
Impact of Neurons on Patient-Derived Cardiomyocytes Using Organ-On-A-Chip and iPSC Biotechnologies
 , , , , and
, , , , and 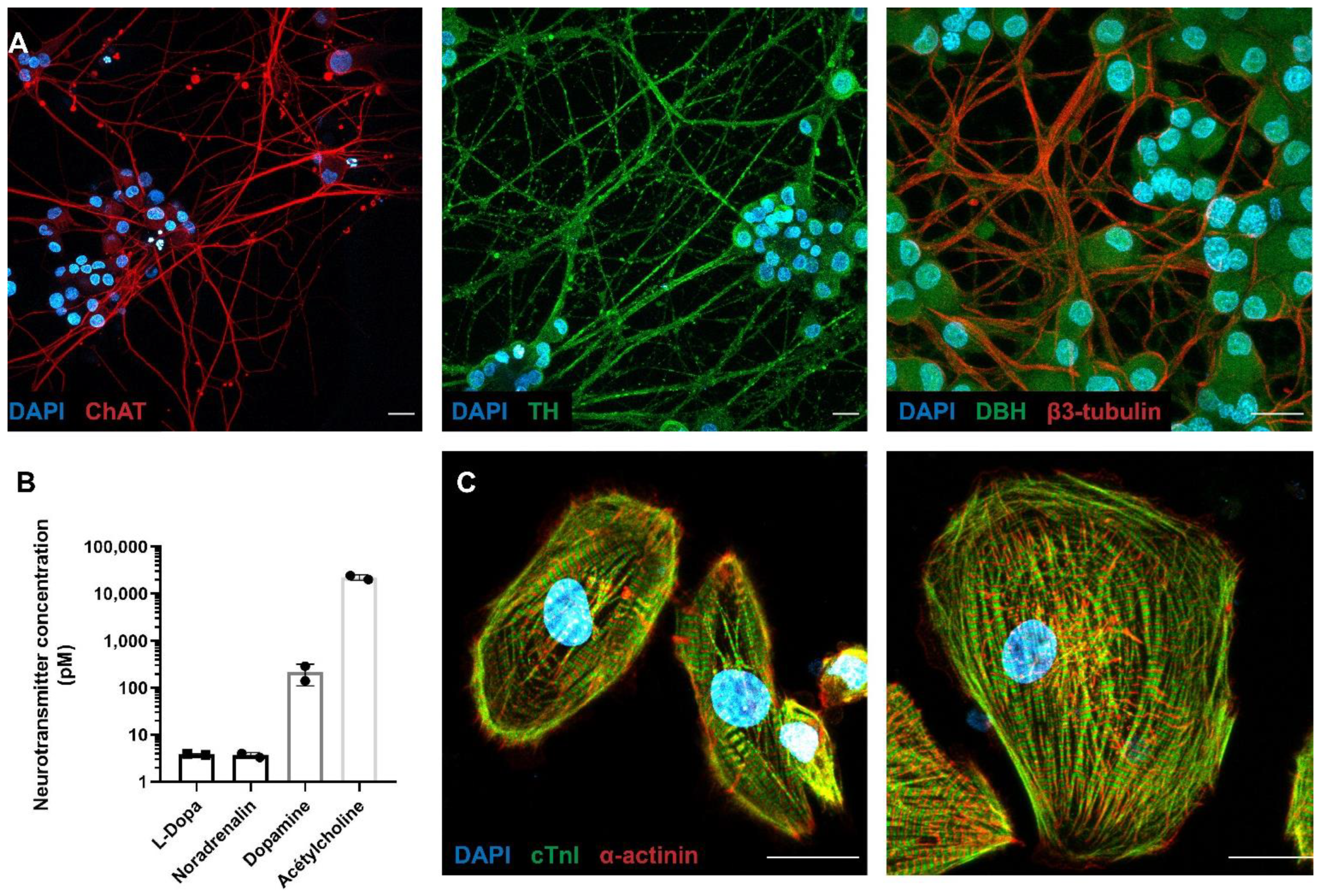{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
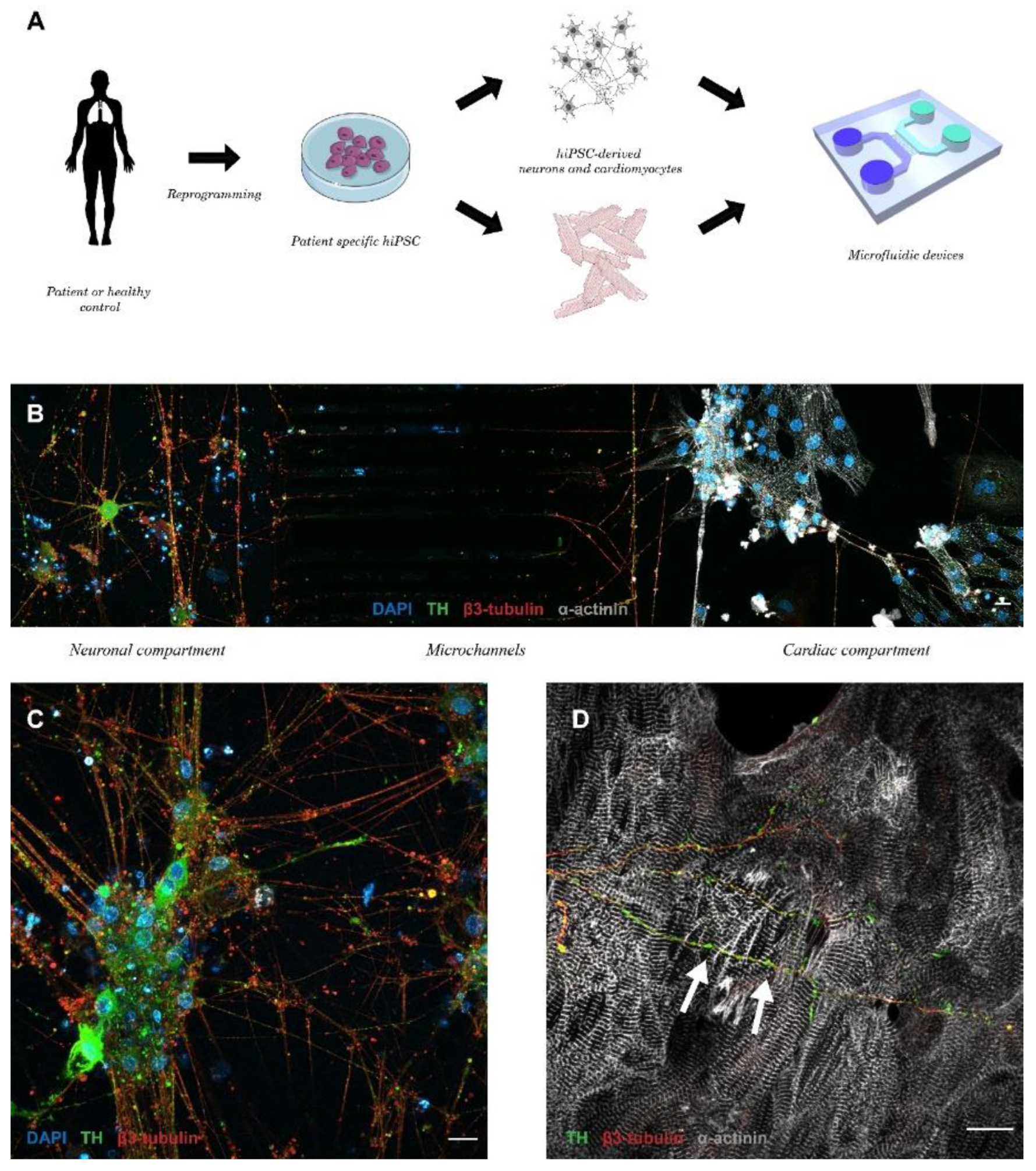{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microfluidic Devices
2.2. Rat PC12 Culture
2.3. hiPSCs Maintenance and Differentiation
2.4. Neurotransmitter Release Quantification
2.5. PC12/hiPSC-CMs Co-Culture
2.6. Immunocytochemistry
2.7. Intracellular Calcium Dynamics
2.8. Measurement of Contractile Function Using Video-Edge Capture
2.9. Statistical Analysis
3. Results
3.1. Molecular Characterization of the PC12 Neurons Connected with hiPSC-CMs
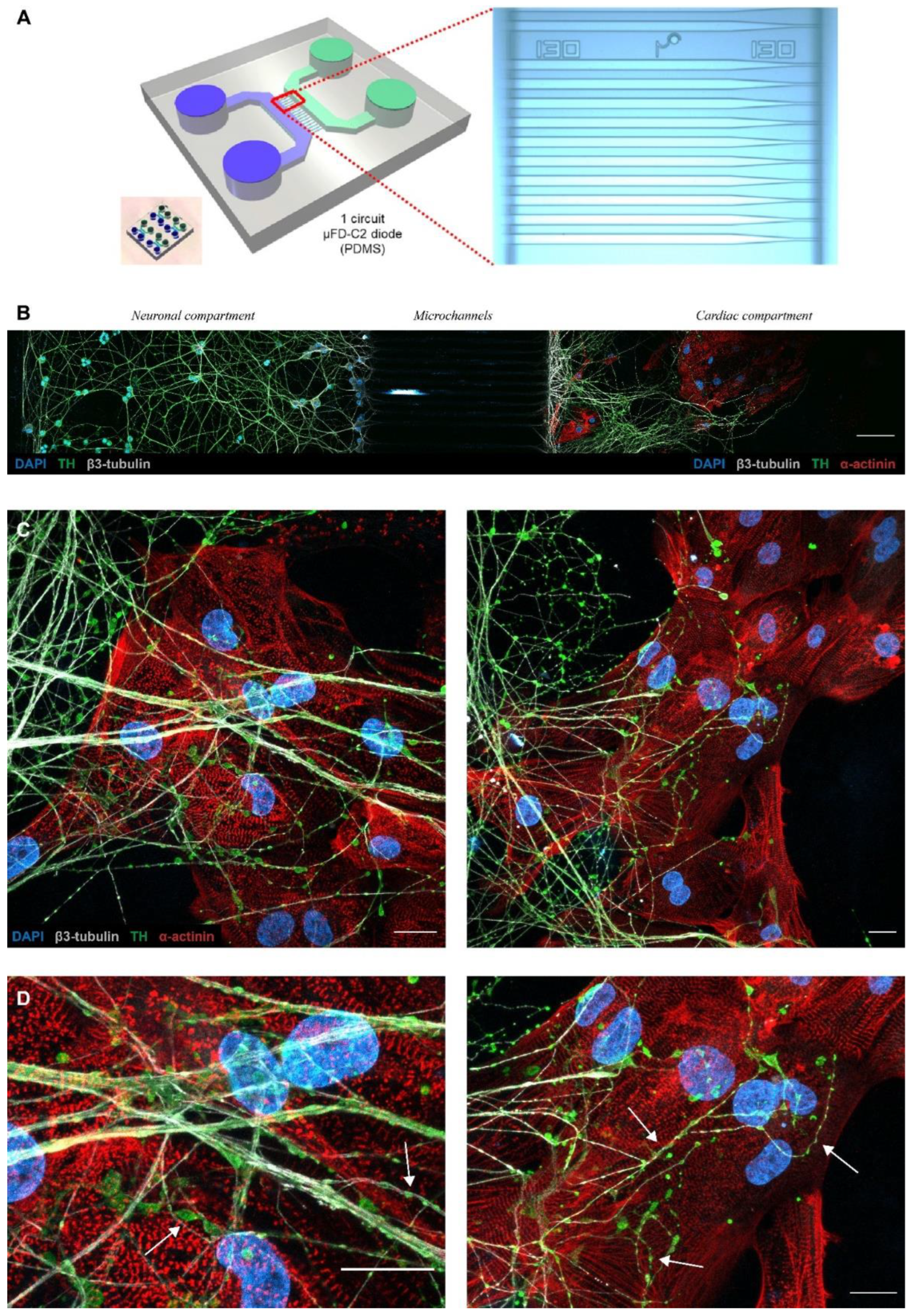
3.2. Microfluidic Devices Allow a Neuro–Cardiac OOC
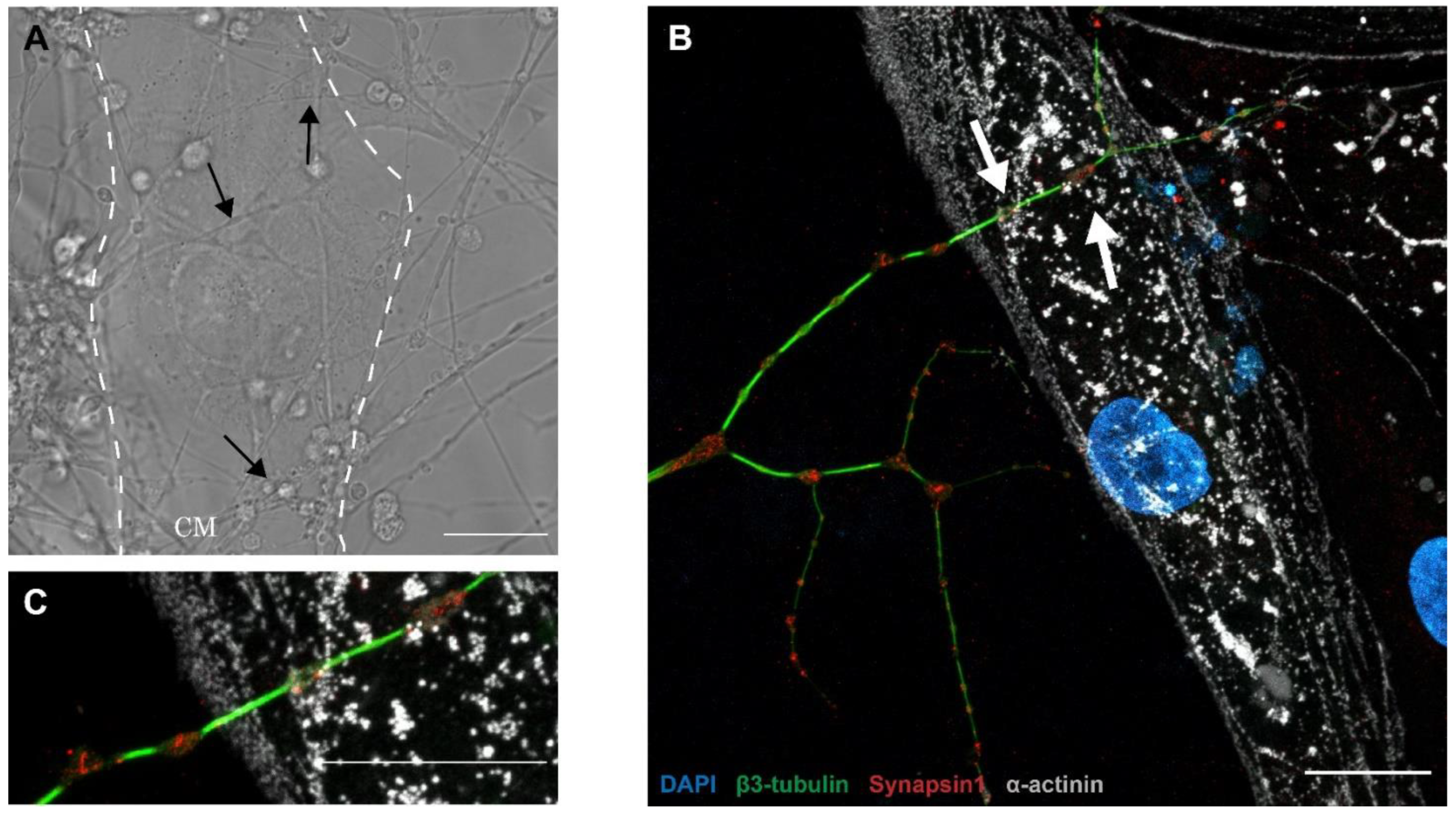
3.3. PC12/hiPSC-CMs OOC Involves Neuro–Cardiac Junctions and Synapse-Like Junctions
3.4. Neurons Modulate the Intracellular Calcium Handling in hiPSC-CMs
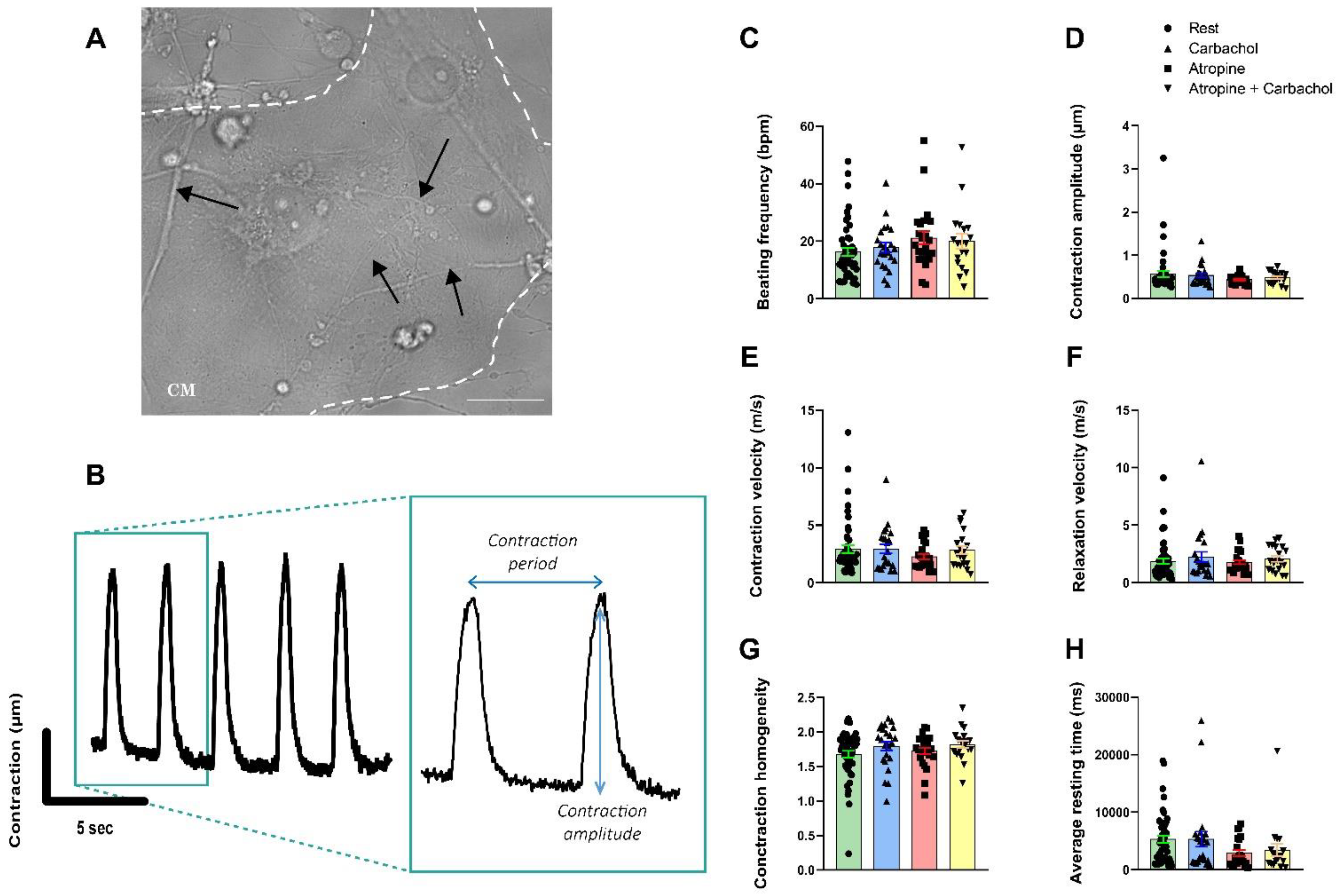
3.5. Neurons Do Not Impact the Contractile Properties in hiPSC-CMs
3.6. Patient-Specific Neuro–Cardiac OOC Using hiPSC-NRs and hiPSC-CMs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crick, S.J.; Anderson, R.H.; Ho, S.Y.; Sheppard, M.N. Localisation and quantitation of autonomic innervation in the porcine heart II: Endocardium, myocardium and epicardium. J. Anat. 1999, 195, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.S.; Challis, R.C.; Fowlkes, C.C.; Hanna, P.; Tompkins, J.D.; Jordan, M.C.; Hiyari, S.; Gabris-Weber, B.A.; Greenbaum, A.; Chan, K.Y.; et al. Identification of peripheral neural circuits that regulate heart rate using optogenetic and viral vector strategies. Nat. Commun. 