
Response Facilitation Induced by Insulin-like Growth Factor-I in the Primary Somatosensory Cortex of Mice Was Reduced in Aging

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Recordings and Tactile Stimulation
2.2. Drugs
2.3. Immunohistochemistry
2.4. Behavioral Experiments
2.5. Data Analysis and Statistics
3. Results
3.1. Whisker Responses Increased in the Presence of IGF-I in Young and Old Animals
3.2. Response Facilitation in Young and Old Mice. Effect of IGF-I Cortical Application
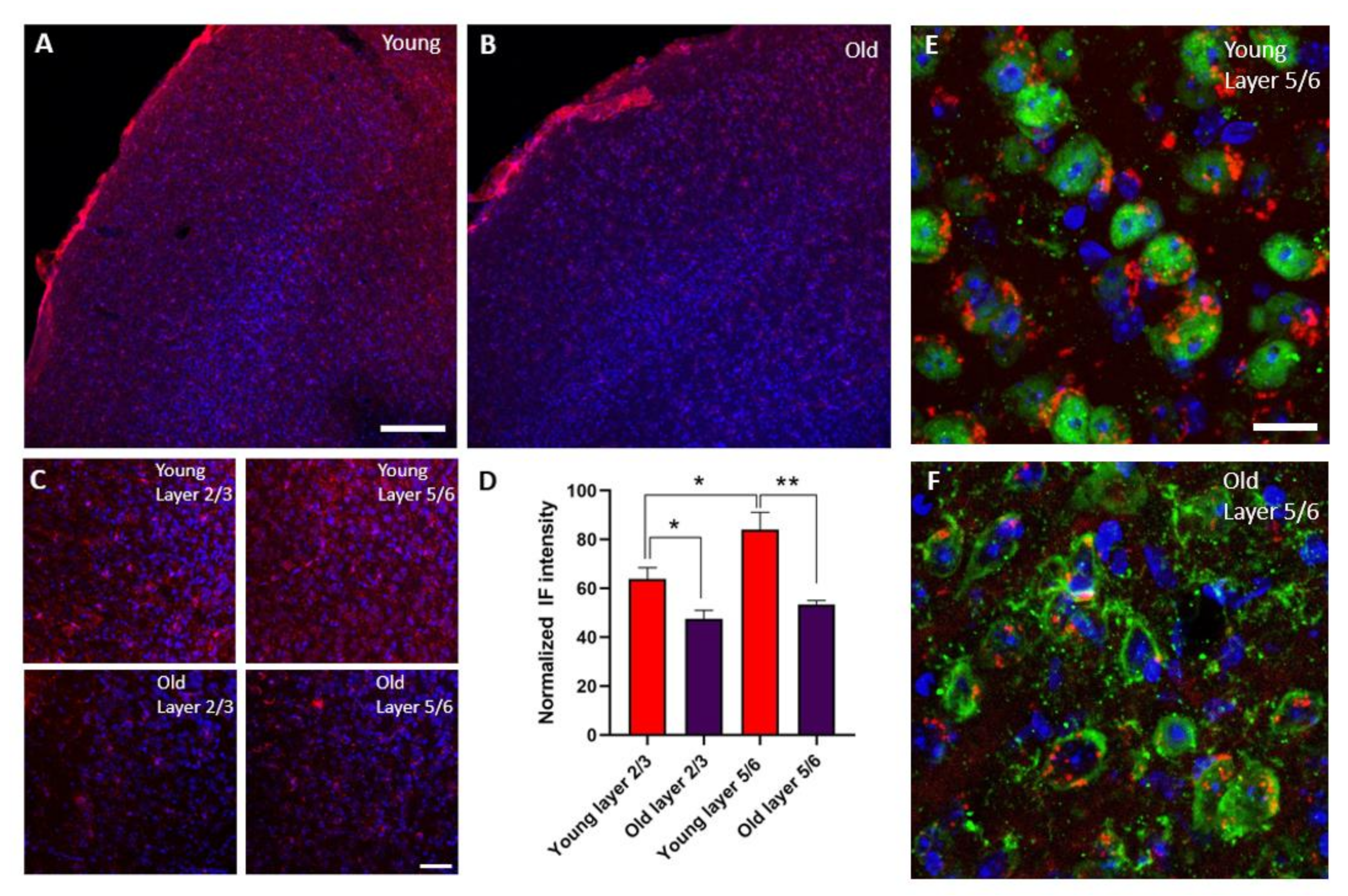
3.3. The Expression of IGR-IR Was Reduced in the S1 Cortex of Old Mice
3.4. The Performance of a Whisker Discrimination Task Is Impaired in Old Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ishii, D.N.; Lupien, S.B. Insulin-like growth factor replacement therapy for diabetic neuropathy: Experimental basis. Exp. Diabesity Res. 2003, 4, 257–269. [Google Scholar] [CrossRef]
- Fernandez, A.M.; Santi, A.; Torres-Aleman, I. Insulin Peptides as Mediators of the Impact of Life Style in Alzheimer’s disease. Brain Plast. 2018, 4, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, A.M.; Torres-Aleman, I. The many faces of insulin-like peptide signalling in the brain. Nat. Rev. Neurosci. 2012, 13, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Al-Samerria, S.; Radovick, S. The Role of Insulin-like Growth Factor-1 (IGF-1) in the Control of Neuroendocrine Regulation of Growth. Cells 2021, 10, 2664. [Google Scholar] [CrossRef] [PubMed]
- Dyer, A.H.; Vahdatpour, C.; Sanfeliu, A.; Tropea, D. The role of Insulin-Like Growth Factor 1 (IGF-1) in brain development, maturation and neuroplasticity. Neuroscience 2016, 325, 89–99. [Google Scholar] [CrossRef]
- Barros-Zulaica, N.; Villa, A.E.P.; Nunez, A. Response Adaptation in Barrel Cortical Neurons Facilitates Stimulus Detection during Rhythmic Whisker Stimulation in Anesthetized Mice. eNeuro 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Carro, E.; Nuñez, A.; Busiguina, S.; Torres-Aleman, I. Circulating insulin-like growth factor I mediates effects of exercise on the brain. J. Neurosci. 2000, 20, 2926–2933. [Google Scholar] [CrossRef]
- Gazit, N.; Vertkin, I.; Shapira, I.; Helm, M.; Slomowitz, E.; Sheiba, M.; Mor, Y.; Rizzoli, S.; Slutsky, I. IGF-1 Receptor Differentially Regulates Spontaneous and Evoked Transmission via Mitochondria at Hippocampal Synapses. Neuron 2016, 89, 583–597. [Google Scholar] [CrossRef] [Green Version]
- Nuñez, A.; Carro, E.; Torres-Aleman, I. Insulin-like growth factor I modifies electrophysiological properties of rat brain stem neurons. J. Neurophysiol. 2003, 89, 3008–3017. [Google Scholar] [CrossRef] [Green Version]
