
Current Nutritional and Pharmacological Approaches for Attenuating Sarcopenia


1
Institute for Liberal Arts, Environment and Society, Tokyo Institute of Technology, Meguro-ku, Tokyo 152-8550, Japan
2
Department of Physical Therapy, Health Sciences University of Hokkaido, Kanazawa, Ishikari-Tobetsu, Hokkaido 061-0293, Japan
3
Laboratory of Nutrition Science, Graduate School of Life and Environmental Sciences, Kyoto Prefectural University, Kyoto 606-8522, Japan
*
Author to whom correspondence should be addressed.
Cells 2023, 12(19), 2422; https://doi.org/10.3390/cells12192422
Submission received: 28 August 2023
/
Revised: 27 September 2023
/
Accepted: 5 October 2023
/
Published: 9 October 2023
(This article belongs to the Special Issue Cellular Plasticity of the Neuromuscular System)
Abstract
:Sarcopenia is characterized by a gradual slowing of movement due to loss of muscle mass and quality, decreased power and strength, increased risk of injury from falls, and often weakness. This review will focus on recent research trends in nutritional and pharmacological approaches to controlling sarcopenia. Because nutritional studies in humans are fairly limited, this paper includes many results from nutritional studies in mammals. The combination of resistance training with supplements containing amino acids is the gold standard for preventing sarcopenia. Amino acid (HMB) supplementation alone has no significant effect on muscle strength or muscle mass in sarcopenia, but the combination of HMB and exercise (whole body vibration stimulation) is likely to be effective. Tea catechins, soy isoflavones, and ursolic acid are interesting candidates for reducing sarcopenia, but both more detailed basic research on this treatment and clinical studies in humans are needed. Vitamin D supplementation has been shown not to improve sarcopenia in elderly individuals who are not vitamin D-deficient. Myostatin inhibitory drugs have been tried in many neuromuscular diseases, but increases in muscle mass and strength are less likely to be expected. Validation of myostatin inhibitory antibodies in patients with sarcopenia has been positive, but excessive expectations are not warranted.
1. Introduction
Skeletal muscle contraction is the driving force behind physical movement and plays an important role in homeostasis. Sarcopenia occurs as a consequence of age-related loss of muscle strength, quality and quantity. It often refers to cellular processes (mitochondrial dysfunction, inflammation, and denervation) and the resulting loss of muscle strength, function and mobility, and increased risk of falls. Sarcopenia is considered ‘primary’ (or age-related) when there is no apparent cause other than age [1]. Secondary sarcopenia usually occurs when multiple causes are present. This condition is commonly observed in the elderly and is due to diabetes, stroke, hip fracture, chronic obstructive pulmonary disease, and chronic heart failure [2]. Muscle mass loss is more pronounced in lower limb muscle groups than in upper limb muscle groups and, at the myofiber level, is characterized by selective type II muscle fiber atrophy and fibronecrosis.
The mechanism of age-related muscle atrophy is thought to be influenced simultaneously by several factors, including poor muscle regeneration, hormonal changes, nutritional deficiencies, and increased oxidative stress. Although the specific contribution of these factors remains to be elucidated, it has been reported that muscle hypertrophy regulators become less responsive with age [3], and dysfunction of autophagy is also evident [4,5]. In contrast, no apparent changes in negative regulators (atrophy gene-1 (atrogin-1), myostatin, NF-κB (nuclear factor-κB)) have occurred in ageing mammalian muscle [6,7]. Strategies such as nutritional supplementation and physical training (both aerobic and resistance exercise) are important interventions for muscle atrophy in the elderly.
Dietary protein provides amino acids necessary for muscle protein synthesis and, as an anabolic stimulus, acts directly on protein synthesis. There is a considerable literature on the role of mixtures containing all essential amino acids (EAAs) in muscle metabolism and prevention of sarcopenia, both in experimental models and in humans [8,9]. The administration of multiple essential amino acids also increases muscle mass and protein synthesis, even under normal conditions without resistance training [8]. In addition, a number of reviews suggest that nutritional interventions such as high protein intake and intake of leucine, a branched-chain amino acid, combined with resistance training would be effective in reducing muscle fiber atrophy in sarcopenia [10,11]. On the other hand, mild calorie restriction (CR) attenuates sarcopenia in all mammals quite effectively [12,13]. Recent studies have also indicated that new supplements (e.g., ursolic acid, resveratrol, and soy isoflavones) could be applied to prevent muscle atrophy [14,15]. Furthermore, despite many negative reports, pharmacological sarcopenia treatment strategies have also attracted the attention of many researchers. This chapter details recent nutritional and pharmacological approaches to control sarcopenia using the database of PubMed.
2. Nutritional Approach
2.1. HMB
Protein supplementation/a protein-rich diet is conditionally recommended by clinical practice guidelines for sarcopenia based on the International Conference on Sarcopenia and Frailty Research (ICFSR) [16]. In addition, in the consensus statements of Asian Working Group for Sarcopenia, high-quality protein, amino acids such as leucine and L-carnitine, or oral nutritional supplement (ONS) containing beta-hydroxy-beta-methylbutyrate (HMB) may be considered and should be taken according to the specific prescribing information [17]. HMB is a metabolite of leucine, a branched-chain amino acid, and HMB stimulates protein synthesis via mTOR and decreases myonuclear apoptosis and proteasome expression. Supplementation with HMB is demonstrated to increase fat oxidation and mitochondrial biogenesis [18]. The effects of HMB supplementation in humans has been examined by many researchers. Deutz et al. [19] reported positive results using 10 days of HMB (3 g) supplementation for unloading patients (bed rest). Specifically, HMB intake prevented muscle weakness during unloading and maintained muscle strength during the rehabilitation period. The HMB intake used in this study was a dose commonly used in academic research, and the subjects were diet-controlled. In addition, Hsieh et al. [20] observed the positive effects of HMB intake (a decrease in blood urea and urinary urea excretion) during a 2-to-4-week intervention period. Notably, they suggested a decrease in proteolysis despite no significant change in anthropometric parameters due to HMB supplementation. However, their study had some problems, including a short duration of supplementation. In contrast, a meta-analysis of several randomized controlled trials (RCTs) showed that HMB supplementation for the elderly prevented loss of lean body mass without changing fat mass, and HMB may be more effective by combining it with other nutrients such as lysine and arginine. Specifically, 12 weeks of HMB/Arg/Lys mixture (2/5/1.5 g per day) has been reported to increase whole body protein synthesis, leg and grip strength, and limb circumference [21]. However, their study found no increase in muscle strength, thus negating the effect of HMB supplementation on muscle strength. Furthermore, a high-quality systematic review suggested no significant effect of HMB supplementation on muscle strength [22]. However, this meta-analysis showed a great deal of evidence suggesting the effect of HMB supplementation on muscle mass. In summary, HMB supplementation for older adults has a positive effect on muscle mass but has no clear effect on physical performance and muscle strength. From the two reviews, where evidence is somewhat lacking, it has not been determined whether HMB alone is effective in improving muscle strength, physical performance, or muscle mass [22], and further research is needed regarding the effects of HMB supplementation on reducing sarcopenia.
Whether HMB alone improves symptoms of sarcopenia has not been clearly elucidated, but HMB and vibration stimulation appear to significantly improve sarcopenia. Whole-body and local vibration are often used to improve muscle weakness in the elderly. An RCT of men with sarcopenia (mean age 88.6 years) showed that vibration (12–16 Hz, 3–5 mm) for 8 weeks significantly improved muscle strength and physical performance (walking speed, 5-stance test, and timed stand test) [23]. Although systematic reviews and meta-analyses have shown that vibration stimulation therapy is an effective strategy to improve physical performance and muscle strength in patients with sarcopenia, caution is necessary when interpreting the results because of the limited number of publications on this topic. Wang et al. [24] recently showed that LMHFV significantly improves skeletal muscle quality by suppressing myostatin expression and reducing intramuscular fat mass in aging mice (SAMP8). In particular, vibration stimulation combined with HMB significantly increased lean body mass and improved muscle performance (single contraction and muscle strength).
2.2. Polyphenol
2.2.1. Catechin
Tea (Camellia sinesis), especially green tea, which is primarily composed of catechins, contains many phenolic hydroxyl groups (-OH). There are four monomers of catechins: epigallocatechin gallate (EGCG), epicatechin (EC), epigallocatechin (EGC), and epicatechin gallate (ECG). Green tea catechins have been shown to act on skeletal muscle cells and may inhibit muscle mass loss. Supplementation with EGCG (5 mg/kg, 4 times/week) for 8 weeks appears to reduce fibrosis and necrotic muscle fibers in muscular dystrophic mice [25]. EGCG supplementation also decreases protein carbonyl content (an oxidative stress marker) in aged male rats [26]. However, some reports suggest that the effects on muscle adaptation depend on the type of catechin being supplemented. In an experiment in which 20-month-old male mice were fed EC or EGCG (0.25% in drinking water) and a standard diet, survival significantly increased from 39% to 69% after 37 weeks of EC consumption, whereas no significant effect was observed with EGCG [27]. In addition, EC slowed skeletal muscle degeneration and improved physical activity in aged mice, indicating that the effects of EC on skeletal muscle may be more pronounced than those of EGCG.
A recent double-blind, controlled trial was performed to examine the effects of green tea in humans using 62 men. They randomized four groups to receive epicatechin for 8 weeks: resistance training (RT), 1 mg of epicatechin (EC) per kg of body weight, resistance training plus epicatechin (RT + EC), and placebo (PL). The results showed significant increases in follistatin, follistatin/myostatin ratio, and muscle strength (leg press and chest press) in the RT + EC group compared to those in the other three groups. In a randomized controlled trial, 128 women with sarcopenia were randomly assigned to four groups: exercise and tea catechin intake (350 mL/day), exercise, tea catechin supplementation, and 3 months of health education. After the intervention, a significant effect on the composite variables of leg muscle mass and normal walking speed was observed in the exercise and catechin intake group compared the health education group [28].
Lychee extract (oligonol), rich in phenolic compounds (flavonols), prevents kidney damage caused by diabetes and obesity caused by a high-fat diet [29]. The addition of oligonol decreased the expression of PGC-1α and Mfn2 and increased the expression of LC3-II, p62, and ATG13, and showed autophagosome and lysosome accumulation [30]. Although numerous studies have investigated the effects of green tea extract supplementation (EC, EGCG, oligonol, etc.) on mitochondria and muscle fibers, almost all studies have been conducted in experimental animals. The safety, bioavailability, and efficacy of green tea should be tested in patients with sarcopenia. There is insufficient evidence for the effect of catechins on sarcopenia patients, as most clinical studies examining the effect of green tea extract in sarcopenia patients have not been RCT studies.
