
Bacillus subtilis Modulated the Expression of Osteogenic Markers in a Human Osteoblast Cell Line
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. HPLC Quantification of Vitamin K2 or Menaquinones (MK) from Probiotics
2.3. XTT Assay
2.4. Alizarin Red (AR) Staining
2.5. ALP Staining
2.6. WB
2.7. Immunofluorescence Staining
2.8. Image Analysis
2.9. RNA Extraction and Quantification
2.10. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.11. Statistical Analysis
3. Results
3.1. MKs Production
- ▪
- P1: 81 and 0.5 µg/g;
- ▪
- P2: 0.59 and 7 µg/g.
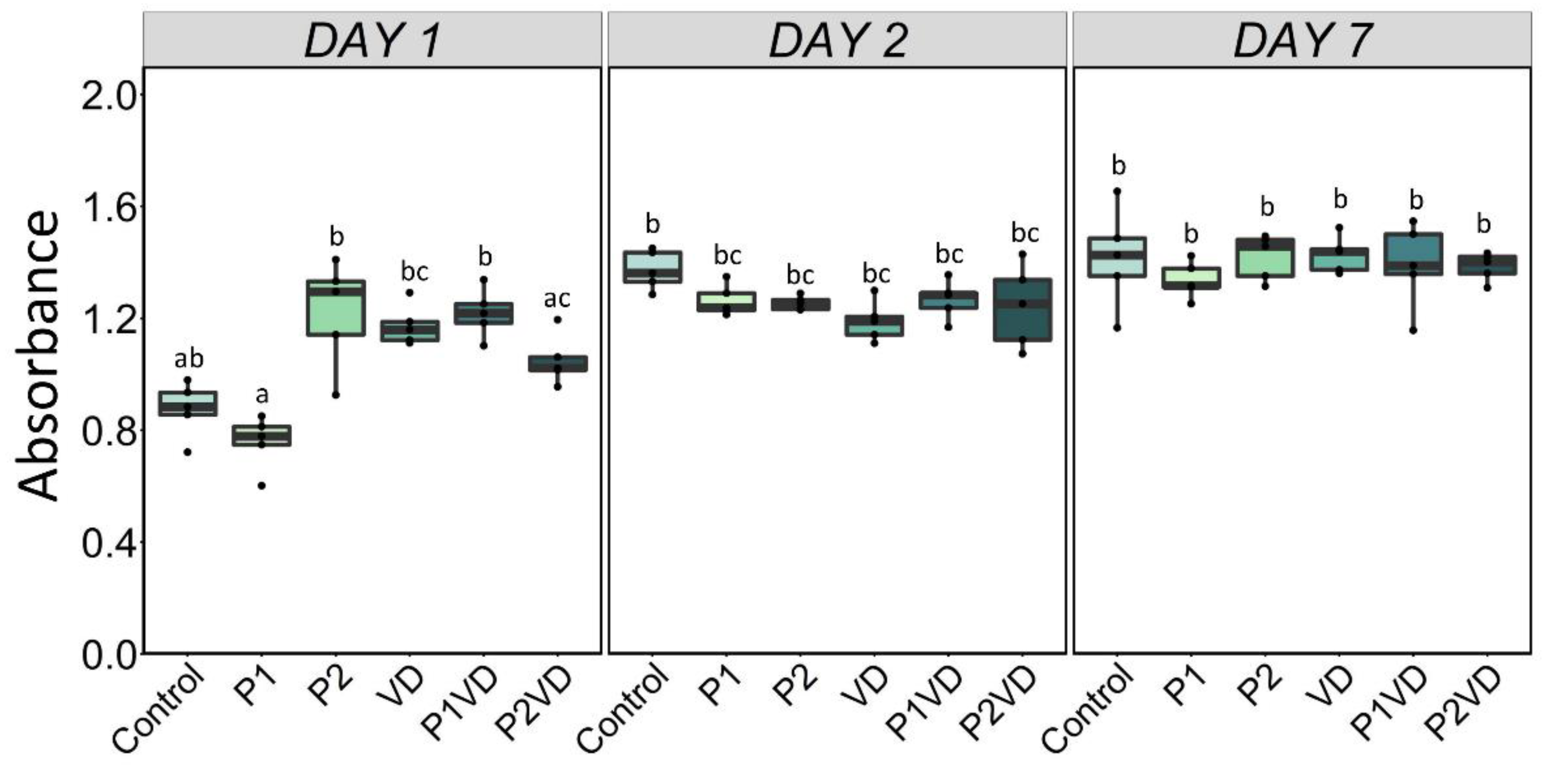
3.2. Cell Viability
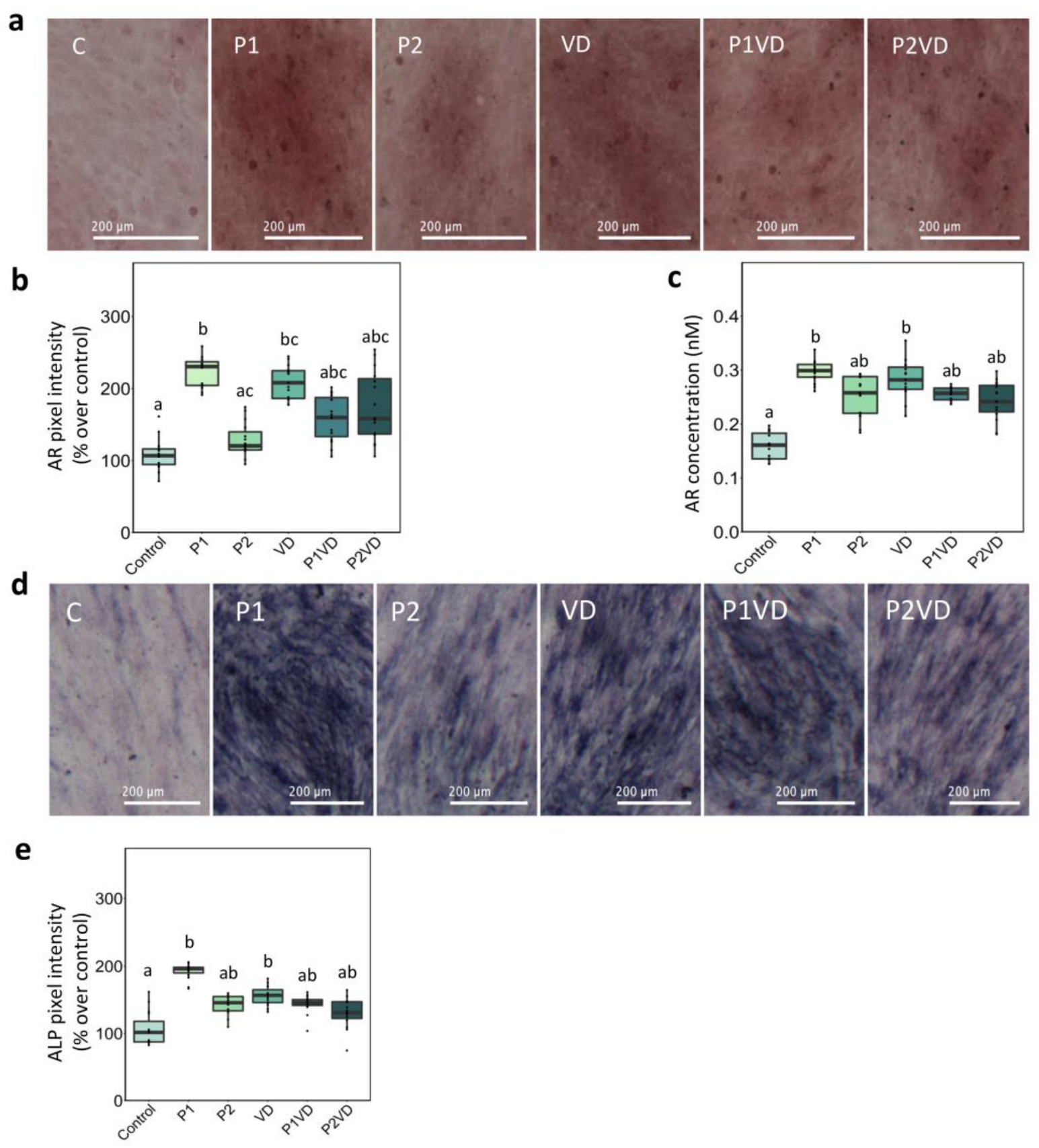
3.3. Calcium Nodules and ALP Production
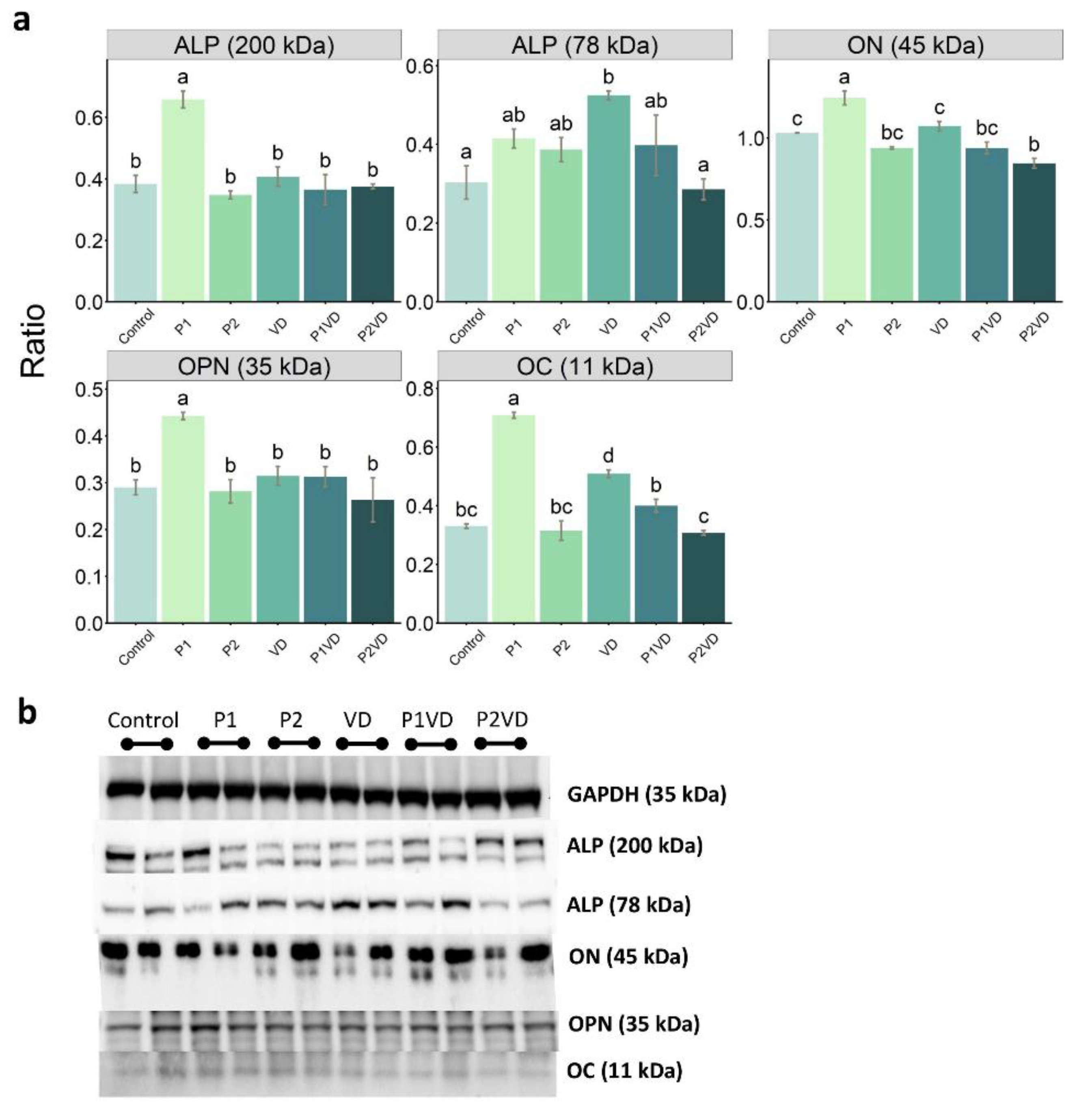
3.4. Protein Expression
3.5. ALP Immunofluorescence
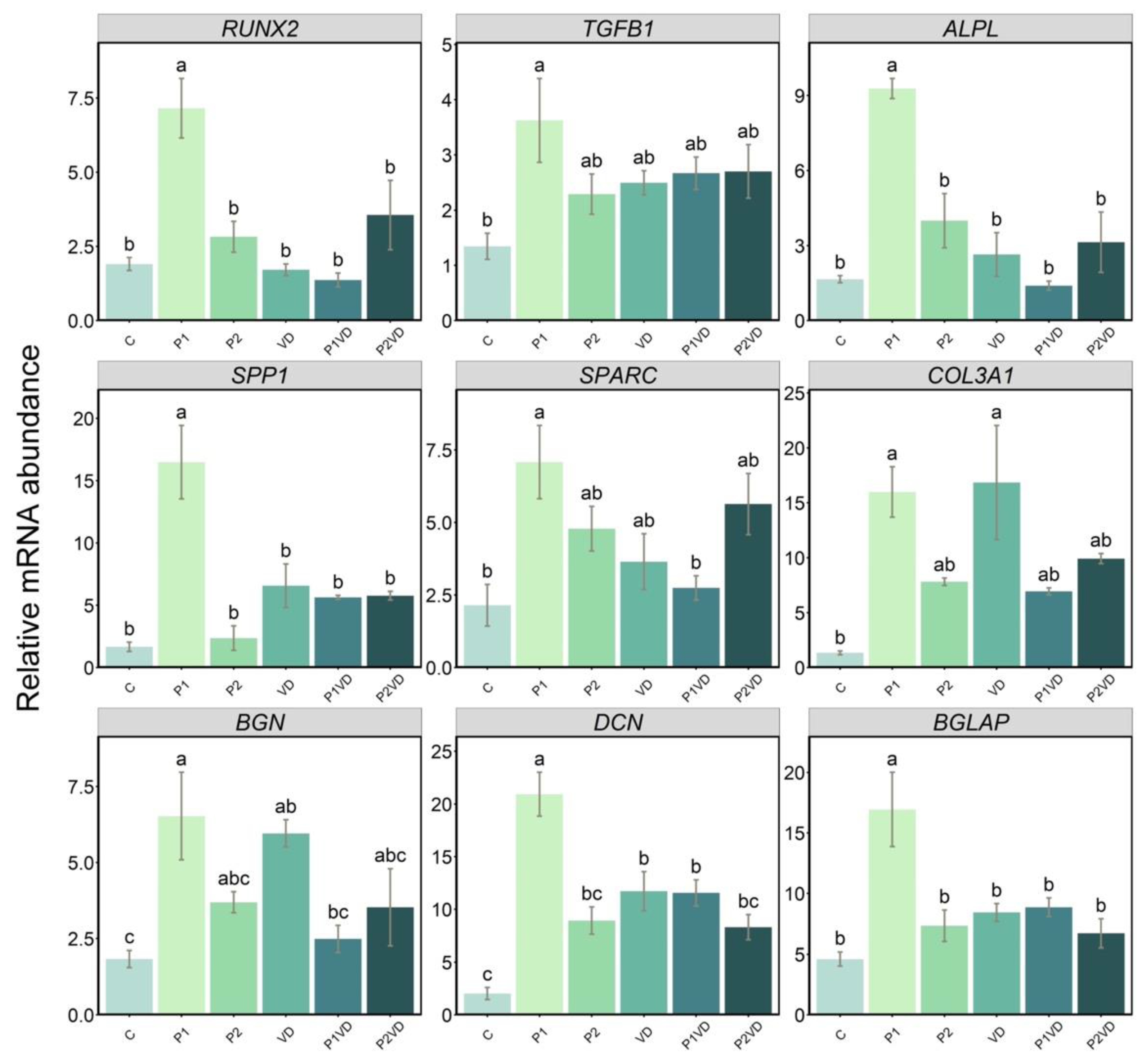
3.6. Gene Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jakab, L. Bone tissue: Rebuilding and inflammation. Orv. Hetil. 2014, 155, 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Kanis, J.A.; Norton, N.; Harvey, N.C.; Jacobson, T.; Johansson, H.; Lorentzon, M.; McCloskey, E.V.; Willers, C.; Borgström, F. SCOPE 2021: A New Scorecard for Osteoporosis in Europe. Arch. Osteoporos. 2021, 16, 82. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of Bone Development and Extracellular Matrix Protein Genes by RUNX2. Cell Tissue Res. 2009, 339, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruderer, M.; Richards, R.G.; Alini, M.; Stoddart, M.J. Role and Regulation of RUNX2 in Osteogenesis. Eur. Cells Mater. 2014, 28, 269–286. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.J.; Fernandes, M.H. Human Bone Cell Cultures in Biocompatibility Testing. Part II: Effect of Ascorbic Acid, β-Glycerophosphate and Dexamethasone on Osteoblastic Differentiation. Biomaterials 2000, 21, 1095–1102. [Google Scholar] [CrossRef]
