
Cellular Sources and Neuroprotective Roles of Interleukin-10 in the Facial Motor Nucleus after Axotomy
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Genomic DNA and Polymerase Chain Reaction (PCR)
2.3. Adoptive Transfer
2.4. Induction of Cre Recombinase
2.5. Facial Nerve Axotomy
2.6. Motoneuron Counting
2.7. Immunohistochemistry
2.7.1. IL-10/GFP Reporter
2.7.2. Immunohistochemistry Performed on Wild-Type Tissue
2.8. Fluorescent In Situ Hybridization
3. Results
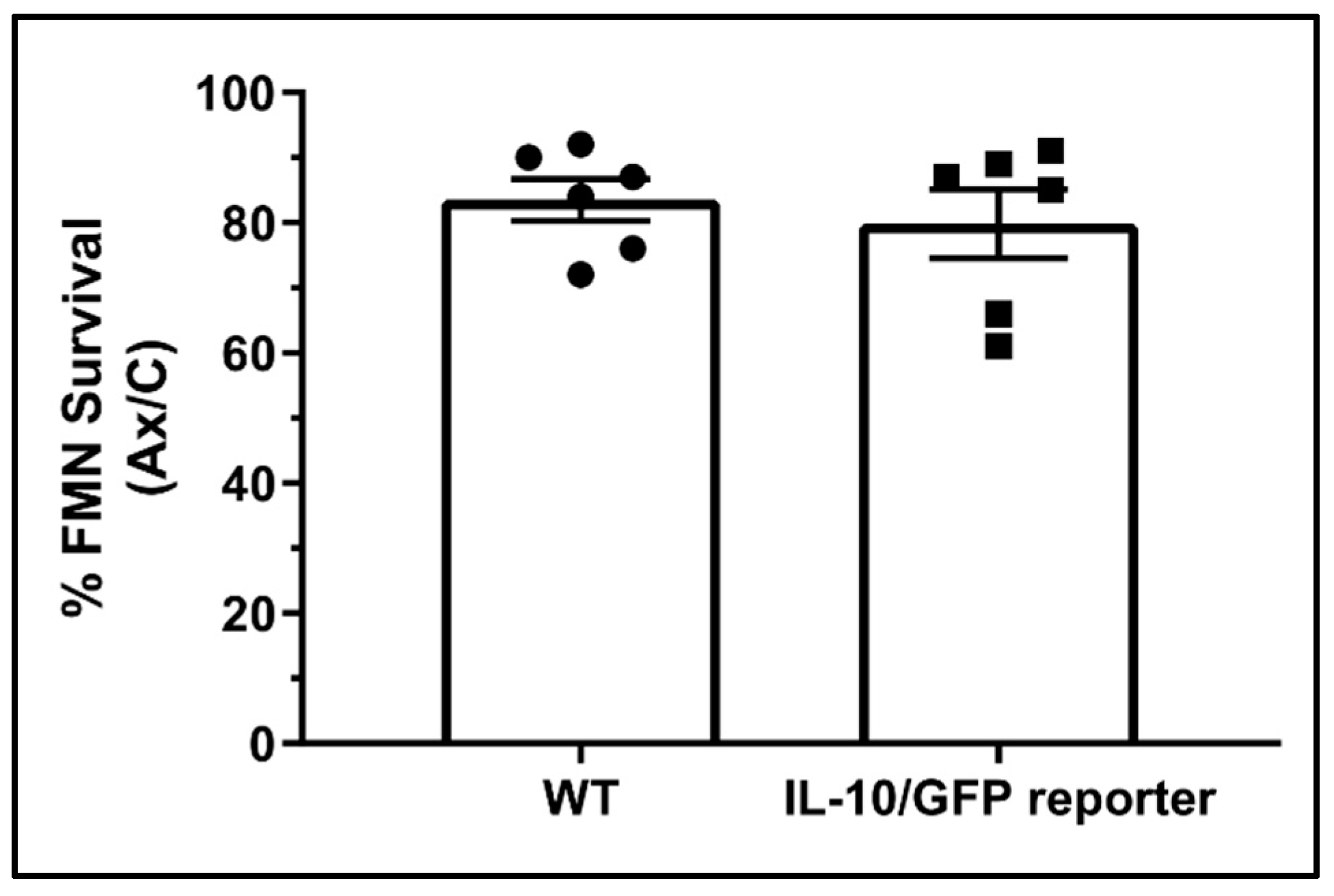
3.1. Contribution of IL-10 from Peripheral Immune Cells to Central Facial Motoneuron Survival
3.2. IL-10/GFP Reporter Characterization
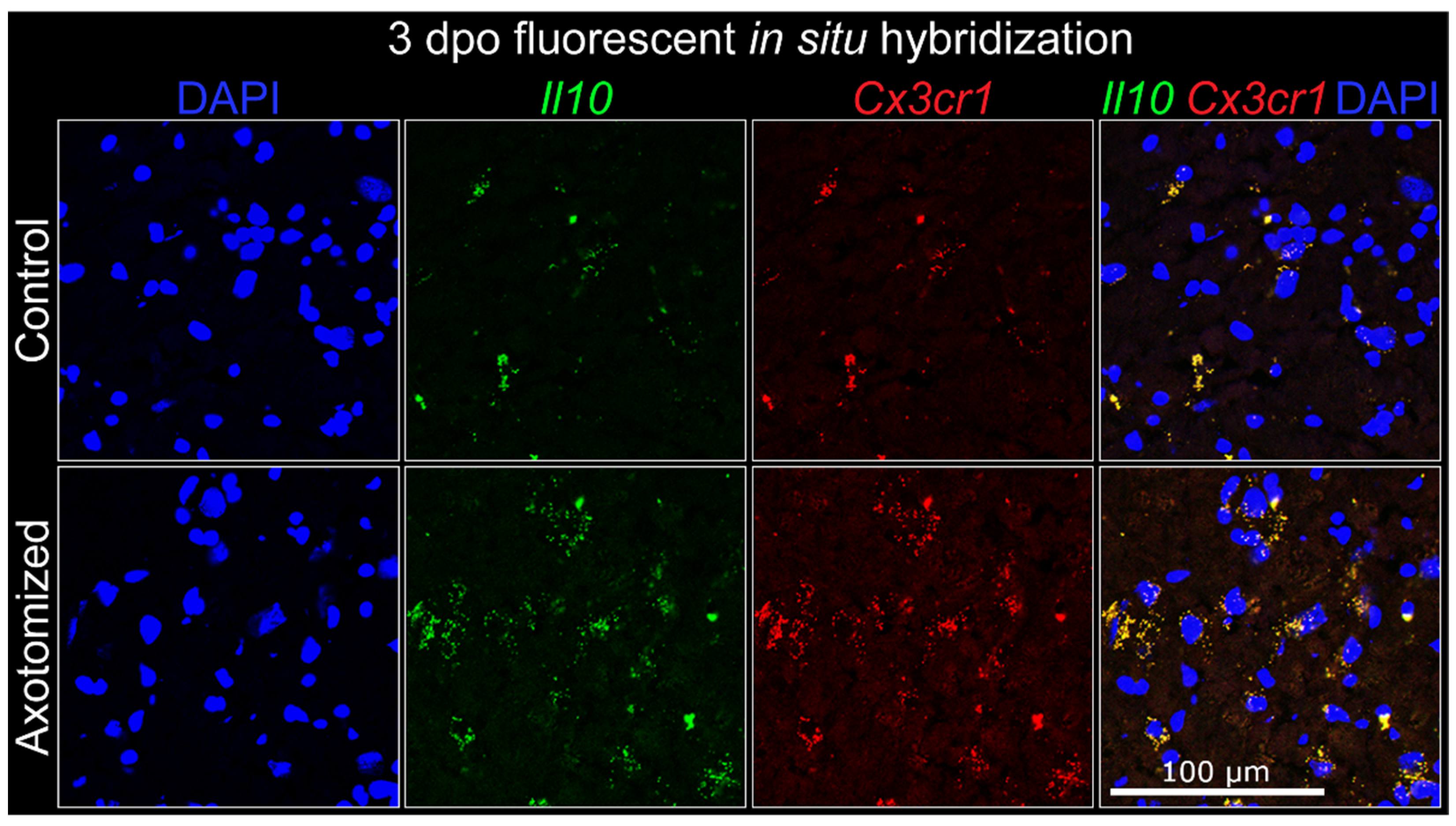
3.3. Central Cellular Localization of IL-10
3.3.1. Neurons
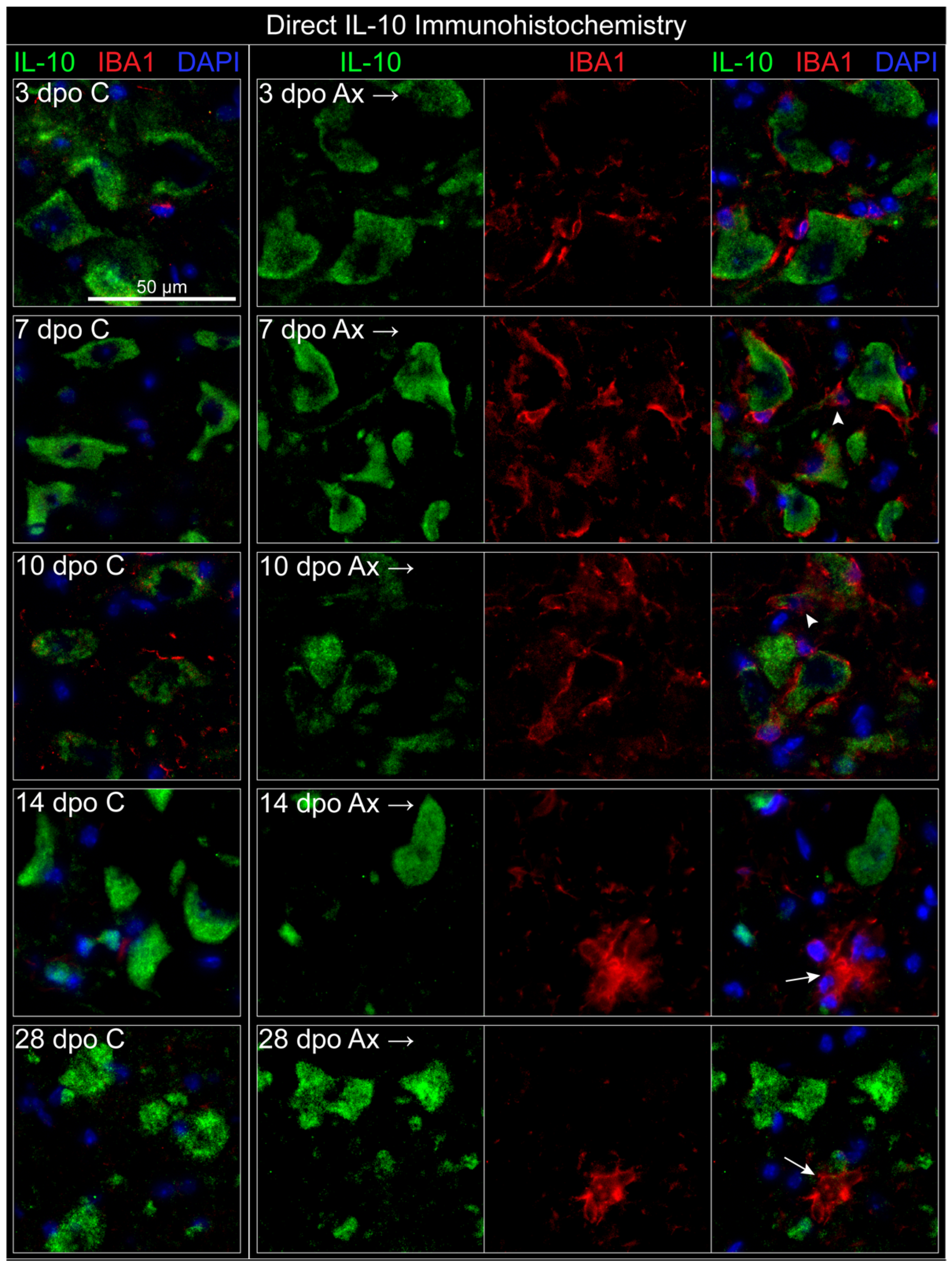
3.3.2. Microglia
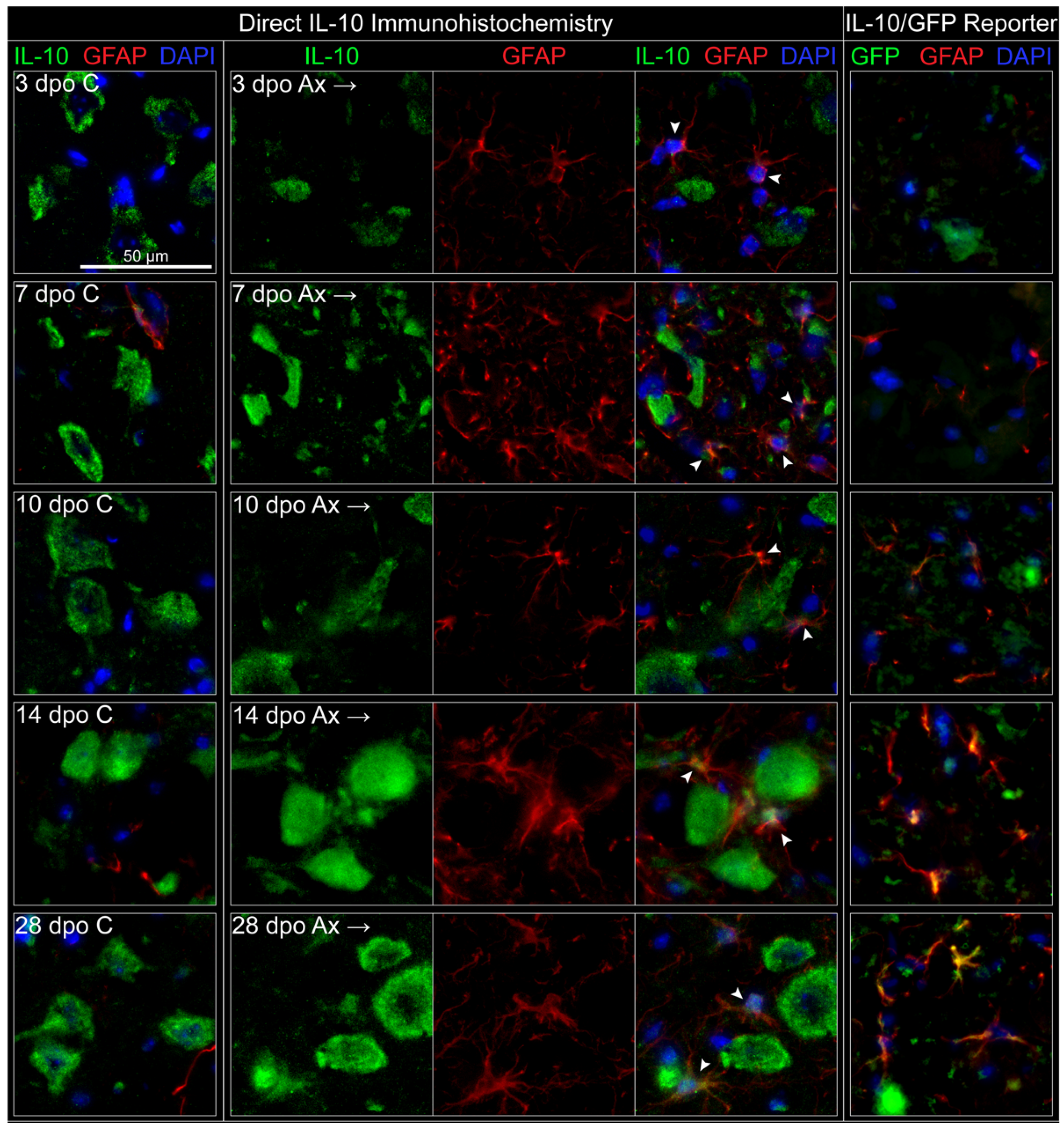
3.3.3. Astrocytes
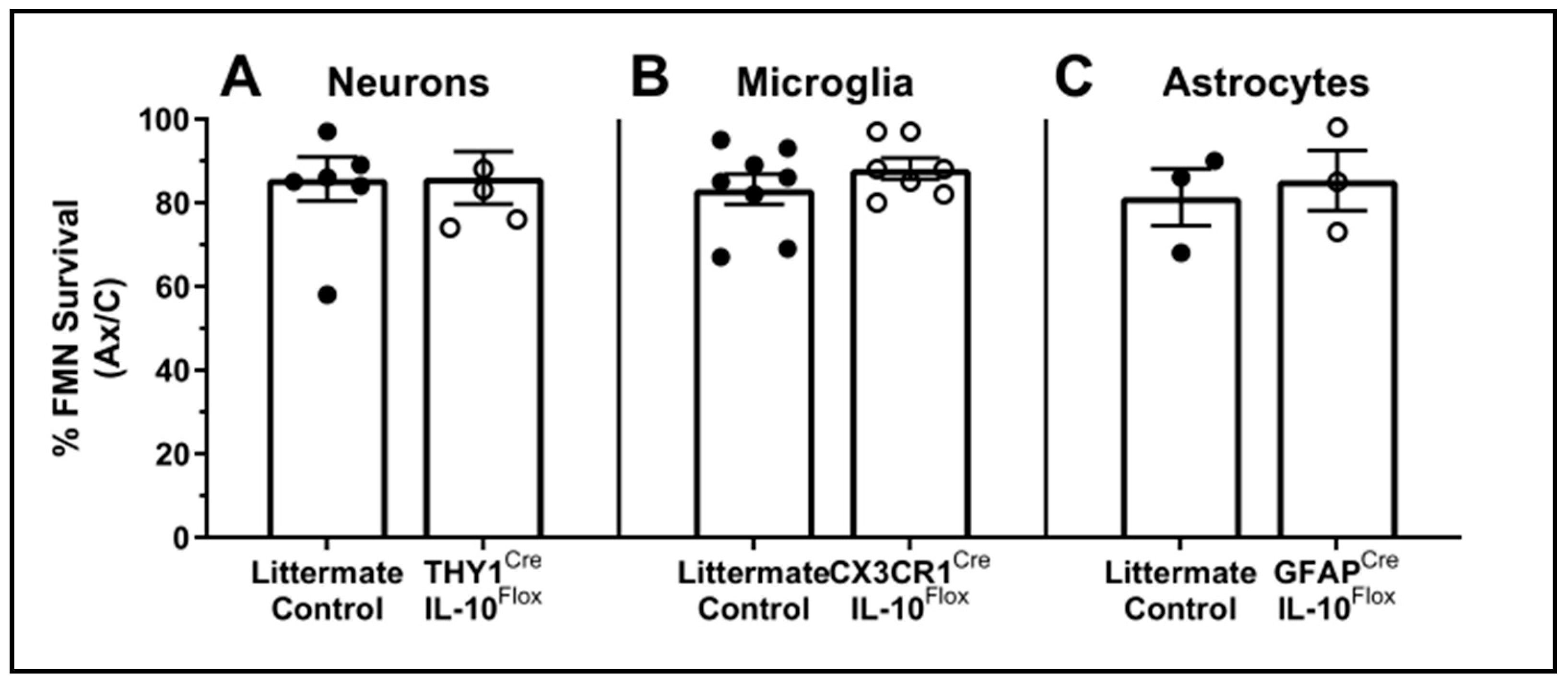
3.4. Cell-Specific Conditional IL-10 Knockout
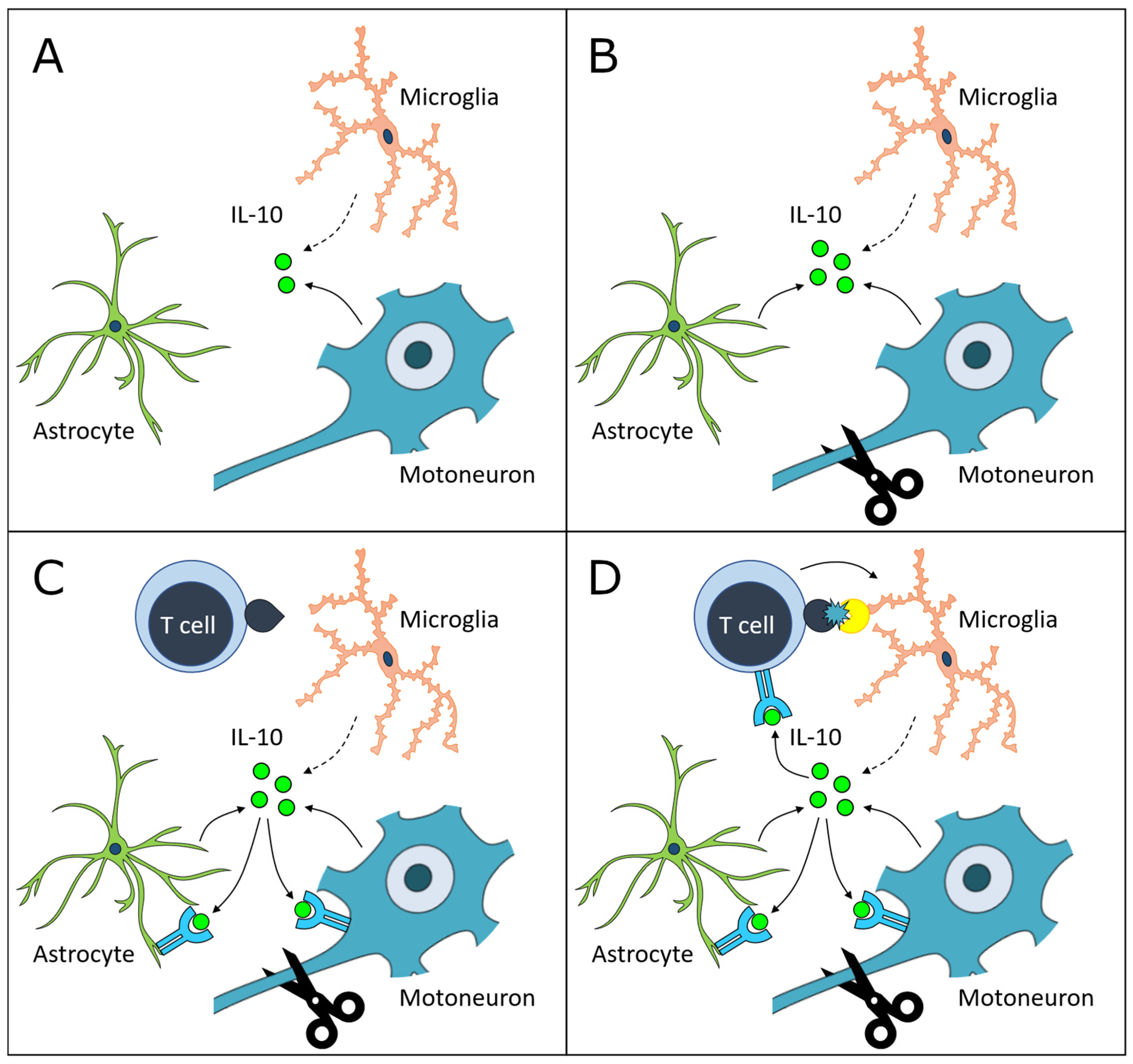
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xin, J.; Wainwright, D.A.; Mesnard, N.A.; Serpe, C.J.; Sanders, V.M.; Jones, K.J. IL-10 within the CNS is necessary for CD4+ T cells to mediate neuroprotection. Brain Behav. Immun. 2011, 25, 820–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serpe, C.J.; Kohm, A.P.; Huppenbauer, C.B.; Sanders, V.M.; Jones, K.J. Exacerbation of facial motoneuron loss after facial nerve transection in severe combined immunodeficient (scid) mice. J. Neurosci. 1999, 19, RC7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serpe, C.J.; Sanders, V.M.; Jones, K.J. Kinetics of facial motoneuron loss following facial nerve transection in severe combined immunodeficient mice. J. Neurosci. Res. 2000, 62, 273–278. [Google Scholar] [CrossRef]
- Serpe, C.J.; Coers, S.; Sanders, V.M.; Jones, K.J. CD4+ T, but not CD8+ or B, lymphocytes mediate facial motoneuron survival after facial nerve transection. Brain Behav. Immun. 2003, 17, 393–402. [Google Scholar] [CrossRef]
