Growth, Secondary Metabolites and Enzyme Activity Responses of Two Edible Fern Species to Drought Stress and Rehydration in Northeast China

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experiment and Stress Treatment
2.3. Growth Traits
2.4. Hydrogen Peroxide (H2O2), Malondialdehyde (MDA) Content and Ion Leakage (IL)
2.5. Osmotic Regulators
2.6. Secondary Metabolites
2.7. Antioxidant Enzyme Activities
2.8. Statistical Analysis
3. Results
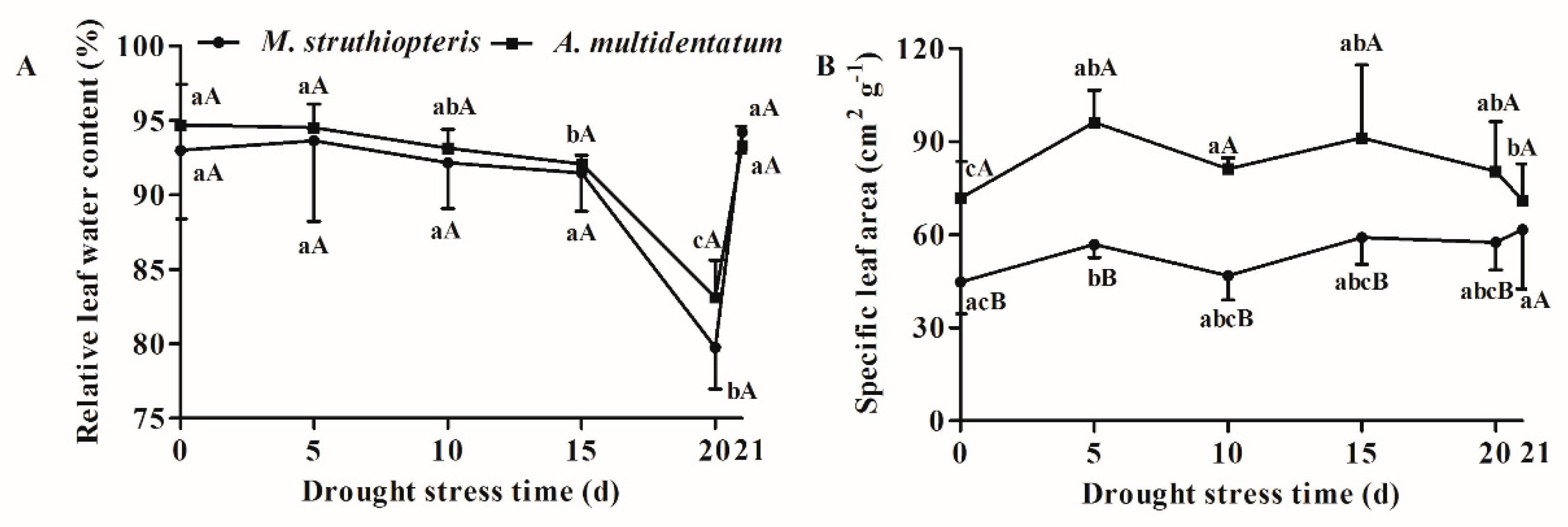
3.1. Growth Traits
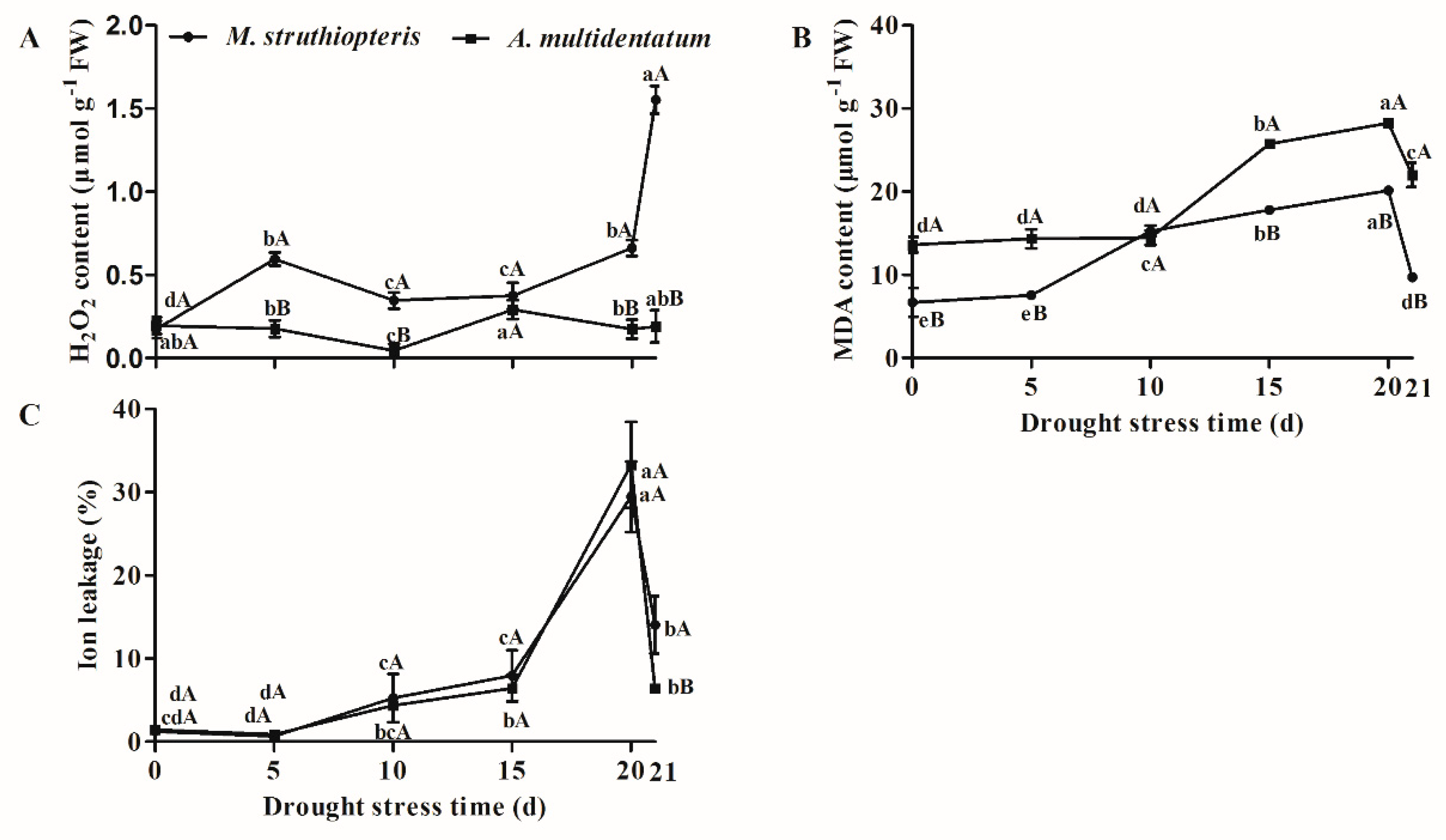
3.2. H2O2, MDA Content and IL
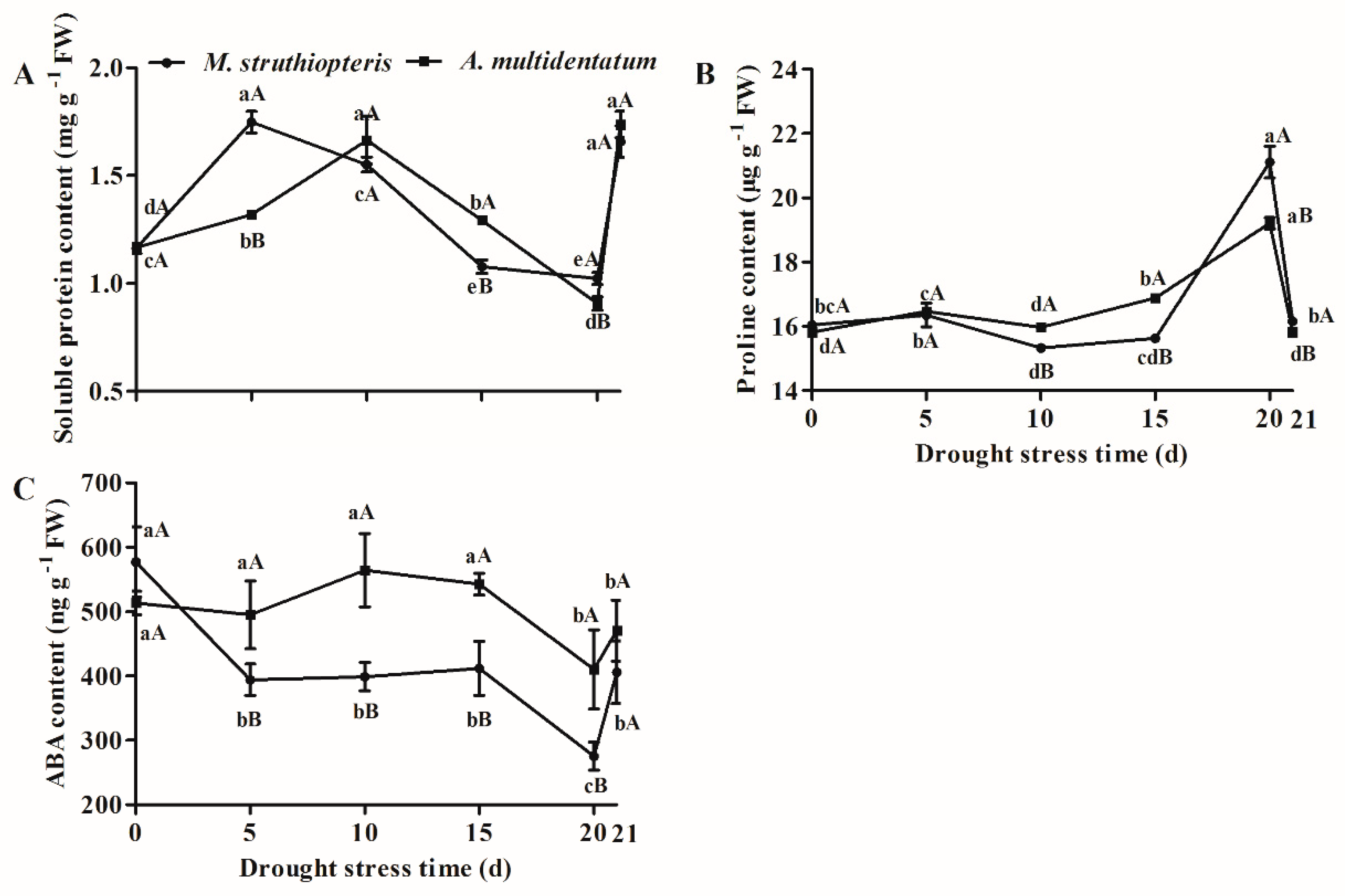
3.3. Osmotic Regulators
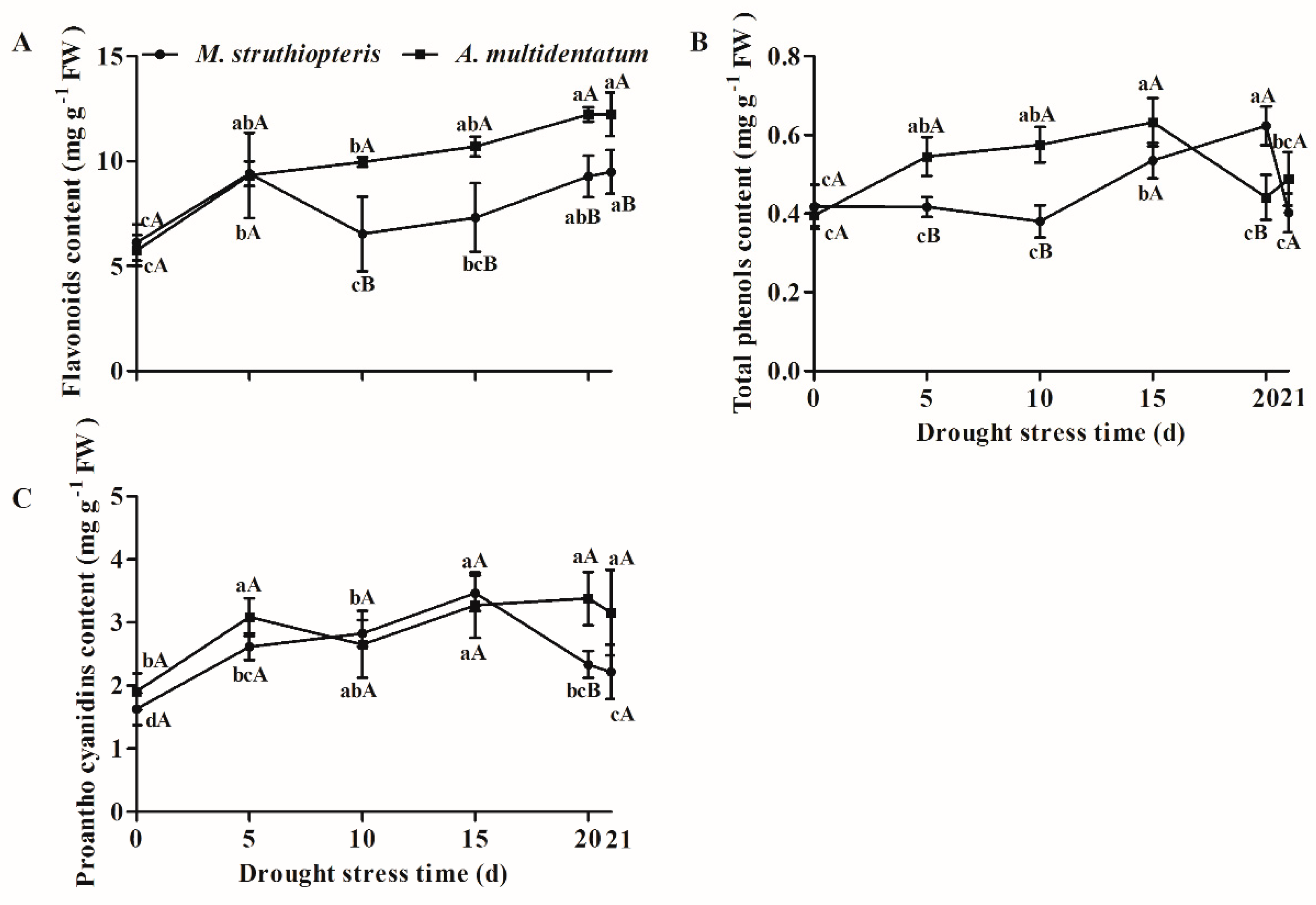
3.4. Secondary Metabolites
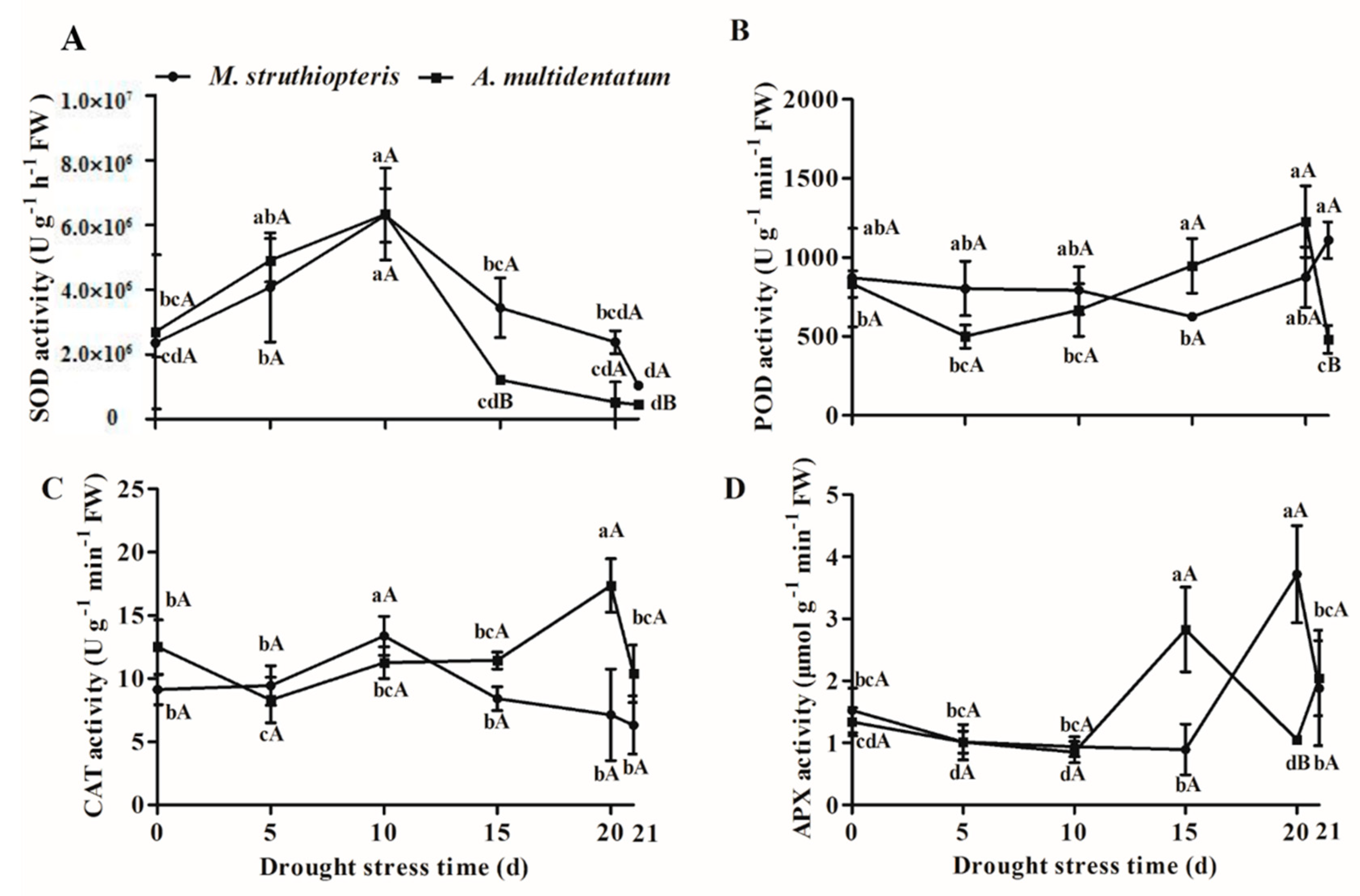
3.5. Antioxidant Enzyme Activities
4. Discussion
4.1. The Responses of Growth Traits to Drought Stress
4.2. The Oxidative Damage of Drought Stress to the Two Fern Species
4.3. The Regulation of Osmotic Regulators to Drought Stress
4.4. The Effects of the Antioxidant System
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Liu, X.; Wang, W.; Zhang, T.; Zeng, X.; Xu, G.; Wu, G.; Kang, H. Spatiotemporal variability of drought in the northern part of Northeast China. Hydrol. Process. 2018, 32, 1449–1460. [Google Scholar] [CrossRef]
- Mishra, A.K.; Singh, V.P. A review of drought concepts. J. Hydrol. 2010, 391, 204–216. [Google Scholar] [CrossRef]
- Li, M.; Hu, W.; Zhang, L.; Wang, G.; Chai, X. Risk analysis of meteorological drought in Northeast China based on standardized precipitation evapotranspiration index. J. Arid. Land Resour. Environ. 2018, 134–139. [Google Scholar] [CrossRef]
- Cai, S.; Zuo, D.; Xu, Z.; Yang, X. Spatiotemporal characteristics of drought in Northeast China based on SPEI. South-North Water Transf. Water Sci. Technol. 2017, 15, 15–21. [Google Scholar] [CrossRef]
- Shen, G.; Zhen, H.; Lei, Z. Spatiotemporal analysis of meteorological drought (1961–2014) in Northeast China using a standardized precipitation evaptranspiration index. Acta Ecol. Sin. 2017, 37, 5882–5893. [Google Scholar] [CrossRef]
- Cheng, L.; Han, M.; Yang, L.-M.; Yang, L.; Sun, Z.; Zhang, T. Changes in the physiological characteristics and baicalin biosynthesis metabolism of Scutellaria baicalensis Georgi under drought stress. Ind. Crop. Prod. 2018, 122, 473–482. [Google Scholar] [CrossRef]
- Selmar, D.; Kleinwächter, M. Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Ind. Crop. Prod. 2013, 42, 558–566. [Google Scholar] [CrossRef]
- Catola, S.; Marino, G.; Emiliani, G.; Huseynova, T.; Musayev, M.; Akparov, Z.; Maserti, B.E. Physiological and metabolomic analysis of Punica granatum (L.) under drought stress. Planta 2016, 243, 441–449. [Google Scholar] [CrossRef]
- Ju, Y.-L.; Yue, X.-F.; Zhao, X.-f.; Zhao, H.; Fang, Y.-l. Physiological, micro-morphological and metabolomic analysis of grapevine (Vitis vinifera L.) leaf of plants under water stress. Plant Physiol. Biochem. 2018, 130, 501–510. [Google Scholar] [CrossRef]