2019, 10, 1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ieda, M.; Kanazawa, H.; Kimura, K.; Hattori, F.; Ieda, Y.; Taniguchi, M.; Lee, J.; Matsumura, K.; Tomita, Y.; Miyoshi, S.; et al. Sema3a maintains normal heart rhythm through sympathetic innervation patterning. Nat. Med. 2007, 13, 604–612. [Google Scholar] [CrossRef]
- Kimura, K.; Ieda, M.; Fukuda, K. Development, maturation, and transdifferentiation of cardiac sympathetic nerves. Circ. Res. 2012, 110, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Dowell, R.T. Postnatal development of rat heart during 6-hydroxydopamine or propranolol treatment. Proc. Soc. Exp. Biol. Med. 1985, 178, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Kreipke, R.E.; Birren, S.J. Innervating sympathetic neurons regulate heart size and the timing of cardiomyocyte cell cycle withdrawal. J. Physiol. 2015, 593, 5057–5073. [Google Scholar] [CrossRef] [Green Version]
- Shcherbakova, O.G.; Hurt, C.M.; Xiang, Y.; Dell’Acqua, M.L.; Zhang, Q.; Tsien, R.W.; Kobilka, B.K. Organization of β-adrenoceptor signaling compartments by sympathetic innervation of cardiac myocytes. J. Cell Biol. 2007, 176, 521–533. [Google Scholar] [CrossRef]
- Zaglia, T.; Milan, G.; Franzoso, M.; Bertaggia, E.; Pianca, N.; Piasentini, E.; Voltarelli, V.A.; Chiavegato, D.; Brum, P.C.; Glass, D.J.; et al. Cardiac sympathetic neurons provide trophic signal to the heart via beta2-adrenoceptor-dependent regulation of proteolysis. Cardiovasc. Res. 2013, 97, 240–250. [Google Scholar] [CrossRef] [Green Version]
- Taskiran Mustafa, F.-H.T.; Verner, R.; Henrik, L.B.W.; Jannik, H. Decreased Myocardial Perfusion Reserve in Diabetic Autonomic Neuropathy. Diabetes 2002, 51, 3306–3310. [Google Scholar] [CrossRef]