- Byram, S.C.; Carson, M.J.; DeBoy, C.A.; Serpe, C.J.; Sanders, V.M.; Jones, K.J. CD4-positive T cell-mediated neuroprotection requires dual compartment antigen presentation. J. Neurosci. 2004, 24, 4333–4339. [Google Scholar] [CrossRef] [Green Version]
- Wainwright, D.A.; Mesnard, N.A.; Xin, J.; Sanders, V.M.; Jones, K.J. Effects of facial nerve axotomy on Th2-associated and Th1-associated chemokine mRNA expression in the facial motor nucleus of wild-type and presymptomatic SOD1 mice. J. Neurodegener. Regen. 2009, 2, 39–44. [Google Scholar]
- Wainwright, D.A.; Xin, J.; Mesnard, N.A.; Beahrs, T.R.; Politis, C.M.; Sanders, V.M.; Jones, K.J. Exacerbation of facial motoneuron loss after facial nerve axotomy in CCR3-deficient mice. ASN Neuro 2009, 1, e00024. [Google Scholar] [CrossRef]
- Wainwright, D.A.; Xin, J.; Mesnard, N.A.; Politis, C.M.; Sanders, V.M.; Jones, K.J. Effects of facial nerve axotomy on Th2- and Th1-associated chemokine expression in the facial motor nucleus of wild-type and presymptomatic mSOD1 mice. J. Neuroimmunol. 2009, 216, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Runge, E.M.; Iyer, A.K.; Setter, D.O.; Kennedy, F.M.; Sanders, V.M.; Jones, K.J. CD4+ T cell expression of the IL-10 receptor is necessary for facial motoneuron survival after axotomy. J. Neuroinflamm. 2020, 17, 121. [Google Scholar] [CrossRef]
- Roers, A.; Siewe, L.; Strittmatter, E.; Deckert, M.; Schlüter, D.; Stenzel, W.; Gruber, A.D.; Krieg, T.; Rajewsky, K.; Muller, W. T cell-specific inactivation of the interleukin 10 gene in mice results in enhanced T cell responses but normal innate responses to lipopolysaccharide or skin irritation. J. Exp. Med. 2004, 200, 1289–1297. [Google Scholar] [CrossRef]
- Olmstead, D.N.; Mesnard-Hoaglin, N.A.; Batka, R.J.; Haulcomb, M.M.; Miller, W.M.; Jones, K.J. Facial nerve axotomy in mice: A model to study motoneuron response to injury. J. Vis. Exp. 2015, e52382. [Google Scholar] [CrossRef] [PubMed]