- Zegarra-Valdivia, J.A.; Pignatelli, J.; Fernandez de Sevilla, M.E.; Fernandez, A.M.; Munive, V.; Martinez-Rachadell, L.; Nuñez, A.; Torres Aleman, I. Insulin-like growth factor I modulates sleep through hypothalamic orexin neurons. FASEB J. 2020, 34, 15975–15990. [Google Scholar] [CrossRef] [PubMed]
- Zegarra-Valdivia, J.A.; Santi, A.; Fernandez de Sevilla, M.E.; Nuñez, A.; Torres-Aleman, I. Serum Insulin-Like Growth Factor I Deficiency Associates to Alzheimer’s Disease Co-Morbidities. J. Alzheimers Dis. 2019, 69, 979–987. [Google Scholar] [CrossRef]
- Zegarra-Valdivia, J.A.; Chaves-Coira, I.; Fernandez de Sevilla, M.E.; Martinez-Rachadell, L.; Esparza, J.; Torres-Aleman, I.; Nuñez, A. Reduced Insulin-Like Growth Factor-I Effects in the Basal Forebrain of Aging Mouse. Front. Aging Neurosci. 2021, 13, 682388. [Google Scholar] [CrossRef]
- Trueba-Sáiz, A.; Cavada, C.; Fernandez, A.M.; Leon, T.; González, D.A.; Fortea Ormaechea, J.; Lleó, A.; Del Ser, T.; Nuñez, A.; Torres-Aleman, I. Loss of serum IGF-I input to the brain as an early biomarker of disease onset in Alzheimer mice. Transl. Psychiatry 2013, 3, e330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maher, F.O.; Clarke, R.M.; Kelly, A.; Nally, R.E.; Lynch, M.A. Interaction between interferon gamma and insulin-like growth factor-1 in hippocampus impacts on the ability of rats to sustain long-term potentiation. J. Neurochem. 2006, 96, 1560–1571. [Google Scholar] [CrossRef]
- Trejo, J.L.; Piriz, J.; Llorens-Martin, M.V.; Fernandez, A.M.; Bolós, M.; LeRoith, D.; Nuñez, A.; Torres-Aleman, I. Central actions of liver-derived insulin-like growth factor I underlying its pro-cognitive effects. Mol. Psychiatry 2007, 12, 1118–1128. [Google Scholar] [CrossRef]
- Castro-Alamancos, M.A.; Torres-Alemán, I. Learning of the conditioned eye-blink response is impaired by an antisense insuline-like growth factor I oligonucleotide. Proc. Natl. Acad. Sci. USA 1994, 91, 10203–10207. [Google Scholar] [CrossRef] [Green Version]
- Carvell, G.E.; Simons, D.J. Biometric analyses of vibrissal tactile discrimination in the rat. J. Neurosci. 1990, 10, 2638–2648. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, S.P.; Wolfe, J.; Feldman, D.E. Sparse temporal coding of elementary tactile features during active whisker sensation. Nat. Neurosci. 2009, 12, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Berg, R.W.; Kleinfeld, D. Vibrissa movement elicited by rhythmic electrical microstimulation to motor cortex in the aroused rat mimics exploratory whisking. J. Neurophysiol. 2003, 90, 2950–2963. [Google Scholar] [CrossRef]
- Von Heimendahl, M.; Itskov, P.M.; Arabzadeh, E.; Diamond, M.E. Neuronal activity in rat barrel cortex underlying texture discrimination. PLoS Biol. 2007, 5, e305. [Google Scholar] [CrossRef] [Green Version]