- Mann, D.L.; Bristow, M.R. Mechanisms and models in heart failure: The biomechanical model and beyond. Circulation 2005, 111, 2837–2849. [Google Scholar] [CrossRef]
- Hong, J.; Adam, R.J.; Gao, L.; Hahka, T.; Xia, Z.; Wang, D.; Nicholas, T.A.; Zucker, I.H.; Lisco, S.J.; Wang, H. Macrophage activation in stellate ganglia contributes to lung injury-induced arrhythmogenesis in male rats. Acta Physiol. 2021, 232, e13657. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.J.; Kimber, S.; Johnstone, D.E.; Shukla, R.C.; Horacek, B.M.; Forbes, C.; Armour, J.A. The Effects of Unilateral Stellate Ganglion Blockade on Human Cardiac Function During Rest and Exercise. J. Cardiovasc. Electrophysiol. 1993, 4, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Cho, G.-S.; Li, Z.; Hong, I.; Zhu, R.; Kim, M.-J.; Kim, Y.J.; Tampakakis, E.; Tung, L.; Huganir, R.; et al. Functional Coupling with Cardiac Muscle Promotes Maturation of hPSC-Derived Sympathetic Neurons. Cell Stem Cell 2016, 19, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prando, V.; Da Broi, F.; Franzoso, M.; Plazzo, A.P.; Pianca, N.; Francolini, M.; Basso, C.; Kay, M.W.; Zaglia, T.; Mangillo, M. Dynamics of neuroeffector coupling at cardiac sympathetic synapses: Dynamics of neurocardiac communication. J. Physiol. 2018, 596, 2055–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duc, P.; Vignes, M.; Hugon, G.; Sebban, A.; Carnac, G.; Malyshev, E.; Charlot, B.; Rage, F. Human neuromuscular junction on micro-structured microfluidic devices implemented with a custom micro electrode array (MEA). Lab Chip 2021, 21, 4223–4236. [Google Scholar] [CrossRef]