- Kiernan, J.A. Fluorescence of GFP in sections of fixed tissue. Biotech. Histochem. 2006, 81, 159. [Google Scholar] [CrossRef] [PubMed]
- Swenson, E.S.; Price, J.G.; Brazelton, T.; Krause, D.S. Limitations of green fluorescent protein as a cell lineage marker. Stem Cells 2007, 25, 2593–2600. [Google Scholar] [CrossRef] [PubMed]
- Madan, R.; Demircik, F.; Surianarayanan, S.; Allen, J.L.; Divanovic, S.; Trompette, A.; Yogev, N.; Gu, Y.; Khodoun, M.; Hildeman, D.; et al. Nonredundant roles for B cell-derived IL-10 in immune counter-regulation. J. Immunol. 2009, 183, 2312–2320. [Google Scholar] [CrossRef] [Green Version]
- Kühn, R.; Löhler, J.; Rennick, D.M.; Rajewsky, K.; Muller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 1993, 75, 263–274. [Google Scholar] [CrossRef]
- Mizuno, T.; Sawada, M.; Marunouchi, T.; Suzumura, A. Production of interleukin-10 by mouse glial cells in culture. Biochem. Biophys. Res. Commun. 1994, 205, 1907–1915. [Google Scholar] [CrossRef]
- Ledeboer, A.; Brevé, J.J.P.; Wierinckx, A.; Van Der Jagt, S.; Bristow, A.F.; Leysen, J.E.; Tilders, F.J.H.; Van Dam, A.-M. Expression and regulation of interleukin-10 and interleukin-10 receptor in rat astroglial and microglial cells. Eur. J. Neurosci. 2002, 16, 1175–1185. [Google Scholar] [CrossRef]
- Chabot, S.; Williams, G.; Hamilton, M.; Sutherland, G.; Yong, V.W. Mechanisms of IL-10 production in human microglia-T cell interaction. J. Immunol. 1999, 162, 6819–6828. [Google Scholar]
- Ma, S.-F.; Chen, Y.-J.; Zhang, J.-X.; Shen, L.; Wang, R.; Zhou, J.-S.; Hu, J.-G.; Lü, H.-Z. Adoptive transfer of M2 macrophages promotes locomotor recovery in adult rats after spinal cord injury. Brain Behav. Immun. 2015, 45, 157–170. [Google Scholar] [CrossRef]