- Li, L.; Liu, Y.; Liu, Y.; He, B.; Wang, M.; Yu, C.; Weng, M. Physiological response and resistance of three cultivars of Acer rubrum L. to continuous drought stress. Acta Ecol. Sin. 2015, 35, 196–202. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, Y.; Taleei, A.; Ranjbar, M. Long-term water deficit modulates antioxidant capacity of peppermint (Mentha piperita L.). Sci. Hortic. 2018, 237, 36–43. [Google Scholar] [CrossRef]
- Khan, M.N.; Zhang, J.; Luo, T.; Liu, J.; Ni, F.; Rizwan, M.; Fahad, S.; Hu, L. Morpho-physiological and biochemical responses of tolerant and sensitive rapeseed cultivars to drought stress during early seedling growth stage. Acta Physiol. Plant. 2019, 41, 25. [Google Scholar] [CrossRef]
- Oral, O.; Kutlu, T.; Aksoy, E.; Fıçıcıoğlu, C.; Uslu, H.; Tuğrul, S. The effects of oxidative stress on outcomes of assisted reproductive techniques. J. Assist. Reprod. Genet. 2006, 23, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-h.; Xu, X.-f.; Sun, Y.-m.; Zhang, J.-l.; Li, C.-z. Influence of drought hardening on the resistance physiology of potato seedlings under drought stress. J. Integr. Agric. 2018, 17, 336–347. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Bozin, B.; Mimica-Dukic, N.; Samojlik, I.; Goran, A.; Igic, R. Phenolics as antioxidants in garlic (Allium sativum L. Alliaceae). Food Chem. 2008, 111, 925–929. [Google Scholar] [CrossRef]
- Mackova, J.; Vaskova, M.; Macek, P.; Hronkova, M.; Schreiber, L.; Santrucek, J. Plant response to drought stress simulated by ABA application: Changes in chemical composition of cuticular waxes. Environ. Exp. Bot. 2013, 86, 70–75. [Google Scholar] [CrossRef]
- Ren, F.; Zhang, R.; Chen, Q.; Bai, Y.; Huang, F.; Li, X. Progress in ABA and SA improving plant drought resistance and salt resistance. Biotechnol. Bull. 2012, 17–21. [Google Scholar] [CrossRef]
- Liu, F.L.; Jensen, C.R.; Shahanzari, A.; Andersen, M.N.; Jacobsen, S.E. ABA regulated stomatal control and photosynthetic water use efficiency of potato (Solanum tuberosum L.) during progressive soil drying. Plant Sci. 2005, 168, 831–836. [Google Scholar] [CrossRef]
- Ban, S.G.; Selak, G.V.; Leskovar, D.I. Short- and long-term responses of pepper seedlings to ABA exposure. Sci. Hortic. 2017, 225, 243–251. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, F.; Li, M.; Liang, D.; Zou, J. Physiological responses of kiwifruit plants to exogenous ABA under drought conditions. Plant Growth Regul. 2011, 64, 63–74. [Google Scholar] [CrossRef]
- Liao, J.-X.; Jiang, M.-X.; Huang, H.-D. Growth characteristics of Adiantum reniforme var. sinensis and A-capillus-veneris in response to light and soil moisture. Nord. J. Bot. 2013, 31, 500–504. [Google Scholar] [CrossRef]
- Liao, J.X.; Zhang, H.; Mo, L.; Huang, Y.Q.; Sun, Y.J.; Li, Y.Q. Differences in growth and biomass allocation of Adiantum flabellulatum and A. capillus-veneris as a result of light and water availability. Bot. Lett. 2017, 164, 393–400. [Google Scholar] [CrossRef]
- Nishida, K.; Hanba, Y.T. Photosynthetic response of four fern species from different habitats to drought stress: Relationship between morpho-anatomical and physiological traits. Photosynthetica 2017, 55, 689–697. [Google Scholar] [CrossRef]