- Stein, G.S.; Lian, J.B.; van Wijnen, A.J.; Stein, J.L.; Montecino, M.; Javed, A.; Zaidi, S.K.; Young, D.W.; Choi, J.-Y.; Pockwinse, S.M. Runx2 Control of Organization, Assembly and Activity of the Regulatory Machinery for Skeletal Gene Expression. Oncogene 2004, 23, 4315–4329. [Google Scholar] [CrossRef] [Green Version]
- Setzer, B.; Bächle, M.; Metzger, M.C.; Kohal, R.J. The Gene-Expression and Phenotypic Response of HFOB 1.19 Osteoblasts to Surface-Modified Titanium and Zirconia. Biomaterials 2009, 30, 979–990. [Google Scholar] [CrossRef]
- Aghaloo, T.; Cowan, C.M.; Chou, Y.-F.; Zhang, X.; Lee, H.; Miao, S.; Hong, N.; Kuroda, S.; Wu, B.; Ting, K.; et al. Nell-1-Induced Bone Regeneration in Calvarial Defects. Am. J. Clin. Pathol. 2006, 169, 903–915. [Google Scholar] [CrossRef] [Green Version]
- de C. Foureaux, R.; Messora, M.R.; de Oliveira, L.F.F.; Napimoga, M.H.; Pereira, A.N.J.; Ferreira, M.S.; Pereira, L.J. Effects of Probiotic Therapy on Metabolic and Inflammatory Parameters of Rats with Ligature-Induced Periodontitis Associated with Restraint Stress. J. Periodontol. 2014, 85, 975–983. [Google Scholar] [CrossRef]
- Nilsson, A.G.; Sundh, D.; Bäckhed, F.; Lorentzon, M. Lactobacillus reuteri Reduces Bone Loss in Older Women with Low Bone Mineral Density: A Randomized, Placebo-Controlled, Double-Blind, Clinical Trial. J. Intern. Med. 2018, 284, 307–317. [Google Scholar] [CrossRef]