- Chiew, G.G.Y.; Wei, N.; Sultania, S.; Lim, S.; Luo, K.Q. Bioengineered three-dimensional co-culture of cancer cells and endothelial cells: A model system for dual analysis of tumor growth and angiogenesis: Analysis of Tumor and Vessel Growth in a 3D Co-Culture. Biotechnol. Bioeng. 2017, 114, 1865–1877. [Google Scholar] [CrossRef]
- Arrigoni, C.; Bersini, S.; Gilardi, M.; Moretti, M. In Vitro Co-Culture Models of Breast Cancer Metastatic Progression towards Bone. Int. J. Mol. Sci. 2016, 17, 1405. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, A.; Nakafutami, S.; Tani, H.; Mori, M.; Takayama, Y.; Moriguchi, H.; Kotani, K.; Miwa, K.; Lee, J.; Noshiro, M.; et al. Device for co-culture of sympathetic neurons and cardiomyocytes using microfabrication. Lab Chip 2011, 11, 2268–2275. [Google Scholar] [CrossRef]
- Takeuchi, A.; Shimba, K.; Mori, M.; Takayama, Y.; Moriguchi, H.; Kotani, K.; Lee, J.; Noshiro, M.; Jimbo, Y. Sympathetic neurons modulate the beat rate of pluripotent cell-derived cardiomyocytes in vitro. Integr. Biol. 2012, 4, 1532–1539. [Google Scholar] [CrossRef]
- Acimovic, I.; Vilotic, A.; Pesl, M.; Lacampagne, A.; Dvorak, P.; Rotrekl, V.; Meli, A.C. Human pluripotent stem cell-derived cardiomyocytes as research and therapeutic tools. BioMed Res. Int. 2014, 2014, 512831. [Google Scholar] [CrossRef]
- Moreau, A.; Reisqs, J.B.; Delanoe-Ayari, H.; Pierre, M.; Janin, A.; Deliniere, A.; Bessiere, F.; Meli, A.C.; Charrabi, A.; Lafont, E.; et al. Deciphering DSC2 arrhythmogenic cardiomyopathy electrical instability: From ion channels to ECG and tailored drug therapy. Clin. Transl. Med. 2021, 11, e319. [Google Scholar] [CrossRef] [PubMed]
