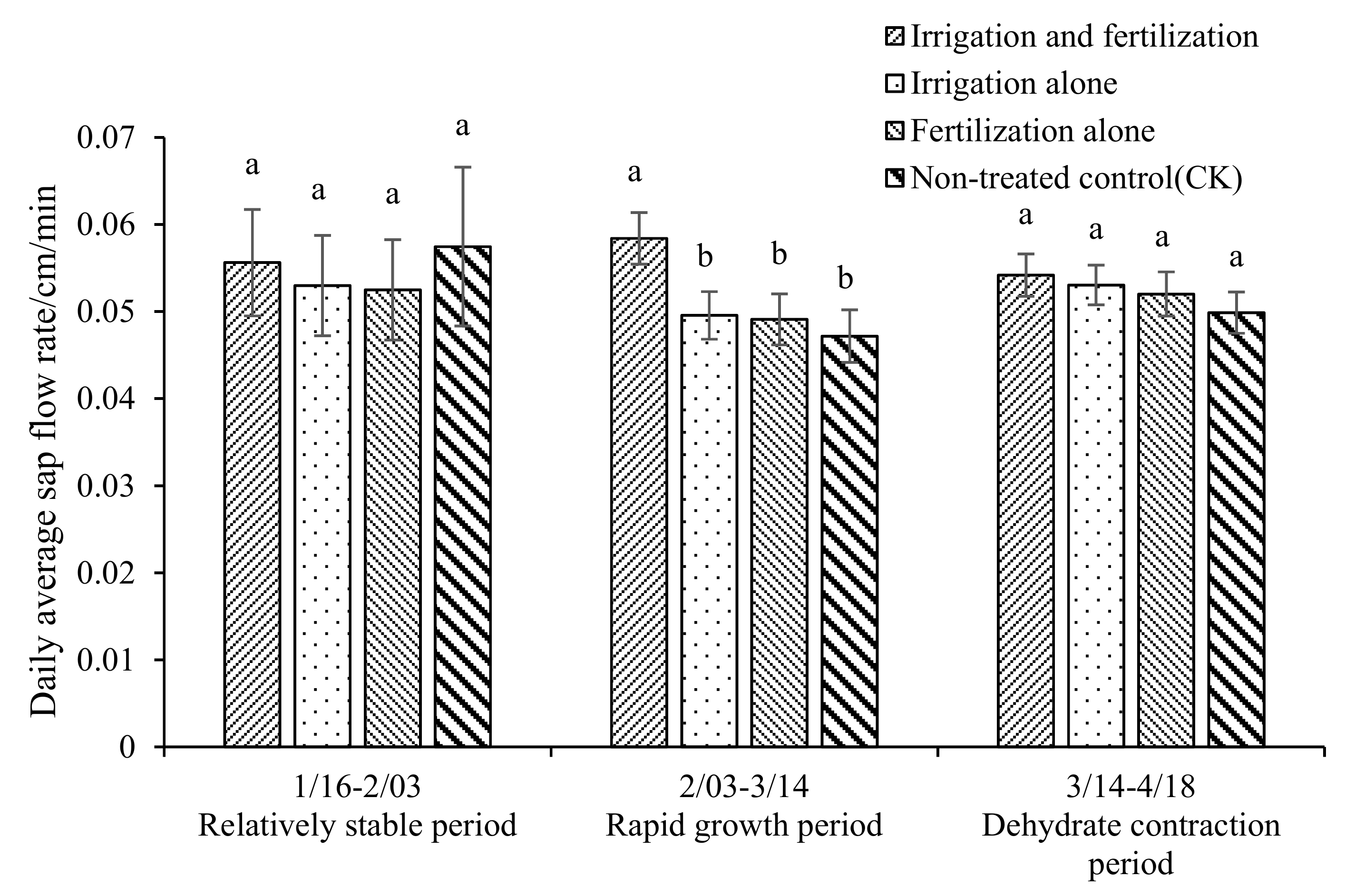
3.3. Characteristics of Growth and Water Use Under Water and Fertilizer Management in Dry Season
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Turnbull, J.W. Development of sustainable forestry plantations in China: A review. Int. For. Rev. 2008, 10, 95–96. [Google Scholar] [CrossRef]
- Bai, G.X.; Wang, Y.Y.; Dai, L.M.; Liu, S.R.; Tang, L.N.; Shao, G.F. Market-oriented forestry in China promotes forestland productivity. New For. 2015, 46, 1–6. [Google Scholar] [CrossRef]
- Trømborg, E.; Buongiorno, J.; Solberg, B. The global timber market: Implications of changes in economic growth, timber supply, and technological trends. For. Policy Econ. 2007, 1, 53–69. [Google Scholar] [CrossRef]
- Warde, P. Fear of Wood Shortage and the Reality of the Woodland in Europe, c.1450–1850. Hist. Workshop J. 2006, 62, 28–57. [Google Scholar] [CrossRef]
- Xie, Y.J.; Arnold, R.J.; Wu, Z.H.; Chen, S.F.; Du, A.P.; Luo, J.Z. Advances in eucalypt research in China. Front. Agric. Sci.Eng. 2017, 4, 380–390. [Google Scholar] [CrossRef]
- Liu, S.R.; Yang, Y.J.; Wang, H. Development strategy and management countermeasures of planted forests in China: Transforming from timber-centered single objective management towards multi-purpose management for enhancing quality and benefits of ecosystem services. Acta Ecol. Sin. 2018, 38, 1–10. [Google Scholar] [CrossRef]
- Huang, G.Q.; Zhao, Q.G. The history, status quo, ecological problems and countermeasures of Eucalyptus plantations in Guangxi. Acta Ecol. Sin. 2014, 34, 5142–5152. [Google Scholar] [CrossRef]
- Albaugh, J.M.; Dye, P.J.; King, J.S. Eucalyptus and Water Use in South Africa. Int. J. For. Res. 2013, 2013, 1–11. [Google Scholar] [CrossRef]
- Nouvellon, Y.; Laclau, J.P.; Epron, D.; Le Maire, G.; Bonnefond, J.M.; Goncalves, J.L.; Bouillet, J.P. Production and carbon allocation in monocultures and mixed-species plantations of Eucalyptus grandis and Acacia mangium in Brazil. Tree Physiol. 2012, 32, 680–695. [Google Scholar] [CrossRef]
- Ryan, M.G.; Stape, J.L.; Binkley, D.; Fonseca, S.; Loos, R.A.; Takahashi, E.N.; Silva, C.R.; Silva, S.R.; Hakamada, R.E.; Ferreira, J.M.; et al. Factors controlling Eucalyptus productivity: How water availability and stand structure alter production and carbon allocation. For. Ecol. Manag. 2010, 259, 1695–1703. [Google Scholar] [CrossRef]
- Ding, L.; Zheng, J.; Hui, F.U. Progress in Research of Climate Change in South China Based on Historical Documents. Trop. Geogr. 2015, 35, 890–894. [Google Scholar] [CrossRef]
- Nodzu, M.I.; Tachibana, Y.; Yamanaka, M.D. Climatological Description of Seasonal Variations in Lower-Tropospheric Temperature Inversion Layers over the Indochina Peninsula. J. Clim. 2006, 19, 3307–3319. [Google Scholar] [CrossRef]