- Takimoto, T.; Hatanaka, M.; Hoshino, T.; Takara, T.; Tanaka, K.; Shimizu, A.; Morita, H.; Nakamura, T. Effect of Bacillus subtilis C-3102 on Bone Mineral Density in Healthy Postmenopausal Japanese Women: A Randomized, Placebo-Controlled, Double-Blind Clinical Trial. Biosci. Microbiota Food Health 2018, 37, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzoli, R.; Biver, E. Are Probiotics the New Calcium and Vitamin D for Bone Health? Curr. Osteoporos. Rep. 2020, 18, 273–284. [Google Scholar] [CrossRef]
- Sojan, J.M.; Gioacchini, G.; Giorgini, E.; Orlando, P.; Tiano, L.; Maradonna, F.; Carnevali, O. Zebrafish Caudal Fin as a Model to Investigate the Role of Probiotics in Bone Regeneration. Sci. Rep. 2022, 12, 8057. [Google Scholar] [CrossRef] [PubMed]
- Narva, M.; Halleen, J.; Väänänen, K.; Korpela, R. Effects of Lactobacillus helveticus Fermented Milk on Bone Cells In Vitro. Life Sci. 2004, 75, 1727–1734. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Hsieh, Y.-M.; Huang, C.-C.; Tsai, C.-C. Inhibitory Effects of Probiotic Lactobacillus on the Growth of Human Colonic Carcinoma Cell Line HT-29. Molecules 2017, 22, 107. [Google Scholar] [CrossRef] [Green Version]
- Nozari, S.; Faridvand, Y.; Etesami, A.; Ahmad Khan Beiki, M.; Miresmaeili Mazrakhondi, S.A.; Abdolalizadeh, J. Potential Anticancer Effects of Cell Wall Protein Fractions from Lactobacillus paracasei on Human Intestinal Caco-2 Cell Line. Lett. Appl. Microbiol. 2019, 69, 148–154. [Google Scholar] [CrossRef]
- Brognara, L.; Salmaso, L.; Mazzotti, A.; Di Martino, A.; Faldini, C.; Cauli, O. Effects of Probiotics in the Management of Infected Chronic Wounds: From Cell Culture to Human Studies. Curr. Clin. Pharmacol. 2020, 15, 193–206. [Google Scholar] [CrossRef]