- Duan, W.; Zhang, Y.-P.; Hou, Z.; Huang, C.; Zhu, H.; Zhang, C.-Q.; Yin, Q. Novel Insights into NeuN: From Neuronal Marker to Splicing Regulator. Mol. Neurobiol. 2016, 53, 1637–1647. [Google Scholar] [CrossRef]
- Darlot, F.; Vinit, S.; Matarazzo, V.; Kastner, A. Sustained cell body reactivity and loss of NeuN in a subset of axotomized bulbospinal neurons after a chronic high cervical spinal cord injury. Eur. J. Neurosci. 2017, 46, 2729–2745. [Google Scholar] [CrossRef] [PubMed]
- Ito, D.; Imai, Y.; Ohsawa, K.; Nakajima, K.; Fukuuchi, Y.; Kohsaka, S. Microglia-specific localisation of a novel calcium binding protein, Iba1. Mol. Brain Res. 1998, 57, 1–9. [Google Scholar] [CrossRef]
- Hoogland, I.C.; Houbolt, C.; Van Westerloo, D.J.; Van Gool, W.A.; Van de Beek, D. Systemic inflammation and microglial activation: Systematic review of animal experiments. J. Neuroinflamm. 2015, 12, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blinzinger, K.; Kreutzberg, G. Displacement of synaptic terminals from regenerating motoneurons by microglial cells. Z. Zellforsch. Mikrosk. Anat. 1968, 85, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Torvik, A.; Skjorten, F. Electron microscopic observations on nerve cell regeneration and degeneration after axon lesions. II. Changes in the glial cells. Acta Neuropathol. 1971, 17, 265–282. [Google Scholar] [CrossRef]
- Raivich, G.; Jones, L.L.; Kloss, C.U.A.; Werner, A.; Neumann, H.; Kreutzberg, G.W. Immune surveillance in the injured nervous system: T-lymphocytes invade the axotomized mouse facial motor nucleus and aggregate around sites of neuronal degeneration. J. Neurosci. 1998, 18, 5804–5816. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Choudhury, G.R.; Winters, A.; Yang, S.-H.; Jin, K. Cerebral regulatory T cells restrain microglia/macrophage-mediated inflammatory responses via IL-10. Eur. J. Immunol. 