2.2.2. Isoflavones
Isoflavones, a type of flavonoid found in soybeans, are structurally similar to estrogens and exert their physiological functions, such as antioxidant effects, by binding to estrogen receptors. Estrogen receptors (ERs) include ERα and ERβ, and daidzein, a soy isoflavone, acts on ERβ, but not ERα. In post-oophorectomy mice, muscle mass increases when isoflavones are added to a high-fat diet for 12 weeks [31]. In contrast, long-term (120 days) isoflavone intake decreases fat accumulation in the skeletal muscle of male mice [32]. Notably, isoflavones affect muscle mass and function in both male and female mice. Many researchers examined the effects of isoflavones on muscle mass but have not observed the changes in muscle fiber size. Moreover, accumulating large amounts of fat and connective tissue in atrophied tissue may render muscle weight-based assessment methods inaccurate. Abe et al. [33] showed that isoflavone administration (20% of the diet) significantly reduced muscle fiber atrophy in the tibialis anterior muscle on day 4 after denervation. Furthermore, isoflavone treatment caused increased expression of IRS-1 and p-Akt1 proteins in mouse denervated muscle.
Most in vivo isoflavone supplementation experiments have used isoflavones in amounts greater than 1% of dietary intake [31,32], but it is difficult for humans to consume such large amounts of isoflavones in every meal. Therefore, testing the effects of isoflavones beyond 1% of dietary intake may be meaningless. Our group found that administration of 0.6% isoflavone (AglyMax, an isoflavone aglycon mixture of daidzein, genistin, and glycitein in a 7:1:2 ratio) inhibited muscle fiber atrophy after denervation in mice [34]. This effect is thought to be due to a reduction in apoptosis-dependent signaling rather than atrogin-1 pathway-dependent signaling.
Intake of daidzein may be beneficial for older women. For example, 8-prenylnaringenin has estrogenic effects similar to daidzein and appears to inhibit denervation-induced skeletal muscle atrophy by promoting Akt phosphorylation. Quercetin, a type of flavonoid, is also abundant in vegetables and fruits and has a scavenging effect on reactive oxygen species (ROS). Since ROS activate NFκB and Foxo pathways and induce the expression of E3 ubiquitin ligase, quercetin’s ability to suppress muscle atrophy has attracted attention. For example, the ingestion of quercetin in hindlimb-unloaded mice reduces the expression of atrogin-1 and MuRF-1 in gastrocnemius muscle and prevents the loss of skeletal muscle mass [35]. Future studies are required to elucidate whether daidzein, quercetin, and AglyMax inhibit muscle aging.
2.3. Ursolic Acid (UA)
Ursolic acid (UA) is a natural pentacyclic triterpenoid carboxylic acid found in plants and fruits such as apples, blueberry and cranberry. UA has anti-carcinogenic, antioxidant, anti-inflammatory, and anti-obesity effects, and UA has also been shown to have beneficial effects in animal models of hyperlipidemia and diabetes. A systematic review involving animal studies concluded that the supplementation with UA increases muscle strength and muscle mass by increasing mTOR pathway activation in muscle satellite cells. UA has been reported to reduce the expression levels of two muscle atrophy genes (atrogin-1 and MuRF1) and reduce muscle atrophy caused by fasting and atrophy-induced stress [36]. UA supplementation also increases Akt phosphorylation in skeletal muscle in vivo but UA may not act directly on skeletal muscle because UA alone is not sufficient to promote IGF-I and insulin receptor activation. Yu et al. [37] orally administered UA (100 mg/kg) to mice models of chronic kidney disease (CKD) for 3 weeks. They found that UA administration reduced the expression of inflammatory cytokines (e.g., IL-6 and TNF-α) and ubiquitin E3 ligases (e.g., MuRF-1, atrogin-1, and MUSA1) and suppressed CKD-induced muscle atrophy (tibialis anterior muscle).
Moreover, supplementation with UA is highly effective for improving muscle tissue in combination with resistance and endurance exercise. Endurance training and UA (ET + UA) decreased the body weight, insulin resistance, and insulin and glucose concentrations more than the other groups [38]. Furthermore, the ATF4 protein level was significantly reduced in ET + UA compared with DM. ET + UA reduced the amount of p53 protein similar to the significant decrease in p21 protein, which was diminished more than that in any other group. In contrast, the amount of p21 in the RT + UA group was not more significant than that in UA group. Collectively, these findings show that exercise and UA supplementation impeded the interactions among p53/ATF4/p21.
The effects of UA intake on skeletal muscle have not been well tested in humans. UA supplementation (loquat leaf extract intake, 50 mg/day) in healthy adults did not significantly improve muscle mass and strength [39]. In addition, the combination of strength training and 8 weeks of UA supplementation increased muscle strength but not lean body mass compared with strength training alone [40]. Further studies are needed to clarify the effects of UA supplementation on sarcopenia in humans.
2.4. Omega-3 PUFAs
Lipids such as omega-3 fatty acids and n-3 polyunsaturated fatty acids have been suggested to affect muscle health indirectly by preventing low-grade inflammation [41]. Intriguingly, significant lower levels of serum 3 fatty acids are associated with sarcopenia, higher dietary lipid intake correlates with lower odds of sarcopenia [42]. More recently, AWGS consensus guidelines found scarce evidence to support a statement around dietary/supplementary lipid intake in muscle health [17]. Omega-3 PUFAs include alpha-linolenic acid (ALA), eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA), and docosapentaenoic acid (DPA). Concentrations of DHA and EPA in cell membranes are important to ensure structural cell function. The primary dietary sources of DHA and EPA are seafood and fatty fish (e.g., mackerel). ALA is found in plant foods such as vegetable oils, chia seeds, and nuts. Although 500 mg of DHA and EPA are recommended daily, most U.S. adults do not achieve this intake. Omega-3 PUFAs are of great interest because of their anti-inflammatory properties and low risk of adverse events. Thus, supplementation with omega-3 PUFAs is beneficial for a variety of aging processes, including cognition, eye health, and bone metabolism. Omega-3 PUFAs may also have beneficial effects in muscles, including hypertrophy of muscle fiber and improvement of mitochondrial function. Furthermore, RCT studies have indicated that supplementation with omega-3 PUFAs promotes muscle protein synthesis in the elderly [43]. The relationship between dietary intake of omega-3 PUFAs and physical performance has also been studied in a cross-sectional study: 2893 men and women (aged 59–73 years) included in the Hertfordshire Cohort study were examined to determine the association between grip strength and dietary intake. The study showed a relationship between each additional serving of fatty fish and increased grip strength in both sexes [44].
The anti-inflammatory effects of omega-3 PUFAs are generally accepted. Supplementation of omega-3 PUFAs in middle-aged and older adults causes significant IL6 and CRP reductions. In addition, an RCT study with elderly subjects showed that treatment with EPA and DHA (for 8 weeks) significantly reduced blood levels of TNFα, IL-6, and IL-1β [45]. Suppression of inflammation by omega-3 PUFAs may be one mechanism to combat sarcopenia because chronic low-level inflammation is involved in its development. In contrast, there are reports stating that supplementation with omega-3 PUFAs does not affect muscle protein synthesis [46], and further studies are needed in elderly subjects with sarcopenia. Moreover, Smith et al. found more amino acid and insulin-stimulated activation of the mTOR-p70S6K pathway after 8 weeks of omega-3 PUFA supplementation compared with a placebo group. Omega-3 PUFA supplementation also produces important changes in gene expression that promote skeletal muscle anabolism by decreasing the expression of the mTOR inhibitory pathway. Thus, omega-3 PUFAs have the potential to overcome age-related anabolic resistance through the activation of mTOR signaling. Figure 1 represents an overview of nutritional intervention for sarcopenia. Table 1 summarizes the nutritional approaches for attenuating muscle atrophy.
3. Pharmacological Approach
ICFSR clinical practice guidelines indicate that pharmacological interventions are not recommended as the first-line therapy for the management of sarcopenia [16].
3.1. Vitamin D
Vitamin D is an important regulator of bone metabolism, including phosphorus and calcium homeostasis. Vitamin D deficiency and loss of muscle mass often occur together; it is estimated that approximately 50% of individuals over 65 years of age have hypovitaminosis D levels. Although the mechanism remains unclear, vitamin D appears to influence muscle function and strength through the vitamin D receptor. Notably, the expression of vitamin D receptors in skeletal muscle decreases with age.
A double-blind study of nursing home residents and older adults living in areas with low vitamin D has shown that supplementation with vitamin D reduces the risk of falls and improves muscle performance and strength [47]. Many epidemiological studies have also shown that lower extremity function is impaired in elderly patients with hypovitaminosis D status, including delayed walking time and time from sitting to standing [48]. To prevent sarcopenia, it may be useful to clarify the effects of vitamin D on skeletal muscle function, strength, and muscle mass. However, a recent review [49] using eligible RCTs clearly denied the positive effect of vitamin D monotherapy in older adults (age 50 years and older). A random-effects inverse-variable model was used to calculate the mean difference between groups for handgrip strength (HGS), timed up-and-go (TUG), and appendicular lean mass (ALM). Significant reductions in short physical performance battery scores were observed for vitamin D supplementation compared with the placebo. Vitamin D supplementation did not affect TUG, HGS, ALM, general muscle strength, or general physical performance. Therefore, vitamin D supplementation has been shown not to improve sarcopenia in elderly individuals who are not vitamin D-deficient. This conclusion would be consistent with the ICFSR clinical practice guideline that there is insufficient evidence to determine whether vitamin D replacement therapy is effective in elderly patients with sarcopenia [16].
3.2. Myostatin Inhibition
Myostatin, a novel member of the transforming growth factor beta superfamily, negatively regulates muscle growth [50]. Myostatin expression in muscle is clearly increased in CKD mice and rats [51,52]. However, conflicting results have been obtained as to whether myostatin expression is increased in sarcopenic muscle [6,53]. Many investigators have evaluated the effects of myostatin inhibition in muscle disease models such as cancer cachexia, CKD, Duchenne muscular dystrophy (DMD), and amyotrophic lateral sclerosis [54]. For example. subcutaneous injection of anti-myostatin peptides in CKD mice restored weight loss (5–7% increase) and muscle mass loss (~10% increase) in the hindlimbs, including the TA, gastrocnemius, EDL, and soleus muscles. Myostatin inhibition also suppressed circulating inflammatory cytokines (TNF-α, M-CSF-1, etc.) and the mRNA expression of these cytokines in CKD mice.