- Isazadeh, A.; Hajazimian, S.; Shadman, B.; Safaei, S.; Bedoustani, A.B.; Chavoshi, R.; Shanehbandi, D.; Mashayekhi, M.; Nahaei, M.; Baradaran, B. Anti-Cancer Effects of Probiotic Lactobacillus acidophilus for Colorectal Cancer Cell Line Caco-2 through Apoptosis Induction. Pharm. Sci. 2020, 27, 262–267. [Google Scholar] [CrossRef]
- Subramaniam, M.; Jalal, S.M.; Rickard, D.J.; Harris, S.A.; Bolander, M.E.; Spelsberg, T.C. Further Characterization of Human Fetal Osteoblastic HFOB 1.19 and HFOB/ER? Cells: Bone Formation In Vivo and Karyotype Analysis Using Multicolor Fluorescent In Situ Hybridization. J. Cell. Biochem. 2002, 87, 9–15. [Google Scholar] [CrossRef]
- Beck, G.R.; Zerler, B.; Moran, E. Phosphate Is a Specific Signal for Induction of Osteopontin Gene Expression. Proc. Natl. Acad. Sci. USA 2000, 97, 8352–8357. [Google Scholar] [CrossRef]
- Ansari, S.; Ito, K.; Hofmann, S. Alkaline Phosphatase Activity of Serum Affects Osteogenic Differentiation Cultures. ACS Omega 2022, 7, 12724–12733. [Google Scholar] [CrossRef] [PubMed]
- Strzelecka-Kiliszek, A.; Bozycki, L.; Mebarek, S.; Buchet, R.; Pikula, S. Characteristics of Minerals in Vesicles Produced by Human Osteoblasts HFOB 1.19 and Osteosarcoma Saos-2 Cells Stimulated for Mineralization. J. Inorg. Biochem. 2017, 171, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Bozycki, L.; Mroczek, J.; Bessueille, L.; Mebarek, S.; Buchet, R.; Pikula, S.; Strzelecka-Kiliszek, A. Annexins A2, A6 and Fetuin-A Affect the Process of Mineralization in Vesicles Derived from Human Osteoblastic HFOB 1.19 and Osteosarcoma Saos-2 Cells. Int. J. Mol. Sci. 2021, 22, 3993. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Fu, Y.; Zhao, X.-H. In Vitro Responses of HFOB1.19 Cells towards Chum Salmon (Oncorhynchus keta) Skin Gelatin Hydrolysates in Cell Proliferation, Cycle Progression and Apoptosis. J. Funct. Foods 2013, 5, 279–288. [Google Scholar] [CrossRef]