2015, 45, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Harrison, J.K.; Jiang, Y.; Chen, S.; Xia, Y.; Maciejewski, D.; McNamara, R.K.; Streit, W.J.; Salafranca, M.N.; Adhikari, S.; Thompson, D.A.; et al. Role for neuronally derived fractalkine in mediating interactions between neurons and CX3CR1-expressing microglia. Proc. Natl. Acad. Sci. USA 1998, 95, 10896–10901. [Google Scholar] [CrossRef] [Green Version]
- Caroni, P. Overexpression of growth-associated proteins in the neurons of adult transgenic mice. J. Neurosci. Methods 1997, 71, 3–9. [Google Scholar] [CrossRef]
- Villacampa, N.; Almolda, B.; Vilella, A.; Campbell, I.L.; González, B.; Castellano, B. Astrocyte-targeted production of IL-10 induces changes in microglial reactivity and reduces motor neuron death after facial nerve axotomy. Glia 2015, 63, 1166–1184. [Google Scholar] [CrossRef]
- Fouda, A.Y.; Kozak, A.; Alhusban, A.; Switzer, J.A.; Fagan, S.C. Anti-inflammatory IL-10 is upregulated in both hemispheres after experimental ischemic stroke: Hypertension blunts the response. Exp. Transl. Stroke Med. 2013, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chistyakov, D.V.; Azbukina, N.V.; Lopachev, A.V.; Kulichenkova, K.N.; Astakhova, A.A.; Sergeeva, M.G. Rosiglitazone as a Modulator of TLR4 and TLR3 Signaling Pathways in Rat Primary Neurons and Astrocytes. Int. J. Mol. Sci. 2018, 19, 113. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Lin, W.; Zhang, Y.; Lin, L.; Chen, J.; Zeng, Y.; Zheng, M.; Zhuang, Z.; Du, H.; Chen, R.; et al. IL-10 Promotes Neurite Outgrowth and Synapse Formation in Cultured Cortical Neurons after the Oxygen-Glucose Deprivation via JAK1/STAT3 Pathway. Sci. Rep. 2016, 6, 30459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, P.S.; Dhull, D.K.; Nalini, A.; Vijayalakshmi, K.; Sathyaprabha, T.N.; Alladi, P.A.; Raju, T.R. Astroglia acquires a toxic neuroinflammatory role in response to the cerebrospinal fluid from amyotrophic lateral sclerosis patients. J. Neuroinflamm. 2016, 13, 212. [Google Scholar] [CrossRef] [Green Version]