The effects of pharmacological inhibition of myostatin in DMD patients have been examined by several researchers but positive effects have been difficult to obtain and have little potential for clinical application. Furthermore, a randomized clinical trial of the same compound in DMD patients showed improved muscle mass and performance (6 min walk test), but was stopped early owing to non-muscular side effects (nosebleeds, capillary dilation, etc.) [55]. LY2495655, a phase II humanized antibody against myostatin, showed a dose-dependent and significant increase in lean mass compared with the placebo at weeks 8 and 16 after an RCT of total hip replacements in 400 patients [56]. Becker et al. [57] conducted a randomized phase II trial for the subcutaneous injection of a myostatin antibody (LY2495655: LY, 315 mg) in frail elderly subjects worldwide (e.g., the US, France, and Australia), aged 75 years and older. After 24 weeks, the LY group showed a significant increase in the least-squares mean change in aLBM compared with the placebo group. In addition, functional characteristics, such as stair climbing time and chair standing with arms, improved from baseline after 24 weeks of LY treatment in elderly subjects [57]. Based on these results, antibody inhibition of myostatin may be effective in treating sarcopenia, but the results of many clinical trials do not support this notion. Although beneficial effects in terms of fat loss and glucose metabolism have been consistently observed in many clinical trials with antimyostatin drugs, many have been disappointing in terms of increased muscle mass and functional improvement [58].
Researchers are particularly interested in the role of GDF11, which bears many similarities to myostatin in terms of the amino acid sequence and receptor and signaling pathways. For example, GDF11 has been reported to inhibit myocardial hypertrophy and decrease with age [59]. Additionally, supplementation of aged mice with GDF11 had a positive effect on aging satellite cells and improved skeletal muscle regeneration [60]. Some of the early studies reported the rejuvenating effects of GDF11 in aging mice and suggested that GDF11 can restore the skeletal muscle, heart tissue, and cerebrovascular system to a young and healthy state [61]. However, many subsequent studies have shown that blood GDF11 levels are either unaffected by aging or increase, with contradicts the previous studies.
Early studies also suggested that GDF11, like myostatin, inhibits satellite cell proliferation, induces differentiation, and inhibits muscle regeneration. However, increased GDF11 was found to be a risk factor for frailty and cardiovascular disease [61], demonstrating that exogenous GDF11 induces skeletal and cardiac muscle decline in mice. These contradictory findings regarding the role of GDF11 may be the result of using E. coli to create the recombinant GDF11 protein [59,60]. E. coli does not have an oxidative environment and thus cannot form the disulfide bond that connects the two GDF11 monomers. The use of GDF11 recombinant protein produced in E. coli in experiments with proper physiological function verification may lead to more reliable results.
3.3. Anabolic Steroids
The current evidence is insufficient to recommend anabolic hormones for the management of sarcopenia in ICFSR consensus guidelines [16]. Testosterone levels in men decrease by 1% per year after age 30, and bioavailable testosterone levels decrease by 2% per year [62]. Blood levels of testosterone in women decline rapidly between the ages of 20 and 45. With renal impairment (progression of CKD stage), testosterone levels markedly decrease [63], and the decrease in testosterone levels in CKD patients is significantly correlated with levels of sperm cytoplasmic droplets and total neutral glucosidase activity [63]. A systematic review [64] has shown that testosterone supplementation attenuates sarcopenia characteristics such as decreased muscle mass [65] and grip strength [66]. In a placebo-controlled study of long-term (6 months) supraphysiologic testosterone administration, leg lean body mass and leg and arm muscle strength increased [67]. Older men receiving high doses of testosterone show significant increases in muscle strength, but the potential risks outweigh the benefits. Increased risk of thrombotic complications, sleep apnea, and prostate cancer are associated with testosterone therapy in older men.
New nonsteroidal compounds called selective androgen receptor modulators SARMs exhibit tissue-selective activity, improved pharmacokinetic properties, and fewer systemic side effects. The effects of SARMs have been tested in healthy older men and patients in cancer-related sarcopenia [68,69]. In fact, a phase II double-blind study of enobosam, a nonsteroidal SARM, showed improvements in physical function (stair climbing speed) and LBM in healthy postmenopausal elderly men [70]. This study showed that as little as 1 mg of enobosarm increased LBM in patients with advanced cancer. Another SARM, GSK2881078, administered once daily for 50 days to healthy elderly men and women, significantly increased lean body mass without significant side effects, although alanine aminotransferase was transiently elevated [70]. More rigorous clinical trials are necessary to determine whether these agents are effective in normal and CKD-induced sarcopenia.
Supasyndh et al. [71] evaluated supplementation with oxymetholone, a milder anabolic steroid, in a 24-week RCT with 43 hemodialysis patients and observed significant increases in fat-free volume, subjective scores of physical function, and grip strength. Hemodialysis patients showed atrophy of type I and type II muscle fibers and decreased IGF-IR mRNA levels after 24 weeks of placebo treatment, but this was suppressed by oxymetholone, which induced hypertrophy of type I muscle fibers. Recently, Kim et al. studied the effects of oxymetholone (50 mg/kg) on old mice after 28 days of chronic forced exercise [72]. Glutathione, superoxide dismutase, and ROS were partially abundant owing to decreased catalase activity, but were restored to baseline levels by oxymetholone supplementation. In addition, oxymetholone treatment suppressed the expression of myostatin, sirtuin 1, and muscle atrophy genes and induced fiber hypertrophy in the gastrocnemius and soleus muscles [72]. Thus, there are few reports of oxymetholone treating sarcopenia or CKD in mouse models or in humans. These results should be clarified to assess the benefits and risks to sarcopenia due to normal or CKD with treatment with anabolic steroids, androgens, or similar substances.
3.4. Glucose-Lowering Drugs
3.4.1. Metformin
Metformin is the most commonly prescribed antidiabetic drug and has been reported to activate AMPK and stimulate PGC1α gene expression in skeletal muscle. This promotes the transcription of genes associated with fatty acid oxidation and suppresses intramuscular fat accumulation. Activation of the AMPK pathway also promotes angiogenesis, mitochondrial biosynthesis, and the conversion of myofiber types from glycolytic to oxidative fibers [73,74,75]. Furthermore, it has been reported that Metformin suppresses the expression of mTORC1-related genes in the skeletal muscle of elderly individuals with impaired glucose tolerance [76]. Since autophagy is dysfunctional in sarcopenia, activating AMPK upstream of the autophagy pathway and suppressing mTORC1 may promote metabolism, resulting in an anti-aging effect.
Clinical trials have shown that Metformin treatment (3 × 500 mg for 16 weeks) in elderly patients with diabetes improved walking speed by 0.13 m/s [77]. Because most of the reports from clinical trials have been in patients with diabetes, there is insufficient evidence for Metformin’s treatment of sarcopenia. However, it may have a therapeutic effect through the aforementioned signaling pathways.
3.4.2. Thiazolidinediones (TZDs)
TZDs increase the expression of adiponectin and its receptors in the blood and increase the expression of genes related to mitochondrial function and fatty acid oxidation by activating AMPK and ACC in skeletal muscle [78]. This results in increased skeletal muscle insulin sensitivity [79]. In a clinical trial in non-diabetic patients, men treated with pioglitazone showed a decrease in thigh muscle mass compared with men who did not receive pioglitazone [80]. TZDs may not be effective enough for the treatment of sarcopenia.
3.4.3. Sulfonylureas
Sulfonylurea acts on pancreatic beta cells to promote insulin secretion and has been reported to decrease muscle protein in rat flexor digitorum brevis muscle through caspase-3-dependent or -independent pathways in in vitro experiments [81]. In a cross-sectional study in diabetic patients, sarcopenia parameters (skeletal muscle mass, skeletal muscle mass index (SMI), muscle strength, and gait speed) were reported to worsen more with sulfonylureas compared with DPP-4i [82], and there were no changes in skeletal muscle mass in diabetic patients in clinical trials [83]. The effect on sarcopenia is not clear, but likely does not contribute to its improvement.
3.4.4. Dipeptidyl Peptidase-4 Inhibitor (DPP-4i)
DPP-4i promotes insulin secretion and inhibits glucagon release. In skeletal muscle, DPP-4i has been reported to increase GLUT4 expression in soleus and gastrocnemius muscles [84], promote glucose uptake [85], and enhance mitochondrial biosynthesis to improve exercise capacity [86] in animal models. Epidemiological studies have reported that DPP-4i treatment of elderly diabetic patients shows improved sarcopenia parameters compared with other diabetes medications [78], but may not be sufficient to improve non-diabetic sarcopenia.
3.4.5. Glucagon-like Peptide-1 Receptor Agonist (GLP-1 Ras)
GLP-1 RAs bind specifically to GLP-1 receptors and stimulate insulin secretion from pancreatic beta cells. For skeletal muscle, they increase GLUT4 gene expression, glucose uptake, and glycogen synthesis [87]. GLP-1 RAs have been reported to promote skeletal muscle oxygen consumption and increase insulin sensitivity [88]. In a study of diabetic patients, six months of treatment with teneligliptin (20 mg/day), a DPP-4i, decreased the SMI more efficiently than that with dulaglutide (0.75 mg/week), a GLP-1 RA [89]. Although 24 weeks of dulaglutide (3 g/day) treatment in obese elderly patients with diabetes did not show an exacerbation of sarcopenia parameters [90], it is unlikely that GLP-1 RAs improve sarcopenia.
3.4.6. Sodium-Glucose Transporter Protein 2 Inhibitors (SGLT2i)
SGLT2 is expressed in the renal proximal tubules and is responsible for glucose reabsorption. Thus, SGLT2i has a hypoglycemic effect by promoting the excretion of glucose into the urine. For skeletal muscle, long-term administration of SGLT2i has been reported to improve insulin sensitivity, inhibit muscle catabolism, and improve the quality of muscle function [91]. According to a study showing improvement in maximum grip strength, improvement in mitochondrial function is considered to be the molecular mechanism that explains this effect [92]. However, it has been suggested that muscle proteins may be degraded and sarcopenia may occur because of the supply of amino acids to the liver [93]. Results of clinical trials using SGLT2i have shown that it reduces the SMI [94,95], with few reports showing improvement in sarcopenia parameters [96]. Therefore, SGLT2i is unlikely to improve sarcopenia.