- Wang, F.M.; Li, J.; Zou, B.; Xu, X.; Li, Z. Effect of Prescribed Fire on Soil Properties and N Transformation in Two; Vegetation Types in South China. Environ. Manag. 2013, 51, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.C. The Characteristics of Growth, Carbon Sequestration and Water Consumption in Eucalyptus urophylla × E. grandis Plantations under Three Different Site Preparation Measures; Chinese Academy of Forestry: Beijing, China, 2015. [Google Scholar]
- Fan, W.B.; Zhao, C.J.; Lin, Z.; Chen, H. Growth Characteristics of Eucalyptus Plantation and Their Responses to Climate Environment in Western Hainan Island. For. Resour. Manag. 2013, 2013, 77–82. [Google Scholar] [CrossRef]
- Wang, Z.C.; XU, Y.X.; Zhu, W.K.; Du, A.P. The transpiration water consumption of two common fast-growing forests in Leizhou Peninsula. Acta Ecol. Sin. 2019, 39, 2147–2155. [Google Scholar] [CrossRef]
- Wang, Z.C.; Zhu, W.K.; Du, A.P. Variation in stem sap flow of Eucalyptus urophylla × E. grandis during rainy and dry seasons. J. Zhejiang A F Univ. 2017, 34, 319–325. [Google Scholar] [CrossRef]
- Kramer, P.J.; Kramer, P.J. Water Relations of Plants. Bot. Gaz. 1913, 55, 264. [Google Scholar] [CrossRef]
- Ford, B.J. Physiology of Woody Plants. Bot. J. Linnean Soc. 2010, 153, 243. [Google Scholar] [CrossRef]
- Yang, L.C.; Berninger, F.; Koskela, J.; Sonninen, E. Drought responses of Eucalyptus microtheca provences depend on seasonality of rainfall in their place of origin. Funct. Plant Biol. 2000, 27, 231–238. [Google Scholar] [CrossRef]
- Ding, Y.L.; Chen, H.S.; Nie, Y.P.; Wang, S.; Zhang, H.L.; Wang, K.L. Water use strategy of Eucalyptus urophylla ×E. grandis on karst hillslope based on isotope analysis. Chin. J. Appl. Ecol. 2016, 27, 2729–2736. [Google Scholar] [CrossRef]
- Hubbard, R.M.; Ryan, M.G.; Giardina, C.P.; Barnard, H. The effect of fertilization on sap flux and canopy conductance in a Eucalyptus saligna experimental forest. Glob. Chang. Biol. 2010, 10, 427–436. [Google Scholar] [CrossRef]
- Li, Y.Y.; Shi, H.; Zhang, A.B.; Tan, H.C. Daily Changes in Radial Growth of Several Tree Stems and Their Response to Environmental Factors in Loess Hilly Region. J. Soil Water Conserv. 2007, 21, 170–173. [Google Scholar] [CrossRef]
- Ortuño, M.F.; Conejero, W.; Moreno, F.; Moriana, A.; Intrigliolo, D.S.; Biel, C.; Mellisho, C.D.; Pérezpastor, A.; Domingo, R.; Ruizsánchez, M.C. Could trunk diameter sensors be used in woody crops for irrigation scheduling? A review of current knowledge and future perspectives. Agric. Water Manag. 2010, 97, 1–11. [Google Scholar] [CrossRef]
- Deslauriers, A.; Morin, H.; Urbinati, C.; Carrer, M. Daily weather response of balsam fir (Abies balsamea (L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada). Trees 2003, 17, 477–484. [Google Scholar] [CrossRef]