- Wu, Y.C.; He, Y.J.; Gao, J. Drought tolerance of three kinds of wild ornamental ferns. Hubei For. Sci. Technol. 2015, 44, 17–21. [Google Scholar]
- Li, Q.J.; Zhong, Q.Y.; Shao, L.; Ji, H.; Chen, L.Q. Effect of PEG stress on drought resistance physiological indices of two ferns’ prothallus. Chin. Agric. Sci. Bull. 2014, 30, 155–160. [Google Scholar]
- Sun, H.; Wang, Q.; Guan, Y.; Liu, B.D. Effects of Microlepia strigosa under drought stress on physiological change laws. Plant Sci. J. 2013, 31, 576–582. [Google Scholar] [CrossRef]
- Dong, Y.L.; Chang, Y. Effects of drought stress on chlorophyll fluorescence characteristics of five species of ornamental ferns. North. Hortic. 2013, 15, 66–69. [Google Scholar]
- Zhu, L.-J.; Yan, F.; Chen, J.-P.; Zhang, N.; Zhang, X.; Yao, X.-S. 8-O-4′ Neolignan glycosides from the aerial parts of Matteuccia struthiopteris. Chin. Chem. Lett. 2016, 27, 63–65. [Google Scholar] [CrossRef]
- Han, X.-Z.; Ma, R.; Chen, Q.; Jin, X.; Jin, Y.-Z.; An, R.-B.; Piao, X.-M.; Lian, M.-L.; Quan, L.-H.; Jiang, J. Anti-inflammatory action of Athyrium multidentatum extract suppresses the LPS-induced TLR4 signaling pathway. J. Ethnopharmacol. 2018, 217, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Fu, P. Clavis Plantarum Chinae Boreali-Orientalis; Science Press: Beijing, China, 1995; pp. 35–46. [Google Scholar]
- Qi, G.; Yang, L.; Xiao, C.; Shi, J.; Mi, Y.; Liu, X. Nutrient values and bioactivities of the extracts from three fern species in China: A comparative assessment. Food Funct. 2015, 6, 2918–2929. [Google Scholar] [CrossRef]
- Xu, J.; Wei, L.; Xu, H.; Gao, D. Effects of different ecological factors on growth and photosynthetic characteristics of Matteuccia struthiopteris. Chin. Agric. Sci. Bull. 2017, 33, 19–24. [Google Scholar]
- Zhang, Y.; Yang, L. Investigation on Athyrium multidentatum (Doll.) Ching in different habitats. Chin. Wild Plant Resour. 2011, 30, 5–8. [Google Scholar]
- Xiao, Q.; Ye, W.; Zhu, Z.; Chen, Y.; Zhen, H. A simple non-destructive method to measure leaf area using digital camera and Photoshop software. Chin. J. Ecol. 2005, 711–714. [Google Scholar] [CrossRef]
- Li, X.Y.; Liu, G.S.; Yang, Y.F.; Zhao, C.H.; Yu, Q.W.; Song, S.X. Relationship between hyperspectral parameters and physiological and biochemical indexes of flue-cured tobacco leaves. Agric. Sci. China 2007, 665–672. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implication of hydrogen peroxide-ascorbate system on membrane permeability of water stressed Vigna seedlings. New Phytol. 1985, 99, 355–360. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Liu, J.J.; Wei, Z.; Li, J.H. Effects of copper on leaf membrane structure and root activity of maize seedling. Bot. Stud. 2014, 55. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Emerich, D.W.; Sanchezdiaz, M. Water-stress induced changes in concentrations of proline and total soluble sugars in Nodulated Alfalfa (Medicag0-astiva) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Ali, A.; Ong, M.K.; Forney, C.F. Effect of ozone pre-conditioning on quality and antioxidant capacity of papaya fruit during ambient storage. Food Chem. 2014, 142, 19–26. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought Stress Effects on Growth, ROS Markers, Compatible Solutes, Phenolics, Flavonoids, and Antioxidant Activity in Amaranthus tricolor. Appl. Biochem. Biotechnol. 2018, 186, 999–1016. [Google Scholar] [CrossRef]