- Ruiz-Gaspà, S.; Guañabens, N.; Enjuanes, A.; Peris, P.; Martinez-Ferrer, A.; Martinez de Osaba, M.J.; Gonzalez, B.; Alvarez, L.; Monegal, A.; Combalia, A.; et al. Lithocholic Acid Downregulates Vitamin D Effects in Human Osteoblasts. Eur. J. Clin. Investig. 2010, 40, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Ayansola, H.; Ma, Y.; Ito, K.; Guo, Y.; Zhang, B. Advances in Enhanced Menaquinone-7 Production from Bacillus subtilis. Front. Bioeng. Biotechnol. 2021, 9, 695526. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.-J.; Gao, H.; Jin, J.-S.; Ahn, B.-Y. A Comparatively Study of Menaquinone-7 Isolated from Cheonggukjang with Vitamin K1 and Menaquinone-4 on Osteoblastic Cells Differentiation and Mineralization. Food Chem. Toxicol. 2019, 131, 110540. [Google Scholar] [CrossRef]
- Tang, H.; Zhu, Z.; Zheng, Z.; Wang, H.; Li, C.; Wang, L.; Zhao, G.; Wang, P. A Study of Hydrophobins-Modified Menaquinone-7 on Osteoblastic Cells Differentiation. Mol. Cell. Biochem. 2021, 476, 1939–1948. [Google Scholar] [CrossRef]
- Sun, L.; Wu, L.; Bao, C.; Fu, C.; Wang, X.; Yao, J.; Zhang, X.; van Blitterswijk, C.A. Gene Expressions of Collagen Type I, ALP and BMP-4 in Osteo-Inductive BCP Implants Show Similar Pattern to That of Natural Healing Bones. Mater. Sci. Eng. C 2009, 29, 1829–1834. [Google Scholar] [CrossRef]
- Yuan, Y.; Jin, S.; Qi, X.; Chen, X.; Zhang, W.; Yang, K.; Zhong, H. Osteogenesis Stimulation by Copper-Containing 316L Stainless Steel via Activation of Akt Cell Signaling Pathway and Runx2 Upregulation. J. Mater. Sci. Technol. 2019, 35, 2727–2733. [Google Scholar] [CrossRef]