- Sleiman, Y.; Souidi, M.; Kumar, R.; Yang, E.; Jaffre, F.; Zhou, T.; Bernardin, A.; Reiken, S.; Cazorla, O.; Kajava, A.V.; et al. Modeling polymorphic ventricular tachycardia at rest using patient-specific induced pluripotent stem cell-derived cardiomyocytes. EBioMedicine 2020, 60, 103024. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Klos, M.; Wilson, G.F.; Herman, A.M.; Lian, X.; Raval, K.K.; Barron, M.R.; Hou, L.; Soerens, A.G.; Yu, J.; et al. Extracellular Matrix Promotes Highly Efficient Cardiac Differentiation of Human Pluripotent Stem CellsNovelty and Significance: The Matrix Sandwich Method. Circ. Res. 2012, 111, 1125–1136. [Google Scholar] [CrossRef]
- Tohyama, S.; Hattori, F.; Sano, M.; Hishiki, T.; Nagahata, Y.; Matsuura, T.; Hashimoto, H.; Suzuki, T.; Yamashita, H.; Satoh, Y.; et al. Distinct metabolic flow enables large-scale purification of mouse and human pluripotent stem cell-derived cardiomyocytes. Cell Stem Cell 2013, 12, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Winbo, A.; Ramanan, S.; Eugster, E.; Jovinge, S.; Skinner, J.R.; Montgomery, J.M. Functional Coculture of Sympathetic Neurons and Cardiomyocytes Derived from Human Induced Pluripotent Stem Cells. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H927–H937. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H.; Foster, G.; Cone, E.J.; Kumar, S.D. Selecting an appropriate isotopic internal standard for gas chromatography/mass spectrometry analysis of drugs of abuse--pentobarbital example. J. Forensic Sci. 1995, 40, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Fauconnier, J.; Meli, A.C.; Thireau, J.; Roberge, S.; Shan, J.; Sassi, Y.; Reiken, S.R.; Rauzier, J.M.; Marchand, A.; Chauvier, D.; et al. Ryanodine receptor leak mediated by caspase-8 activation leads to left ventricular injury after myocardial ischemia-reperfusion. Proc. Natl. Acad. Sci. USA 2011, 108, 13258–13263. [Google Scholar] [CrossRef] [Green Version]