- Zhang, D.; Ren, J.; Liu, H.M.; Wang, H.M. Responses of main secondary metabolites and DPPH free radical scavenging activity of the Korean Pine to drought stress. Bull. Bot. Res. 2016, 36, 542–548. [Google Scholar]
- Azevedo, R.A.; Alas, R.M.; Smith, R.J.; Lea, P.J. Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and roots of wild-type and a catalase-deficient mutant of barley. Physiol. Plant. 1998, 104, 280–292. [Google Scholar] [CrossRef]
- Zhou, W.; Leul, M. Uniconazole-induced tolerance of rape plants to heat stress in relation to changes in hormonal levels, enzyme activities and lipid peroxidation. Plant Growth Regul. 1999, 27, 99–104. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: II. Purification and quantitative relationship with water-soluble protein in seedlings. Plant Physiol. 1977, 59, 315–318. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific peroxidase in Spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Okunlola, G.O.; Olatunji, O.A.; Akinwale, R.O.; Tariq, A.; Adelusi, A.A. Physiological response of the three most cultivated pepper species (Capsicum spp.) in Africa to drought stress imposed at three stages of growth and development. Sci. Hortic. 2017, 224, 198–205. [Google Scholar] [CrossRef]
- Hasan, M.M.-U.; Ma, F.; Prodhan, Z.H.; Li, F.; Shen, H.; Chen, Y.; Wang, X. Molecular and physio-biochemical characterization of cotton species for assessing drought stress tolerance. Int. J. Mol. Sci. 2018, 19, 2636. [Google Scholar] [CrossRef]
- Ravikumar, G.; Manimaran, P.; Voleti, S.R.; Subrahmanyam, D.; Sundaram, R.M.; Bansal, K.C.; Viraktamath, B.C.; Balachandran, S.M. Stress-inducible expression of AtDREB1A transcription factor greatly improves drought stress tolerance in transgenic indica rice. Transgenic Res. 2014, 23, 421–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dossa, K.; Yehouessi, L.W.; Likeng-Li-Ngue, B.C.; Diouf, D.; Liao, B.S.; Zhang, X.R.; Cisse, N.; Bell, J.M. Comprehensive screening of some west and central African sesame genotypes for drought resistance probing by agromorphological, physiological, biochemical and seed quality traits. Agronomy 2017, 7, 83. [Google Scholar] [CrossRef]
- Budak, H.; Kantar, M.; Kurtoglu, K.Y. Drought tolerance in modern and wild wheat. Sci. World J. 2013, 2013, 548246. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.W.; Sun, X.W.; Li, R.X.; Liu, G. The investigate and analyze in habitat of Matteuccia struthiopteris Todaro. Territ. Nat. Resour. Study 2007, 92–93. [Google Scholar]
- Ghalambor, C.K.; McKay, J.K.; Carroll, S.P.; Reznick, D.N. Adaptive versus non-adaptive phenotypic plasticity and the potential for contemporary adaptation in new environments. Funct. Ecol. 2007, 21, 394–407. [Google Scholar] [CrossRef] [Green Version]