- Boonrungsiman, S.; Gentleman, E.; Carzaniga, R.; Evans, N.D.; McComb, D.W.; Porter, A.E.; Stevens, M.M. The Role of Intracellular Calcium Phosphate in Osteoblast-Mediated Bone Apatite Formation. Proc. Natl. Acad. Sci. USA 2012, 109, 14170–14175. [Google Scholar] [CrossRef] [Green Version]
- Mann, V.; Grimm, D.; Corydon, T.J.; Krüger, M.; Wehland, M.; Riwaldt, S.; Sahana, J.; Kopp, S.; Bauer, J.; Reseland, J.E.; et al. Changes in Human Foetal Osteoblasts Exposed to the Random Positioning Machine and Bone Construct Tissue Engineering. Int. J. Mol. Sci. 2019, 20, 1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sojan, J.M.; Raman, R.; Muller, M.; Carnevali, O.; Renn, J. Probiotics Enhance Bone Growth and Rescue BMP Inhibition: New Transgenic Zebrafish Lines to Study Bone Health. Int. J. Mol. Sci. 2022, 23, 4748. [Google Scholar] [CrossRef] [PubMed]
- Atkins, G.J.; Welldon, K.J.; Wijenayaka, A.R.; Bonewald, L.F.; Findlay, D.M. Vitamin K Promotes Mineralization, Osteoblast-to-Osteocyte Transition, and an Anticatabolic Phenotype by γ-Carboxylation-Dependent and -Independent Mechanisms. Am. J. Physiol. Cell Physiol. 2009, 297, C1358–C1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Zhang, S.; Liu, J.; Liu, Y.; Liang, Q. Vitamin K2 Stimulates MC3T3-E1 Osteoblast Differentiation and Mineralization through Autophagy Induction. Mol. Med. Rep. 2019, 19, 3676–3684. [Google Scholar] [CrossRef]
- Zhang, Y.-L.; Yin, J.-H.; Ding, H.; Zhang, W.; Zhang, C.-Q.; Gao, Y.-S. Vitamin K2 Prevents Glucocorticoid-Induced Osteonecrosis of the Femoral Head in Rats. Int. J. Biol. Sci. 2016, 12, 347–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, S.; Yamada, A.; Tanabe, R.; Nakaoka, K.; Hosoi, T.; Goseki-Sone, M. Menaquinone-4 (Vitamin K2) up-Regulates Expression of Human