3.4.7. Insulin
Insulin acts on skeletal muscle to promote protein synthesis. It also takes up glucose and promotes glycogen synthesis. Anti-insulinemia for younger patients stimulates muscle protein synthesis, but the response is found to be blunted in the elderly [97]. Observational studies have reported that insulin administration prevents a decrease in SMI in diabetic patients [98]. In addition, a 3-year follow-up study of 118 diabetic patients has shown that insulin produces a positive effect on skeletal muscle mass compared with other oral hypoglycemic agents [99]. However, all of these reports were in diabetic patients and would not improve sarcopenia. Figure 2 represents an overview of glucose-lowering drugs for sarcopenia. Table 2 summarizes the pharmacological approach for attenuating muscle atrophy.
4. Conclusions
Recent advances in our understanding of muscle biology have revealed molecular mechanisms of sarcopenia and candidate nutritional therapies. Although the combination of resistance training and supplements containing amino acids is usually recommended to prevent age-related muscle weakness and loss [10,11], protein (amino acid)-only supplements had no effect on the symptoms of sarcopenia. Many candidate substances, such as catechins, soy isoflavones, and ursolic acid, have the potential to combat sarcopenia, but even in rodent models of sarcopenia, no systematic and basic research has been conducted on this treatment. Myostatin is a potent inhibitor gene of muscle hypertrophy, and its inhibitors have been tried in many neuromuscular diseases. The effect of myostatin inhibitors on sarcopenia has been positive so far, but do not over-expect it. A more recent scoping review highlights the importance of future research focused on the potential use of Zn in the prevention and management of malnutrition, sarcopenia, and frailty in the elderly [101]. Little is known about whether this micronutrient’s intake improves sarcopenia, which will be an interesting area of research in the future.
Author Contributions
Writing—original draft preparation, K.S. and K.H.; writing—review and editing, A.Y.; funding acquisition, W.A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a research Grant-in-Aid for Scientific Research C (No. 23K10659) from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Data Availability Statement
We confirm that our MS excluded unpublished data and figures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cruz-Jentoft, A.J.; Baeyens, J.P.; Bauer, J.M.; Boirie, Y.; Cederholm, T.; Landi, F.; Martin, F.C.; Michel, J.P.; Rolland, Y.; Schneider, S.M.; et al. Sarcopenia: European consensus on definition and diagnosis: Report of the European working group on sarcopenia in older people. Age Ageing 2010, 39, 412–423. [Google Scholar] [CrossRef] [PubMed]
- von Haehling, S.; Morley, J.E.; Anker, S.D. An overview of sarcopenia: Facts and numbers on prevalence and clinical impact. J. Cachexia Sarcopenia Muscle 2010, 1, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Parkington, J.D.; LeBrasseur, N.K.; Siebelt, A.P.; Fielding, R.A. Contraction-mediated mTOR, p70S6K, and ERK1/2 phosphorylation in aged skeletal muscle. J. Appl. Physiol. 2004, 97, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Carnio, S.; LoVerso, F.; Baraibar, M.A.; Longa, E.; Khan, M.M.; Maffei, M.; Reischl, M.; Canepari, M.; Loefler, S.; Kern, H.; et al. Autophagy impairment in muscle induces neuromuscular junction degeneration and precocious aging. Cell Rep. 2014, 8, 1509–1521. [Google Scholar] [CrossRef]
- Sakuma, K.; Kinoshita, M.; Ito, Y.; Aizawa, M.; Aoi, W.; Yamaguchi, A. p62/SQSTM1 but not LC3 is accumulated in sarcopenic muscle of mice. J. Cachexia Sarcopenia Muscle 2016, 7, 204–212. [Google Scholar] [CrossRef]
- Sakuma, K.; Aoi, W.; Yamaguchi, A. Molecular mechanism of sarcopenia and cachexia: Recent research advances. Pflügers Arch. 2017, 469, 573–591. [Google Scholar] [CrossRef]
- Sandri, M.; Barberi, L.; Bijlsma, A.Y.; Blaauw, B.; Dyar, K.A.; Milan, G.; Mammucari, C.; Meskers, C.G.; Pallafacchina, G.; Paoli, A.; et al. Signaling pathways regulating muscle mass in ageing skeletal muscle. The role of IGF-1-Akt-mTOR-FoxO pathway. Biogerontology 2013, 14, 303–323. [Google Scholar] [CrossRef]
- Nicastro, H.; Artioli, G.G.; Dos Santos Costa, A.; Sollis, M.Y.; Da Luz, C.R.; Blachier, F.; Lancha, A.H., Jr. An overview of the ther apeutic effects of leucine supplementation on skeletal muscle under atrophic conditions. Amino Acids 2011, 40, 287–300. [Google Scholar] [CrossRef]
- Ruocco, C.; Segala, A.; Valerio, A.; Nisoli, E. Essential amino acid formulations to prevent mitohchondrial dysfunction and oxidative stress. Curr. Opin. Clin. Nutr. Metab. Care 2021, 24, 88–95. [Google Scholar] [CrossRef]
- Dickinson, J.M.; Reidy, P.T.; Gundermann, D.M.; Borack, M.S.; Walker, D.K.; D’Lugos, A.C.; Volpi, E.; Rasmussen, B.B. The impact of postexercise essential amino acid ingestion on the ubiquitin proteasome and autophagosomal-lysosomal systems in skeletal muscle of older men. J. Appl. Physiol. 2017, 122, 620–630. [Google Scholar] [CrossRef]
- Timmerman, K.L.; Volpi, E. Amino acid metabolism and regulatory effets in aging. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Mattison, J.A.; Roth, G.S.; Beasley, T.M.; Tilmont, E.M.; Handy, A.M.; Herbert, R.L.; Longo, D.L.; Allison, D.B.; Young, J.E.; Bryant, M.; et al. Impact of caloric restriction on health and survival in rhesus monkeys from the NIA study. Nature 2012, 489, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, S.E.; Seo, A.Y.; Marzetti, E.; Lees, H.A.; Leeuwenburgh, C. Skeletal muscle autophagy and apoptosis during aging: Effects of calorie restriction and life-long exercise. Exp. Gerontol. 2010, 45, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, K.; Yamaguchi, A. Recent advances in pharmacological, hormonal, nutritional intervention for sarcopenia. Pflügers Arch. 2018, 470, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, K.; Yamaguchi, A.; Shimizu, M. Sarcopenia: Molecular mechanism and current nutritional approach. In Encyclopedia of Human Nutrition; Caballero, B., Ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2023; Volume 3, pp. 633–644. [Google Scholar]
- Dent, E.; Morley, J.E.; Cruz-Jentoft, A.J.; Arai, H.; Kritchevsky, S.B.; Guralnik, J.; Bauer, J.M.; Pahor, M.; Clark, B.C.; Cesari, M.; et al. International clinical practice guidelines for sarcopenia (ICFSR): Screening, diagnosis and management. J. Nutr. Health Aging 2018, 22, 1148–1161. [Google Scholar] [CrossRef]
- Chen, L.K.; Arai, H.; Assantachai, P.; Akishita, M.; Chew, S.T.H.; Dumlao, L.C.; Duque, G.; Woo, J. Roles of nutrition in muscle health of community-dwelling older adults: Evidence-based expert consensus from Asian Working for Sarcopenia. J. Cachexia Sarcopenia Muscle 2022, 13, 1653–1672. [Google Scholar] [CrossRef]
- He, X.; Duan, Y.; Yao, K.; Li, F.; Hou, Y.; Wu, G.; Yin, Y. β-hydroxyl-β-methylbutyrate, mitochondrial biogenesis, and skeletal muscle health. Amino Acids 2016, 48, 653–664. [Google Scholar] [CrossRef]
- Deutz, N.E.P.; Pereira, S.L.; Hays, N.P.; Oliver, J.S.; Edens, N.K.; Evans, C.M.; Wolfe, R.R. Effect of β-hydroxy-β-methylbutyrate (HMB) on lean body mass during 10 days of bed rest in older adults. Clin. Nutr. 2013, 32, 704–712. [Google Scholar] [CrossRef]
- Hsieh, L.C.; Chow, C.J.; Chang, W.C.; Liu, T.H.; Chang, C.K. Effect of beta-hydrox-beta-methylbutyrate on protein metabolism in bed-ridden elderly receiving tube feeding. Asia Pac. J. Clin. Nutr. 2010, 19, 200–208. [Google Scholar]
- Flakoll, P.; Sharp, R.; Baier, S.; Levenhagen, D.; Carr, C.; Nissen, S. Effect of beta-hydroxyl-beta-methylbutyrate, arginine, and lysine supplementation on strength, functionality, body composition, and protein metabolism in elderly women. Nutrition 2004, 20, 445–451. [Google Scholar] [CrossRef]
- Wu, H.; Xia, Y.; Jiang, J.; Du, H.; Guo, X.; Liu, X.; Li, C.; Huang, G.; Niu, K. Effect of beta-hydroxyl-beta-methylbutyrate supplementation on muscle loss in older adults: A systematic review and meta-analysis. Arch. Gerontol. Geriatr. 2015, 61, 168–175. [Google Scholar] [CrossRef]