- Auslander, M.; Nevo, E.; Inbar, M. The effects of slope orientation on plant growth, developmental instability and susceptibility to herbivores. J. Arid Environ. 2003, 55, 405–416. [Google Scholar] [CrossRef]
- Bongers, F.J.; Olmo, M.; Lopez-Iglesias, B.; Anten, N.P.R.; Villar, R. Drought responses, phenotypic plasticity and survival of Mediterranean species in two different microclimatic sites. Plant Biol. 2017, 19, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Lortie, C.J.; Aarssen, L.W. The specialization hypothesis for phenotypic plasticity in plants. Int. J. Plant Sci. 1996, 157, 484–487. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, F.; Liu, L.; Zhu, S. Physiological responses of Leucaena leucocephala seedlings to drought stress. In Proceedings of the 2012 International Conference on Modern Hydraulic Engineering, Nanjing, China, 9–11 March 2012; Zhou, B., Ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar] [CrossRef]
- Marshall, J.G.; Rutledge, R.G.; Blumwald, E.; Dumbroff, E.B. Reduction in turgid water volume in jack pine, white spruce and black spruce in response to drought and paclobutrazol. Tree Physiol. 2000, 20, 701–707. [Google Scholar] [CrossRef] [Green Version]
- Borrajo, C.I.; Sanchez-Moreiras, A.M.; Reigosa, M.J. Morpho-physiological responses of tall wheatgrass populations to different levels of water stress. PLoS ONE 2018, 13, e0209281. [Google Scholar] [CrossRef]
- Pagter, M.; Bragato, C.; Brix, H. Tolerance and physiological responses of Phragmites australis to water deficit. Aquat. Bot. 2005, 81, 285–299. [Google Scholar] [CrossRef]
- Meher; Shivakrishna, P.; Ashok Reddy, K.; Manohar Rao, D. Effect of PEG-6000 imposed drought stress on RNA content, relative water content (RWC), and chlorophyll content in peanut leaves and roots. Saudi J. Biol. Sci. 2018, 25, 285–289. [Google Scholar] [CrossRef]
- Bayoumi, T.Y.; Eid, M.H.; Metwali, E.M. Application of physiological and biochemical indices as a screening technique for drought tolerance in wheat genotypes. Afr. J. Biotechnol. 2008, 7, 2341–2352. [Google Scholar] [CrossRef]
- Deng, H.M.; Long, C.Y.; Cai, S.Z.; Song, Y.; Yan, R.X.; Che, Y.R.; Wang, C.J.; Xiao, Y. Morphology and physiological characteristics of Stachys lanata seedling under water stress. Acta Bot. Boreali-Occident. Sin. 2018, 38, 1099–1108. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Ač, A.; Marek, M.V.; Kalina, J.; Urban, O. Differences in pigment composition, photosynthetic rates and chlorophyll fluorescence images of sun and shade leaves of four tree species. Plant Physiol. Biochem. 2007, 45, 577–588. [Google Scholar] [CrossRef]
- Lindsey, A.J.; Barker, D.J.; Metzger, J.D.; Mullen, R.W.; Thomison, P.R. Physiological and morphological response of a drought-tolerant maize hybrid to agronomic management. Agron. J. 2018, 110, 1354–1362. [Google Scholar] [CrossRef]
- Diaz-Lopez, L.; Gimeno, V.; Simon, I.; Martinez, V.; Rodriguez-Ortega, W.M.; Garcia-Sanchez, F. Jatropha curcas seedlings show a water conservation strategy under drought conditions based on decreasing leaf growth and stomatal conductance. Agric. Water Manag. 2012, 105, 48–56. [Google Scholar] [CrossRef]