Intestinal Alkaline Phosphatase in Caco-2 Cells. Nutr. Res. 2016, 36, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Horie-Inoue, K.; Inoue, S. Steroid and Xenobiotic Receptor Mediates a Novel Vitamin K2 Signaling Pathway in Osteoblastic Cells. J. Bone Miner. Metab. 2008, 26, 9–12. [Google Scholar] [CrossRef]
- Koshihara, Y.; Hoshi, K.; Okawara, R.; Ishibashi, H.; Yamamoto, S. Vitamin K Stimulates Osteoblastogenesis and Inhibits Osteoclastogenesis in Human Bone Marrow Cell Culture. J. Endocrinol. 2003, 176, 339–348. [Google Scholar] [CrossRef] [Green Version]
- Weng, S.-J.; Yan, D.-Y.; Gu, L.-J.; Chen, L.; Xie, Z.-J.; Wu, Z.-Y.; Tang, J.-H.; Shen, Z.-J.; Li, H.; Bai, B.-L.; et al. Combined Treatment with Vitamin K2 and PTH Enhanced Bone Formation in Ovariectomized Rats and Increased Differentiation of Osteoblast In Vitro. Chem. Biol. Interact. 2019, 300, 101–110. [Google Scholar] [CrossRef]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Steroid and Xenobiotic Receptor SXR Mediates Vitamin K2-Activated Transcription of Extracellular Matrix-Related Genes and Collagen Accumulation in Osteoblastic Cells. J. Biol. Chem. 2006, 281, 16927–16934. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proliferation | ECM Maturation | ECM Mineralization |
|---|---|---|
| TGFB1 | SPP1 | ITGB1 |
| RUNX2 | BGLAP | ITGB3 |
| RUNX3 | SPARC | ITGB5 |
| NELL1 | ALPL | RUNX3 |
| BMP7 | IBSP | DCN |
| IBSP | COL3A1 | COL3A1 |
| COL1A1 | ||
| COL2A1 | ||
| ALPL |
| Groups | Composition |
|---|---|
| Control (C) | DMEM + FBS 10% + 0.03 mg/mL G418 |
| P1 | DMEM + FBS 10% + 0.03 mg/mL G418 + P1 extract 1 µL/mL |
| P2 | DMEM + FBS 10% + 0.03 mg/mL G418 + P2 extract 1 µL/mL |
| VD | DMEM + FBS 10% + 0.03 mg/mL G418 + 100 nM VD |