- Granier, A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiol. 1987, 3, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Bouriaud, O.; Leban, J.M.; Bert, D.; Deleuze, C. Intra-annual variations in climate influence growth and wood density of Norway spruce. Tree Physiol. 2005, 25, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Wang, Y.H.; Yu, P.T.; Liu, H.L.; Shi, Z.J.; Guan, W. Growth in stem diameter of Larix principis-rupprechtii and its response to meteorological factors in the south of Liupan Mountain, China. Acta Ecol. Sin. 2007, 27, 432–440. [Google Scholar] [CrossRef]
- Jiang, Y.; Yang, Y.G.; Dong, M.Y.; Zhang, W.T.; Ren, F.P. Stem radius growth of Picea meyeri and Larix principis-rupprechtii nearby the tree-line of Luya Mountain. Chin. J. Appl. Ecol. 2009, 20, 1271–1277. [Google Scholar] [CrossRef]
- Guan, W.; Xiong, W.; Wang, Y.H.; Yu, P.T.; He, C.Q.; Du, A.P.; Liu, H.L. Stem Diameter Growth of Larix principis-rupprechtii and Its Response to Meteorological Factors in the North of Liupan Mountain. Scientia Silvae Sinicae 2007, 43, 1–6. [Google Scholar] [CrossRef]
- Dong, M.Y.; Jiang, Y.; Yang, H.C.; Wang, M.C.; Zhang, W.T. Dynamics of stem radial growth of Picea meyeri during the growing season at the treeline of Luya Mountain, China. Chin. J. Plant Ecol. 2012, 36, 956–964. [Google Scholar] [CrossRef]
- MäKinen, H.; Seo, J.-W.; Nöjd, P.; Schmitt, U.; Jalkanen, R. Seasonal dynamics of wood formation: A comparison between pinning, microcoring and dendrometer measurements. Eur. J. For. Res. 2008, 127, 235–245. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Morin, H. Application of the Gompertz equation for the study of xylem cell development. Dendrochronologia 2004, 21, 33–39. [Google Scholar] [CrossRef]
- Subramanian, N.; Nilsson, U.; Mossberg, M.; Bergh, J. Impacts of climate change, weather extremes and alternative strategies in managed forests. Ecoscience 2018, 26, 1–18. [Google Scholar] [CrossRef]
- Deng, M.J.; Wang, X.A.; Han, B.B.; Li, J. Characteristics of Radial Growth of Larix chinensis in Different Time Scales and Its Relationship with Environment Factors in the North Slope of Qinling Mountains. Chin. Agric. Sci. Bull. 2016, 32, 1–6. [Google Scholar] [CrossRef]
- Deslauriers, A.; Rossi, S.; Anfodillo, T. Dendrometer and intra-annual tree growth: What kind of information can be inferred? Dendrochronologia 2008, 25, 113–124. [Google Scholar] [CrossRef]
- Wimmer, R.; Downes, G.M.; Evans, R. High-resolution analysis of radial growth and wood density in Eucalyptus nitens, grown under different irrigation regimes. Ann. For. Sci. 2002, 59, 519–524. [Google Scholar] [CrossRef]
- Dietrich, L.; Zweifel, R.; Kahmen, A. Daily stem diameter variations can predict the canopy water status of mature temperate trees. Tree Physiol. 2018, 38, 941–952. [Google Scholar] [CrossRef]