- Garabedian, C.E.; Jones, S.R.; Merzenich, M.M.; Dale, A.; Moore, C.I. Band-pass response properties of rat SI neurons. J. Neurophysiol. 2003, 90, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Megevand, P.; Troncoso, E.; Quairiaux, C.; Muller, D.; Michel, C.M.; Kiss, J.Z. Long-term plasticity in mouse sensorimotor circuits after rhythmic whisker stimulation. J. Neurosci. 2009, 29, 5326–5335. [Google Scholar] [CrossRef] [PubMed]
- Barros-Zulaica, N.; Castejon, C.; Nunez, A. Frequency-specific response facilitation of supra and infragranular barrel cortical neurons depends on NMDA receptor activation in rats. Neuroscience 2014, 281, 178–194. [Google Scholar] [CrossRef]
- Derdikman, D.; Yu, C.; Haidarliu, S.; Bagdasarian, K.; Arieli, A.; Ahissar, E. Layer-specific touch-dependent facilitation and depression in the somatosensory cortex during active whisking. J. Neurosci. 2006, 26, 9538–9547. [Google Scholar] [CrossRef] [Green Version]
- Torres-Aleman, I. Role of insulin-like growth factors in neuronal plasticity and neuroprotection. Adv. Exp. Med. Biol. 2005, 567, 243–258. [Google Scholar]
- Kenyon, C. A conserved regulatory system for aging. Cell 2001, 105, 165–168. [Google Scholar] [CrossRef] [Green Version]
- Junnila, R.K.; List, E.O.; Berryman, D.E.; Murrey, J.W.; Kopchick, J.J. The GH/IGF-1 axis in ageing and longevity. Nat. Rev. Endocrinol. 2013, 9, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Morrison, J.H.; Baxter, M.G. The ageing cortical synapse: Hallmarks and implications for cognitive decline. Nat. Rev. Neurosci. 2012, 13, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Petralia, R.S.; Mattson, M.P.; Yao, P.J. Communication breakdown: The impact of ageing on synapse structure. Ageing Res. Rev. 2014, 14, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Burke, S.N.; Barnes, C.A. Neural plasticity in the ageing brain. Nat. Rev. Neurosci. 2006, 7, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.P.; Fernandez, A.M.; Haas, C.; Zimmer, E.; Portela, L.V.; Torres-Aleman, I. Reduced brain insulin-like growth factor I function during aging. Mol. Cell Neurosci. 2012, 49, 9–12. [Google Scholar] [CrossRef]
- Ashpole, N.M.; Sanders, J.E.; Hodges, E.L.; Yan, H.; Sonntag, W.E. Growth hormone, insulin-like growth factor-1 and the aging brain. Exp. Gerontol. 2015, 68, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Piriz, J.; Torres-Aleman, I.; Nunez, A. Independent alterations in the central and peripheral somatosensory pathways in rat diabetic neuropathy. Neuroscience 2009, 160, 402–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piriz, J.; Muller, A.; Trejo, J.L.; Torres-Aleman, I. IGF-I and the aging mammalian brain. Exp. Gerontol. 2011, 46, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Ahn, J.H.; Park, J.H.; Yan, B.C.; Kim, I.H.; Lee, D.H.; Cho, J.H.; Chen, B.H.; Lee, J.C.; Cho, J.H.; et al. Decreased insulin-like growth factor-I and its receptor expression in the hippocampus and somatosensory cortex of the aged mouse. Neurochem. Res. 2014, 39, 770–776. [Google Scholar] [CrossRef]