| P1VD | DMEM + FBS 10% + 0.03 mg/mL G418 + P1 extract 1 µL/mL + 100 nM VD |
| P2VD | DMEM + FBS 10% + 0.03 mg/mL G418 + P2 extract 1 µL/mL + 100 nM VD |
| Gene | Forward Primer (5′-3′) | Reverse Primer (3′-5′) |
|---|---|---|
| ACTB | ATTGGCAATGAGCGGTTC | GGATGCCACAGGACTCCAT |
| RPLP0 | CTGGAAAACAACCCAGCTCT | GAGGTCCTCCTTGGTGAACA |
| RUNX2 | GTGCCTAGGCGCATTTCA | GCTCTTCTTACTGAGAGTGGAAGG |
| ALPL | CCATCCTGTATGGCAATGG | CGCCTGGTAGTTGTTGTGAG |
| TGFB1 | GCAGCACGTGGAGCTGTA | CAGCCGGTTGCTGAGGTA |
| COL3A1 | CTGGACCCCAGGGTCTTC | CATCTGATCCAGGGTTTCCA |
| SPP1 | TTGCAGCCTTCTCAGCCAA | CAAAAGCAAATCACTGCAATTCTC |
| SPARC | GTACATCGCCCTGGATGAGT | CGAAGGGGAGGGTTAAAGAG |
| BGN | CAGCCCGCCAACTAGTCA | GGCCAGCAGAGACACGAG |
| DCN | GGAGACTTTAAGAACCTGAAGAACC | CGTTCCAACTTCACCAAAGG |
| BGLAP | ACACTCCTCGCCCTATTG | GATGTGGTCAGCCAACTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sojan, J.M.; Licini, C.; Marcheggiani, F.; Carnevali, O.; Tiano, L.; Mattioli-Belmonte, M.; Maradonna, F. Bacillus subtilis Modulated the Expression of Osteogenic Markers in a Human Osteoblast Cell Line. Cells 2023, 12, 364. https://doi.org/10.3390/cells12030364
Sojan JM, Licini C, Marcheggiani F, Carnevali O, Tiano L, Mattioli-Belmonte M, Maradonna F. Bacillus subtilis Modulated the Expression of Osteogenic Markers in a Human Osteoblast Cell Line. Cells. 2023; 12(3):364. https://doi.org/10.3390/cells12030364
Chicago/Turabian StyleSojan, Jerry Maria, Caterina Licini, Fabio Marcheggiani, Oliana Carnevali, Luca Tiano, Monica Mattioli-Belmonte, and Francesca Maradonna. 2023. "Bacillus subtilis Modulated the Expression of Osteogenic Markers in a Human Osteoblast Cell Line" Cells 12, no. 3: 364. https://doi.org/10.3390/cells12030364