- Lacampagne, A.; Liu, X.; Reiken, S.; Bussiere, R.; Meli, A.C.; Lauritzen, I.; Teich, A.F.; Zalk, R.; Saint, N.; Arancio, O.; et al. Post-translational remodeling of ryanodine receptor induces calcium leak leading to Alzheimer’s disease-like pathologies and cognitive deficits. Acta Neuropathol. 2017, 134, 749–767. [Google Scholar] [CrossRef]
- Jelinkova, S.; Vilotic, A.; Pribyl, J.; Aimond, F.; Salykin, A.; Acimovic, I.; Pesl, M.; Caluori, G.; Klimovic, S.; Urban, T.; et al. DMD Pluripotent Stem Cell Derived Cardiac Cells Recapitulate in vitro Human Cardiac Pathophysiology. Front. Bioeng. Biotechnol. 2020, 8, 535. [Google Scholar] [CrossRef]
- Souidi, M.; Sleiman, Y.; Acimovic, I.; Pribyl, J.; Charrabi, A.; Baecker, V.; Scheuermann, V.; Pesl, M.; Jelinkova, S.; Skladal, P.; et al. Oxygen Is an Ambivalent Factor for the Differentiation of Human Pluripotent Stem Cells in Cardiac 2D Monolayer and 3D Cardiac Spheroids. Int. J. Mol. Sci. 2021, 22, 662. [Google Scholar] [CrossRef]
- Taylor, A.M.; Blurton-Jones, M.; Rhee, S.W.; Cribbs, D.H.; Cotman, C.W.; Jeon, N.L. A microfluidic culture platform for CNS axonal injury, regeneration and transport. Nat. Methods 2005, 2, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.; Rhee, S.W.; Tu, C.H.; Cribbs, D.H.; Cotman, C.W.; Jeon, N.L. Microfluidic Multicompartment Device for Neuroscience Research. Langmuir 2003, 19, 1551–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habibey, R.; Rojo Arias, J.E.; Striebel, J.; Busskamp, V.A.-O. Microfluidics for Neuronal Cell and Circuit Engineering. Chem. Rev. 2022, 122, 14842–14880. [Google Scholar] [CrossRef] [PubMed]