- Zhu, Y.Q.; Peng, N.; Zhou, M.; Liu, P.P.; Qi, X.L.; Wang, N.; Wang, G.; Wu, Z.P. Tai Chi and whole-body vibrating therapy in sarcopenic men in advanced old age: A clinical randomized controlled trial. Eur. J. Ageing 2019, 16, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cui, C.; Chim, Y.N.; Yao, H.; Shi, L.; Xu, J.; Wang, J.; Wang, R.M.Y.; Leung, K.S.; Chow, S.K.H.; et al. Vibration and beta-hydroxy-beta-methylbutyrate treatment suppresses intramuscular fat infiltration and adipogenic differentiation in sarcopenic mice. J. Cachexia Sarcopenia Muscle 2020, 11, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Nakae, Y.; Hirasaka, K.; Goto, J.; Nikawa, T.; Shono, M.; Yoshida, M.; Stoward, P.J. Subcutaneous injection, from birth, of epigallocatechin-3-gallate, a component of green tea, limits the onset of muscular dystrophy in mdx mice: A quantitative histological, immunohistochemical and electrophysiological study. Histochem. Cell Biol. 2008, 129, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Senthil Kumaran, V.; Arulmathi, K.; Srividhya, R.; Kalaiselvi, P. Repletion of antioxidant status by EGCG and retardation of oxidative damage induced macromolecular anomalies in aged rats. Exp. Gerontol. 2008, 43, 176–183. [Google Scholar] [CrossRef]
- Si, H.; Wang, X.; Zhang, L.; Parnell, L.D.; Admed, B.; LeRoith, T.; Ansah, T.A.; Zhang, L.; Li, J.; Ordovás, J.M.; et al. Dietary epicatechin improves survival and delays skeletal muscle degeneration in aged mice. FASEB J. 2019, 33, 965–977. [Google Scholar] [CrossRef]
- Mafi, F.; Biglari, S.; Ghardashi Afousi, A.; Gaeini, A.A. Improvement in skeletal muscle strength and plasma levels of follistatin and myostatin induced by an 8-week resistance training and epicatechin supplementation in sarcopenic older adults. J. Aging Phys. Act. 2019, 27, 384–391. [Google Scholar] [CrossRef]
- Liu, H.W.; Wei, C.C.; Chen, Y.J.; Chen, Y.A.; Chang, S.J. Flavanol-rich lychee fruit extract alleviates diet-induced insulin resistance via suppressing mTOR/SREBP-1 mediated lipogenesis in liver and restoring insulin signaling in skeletal muscle. Mol. Nutr. Food Res. 2016, 60, 2288–2296. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chen, Y.T.; Liu, H.W.; Chan, Y.C.; Liu, M.Y.; Hu, S.H.; Tseng, W.T.; Wu, H.L.; Wang, M.F.; Chang, S.J. Oligonol alleviates sarcopenia by regulation of signaling pathways involved in protein turnover and mitochondrial quality. Mol. Nutr. Food Res. 2019, 63, e1801102. [Google Scholar] [CrossRef]
- Beekmann, K.; de Haan, L.H.; Actis-Goretta, L.; Houtman, R.; van Bladeren, P.J.; Rietjens, I.M. The effect of glucuronidation on isoflavone induced estrogen receptor (ER) α and ERβ mediated coregulator interactions. J. Steroid Biochem. Mol. Biol. 2015, 154, 245–253. [Google Scholar] [CrossRef]
- Kurrat, A.; Blei, T.; Kluxen, F.M.; Mueller, D.R.; Piechotta, M.; Soukup, S.T.; Kulling, S.E.; Diel, P. Lifelong exposure to dietary isoflavones reduces risk of obesity in ovariectomized Wistar rats. Mol. Nutr. Food Res. 2015, 59, 2407–2418. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Kohno, S.; Yama, T.; Ochi, A.; Suto, T.; Hirasaka, K.; Ohno, A.; Teshima-Kondo, S.; Okumira, Y.; Oarada, M.; et al. Soy glycinin contains a functional inhibitory sequence against muscle-atrophy-associated ubiquitin ligase Cbl-b. Int. J. Endocrinol. 2013, 2013, 907565. [Google Scholar] [CrossRef] [PubMed]
- Tabata, S.; Aizawa, M.; Kinoshita, M.; Ito, Y.; Kawamura, Y.; Takebe, M.; Pan, W.; Sakuma, K. The influence of isoflavone for denervation-induced muscle atrophy. Eur. J. Nutr. 2018, 58, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Mukai, R.; Nakao, R.; Yamamoto, H.; Nikawa, T.; Takeda, E.; Terao, J. Quercetin prevents unloading-derived disused muscle atrophy by attenuating the induction of ubiquitin ligases in tail-suspension mice. J. Nat. Prod. 2010, 73, 1708–1710. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, S.D.; Suneja, M.; Ebert, S.M.; Bongers, K.S.; Fox, D.K.; Malberg, S.E.; Alipour, F.; Shields, R.K.; Adams, C.M. mRNA expression signatures of human skeletal muscle atrophy identify a natural compound that increases muscle mass. Cell Metab. 2011, 13, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Chen, J.A.; Xu, J.; Cao, J.; Wang, Y.; Thomas, S.S.; Hu, Z. Suppression of muscle wasting by the plant-derived compound ursolic acid in a model of chronic kidney disease. J. Cachexia Sarcopenia Muscle 2017, 8, 327–341. [Google Scholar] [CrossRef]
- Zolfaghari, M.; Faramarzi, M.; Hedayati, M.; Ghaffari, M. The effect of resistance and endurance training with ursolic acid on atrophy-related biomarkers in muscle tissue of diabetic male rats induced by streptozotocin and a high-fat diet. J. Food Biochem. 2022, 46, e14202. [Google Scholar] [CrossRef]
- Cho, Y.H.; Lee, S.Y.; Kim, C.M.; Kim, N.D.; Choe, S.; Lee, C.H.; Shin, J.H. Effect of loquat leaf extract on muscle strength, muscle mass, and muscle function in healthy adults: A randomized, double-blinded, and placebo-controlled trial. Evid.-Based Complement. Altern. Med. 2016, 2016, 4301621. [Google Scholar] [CrossRef]
- Bang, H.S.; Seo, D.Y.; Chung, Y.M.; Oh, K.M.; Park, J.J.; Arturo, F.; Jeong, S.H.; Kim, N.; Han, J. Ursolic Acid-induced elevation of serum irisin augments muscle strength during resistance training in men. Korean J. Physiol. Pharmacol. 2014, 18, 441–446. [Google Scholar] [CrossRef]
- Dupont, J.; Dedeyne, L.; Dalle, S.; Koppo, K.; Gielen, E. The role of omega-3 in the prevention and treatment of sarcopenia. Aging Clin. Exp. Res. 2019, 31, 825–836. [Google Scholar] [CrossRef]
- Otsuka, Y.; Iidaka, T.; Horii, C.; Muraki, S.; Oka, H.; Nakamura, K.; Izumo, T.; Rogi, T.; Shibata, H.; Tanaka, S.; et al. Dietary intake of vitamin E and fats associated with sarcopenia in community-dwelling older Japanese people: A cross-sectional study from the fifty survey of the ROAD study. Nutrients 2021, 13, 1730. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.I.; Julliand, S.; Reeds, D.N.; Sinacore, D.R.; Klein, S.; Mittendorfer, B. Fish oil-derived n-3 PUFA therapy increases muscle mass and function in healthy older adults. Am. J. Clin. Nutr. 2015, 102, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.M.; Jameson, K.A.; Batelaan, S.F.; Martin, H.J.; Syddall, H.E.; Dennison, E.M.; Cooper, C.; Sayer, A.A. Hertfordshire Cohort Study Group. Diet and its relationship with grip strength in community-dwelling older men and women: The Hertfordshire cohort study. J. Am. Geriatr. Soc. 2008, 56, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.; Sullenbarger, B.; Prakash, R.; McDaniel, J.C. Supplementation with eicosapentaenoic acid and docosahexaenoic acid reduces high levels of circulating proinflammatory cytokines in aging adults: A randomized, controlled study. Prostaglandins Leukot. Essent. Fat. Acids 2018, 132, 23–29. [Google Scholar] [CrossRef]
- Smith, G.I.; Atherton, P.; Reeds, D.N.; Mohammed, B.S.; Rankin, D.; Rennie, M.J.; Mittendorfer, B. Dietary omega-3 fatty acid supplementation increases the rate of muscle protein synthesis in older adults: A randomized controlled trial. Am. J. Clin. Nutr. 2011, 93, 402–412. [Google Scholar] [CrossRef]
- Annweiler, C.; Schott, A.M.; Berrut, G.; Fantino, B.; Beauchet, O. Vitamin D-related changes in physical performance: A systemic review. J. Nutr. Health Aging 2009, 13, 893–898. [Google Scholar] [CrossRef]
- Wicherts, I.S.; can Schoor, N.M.; Boeke, A.J.; Visser, M.; Deeg, D.J.; Smit, J.; Knol, D.L.; Lips, P. Vitamin D status predicts physical performance and its decline in older persons. J. Clin. Endocrinol. Metab. 2007, 92, 2058–2065. [Google Scholar] [CrossRef]
- Prokopidis, K.; Giannos, P.; Triantafyllidis, K.K.; Kechagias, K.; Mesinovic, J.; Witard, O.C.; Scott, D. Effect of vitamin D monotherapy on indices of sarcopenia in community-dwelling older adults: A systematic review and meta-analysis. J. Cachexia Sarcopnie Muscle 2022, 13, 1642–1652. [Google Scholar] [CrossRef]
- Lee, S.J. Regulation of muscle mass by myostatin. Annu. Rev. Cell Dev. Biol. 2004, 20, 61–86. [Google Scholar] [CrossRef]
- Wang, D.T.; Yang, Y.J.; Huang, R.H.; Zhang, Z.H.; Lin, X. Myostatin activates the ubiquitin-proteasome and autophagy-lysosome systems contributing to muscle wasting in chronic kidney disease. Oxidative Med. Cell. Longev. 2015, 2015, 684965. [Google Scholar] [CrossRef]