- Schäfer, C.; Thurm, E.A.; Rötzer, T.; Kallenbach, C.; Pretzsch, H. Daily stem water deficit of Norway spruce and European beech in intra- and interspecific neighborhood under heavy drought. Scand. J. For. Res. 2018, 33, 568–582. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Winget, C.H. Diurnal and Seasonal Variation in Radii of Tree Stems. Ecology 1964, 45, 149–155. [Google Scholar] [CrossRef]
- Steppe, K.; De Pauw, D.J.W.; Lemeur, R.; Vanrolleghem, P.A. A mathematical model linking tree sap flow dynamics to daily stem diameter fluctuations and radial stem growth. Tree Physiol. 2006, 26, 257. [Google Scholar] [CrossRef]
- Zweifel, R.; Zimmermann, L.; Newbery, D.M. Modeling tree water deficit from microclimate: An approach to quantifying drought stress. Tree Physiol. 2005, 25, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Brække, F.H.; Kozlowski, T.T. Shrinkage and swelling of stems of Pinus resinosa and Betula papyrifera in northern Wisconsin. Plant Soil 1975, 43, 387–410. [Google Scholar] [CrossRef]
- Wang, Z.C.; Du, A.P. Characteristics of Stem Sap Flow of Eucalypt and Its Relation to Environmental Factors. J. Northeast For. Univ. 2016, 44, 24–28. [Google Scholar] [CrossRef]
- De Swaef, T.; De Schepper, V.; Vandegehuchte, M.W.; Steppe, K.; Way, D. Stem diameter variations as a versatile research tool in ecophysiology. Tree Physiol. 2015, 35, 1047–1061. [Google Scholar] [CrossRef] [PubMed]
- Gaafar, R.M. Molecular marker analysis of heading date Hd1 locus in Egyptian rice varieties. Afr. J. Biotechnol. 2010, 9, 3368–3372. [Google Scholar] [CrossRef]
- Kojima, S. Hd3a, a Rice Ortholog of the Arabidopsis FT Gene, Promotes Transition to Flowering Downstream of Hd1 under Short-Day Conditions. Plant Cell Physiol. 2002, 43, 1096–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuny, H.E.; Rathgeber, C.B.K.; Frank, D.; Fonti, P.; Mäkinen, H.; Prislan, P.; Rossi, S.; del Castillo, E.M.; Campelo, F.; Vavrčík, H.; et al. Woody biomass production lags stem-girth increase by over one month in coniferous forests. Nat. Plants 2015, 1, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zweifel, R.; Item, H.; Häsler, R. Stem radius changes and their relation to stored water in stems of young Norway spruce trees. Trees 2000, 15, 50–57. [Google Scholar] [CrossRef]
- Schepper, V.D.; Steppe, K.; Tardieu, F. Development and verification of a water and sugar transport model using measured stem diameter variations. J. Exp. Bot. 2010, 61, 2083–2099. [Google Scholar] [CrossRef] [Green Version]
- Mäkinen, H.; Nöjd, P.; Saranpää, P. Seasonal changes in stem radius and production of new tracheids in Norway spruce. Tree Physiol. 2003, 23, 959–968. [Google Scholar] [CrossRef]
- Goldblum, D.; Rigg, L.S. Tree growth response to climate change at the deciduous–boreal forest ecotone, Ontario, Canada. Revue Canadienne De Recherche Forestière 2005, 35, 2709–2718. [Google Scholar] [CrossRef]