- Ludewig, K.; Hanke, J.M.; Wuthe, B.; Otte, A.; Mosner, E.; Eckstein, R.L.; Donath, T.W. Differential effect of drought regimes on the seedling performance of six floodplain grassland species. Plant Biol. 2018, 20, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef]
- Upadhyaya, H.; Khan, M.H.; Panda, S.K. Hydrogen peroxide induces oxidative stress in detached leaves of Oryza sativa L. Plant Physiol. 2006, 33, 83–95. [Google Scholar]
- Zhao, H.-J.; Tan, J.-F. Role of calcium ion in protection against heat and high irradiance stress-induced oxidative damage to photosynthesis of wheat leaves. Photosynthetica 2005, 43, 473–476. [Google Scholar] [CrossRef]
- Liu, R.; Shi, H.; Wang, Y.; Chen, S.; Deng, J.; Liu, Y.; Li, S.; Chan, Z. Comparative physiological analysis of lotus (Nelumbo nucifera) cultivars in response to salt stress and cloning of NnCIPK genes. Sci. Hortic. 2014, 173, 29–36. [Google Scholar] [CrossRef]
- Chai, Y.; Zhang, S.; Yao, C. Effects of drought stress and rewatering on photosynthetic physiological characteristics of sorghum. In Proceedings of the IEEE 2011 International Conference on Remote Sensing, Environment and Transportation Engineering, Nanjing, China, June 2011. [Google Scholar] [CrossRef]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Goli, S.A.H. Effect of drought stress on total phenolic, lipid peroxidation, and antioxidant activity of Achillea species. Appl. Biochem. Biotechnol. 2016, 178, 796–809. [Google Scholar] [CrossRef]
- Nie, W.; Gong, B.; Chen, Y.; Wang, J.; Wei, M.; Shi, Q. Photosynthetic capacity, ion homeostasis and reactive oxygen metabolism were involved in exogenous salicylic acid increasing cucumber seedlings tolerance to alkaline stress. Sci. Hortic. 2018, 235, 413–423. [Google Scholar] [CrossRef]
- Jin, R.; Shi, H.; Han, C.; Zhong, B.; Wang, Q.; Chan, Z. Physiological changes of purslane (Portulaca oleracea L.) after progressive drought stress and rehydration. Sci. Hortic. 2015, 194, 215–221. [Google Scholar] [CrossRef]
- Filippou, P.; Antoniou, C.; Fotopoulos, V. Effect of drought and rewatering on the cellular status and antioxidant response of Medicago truncatula plants. Plant Signal. Behav. 2011, 6, 270–277. [Google Scholar] [CrossRef]
- Sahin, U.; Ekinci, M.; Ors, S.; Turan, M.; Yildiz, S.; Yildirim, E. Effects of individual and combined effects of salinity and drought on physiological, nutritional and biochemical properties of cabbage (Brassica oleracea var. capitata). Sci. Hortic. 2018, 240, 196–204. [Google Scholar] [CrossRef]
- Ekinci, M.; Ors, S.; Sahin, U.; Yildirim, E.; Dursun, A. Responses to the irrigation water amount of spinach supplemented with organic amendment in greenhouse conditions. Commun. Soil Sci. Plant Anal. 2015, 46, 327–342. [Google Scholar] [CrossRef]
- Anamul Hoque, M.; Okuma, E.; Nasrin Akhter Banu, M.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Exogenous proline mitigates the detrimental effects of salt stress more than exogenous betaine by increasing antioxidant enzyme activities. J. Plant Physiol. 2007, 164, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yuan, J.; Luo, A.; Chen, Y.; Fan, Y. Drought stress and re-watering increase secondary metabolites and enzyme activity in dendrobium moniliforme. Ind. Crop. Prod. 2016, 94, 385–393. [Google Scholar] [CrossRef]