- Peyrin, J.; Deliglise, B.; Saias, L.; Vignes, M.; Gougis, P.; Magnifico, S.; Betuing, S.; Pietri, M.; Caboche, J.; Vanhoutte, P.; et al. Axon diodes for the reconstruction of oriented neuronal networks in microfluidic chambers. Lab Chip 2011, 11, 3663–3673. [Google Scholar] [CrossRef] [PubMed]
- Kruepunga, N.; Hikspoors, J.; Hulsman, C.J.M.; Mommen, G.M.C.; Kohler, S.E.; Lamers, W.H. Development of the sympathetic trunks in human embryos. J. Anat. 2021, 239, 32–45. [Google Scholar] [CrossRef]
- Hilfiker, S.; Pieribone, V.A.; Czernik, A.J.; Kao, H.T.; Augustine, G.J.; Greengard, P. Synapsins as regulators of neurotransmitter release. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1999, 354, 269–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalski, W.J.; Garcia-Pak, I.H.; Li, W.; Uosaki, H.; Tampakakis, E.; Zou, J.; Lin, Y.; Patterson, K.; Kwon, C.; Mukouyama, Y. Sympathetic Neurons Regulate Cardiomyocyte Maturation in Culture. Front. Cell Dev. Biol. 2022, 10, 850645. [Google Scholar] [CrossRef]
- Hakli, M.; Jantti, S.; Joki, T.; Sukki, L.; Tornberg, K.; Aalto-Setala, K.; Kallio, P.; Pekkanen-Mattila, M.; Narkilahti, S. Human Neurons Form Axon-Mediated Functional Connections with Human Cardiomyocytes in Compartmentalized Microfluidic Chip. Int. J. Mol. Sci. 2022, 23, 3148. [Google Scholar] [CrossRef]
- Freeman, K.; Tao, W.; Sun, H.; Soonpaa, M.H.; Rubart, M. In situ three-dimensional reconstruction of mouse heart sympathetic innervation by two-photon excitation fluorescence imaging. J. Neurosci. Methods 2014, 221, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Huh, K.H.; Fuhrer, C. Clustering of nicotinic acetylcholine receptors: From the neuromuscular junction to interneuronal synapses. Mol. Neurobiol. 2002, 25, 79–112. [Google Scholar] [CrossRef]