- Zhang, L.; Rajan, V.; Lin, E.; Hu, Z.; Han, H.Q.; Zhou, X.; Song, Y.; Min, H.; Wang, X.; Du, J.; et al. Pharmacological inhibition of myostatin suppresses systemic inflammation and muscle atrophy in mice with chronic kidney disease. FASEB J. 2011, 25, 1653–1663. [Google Scholar] [CrossRef] [PubMed]
- Carlson, M.E.; Hsu, M.; Conboy, I.M. Imbalance between pSmad3 and Notch induces CDK inhibitors in old muscle stem cells. Nature 2008, 454, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.T.; Chee, A.; Gleeson, B.G.; Naim, T.; Swiderski, K.; Koopman, R.; Lynch, G.S. Antibody-directed myostatin inhibition enhances muscle mass and function in tumor-bearing mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R716–R726. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.; McMillan, H.J.; Mah, J.K.; Tarnopolsky, N.; Selby, K.; McClure, T.; Wilson, D.M.; Sherman, M.L.; Escolar, D.; Attie, K.M. Myostatin inhibitor ACE-031 treatment of ambulatory boys with Duchenne muscular dystrophy: Results of a randomized, placebo-controlled clinical trial. Muscle Nerve 2017, 55, 458–464. [Google Scholar] [CrossRef]
- Woodhouse, L.; Gandhi, R.; Warden, S.J.; Poiraudeau, S.; Myers, S.L.; Benson, C.T.; Hu, L.; Ahmad, Q.I.; Linnemeier, P.; Gomez, E.V.; et al. A phase 2 randomized study investigating the efficacy and safety of myostatin antibody LY2495655 versus placebo in patients undergoing elective total hip arthroplasty. J. Frailty Aging 2016, 5, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Lord, S.R.; Studenski, S.A.; Warden, S.J.; Fielding, R.A.; Recknor, C.P.; Hochberg, M.C.; Ferrari, S.L.; Blain, H.; Binder, E.F.; et al. Myostatin antibody (LY2495655) in older weak fallers: A proof-of-concept, randomised, phase 2 trial. Lancet Diabetes Endocrinol. 2015, 3, 948–957. [Google Scholar] [CrossRef]
- Lee, S.J. Targeting the myostatin signaling pathway to treat muscle loss and metabolic dysfunction. J. Clin. Investig. 2021, 131, e148372. [Google Scholar] [CrossRef]
- Loffredo, F.S.; Steinhauser, M.L.; Jay, S.M.; Gannon, J.; Pancoast, J.R.; Yalamanchi, P.; Sinha, M.; Dall’Osso, C.; Khong, D.; Shadrach, T.; et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell 2013, 153, 828–839. [Google Scholar] [CrossRef]
- Sinha, M.; Jang, Y.C.; Oh, J.; Khong, D.; Wu, E.Y.; Manohar, R.; Miller, C.; Regalado, S.G.; Loffredo, F.S.; Pancoast, J.R.; et al. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle. Science 2014, 344, 649–652. [Google Scholar] [CrossRef]
- Schafer, M.J.; Atkinson, E.J.; Vanderboom, P.M.; Kotajarvi, B.; White, T.A.; Moore, M.M.; Bruce, C.J.; Greason, K.L.; Suri, R.M.; Khosla, S.; et al. Quantification of GDF11 and myostatin in human aging and cardiovascular disease. Cell Metab. 2016, 23, 1207–1215. [Google Scholar] [CrossRef]
- Feldman, H.A.; Longcope, C.; Derby, C.A.; Johannes, C.B.; Araujo, A.B.; Coviello, A.D.; Bremner, W.J.; McKinlay, J.B. Age trends in the level of serum testosterone and other hormones in middle-aged men: Longituidinal results from the Massachusetts male aging study. J. Clin. Endocrinol. Metab. 2002, 87, 589–598. [Google Scholar] [CrossRef]
- Lehtihet, M.; Hylander, B. Semen quality in men with chronic kidney disease and its correlation with chronic kidney disease stages. Andrologia 2015, 47, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Bhasin, S.; Calof, O.; Storer, T.W.; Lee, M.L.; Mazer, N.A.; Jasuja, R.; Montori, V.M.; Gao, W.; Dalton, J.T. Drug insight: Testosterone and selective androgen receptor modulators as anabolic therapies for physical dysfunction in chronic illness and ageing. Nat. Clin. Pract. Endocrinol. Metab. 2006, 2, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Bakhshi, V.; Elliott, M.; Gentili, A.; Godschalk, M.; Mulligan, T. Testosterone improves rehabilitation outcomes in ill older men. J. Am. Geriatr. Soc. 2000, 48, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, A.A.; Sheffield-Moore, M.; Yeckel, C.W.; Gilkison, C.; Jiang, J.; Achacosa, A.; Lieberman, S.A.; Tipton, K.; Wolfe, R.R.; Urban, R.J. Testosterone administration to older men improves muscle function: Molecular and physiological mechanisms. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E601–E607. [Google Scholar] [CrossRef]
- Sinha-Hikim, I.; Cornford, M.; Gaytan, H.; Lee, M.L.; Bhasin, S. Effects of testosterone supplementation on skeletal muscle fiber hypertrophy and satellite cells in community-dwelling older men. J. Clin. Endocrinol. Metab. 2006, 91, 3024–3033. [Google Scholar] [CrossRef]
- Crawford, J.; Prado, C.M.M.; Ann Johnston, M.; Gralla, R.J.; Taylor, R.P.; Hancock, M.L.; Dalton, J.T. Study design and rationale for the phase 3 clinical development program of enobosarm, a selective androgen receptor modulator, for the prevention and treatment of muscle wasting in cancer patients (POWER Trials). Curr. Oncol. Rep. 2016, 18, 37. [Google Scholar] [CrossRef]
- Dalton, J.T.; Barnette, K.G.; Bohl, C.E.; Hancock, M.L.; Rodriguez, D.; Dodson, S.T.; Morton, R.A.; Steiner, M.S. The selective androgen receptor modulator GTx-024 (enobosarm) improves lean body mass and physical function in healthy elderly men and postmenopausal women: Results of a double-blind, placebo-controlled phase II trial. J. Cachexia Sarcopenia Muscle 2011, 2, 153–161. [Google Scholar] [CrossRef]
- Neil, D.; Clark, R.V.; Magee, M.; Billiard, J.; Chan, A.; Xue, Z.; Russell, A. GSK2881078, a SARM, produces dose-dependent increases in lean mass in healthy older men and women. J. Clin. Endocrinol. Metab. 2018, 103, 3215–3224. [Google Scholar] [CrossRef]
- Supasyndh, O.; Satirapoj, B.; Aramwit, P.; Viroonudomphol, D.; Chaiprasert, A.; Thanachatwej, V.; Vanichakarn, S.; Kopple, J.D. Effect of oral anabolic steroid on muscle strength and muscle growth in hemodialysis patients. Clin. J. Am. Soc. Nephrol. 2013, 8, 271–279. [Google Scholar] [CrossRef]
- Kim, K.Y.; Ku, S.K.; Lee, K.W.; Song, C.H.; An, W.G. Muscle-protective effects of Schisandrae Fructus extracts in old mice after chronic forced exercise. J. Ethnopharmacol. 2018, 212, 175–187. [Google Scholar] [CrossRef]
- Kristensen, J.M.; Treebak, J.T.; Schjerling, P.; Goodyear, L.; Wojtaszewski, J.F. Two weeks of metformin treatment induces AMPK-dependent enhancement of insulin-stimulated glucose uptake in mouse soleus muscle. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1099–E1109. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K. AMP-activated protein kinase (AMPK) controls the aging process via an integrated signaling network. Ageing Res. Rev. 2012, 11, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Suwa, M.; Egashira, T.; Nakano, H.; Sasaki, H.; Kumagai, S. Metformin increases the PGC-1alpha protein and oxidative enzyme activities possibly via AMPK phosphorylation in skeletal muscle in vivo. J. Appl. Physiol. 2006, 101, 1685–1692. [Google Scholar] [CrossRef]
- Kulkarni, A.S.; Brutsaert, E.F.; Anghel, V.; Zhang, K.; Bloomgarden, N.; Pollak, M.; Mar, J.C.; Hawkins, M.; Crandall, J.P.; Barzilai, N. Metformin regulates metabolic and nonmetabolic pathways in skeletal muscle and subcutaneous adipose tissues of older adults. Aging Cell 2018, 17, e12723. [Google Scholar] [CrossRef]
- Laksmi, P.W.; Setiati, S.; Tamin, T.Z.; Soewondo, P.; Rochmah, W.; Nafrialdi, N.; Prihartono, J. Effect of metformin on handgrip strength, gait speed, myostatin serum level, and health-related quality of life: A double blind randomized controlled trial among non-diabetic pre-frail elderly patients. Acta Medica Indones. 2017, 49, 118–127. [Google Scholar]
- Rabøl, R.; Boushel, R.; Almdal, T.; Hansen, C.N.; Ploug, T.; Haugaard, S.B.; Prats, C.; Madsbad, S.; Dela, F. Opposite effects of pioglitazone and rosiglitazone on mitochondrial respiration in skeletal muscle of patients with type 2 diabetes. Diabetes Obes. Metab. 2010, 12, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Coletta, D.K.; Sriwijitkamol, A.; Wajcberg, E.; Tantiwong, P.; Li, M.; Prentki, M.; Madiraju, M.; Jenkinson, C.P.; Cersosimo, E.; Musi, N.; et al. Pioglitazone stimulates AMP-activated protein kinase signalling and increases the expression of genes involved in adiponectin signalling, mitochondrial function and fat oxidation in human skeletal muscle in vivo: A randomised trial. Diabetologia 2009, 52, 723–732. [Google Scholar] [CrossRef]