- Biondi, F.; Hartsough, P. Using Automated Point Dendrometers to Analyze Tropical Treeline Stem Growth at Nevado de Colima, Mexico. Sensors 2010, 10, 5827–5844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, J.; Campelo, F.; Rossi, S.; Carvalho, A.; Freitas, H.; Nabais, C. Adjustment capacity of maritime pine cambial activity in drought-prone environments. PLoS ONE 2015, 5, e0126223. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.X.; Bi-Yun, Y.U.; Liang, H.X.; Huang, J.G. Advancement in studies of tree growth and ecophysiology incorporating micro-sampling approach. Chin. J. Plant Ecol. 2017, 41, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Dan, T.W.; Pate, J.S.; Bartle, J.R. Budgets of water use by Eucalyptus kochiitree belts in the semi-arid wheatbelt of Western Australia. Plant Soil 2004, 262, 129–149. [Google Scholar] [CrossRef]
- Drew, D.M.; Downes, G.M.; Grzeskowiak, V.; Naidoo, T. Differences in daily stem size variation and growth in two hybrid eucalypt clones. Trees 2009, 23, 585. [Google Scholar] [CrossRef]
- Ward, E.J.; Domec, J.C.; Laviner, M.A.; Fox, T.R.; Ge, S.; Mcnulty, S.; King, J.; Noormets, A. Fertilization intensifies drought stress: Water use and stomatal conductance of Pinus taeda in a midrotation fertilization and throughfall reduction experiment. For. Ecol. Manag. 2015, 355, 72–82. [Google Scholar] [CrossRef]
- Granier, A.; Bréda, N.; Longdoz, B.; Gross, P.; Ngao, J.M. Ten years of fluxes and stand growth in a young beech forest at Hesse, North-eastern France. Ann. For. Sci. 2008, 65, 704. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Sample Tree Number | DBH/cm | Tree Height/m | Crown Width (EW/m × SN/m) |
|---|---|---|---|---|
| Non-treated control (CK) | 1 | 14.72 | 16.2 | 3.8 × 4.0 |
| 2 | 13.21 | 15.2 | 2.5 × 3.0 | |
| 3 1 | 14.94 13.78 | 16.4 15.3 | 3.2 × 3.6 2.6 × 3.2 | |
| Irrigation alone | ||||
| 2 | 15.94 | 16.2 | 3.5 × 3.6 | |
| 3 1 | 14.89 14.76 | 16.4 15.7 | 3.0 × 3.2 3.3 × 3.5 | |
| Fertilization alone | ||||
| 2 | 13.43 | 16 | 2.8 × 3.1 | |
| 3 1 | 14.91 13.52 | 16.4 16.2 | 3.3 × 3.6 2.1 × 3.7 | |
| Irrigation and fertilization | ||||
| 2 | 13.13 | 15.5 | 2.6 × 3.8 | |
| 3 | 16.24 | 17.2 | 3.4 × 4.1 |
| Item | Atmospheric Temperature °C | Air Relative Humidity % | Wind Speed m·s−1 | Daily Average PAR μmol·(s−1·m−2) | Rainfallmm·d−1 | Water Surface Evaporation mm |
VPD Kpa |
|---|---|---|---|---|---|---|---|
| Mean | 19.50 | 79.58 | 5.75 | 112.43 | 0.25 | 0.34 | 0.42 |
| Minimum | 5.64 | 40.73 | 1.21 | 18.20 | 0.00 | 0.09 | 0.04 |
| Maximum | 27.37 | 98.10 | 12.23 | 217.50 | 2.60 | 5.08 | 0.77 |
| Environmental Factors | Pearson Correlation Analysis | Partial Correlation Analysis | ||
|---|---|---|---|---|
| Pearson Correlation Coefficient | P | Partial Correlation Coefficient | P | |
| Atmospheric temperature | 0.744 ** | 0.000 | 0.282 * | 0.014 |