- Dellu, F.; Contarino, A.; Simon, H.; Koob, G.F.; Gold, L.H. Genetic differences in response to novelty and spatial memory using a two-trial recognition task in mice. Neurobiol. Learn. Mem. 2000, 73, 31–48. [Google Scholar] [CrossRef] [Green Version]
- Noriega-Prieto, J.A.; Maglio, L.E.; Zegarra-Valdivia, J.A.; Pignatelli, J.; Fernandez, A.M.; Martinez-Rachadell, L.; Fernandes, J.; Núñez, Á.; Araque, A.; Torres-Alemán, I.; et al. Astrocytic IGF-IRs Induce Adenosine-Mediated Inhibitory Downregulation and Improve Sensory Discrimination. J. Neurosci. 2021, 41, 4768–4781. [Google Scholar] [CrossRef]
- Manns, I.D.; Sakmann, B.; Brecht, M. Sub- and Suprathreshold Receptive Field Properties of Pyramidal Neurons in Layers 5A and 5B of Rat Somatosensory Barrel Cortex. J. Physiol. 2004, 556, 601–622. [Google Scholar] [CrossRef] [PubMed]
- De Kock, C.P.; Bruno, R.M.; Spors, H.; Sakmann, B. Layer- and cell-type-specific suprathreshold stimulus representation in rat primary somatosensory cortex. J. Physiol. 2007, 581, 139–154. [Google Scholar] [CrossRef] [PubMed]
- De Kock, C.P.; Sakmann, B. High frequency action potential bursts (>or= 100 Hz) in L2/3 and L5B thick tufted neurons in anaesthetized and awake rat primary somatosensory cortex. J. Physiol. 2008, 586, 3353–3364. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Alloway, K.D. Differential response patterns in the si barrel and septal compartments during mechanical whisker stimulation. J. Neurophysiol. 2009, 102, 1632–1646. [Google Scholar] [CrossRef]
- Breese, C.R.; Ingram, R.L.; Sonntag, W.E. Influence of age and long-term dietary restriction on plasma insulin-like growth factor-1 (IGF-1), IGF-1 gene expression, and IGF-1 binding proteins. J. Gerontol. 1991, 46, B180–B187. [Google Scholar] [CrossRef] [PubMed]
- D’Costa, A.P.; Ingram, R.L.; Lenham, J.E.; Sonntag, W.E. The regulation and mechanisms of action of growth hormone and insulin-like growth factor 1 during normal ageing. J. Reprod. Fertil. Suppl. 1993, 46, 87–98. [Google Scholar] [PubMed]
- Bando, H.; Zhang, C.; Takada, Y.; Yamasaki, R.; Saito, S. Impaired secretion of growth hormone-releasing hormone, growth hormone and IGF-I in elderly men. Acta Endocrinol. 1991, 124, 31–36. [Google Scholar] [CrossRef]
- Deak, F.; Sonntag, W.E. Aging, synaptic dysfunction, and insulin-like growth factor (IGF)-1. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 611–625. [Google Scholar] [CrossRef] [Green Version]