- Shea, M.K.; Nicklas, B.J.; Marsh, A.P.; Houston, D.K.; Miller, G.D.; Isom, S.; Miller, M.E.; Carr, J.J.; Lyles, M.F.; Harris, T.B.; et al. The effect of pioglitazone and resistance training on body composition in older men and women undergoing hypocaloric weight loss. Obesity 2011, 19, 1636–1646. [Google Scholar] [CrossRef]
- Mele, A.; Calzolaro, S.; Cannone, G.; Cetrone, M.; Conte, D.; Tricarico, D. Database search of spontaneous reports and pharmacological investigations on the sulfonylureas and glinides-induced atrophy in skeletal muscle. Pharmacol. Res. Perspect. 2014, 2, e00028. [Google Scholar] [CrossRef]
- Rizzo, M.R.; Barbieri, M.; Fava, I.; Desiderio, M.; Coppola, C.; Marfella, R.; Paolisso, G. Sarcopenia in elderly diabetic patients: Role of dipeptidyl peptidase 4 inhibitors. J. Am. Med. Dir. Assoc. 2016, 17, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Nagai, Y.; Kato, H.; Fukuda, H.; Tanaka, Y. Effect of the dipeptidyl peptidase-4 inhibitor sitagliptin on muscle mass and the muscle/fat ratio in patients with type 2 diabetes. J. Clin. Med. Res. 2020, 12, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Giannocco, G.; Oliveira, K.C.; Crajoinas, R.O.; Venturini, G.; Salles, T.A.; Fonseca-Alaniz, M.H.; Maciel, R.M.; Girardi, A.C. Dipeptidyl peptidase IV inhibition upregulates GLUT4 translocation and expression in heart and skeletal muscle of spontaneously hypertensive rats. Eur. J. Pharmacol. 2013, 698, 74–86. [Google Scholar] [CrossRef]
- Sato, H.; Kubota, N.; Kubota, T.; Takamoto, I.; Iwayama, K.; Tokuyama, K.; Moroi, M.; Sugi, K.; Nakaya, K.; Goto, M.; et al. Anagliptin increases insulin-induced skeletal muscle glucose uptake via an NO-dependent mechanism in mice. Diabetologia 2016, 59, 2426–2434. [Google Scholar] [CrossRef] [PubMed]
- Takada, S.; Masaki, Y.; Kinugawa, S.; Matsumoto, J.; Furihata, T.; Mizushima, W.; Kadoguchi, T.; Fukushima, A.; Homma, T.; Takahashi, M.; et al. Dipeptidyl peptidase-4 inhibitor improved exercise capacity and mitochondrial biogenesis in mice with heart failure via activation of glucagon-like peptide-1 receptor signalling. Cardiovasc. Res. 2016, 111, 338–347. [Google Scholar] [CrossRef]
- Green, C.J.; Henriksen, T.I.; Pedersen, B.K.; Solomon, T.P. Glucagon like peptide-1-induced glucose metabolism in differentiated human muscle satellite cells is attenuated by hyperglycemia. PLoS ONE 2012, 7, e44284. [Google Scholar] [CrossRef] [PubMed]
- Choung, J.S.; Lee, Y.S.; Jun, H.S. Exendin-4 increases oxygen consumption and thermogenic gene expression in muscle cells. J. Mol. Endocrinol. 2017, 58, 79–90. [Google Scholar] [CrossRef]
- Yajima, T.; Yajima, K.; Takahashi, H.; Yasuda, K. The effect of dulaglutide on body composition in type 2 diabetes mellitus patients on hemodialysis. J. Diabetes Complicat. 2018, 32, 759–763. [Google Scholar] [CrossRef]
- Perna, S.; Guido, D.; Bologna, C.; Solerte, S.B.; Guerriero, F.; Isu, A.; Rondanelli, M. Liraglutide and obesity in elderly: Efficacy in fat loss and safety in order to prevent sarcopenia. A perspective case series study. Aging Clin. Exp. Res. 2016, 28, 1251–1257. [Google Scholar] [CrossRef]
- Merovci, A.; Solis-Herrera, C.; Daniele, G.; Eldor, R.; Fiorentino, T.V.; Tripathy, D.; Xiong, J.; Perez, Z.; Norton, L.; Abdul-Ghani, M.A.; et al. Dapagliflozin improves muscle insulin sensitivity but enhances endogenous glucose production. J. Clin. Investig. 2014, 124, 509–514. [Google Scholar] [CrossRef]
- Esterline, R.L.; Vaag, A.; Oscarsson, J.; Vora, J. Mechanisms in endocrinology: SGLT2 inhibitors: Clinical benefits by restoration of normal diurnal metabolism? Eur. J. Endocrinol. 2018, 178, R113–R125. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T. Sarcopenia, frailty circle and treatment with sodium-glucose cotransporter 2 inhibitors. J. Diabetes Investig. 2019, 10, 193–195. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Sakai, K.; Saito, K.; Tsutsui, K.; Yamashita, S.; Kato, N. Efficacy and safety of ipragliflozin in Japanese patients with type 2 diabetes receiving conventional therapy: Clinical implication of the importance of exercise habits during treatment with ipragliflozin. Diabetol. Int. 2017, 8, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Matsuba, R.; Matsuba, I.; Shimokawa, M.; Nagai, Y.; Tanaka, Y. Tofogliflozin decreases body fat mass and improves peripheral insulin resistance. Diabetes Obes. Metab. 2018, 20, 1311–1315. [Google Scholar] [CrossRef]
- Sano, M.; Meguro, S.; Kawai, T.; Suzuki, Y. Increased grip strength with sodium-glucose cotransporter 2. J. Diabetes 2016, 8, 736–737. [Google Scholar] [CrossRef]
- Abdulla, H.; Smith, K.; Atherton, P.J.; Idris, I. Role of insulin in the regulation of human skeletal muscle protein synthesis and breakdown: A systematic review and meta-analysis. Diabetologia 2016, 59, 44–55. [Google Scholar] [CrossRef]
- Bouchi, R.; Fukuda, T.; Takeuchi, T.; Nakano, Y.; Murakami, M.; Minami, I.; Izumiyama, H.; Hashimoto, K.; Yoshimoto, T.; Ogawa, Y. Insulin treatment attenuates decline of muscle mass in Japanese patients with type 2 diabetes. Calcif. Tissue Int. 2017, 101, 1–8. [Google Scholar] [CrossRef]
- Ferrari, U.; Then, C.; Rottenkolber, M.; Selte, C.; Seissler, J.; Conzade, R.; Linkohr, B.; Peters, A.; Drey, M.; Thorand, B. Longitudinal association of type 2 diabetes and insulin therapy with muscle parameters in the KORA-Age study. Acta Diabetol. 2020, 57, 1057–1063. [Google Scholar] [CrossRef]
- Sasaki, T.; Sugawara, M.; Fykuda, M. Sodium–glucose cotransporter 2 inhibitor induced changes in body composition and simultaneous changes in metabolic profile: 52-week prospective LIGHT (Luseogliflozin: The Components of Weight Loss in Japanese Patients with Type 2 Diabetes Mellitus) study. J. Diabetes Investig. 2019, 10, 108–117. [Google Scholar] [CrossRef]
- Madushika Abeywickrama, H.; Uchiyama, M.; Sumiyoshi, T.; Okuda, A.; Koyama, Y. The role of zinc on nutritional status, sarcopenia, and frailty in older adults: A scoping review. Nutr. Rev. 2023; in press. [Google Scholar] [CrossRef]
Figure 1.
Nutritional interventions affect different mediators in sarcopenic muscle. Recent findings suggest that the myostatin-Smad pathway inhibits protein synthesis probably due to blocking the functional role of Akt. Treatment with an ursolic acid upregulates the amount of IGF-I and then stimulates protein synthesis by activating the Akt/mTORC1/p70S6K pathway. Administration of omega-3 PUFA or HMB may work to prevent sarcopenia by activating mTORC1. Sarcopenic muscle exhibits a marked defect of autophagy-dependent signaling, which is effectively ameliorated by oligonol supplementation. ALK, activin receptor-like kinase; ActRIIB, activin receptor IIB; IGF-I, insulin-like growth factor I; TSC, tuberous sclerosis complex; TORC1, component of TOR signaling complex 1; Rheb, Ras homolog enriched in brain; mTORC1, mammalian target of rapamycin complex 1; eIF4E, eukaryotic initiation factor 4E; FOXO, forkhead box O; LC3, microtubule-associated protein light chain 3; BNIP, BCL2/adenovirus E1B 19 kd-interacting protein; Atg, autophagy-related genes.
Figure 1.
Nutritional interventions affect different mediators in sarcopenic muscle. Recent findings suggest that the myostatin-Smad pathway inhibits protein synthesis probably due to blocking the functional role of Akt. Treatment with an ursolic acid upregulates the amount of IGF-I and then stimulates protein synthesis by activating the Akt/mTORC1/p70S6K pathway. Administration of omega-3 PUFA or HMB may work to prevent sarcopenia by activating mTORC1. Sarcopenic muscle exhibits a marked defect of autophagy-dependent signaling, which is effectively ameliorated by oligonol supplementation. ALK, activin receptor-like kinase; ActRIIB, activin receptor IIB; IGF-I, insulin-like growth factor I; TSC, tuberous sclerosis complex; TORC1, component of TOR signaling complex 1; Rheb, Ras homolog enriched in brain; mTORC1, mammalian target of rapamycin complex 1; eIF4E, eukaryotic initiation factor 4E; FOXO, forkhead box O; LC3, microtubule-associated protein light chain 3; BNIP, BCL2/adenovirus E1B 19 kd-interacting protein; Atg, autophagy-related genes.

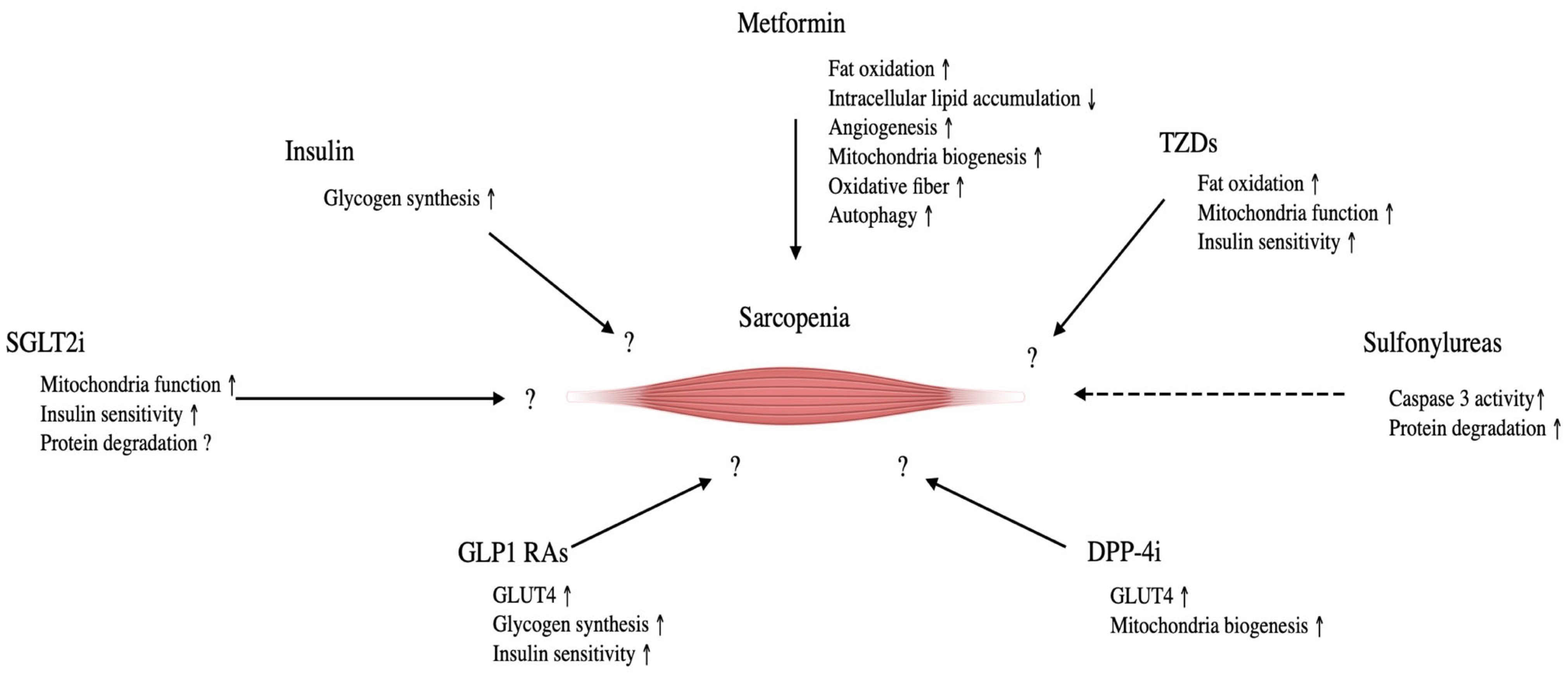
Figure 2.