- Hulshof, S.; Montagne, L.; De Groot, C.J.; Van Der Valk, P. Cellular localization and expression patterns of interleukin-10, interleukin-4, and their receptors in multiple sclerosis lesions. Glia 2002, 38, 24–35. [Google Scholar] [CrossRef]
- Cannella, B.; Raine, C.S. The adhesion molecule and cytokine profile of multiple sclerosis lesions. Ann. Neurol. 1995, 37, 424–435. [Google Scholar] [CrossRef]
- Jander, S.; Pohl, J.; D’Urso, D.; Gillen, C.; Stoll, G. Time course and cellular localization of interleukin-10 mRNA and protein expression in autoimmune inflammation of the rat central nervous system. Am. J. Pathol. 1998, 152, 975–982. [Google Scholar]
- Voskuhl, R.R.; Peterson, R.S.; Song, B.; Ao, Y.; Morales, L.B.J.; Tiwari-Woodruff, S.; Sofroniew, M.V. Reactive astrocytes form scar-like perivascular barriers to leukocytes during adaptive immune inflammation of the CNS. J. Neurosci. 2009, 29, 11511–11522. [Google Scholar] [CrossRef] [Green Version]
- Tone, M.; Powell, M.J.; Tone, Y.; Thompson, S.A.J.; Waldmann, H. IL-10 gene expression is controlled by the transcription factors Sp1 and Sp3. J. Immunol. 2000, 165, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Powell, M.J.; Thompson, S.A.J.; Tone, Y.; Waldmann, H.; Tone, M. Posttranscriptional regulation of IL-10 gene expression through sequences in the 3’-untranslated region. J. Immunol. 2000, 165, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.; Dooley, N.; Ulvestad, E.; Becher, B.; Antel, J. IL-10 production by adult human derived microglial cells. Neurochem. Int. 1996, 29, 55–64. [Google Scholar] [CrossRef]
- Jack, C.S.; Arbour, N.; Manusow, J.; Montgrain, V.; Blain, M.; McCrea, E.; Shapiro, A.; Antel, J. TLR signaling tailors innate immune responses in human microglia and astrocytes. J. Immunol. 