- Fornasiero, E.F.; Raimondi, A.; Guarnieri, F.C.; Orlando, M.; Fesce, R.; Benfenati, F.; Valtorta, F. Synapsins contribute to the dynamic spatial organization of synaptic vesicles in an activity-dependent manner. J. Neurosci. 2012, 32, 12214–12227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosahl, T.W.; Spillane, D.; Missler, M.; Herz, J.; Selig, D.K.; Wolff, J.R.; Hammer, R.E.; Malenka, R.C.; Südhof, T.C. Essential functions of synapsins I and II in synaptic vesicle regulation. Nature 1995, 375, 488–493. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardin, A.A.; Colombani, S.; Rousselot, A.; Andry, V.; Goumon, Y.; Delanoë-Ayari, H.; Pasqualin, C.; Brugg, B.; Jacotot, E.D.; Pasquié, J.-L.; et al. Impact of Neurons on Patient-Derived Cardiomyocytes Using Organ-On-A-Chip and iPSC Biotechnologies. Cells 2022, 11, 3764. https://doi.org/10.3390/cells11233764
Bernardin AA, Colombani S, Rousselot A, Andry V, Goumon Y, Delanoë-Ayari H, Pasqualin C, Brugg B, Jacotot ED, Pasquié J-L, et al. Impact of Neurons on Patient-Derived Cardiomyocytes Using Organ-On-A-Chip and iPSC Biotechnologies. Cells. 2022; 11(23):3764. https://doi.org/10.3390/cells11233764
Chicago/Turabian StyleBernardin, Albin A., Sarah Colombani, Antoine Rousselot, Virginie Andry, Yannick Goumon, Hélène Delanoë-Ayari, Côme Pasqualin, Bernard Brugg, Etienne D. Jacotot, Jean-Luc Pasquié, and et al. 2022. "Impact of Neurons on Patient-Derived Cardiomyocytes Using Organ-On-A-Chip and iPSC Biotechnologies" Cells 11, no. 23: 3764. https://doi.org/10.3390/cells11233764