- Sonntag, W.E.; Deak, F.; Ashpole, N.; Toth, P.; Csiszar, A.; Freeman, W.; Ungvari, Z. Insulin-like growth factor-1 in CNS and cerebrovascular aging. Front. Aging Neurosci. 2013, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Sonntag, W.E.; Ramsey, M.; Carter, C.S. Growth hormone and insulin-like growth factor-1 (IGF-1) and their influence on cognitive aging. Ageing Res. Rev. 2005, 4, 195–212. [Google Scholar] [CrossRef]
- Sonntag, W.E.; Bennett, S.A.; Khan, A.S.; Thornton, P.L.; Xu, X.; Ingram, R.L.; Brunso-Bechtold, J.K. Age and insulin-like growth factor-1 modulate N-methyl-D-aspartate receptor subtype expression in rats. Brain Res. Bull. 2000, 51, 331–338. [Google Scholar] [CrossRef]
- Le Greves, M.; Le Greves, P.; Nyberg, F. Age-related effects of IGF-1 on the NMDA-, GH- and IGF-1-receptor mRNA transcripts in the rat hippocampus. Brain Res. Bull. 2005, 65, 369–374. [Google Scholar] [CrossRef]
- Pirger, Z.; Naskar, S.; Laszlo, Z.; Kemenes, G.; Reglodi, D.; Kemenes, I. Reversal of age-related learning deficiency by the vertebrate PACAP and IGF-1 in a novel invertebrate model of aging: The pond snail (Lymnaea stagnalis). J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 1331–1338. [Google Scholar] [CrossRef]
- Lupien, S.B.; Bluhm, E.J.; Ishii, D.N. Systemic insulin-like growth factor-I administration prevents cognitive impairment in diabetic rats, and brain IGF regulates learning/memory in normal adult rats. J. Neurosci. Res. 2003, 74, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Miki Stein, A.; Munive, V.; Fernandez, A.M.; Nunez, A.; Torres-Aleman, I. Acute exercise does not modify brain activity and memory performance in APP/PS1 mice. PLoS ONE 2017, 12, e0178247. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Magro, N.; Zegarra-Valdivia, J.A.; Troyas-Martinez, S.; Torres-Aleman, I.; Nuñez, A. Response Facilitation Induced by Insulin-like Growth Factor-I in the Primary Somatosensory Cortex of Mice Was Reduced in Aging. Cells 2022, 11, 717. https://doi.org/10.3390/cells11040717
García-Magro N, Zegarra-Valdivia JA, Troyas-Martinez S, Torres-Aleman I, Nuñez A. Response Facilitation Induced by Insulin-like Growth Factor-I in the Primary Somatosensory Cortex of Mice Was Reduced in Aging. Cells. 2022; 11(4):717. https://doi.org/10.3390/cells11040717
Chicago/Turabian StyleGarcía-Magro, Nuria, Jonathan A. Zegarra-Valdivia, Sara Troyas-Martinez, Ignacio Torres-Aleman, and Angel Nuñez. 2022. "Response Facilitation Induced by Insulin-like Growth Factor-I in the Primary Somatosensory Cortex of Mice Was Reduced in Aging" Cells 11, no. 4: 717. https://doi.org/10.3390/cells11040717