2005, 175, 4320–4330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, D.R.; Kim, S.Y.; Kim, K.Y.; Lee, H.G.; Moon, J.H.; Lee, J.S.; Lee, S.H.; Kim, S.U.; Lee, Y.B. Cross talk between P2 purinergic receptors modulates extracellular ATP-mediated interleukin-10 production in rat microglial cells. Exp. Mol. Med. 2008, 40, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Park, K.W.; Lee, H.G.; Jin, B.K.; Lee, Y.B. Interleukin-10 endogenously expressed in microglia prevents lipopolysaccharide-induced neurodegeneration in the rat cerebral cortex in vivo. Exp. Mol. Med. 2007, 39, 812–819. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Peng, X.; Insolera, R.; Fink, D.J.; Mata, M. IL-10 promotes neuronal survival following spinal cord injury. Exp. Neurol. 2009, 220, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Peng, X.; Insolera, R.; Fink, D.J.; Mata, M. Interleukin-10 provides direct trophic support to neurons. J. Neurochem. 2009, 110, 1617–1627. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain (Jackson Strain No.) | Alternately Named | Source |
|---|---|---|
| B6(Cg)-Il10tm1.1Karp/J (014530) | IL-10/GFP reporter | The Jackson Laboratory |
| B6.129P2(Cg)-Cx3cr1tm2.1(cre/ERT2)Litt/WganJ (021160) | Cx3cr1Cre | The Jackson Laboratory |
| B6.129P2-Il10tm1Cgn/J (002251) | IL-10−/− | The Jackson Laboratory |
| B6.Cg-Tg(GFAP-cre/ERT2)505Fmv/J (012849) | GfapCre | The Jackson Laboratory |
| C57BL/6J (000664) | WT | The Jackson Laboratory |
| IL10flox/flox | Dr. Gang Huang, Cincinnati Children’s Hospital; see [10] | |
| Tg(Thy1-cre/ERT2,-EYFP)HGfng/PyngJ (012708) | Thy1Cre | The Jackson Laboratory |
| Gene | 5′ Primer Sequence | 3′ Primer Sequence |
|---|---|---|
| IL10 exon 1 (133 bp in WT) | ATG CCT GGC TCA G | CCA CAT GCT CCT AGA GCT GC |
| IL10 floxed sequence and exon 2 (480 bp WT, 514 bp floxed) | CCA GCA TAG AGA GCT TGC ATT ACA | GAG TCG GTT AGC AGT ATG TTG TCC AG |
| Thy1Cre transgene (300 bp) | TCT GAG TGG CAA AGG ACC TTA GG | CGC TGA ACT TGT GGC CGT TTA CG |