- Sauter, A.; Davies, W.J.; Hartung, W. The long-distance abscisic acid signal in the droughted plant: The fate of the hormone on its way from root to shoot. J. Exp. Bot. 2001, 52, 1991–1997. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R.R.; Gampala, S.S.L.; Rock, C.D. Abscisic acid signaling in seeds and seedlings. Plant Cell 2002, 14 (Suppl. l), S15–S45. [Google Scholar] [CrossRef]
- Dar, N.A.; Amin, I.; Wani, W.; Wani, S.A.; Shikari, A.B.; Wani, S.H.; Masoodi, K.Z. Abscisic acid: A key regulator of abiotic stress tolerance in plants. Plant Gene 2017, 11, 106–111. [Google Scholar] [CrossRef]
- Koffler, B.E.; Luschin-Ebengreuth, N.; Stabentheiner, E.; Mueller, M.; Zechmann, B. Compartment specific response of antioxidants to drought stress in Arabidopsis. Plant Sci. 2014, 227, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, F.G.; Peng, S.M.; Ma, W.J.; Korpelainen, H.; Li, C.Y. Comparative physiological, ultrastructural and proteomic analyses reveal sexual differences in the responses of Populus cathayana under drought stress. Proteomics 2010, 10, 2661–2677. [Google Scholar] [CrossRef] [PubMed]
- Pavlovic, I.; Petrik, I.; Tarkowska, D.; Lepedus, H.; Vujcic Bok, V.; Radic Brkanac, S.; Novak, O.; Salopek-Sondi, B. Correlations between phytohormones and drought tolerance in selected Brassica crops: Chinese cabbage, white cabbage and kale. Int. J. Mol. Sci. 2018, 19, 2866. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.; Carvalho, M.E.A.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges—A little help from the antioxidant networks. Environ. Exp. Bot. 2018. [Google Scholar] [CrossRef]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Amaral Carvalho, M.E.; Piotto, F.A.; Franco, M.R.; Reis Borges, K.L.; Gaziola, S.A.; Camargo Castro, P.R.; Azevedo, R.A. Cadmium toxicity degree on tomato development is associated with disbalances in B and Mn status at early stages of plant exposure. Ecotoxicology 2018, 27, 1293–1302. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.-M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, I.; Alegre, L.; Van Breusegem, F.; Munne-Bosch, S. How relevant are flavonoids as antioxidants in plants? Trends Plant Sci. 2009, 14, 125–132. [Google Scholar] [CrossRef]
- Wang, C.; Lu, J.; Zhang, S.; Wang, P.; Hou, J.; Qian, J. Effects of Pb stress on nutrient uptake and secondary metabolism in submerged macrophyte Vallisneria natans. Ecotoxicol. Environ. Saf. 2011, 74, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Hodaei, M.; Rahimmalek, M.; Arzani, A.; Talebi, M. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crop. Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Dos Santos, I.C.; de Almeida, A.-A.F.; Pirovani, C.P.; Costa, M.G.C.; das Gracas Fernandes da Silva, M.F.; Bellete, B.S.; Freschi, L.; Soares Filho, W.; Coelho Filho, M.A.; Gesteira, A.d.S. Differential accumulation of flavonoids and phytohormones resulting from the canopy/rootstock interaction of citrus plants subjected to dehydration/rehydration. Plant Physiol. Biochem. 2017, 119, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, X.; Wang, D.; Zou, Z.; Liang, Z. Effect of drought stress on growth and accumulation of active constituents in Salvia miltiorrhiza Bunge. Ind. Crop. Prod. 2011, 33, 84–88. [Google Scholar] [CrossRef]