The effect of glucose lowering drugs on sarcopenia and its molecular mechanisms. Tzds, thiazolidinediones; DPP4i, Dipeptidyl pepridaze-4 inhibitors; GLP-1 Ras, Glucogon-like peptide-1 receptor agonist; SLGT2i, Sodium-glucose transporter protein 2 inhibitors. Straight lines indicate a positive effect on sarcopenia; dotted lines indicate negative effects.
Figure 2.
The effect of glucose lowering drugs on sarcopenia and its molecular mechanisms. Tzds, thiazolidinediones; DPP4i, Dipeptidyl pepridaze-4 inhibitors; GLP-1 Ras, Glucogon-like peptide-1 receptor agonist; SLGT2i, Sodium-glucose transporter protein 2 inhibitors. Straight lines indicate a positive effect on sarcopenia; dotted lines indicate negative effects.

{kind=link}
{kind=link}
Table 1.
Summary of nutritional approaches for attenuating muscle atrophy.
| Title 1 | Duration, Intervention Pattern | Species | Outcome | Authors |
|---|---|---|---|---|
| HMB | 3 g, 10 days | Human, unloading patients | Muscle strength ↑ | Deutz et al. [19] |
| 2 to 4 weeks | Human | Anthropometric parameters → | Hsieh et al. [20] | |
| RCT, HMB | Human, elderly | LBM ↑, fat mass → Muscle strength → | Flakoll et al. [21] | |
| HMB | RCT, HMB + Arg + Lys (2/5/1.5 g per day) | Human, elderly | LBM ↑, fat mass → Whole body protein synthesis↑ Leg and grip strength ↑ Limb circumference ↑ Muscle strength → | Flakoll et al. [21] |
| HMB + vibration | RCT, 12–16 Hz, 3–5 mm, 8 weeks | Human, sarcopenia | Muscle strength ↑, Physical performance (walking speed, 5-stance test, timed stand test) ↑ | Zhu et al. [23] |
| LMHFV | Ageing mice (SAMP8) | Myostatin expression ↓ Intramuscular fat mass ↓ | Wang et al. [24] | |
| Catechin | EGCG, 5 mg/kg, 4 times/week, 8 weeks | Muscular dystrophic mice | fibrosis ↓, necrotic fibers ↓ | Nakae et al. [25] |
| EGCG, 100 mg/kg, 30 days | Aged male rats | Oxidative stress marker ↓ | Senthil Kumaran et al. [26] | |
| EC or EGCG (0.25% in drinking water), 37 weeks | 20-month old male mice | Survival rate: EC ↑, EGCG → Muscle degeneration ↓: EC >> EGCG Physical activity ↑: EC >> EGCG | Si et al. [27] | |
| Oligonol, 200 mg/kg, 8 weeks | SAMP8 mice, 32-weeks old | PGC-1α and Mfn2 ↓ LC3-II, p62, and ATG13 ↑ Autophagosome number ↑ | Chang et al. [30] | |
| Catechin + resistance training | RCT, tea catechin (350 mL/day), 3 month | Human, sarcopenia, women | Leg muscle mass ↑ Normal walking speed ↑ | Mafi et al. [28] |
| 120 days | Male mice | Fat accumulation ↓ | Kurrat et al. [32] | |
| 20% of the diet, 4 days | Mouse, denervated | IRS-1 and p-Akt protein ↑ | Abe et al. [33] | |
| Isoflavones | 0.6% of the diet, 2 weeks daidzein/genistein/ glycitein (7:1:2) | Mouse, denervated | Apoptosis-dependent signaling ↓ | Tabata et al. [34] |
| Muscular injection (50 μL; 2.5 pmol of quercetin) | Hindlimb-unloaded mice | Skeletal muscle mass ↑ Atrogin-1 and MuRF1 ↓ | Mukai et al. [35] | |
| Ursolic acid | Orally treatment (100 mg/kg), 3 weeks | Mice models of CKD | Muscle mass ↑ Inflammatory cytokines (IL-6 and TNF-α) ↓ Ubiquitin E3 ligases (MuRF1, atrogin-1, MUSA1) ↓ | Yu et al. [37] |
| 50 mg/kg, loquat leaf extract intake | Human (healthy adults) | Muscle mass and muscle strength → | Cho et al. [39] | |
| Ursolic acid + endurance training | 500 mg/kg, 8 weeks | 21-month old male rats with diabetes by high fat diet | Body weight ↓ Insulin resistance ↓ Insulin and glucose concentration ↓ p53, ATF4, and p21 protein level ↓ | Zolfaghari et al. [38] |
| Ursolic acid + resistance training | 1 capsule (450 mg), 3 times/day, 8 weeks | Healthy male human (age: 29.4 ± 5.1 years) | Muscle strength ↑ Lean body mass → | Bang et al. [40] |
| 1.86 g EPA and 1.5 g DHA/day, 6 months | Human (60-85 year men and women, elderly) | Muscle protein synthesis ↑ | Smith et al. [43] | |
| Omega-3 PUFAs | RCT, EPA and DHA, 8 weeks | Middle-aged and older adults | TNF-α, IL-6, and IL-1β ↓ | Tan et al. [45] |
| RCT, 8 weeks | Healthy older men and women (>65 years) | Muscle protein synthesis → | Smith et al. [46] |
↑: increase, ↓: decrease, → no change.
Table 2.
Summary of pharmacological approach for attenuating muscle atrophy.
| Title 1 | Duration, Intervention Pattern | Species | Outcome | Authors |
|---|---|---|---|---|
| Vitamin D | Systematic review | Nursing home residents and older adults living in areas with low vitamin D | Risk of falls ↓ Muscle performance ↑ Muscle strength ↑ | Annweiler et al. [47] |
| Subcutaneous injection | CKD model mice | Body weight ↑ Muscle mass ↑ Inflammation marker ↓ | Zhang et al. [52] | |
| Myostatin inhibition | RCT, ACE-031, Cohort 1: 0.5 mg/kg every 4 weeks, Cohort 2: 1 mg/kg every 2 weeks | DMD patients | Muscle mass ↑ Performance (6 min walk test) ↑ Non-muscular side effects (nosebleeds, capillary dilation) ↑ | Campbell et al. [55] |
| RCT, LY2495655, 8 and 16 weeks | Patients undergoing elective total hip arthroplasty | LBM ↑ | Woodhouse et al. [56] | |
| Phase 2 trial, LY2495655, 24 weeks | Frail elderly (75 years and older) subjects worldwide (e.g., US, France, Australia) | LBM ↑ Functional characteristics (stair climbing time and chair standing with arms) ↑ | Becker et al. [57] | |
| GDF11 | IP injection, 0.1 mg/kg, 30 days | Aged (22–24 months) mice | Satellite cells ↑ Skeletal muscle regeneration ↑ Muscle physiological parameters (Run time and grip strength) ↑ PGC-1α ↑ | Sinha et al. [60] |
| Anabolic steroids | Testosterone, high dose | Community-dwelling older men, 6 months | LBM ↑ Leg and arm muscle strength ↑ | Sinha-Hikim et al. [67] |
| Double-blind study, SARMs (enobosarm) | Healthy postmenopausal elderly men | LBM ↑ Physical function (stair climbing speed) ↑ | Neil et al. [70] | |
| SARMs (GSK2881078), Once daily, 50 days | Healthy elderly men and women | LBM ↑ | Neil et al. [70] | |
| RCT, oxymetholone, 24-weeks | Hemodyalysis patients | LBM ↑ Physical function and grip strength ↑ CSA of Type I fibers ↑ | Supasyndh et al. [71] | |
| Oxymetholone, 50 mg/kg | 10-month-old mice | Myostatin, sirtuin1, Fiber size (soleus and gastrocnemius muscles) ↑ | Kim et al. [72] | |
| Metformin, 3 × 500 mg, 16 weeks | Elderly patients with diabetes | Walking speed ↑ | Laksmi et al. [77] | |
| Thiazolidinediones (pioglitazone) | Diabetic patients | Thigh muscle mass ↓ | Shea et al. [80] | |
| Sulfonylurea, 24 month | Elderly patients with diabetes | Skeletal muscle mass → Muscle strength → Gait speed → | Rizzo et al. [82] | |
| Glucose-lowering drugs | DPP-4i, 24 month | Elderly patients with diabetes | Skeletal muscle mass ↑ Muscle strength ↑ Gait speed ↑ | Rizzo et al. [82] |
| DPP-4i (tenelingliptin), 20 mg/day, 6 months | Diabetic patients undergoing hemodyalysis | Skeletal muscle mass → Fat mass → | Yajima et al. [89] | |
| GLP-1 RAs (dulaglutide), 0.75 mg/week, 6 months | Diabetic patients undergoing hemodyalysis | Skeletal muscle mass ↓ Fat mass ↓ | Yajima et al. [89] | |
| GLP-1 RAs (dulaglutide), 3 g/day, 6 months | Obese elderly patients with diabetes | Sarcopenic parameters → | Perna et al. [90] | |
| SGLT2i (depaglifrozin), 10 mg/day, 2 weeks | Diabetic patients | Insulin sensitivity ↑ Muscle catabolism ↓ | Merovci et al. [91] | |
| SGLT2i (luseoglifrozin), 2.5–5 mg/day, 1 year | Diabetic patients | Body weight ↓ Body mass index ↓ Waist circumference ↓ | Sasaki et al. [100] | |
| Insulin, 3 years | Diabetic patients | Skeletal muscle mass ↑ | Ferrari et al. [99] |
↑: increase, ↓: decrease, → no change.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sakuma, K.; Hamada, K.; Yamaguchi, A.; Aoi, W. Current Nutritional and Pharmacological Approaches for Attenuating Sarcopenia. Cells 2023, 12, 2422. https://doi.org/10.3390/cells12192422
AMA Style
Sakuma K, Hamada K, Yamaguchi A, Aoi W. Current Nutritional and Pharmacological Approaches for Attenuating Sarcopenia. Cells. 2023; 12(19):2422. https://doi.org/10.3390/cells12192422
Chicago/Turabian StyleSakuma, Kunihiro, Kento Hamada, Akihiko Yamaguchi, and Wataru Aoi. 2023. "Current Nutritional and Pharmacological Approaches for Attenuating Sarcopenia" Cells 12, no. 19: 2422. https://doi.org/10.3390/cells12192422
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.