| Relative humidity of air | 0.359 ** | 0.001 | 0.103 | 0.380 |
| Wind speed | −0.022 | 0.847 | ||
| Photosynthetically active radiation | 0.347 ** | 0.002 | 0.019 | 0.870 |
| Rainfall | −0.041 | 0.718 | ||
| Soil volumetric water content | 0.892 ** | 0.000 | 0.623 ** | 0.000 |
| Soil temperature | 0.81 ** | 0.000 | −0.122 | 0.296 |
| Vapor pressure difference | 0.184 | 0.333 | ||
| Growth Attribute | Treatments | Different Growth Stages | ||
|---|---|---|---|---|
| 1/16–2/03 (Relatively Stable Period) | 2/03–3/14 (Rapid Growth Period) | 3/14–4/18 (Dehydrate Contraction Period) | ||
| Breast diameter increase | Irrigation and fertilization | 0.093 ± 0.006 a | 0.168 ± 0.011 a | 0.03 ± 0.024 a |
| Irrigation alone | 0.075 ± 0.013 ab | 0.134 ± 0.025 b | −0.029 ± 0.027 a | |
| Fertilization alone | 0.054 ± 0.003 b | 0.151 ± 0.008 ab | 0.006 ± 0.012 a | |
| Non-treated control (CK) | 0.073 ± 0.012 ab | 0.111 ± 0.014 b | −0.02 ± 0.014 a | |
| Breast diameter cumulative growth | Irrigation and fertilization | 0.093 ± 0.006 a | 0.261 ± 0.017 a | 0.291 ± 0.028 a |
| Irrigation alone | 0.075 ± 0.013 ab | 0.209 ± 0.035 ab | 0.206 ± 0.025 b | |
| Fertilization alone | 0.054 ± 0.003 b | 0.205 ± 0.01 ab | 0.211 ± 0.018 b | |
| Non-treated control (CK) | 0.073 ± 0.012 ab | 0.184 ± 0.02 b | 0.164 ± 0.03 b | |
| Treatments | Different Growth Stages | |||
|---|---|---|---|---|
| 1/16–2/03 (Relatively Stable Period) | 2/03–3/14 (Rapid Growth Period) | 3/14–4/18 (Dehydrate Contraction Period) | Total | |
| Irrigation and fertilization | 15.10 ± 0.991 a | 12.77 ± 0.694 a | 4.13 ± 0.99 a | 9.9 ± 0.964 a |
| Irrigation alone | 13.99 ± 1.758 a | 12.97 ± 1.641 a | 0.22 ± 0.146 b | 7.09 ± 0.939 b |
| Fertilization alone | 8.64 ± 0.505 b | 12.86 ± 0.613 a | 1.51 ± 0.355 b | 7.09 ± 0.578 b |
| Non-treated control (CK) | 11.75 ± 1.845 ab | 10.79 ± 1.249 a | 0.67 ± 0.355 b | 6.21 ± 1.076 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Du, A.; Xu, Y.; Zhu, W.; Zhang, J. Factors Limiting the Growth of Eucalyptus and the Characteristics of Growth and Water Use under Water and Fertilizer Management in the Dry Season of Leizhou Peninsula, China. Agronomy 2019, 9, 590. https://doi.org/10.3390/agronomy9100590
Wang Z, Du A, Xu Y, Zhu W, Zhang J. Factors Limiting the Growth of Eucalyptus and the Characteristics of Growth and Water Use under Water and Fertilizer Management in the Dry Season of Leizhou Peninsula, China. Agronomy. 2019; 9(10):590. https://doi.org/10.3390/agronomy9100590
Chicago/Turabian StyleWang, Zhichao, Apeng Du, Yuxing Xu, Wankuan Zhu, and Jing Zhang. 2019. "Factors Limiting the Growth of Eucalyptus and the Characteristics of Growth and Water Use under Water and Fertilizer Management in the Dry Season of Leizhou Peninsula, China" Agronomy 9, no. 10: 590. https://doi.org/10.3390/agronomy9100590