- de Abreu, I.N.; Mazzafera, P. Effect of water and temperature stress on the content of active constituents of Hypericum brasiliense Choisy. Plant Physiol. Biochem. 2005, 43, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Helaly, M.N.; El-Hoseiny, H.; El-Sheery, N.I.; Rastogi, A.; Kalaji, H.M. Regulation and physiological role of silicon in alleviating drought stress of mango. Plant Physiol. Biochem. 2017, 118, 31–44. [Google Scholar] [CrossRef]
- Pang, J.; Hao, L.; Zhang, F.; Zhao, P.; Yang, Z. The response of active oxygen species and ascorbic acid in Pugionium cornutum (L.) Gaertn. leaves to drought stress. Plant Physiol. J. 2013, 49, 57–62. [Google Scholar] [CrossRef]
- Ji, Y.; Zhang, X.Q.; Peng, Y.; Liang, X.Y.; Huang, L.K.; Ma, X.; Ma, Y.M. Effects of drought stress on lipid peroxidation, osmotic adjustment and activities of protective enzymes in the roots and leaves of orchardgrass. Acta Prataculturae Sin. 2014, 23, 144–151. [Google Scholar] [CrossRef]
- Chool Boo, Y.; Jung, J. Water Deficit—Induced oxidative stress and antioxidative defenses in rice plants. J. Plant Physiol. 1999, 155, 255–261. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Guo, K.; Fan, D.; Li, G.; Zheng, Y.; Yu, L.; Yang, R. Effect of drought on pigments, osmotic adjustment and antioxidant enzymes in six woody plant species in karst habitats of southwestern China. Environ. Exp. Bot. 2011, 71, 174–183. [Google Scholar] [CrossRef]
- Weydert, C.J.; Cullen, J.J. Measurement of superoxide dismutase, catalase and glutathione peroxidase in cultured cells and tissue. Nat. Protoc. 2010, 5, 51–66. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Gao, S.; He, X.; Li, Y.; Li, P.; Zhang, Y.; Chen, W. Growth, Secondary Metabolites and Enzyme Activity Responses of Two Edible Fern Species to Drought Stress and Rehydration in Northeast China. Agronomy 2019, 9, 137. https://doi.org/10.3390/agronomy9030137
Wang Y, Gao S, He X, Li Y, Li P, Zhang Y, Chen W. Growth, Secondary Metabolites and Enzyme Activity Responses of Two Edible Fern Species to Drought Stress and Rehydration in Northeast China. Agronomy. 2019; 9(3):137. https://doi.org/10.3390/agronomy9030137
Chicago/Turabian StyleWang, Yanlin, Shanshan Gao, Xingyuan He, Yan Li, Peiyang Li, Yue Zhang, and Wei Chen. 2019. "Growth, Secondary Metabolites and Enzyme Activity Responses of Two Edible Fern Species to Drought Stress and Rehydration in Northeast China" Agronomy 9, no. 3: 137. https://doi.org/10.3390/agronomy9030137
APA StyleWang, Y., Gao, S., He, X., Li, Y., Li, P., Zhang, Y., & Chen, W. (2019). Growth, Secondary Metabolites and Enzyme Activity Responses of Two Edible Fern Species to Drought Stress and Rehydration in Northeast China. Agronomy, 9(3), 137. https://doi.org/10.3390/agronomy9030137