| Thy1Cre internal control (200 bp) | CAA ATG TTG CTT GTC TGG TG | GTC AGT CGA GTG CAC AGT TT |
| GfapCre transgene (200 bp) | GCC AGT CTA GCC CAC TCC TT | TCC CTG AAC ATG TCC ATC AG |
| GfapCre internal control (324 bp) | CTA GGC CAC AGA ATT GAA AGA TCT | GTA GGT GGA AAT TCT AGC ATC ATC C |
| Cx3cr1Cre transgene (300 bp) | AAG ACT CAC GTG GAC CTG CT | CGG TTA TTC AAC TTG CAC CA |
| Cx3cr1Cre internal control (695 bp) | AAG ACT CAC GTG GAC CTG CT (common) | AGG ATG TTG ACT TCC GAG TTG |
| Antibody | Manufacturer and Cat. No. | Dilution |
|---|---|---|
| Mouse anti-GFAP 594 (used in IL-10/GFP reporter) | Thermo Fisher A-21295 | 1:500 |
| Rabbit anti-GFP 488 | Thermo Fisher A-21311 | 1:100 |
| Rabbit anti-IBA1 | Thermo Fisher 019-19741 | 1:500 |
| Mouse anti-NeuN 555 | Millipore MAB377A5 | 1:200 |
| Rabbit anti-GFAP (used in perfused WT tissue) | Thermo Fisher PA3-16727 | 1:1000 |
| Goat anti-IL-10 | R&D Systems AF519 | 1:200 |
| Donkey anti-rabbit 568 (used for GFAP, IBA1) | Abcam ab175470 | 1:1000 |
| Donkey anti-goat 488 (used for IL-10) | Abcam ab150129 | 1:1000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Runge, E.M.; Setter, D.O.; Iyer, A.K.; Regele, E.J.; Kennedy, F.M.; Sanders, V.M.; Jones, K.J. Cellular Sources and Neuroprotective Roles of Interleukin-10 in the Facial Motor Nucleus after Axotomy. Cells 2022, 11, 3167. https://doi.org/10.3390/cells11193167
Runge EM, Setter DO, Iyer AK, Regele EJ, Kennedy FM, Sanders VM, Jones KJ. Cellular Sources and Neuroprotective Roles of Interleukin-10 in the Facial Motor Nucleus after Axotomy. Cells. 2022; 11(19):3167. https://doi.org/10.3390/cells11193167
Chicago/Turabian StyleRunge, Elizabeth M., Deborah O. Setter, Abhirami K. Iyer, Eric J. Regele, Felicia M. Kennedy, Virginia M. Sanders, and Kathryn J. Jones. 2022. "Cellular Sources and Neuroprotective Roles of Interleukin-10 in the Facial Motor Nucleus after Axotomy" Cells 11, no. 19: 3167. https://doi.org/10.3390/cells11193167