
The Suppression of Ubiquitin C-Terminal Hydrolase L1 Promotes the Transdifferentiation of Auditory Supporting Cells into Hair Cells by Regulating the mTOR Pathway

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Organ of Corti Explants
2.3. Drug Treatment
2.4. Quantitative Polymerase Chain Reaction
2.5. Immunohistochemistry
2.6. Immunofluorescence
2.7. Western Blot Analysis
2.8. 5-Ethynyl-2′-deoxyuridine Incorporation Assay
2.9. Hematoxylin and Eosin Staining and Histological Analysis
2.10. Statistical Analyses
3. Results
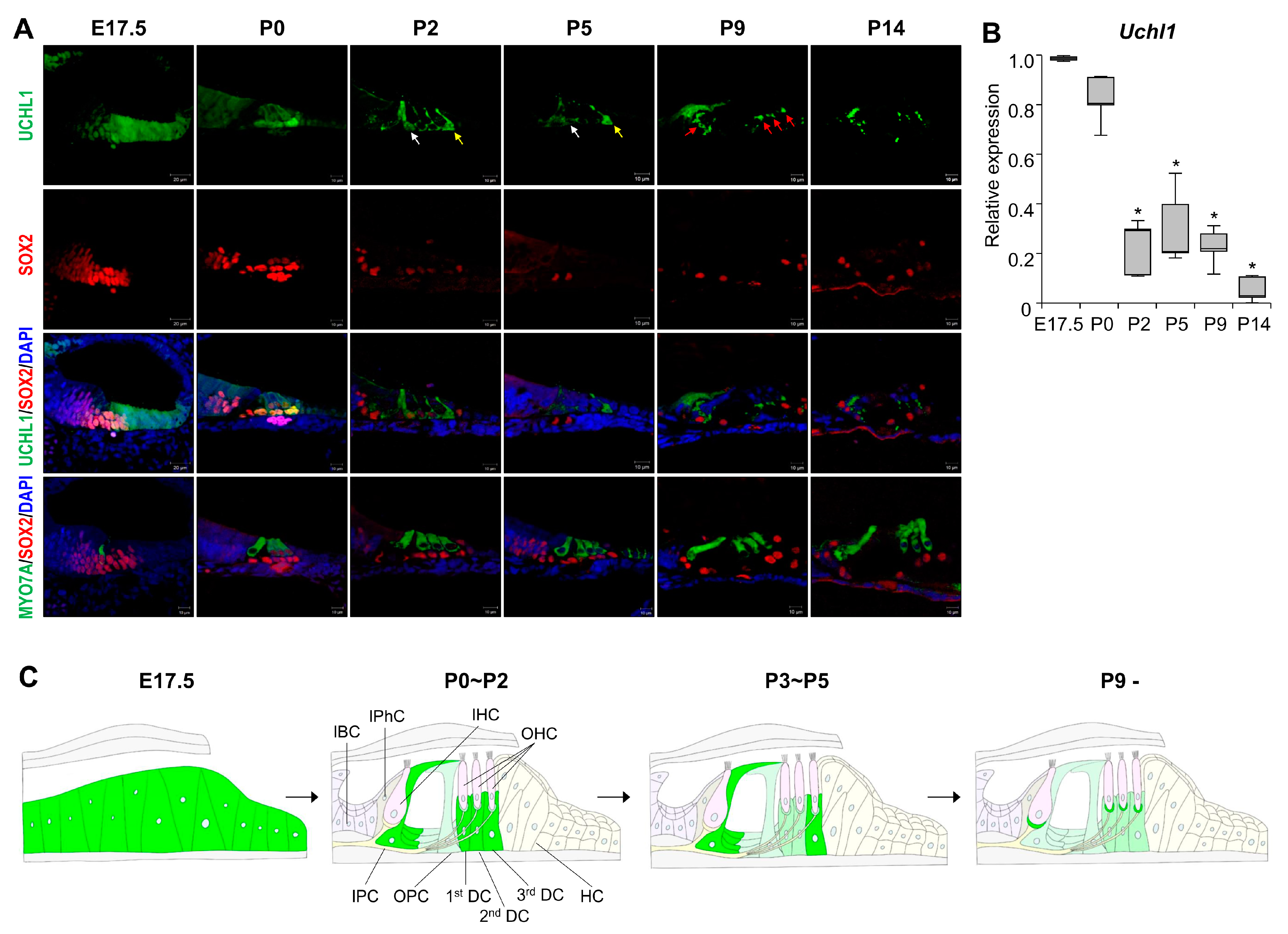
3.1. Dynamic Expression of UCHL1 in the Developing Cochlea
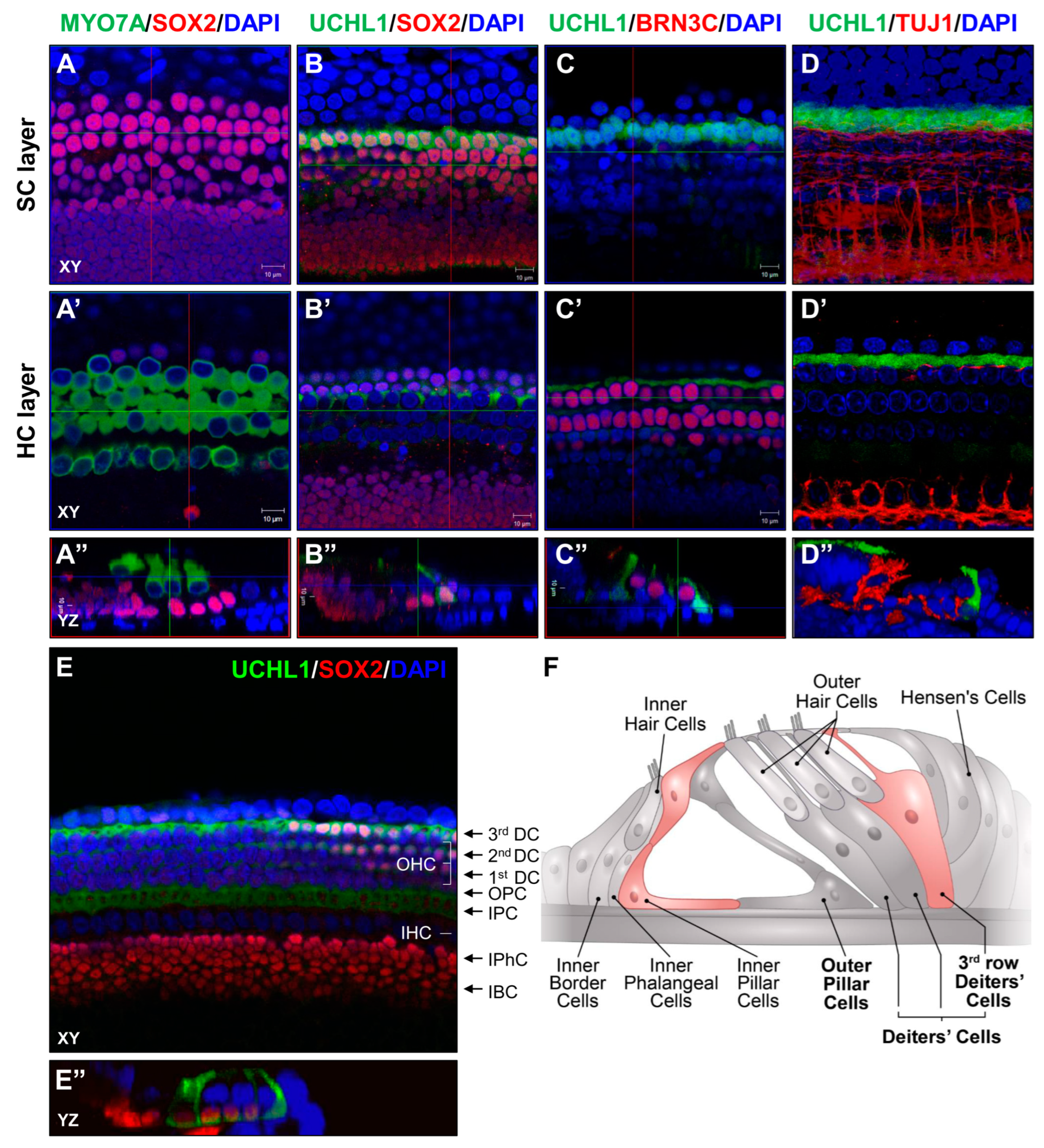
3.2. UCHL1 Expression Is Restricted to the 3rd DCs and IPCs in the Neonatal Organ of Corti
3.3. A Subpopulation of UCHL1-Expressing Cells Switches to HCs under Transdifferentiation
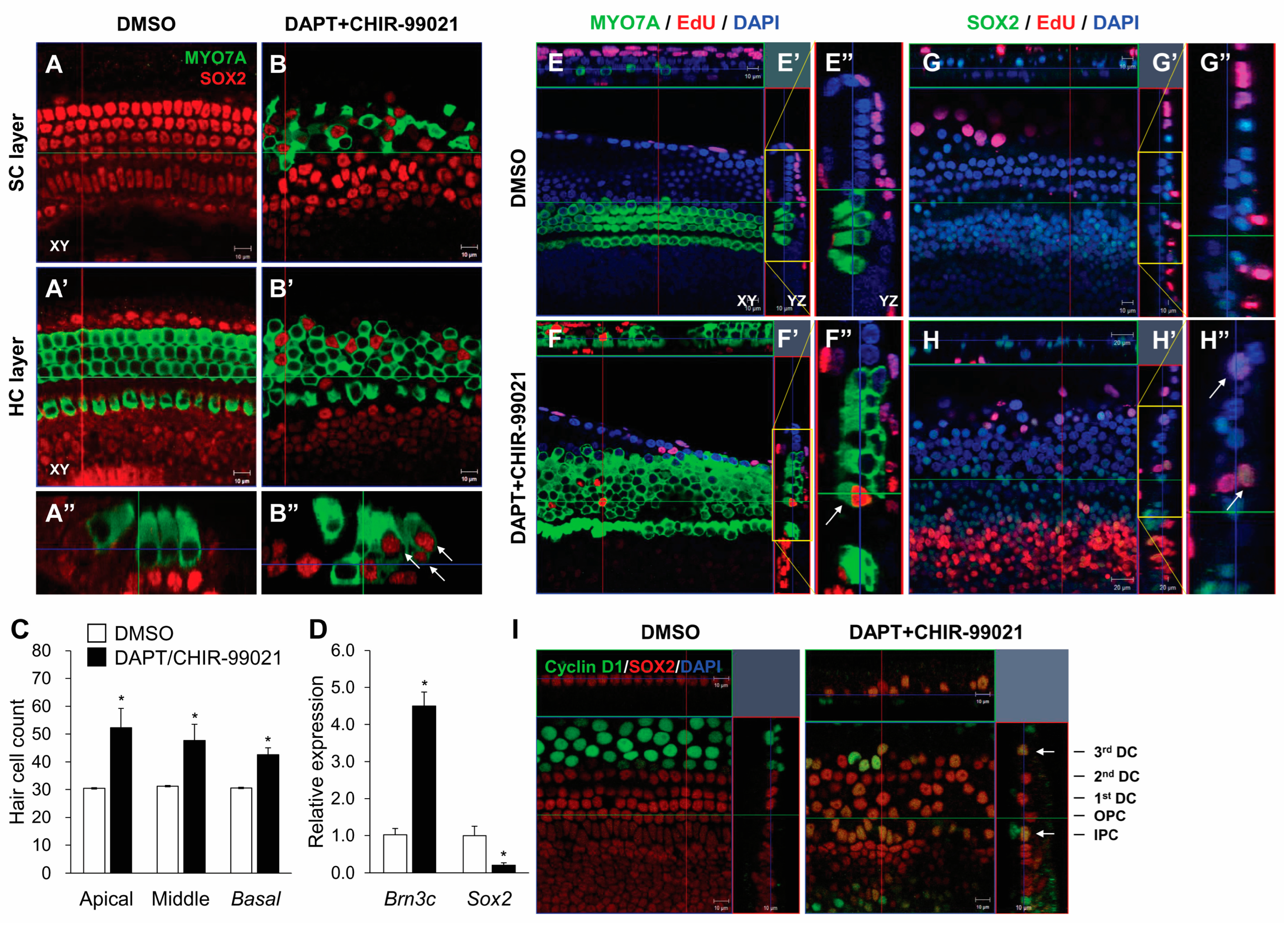
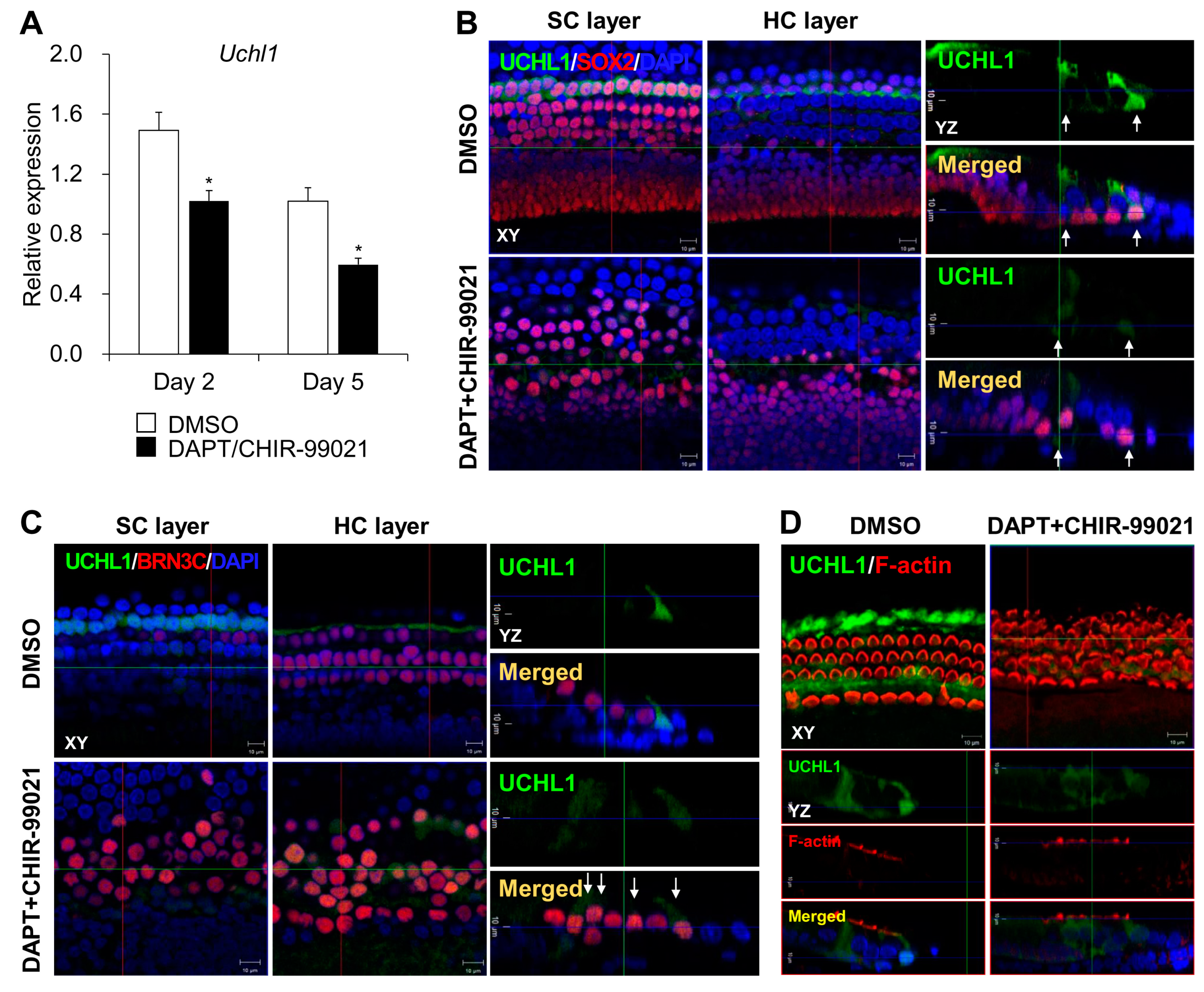
3.4. The UCHL1 Inhibitor LDN-57444 Promotes the Transdifferentiation of SCs into HCs
3.5. mTOR Complex 1 Activation by a UCHL1 Inhibitor Promotes HC Transdifferentiation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qi, J.; Huang, W.; Lu, Y.; Yang, X.; Zhou, Y.; Chen, T.; Wang, X.; Yu, Y.; Sun, J.Q.; Chai, R. Stem Cell-Based Hair Cell Regeneration and Therapy in the Inner Ear. Neurosci. Bull. 2024, 40, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qiang, R.; Dong, Y.; Zhang, Y.; Chen, Y.; Zhou, H.; Gao, X.; Chai, R. Hair cell regeneration from inner ear progenitors in the mammalian cochlea. Am. J. Stem Cells 2020, 9, 25–35. [Google Scholar] [PubMed]
- Gao, J.; Fan, L.; Zhao, L.; Su, Y. The interaction of Notch and Wnt signaling pathways in vertebrate regeneration. Cell Regen. 2021, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Smith-Cortinez, N.; Yadak, R.; Hendriksen, F.G.J.; Sanders, E.; Ramekers, D.; Stokroos, R.J.; Versnel, H.; Straatman, L.V. LGR5-Positive Supporting Cells Survive Ototoxic Trauma in the Adult Mouse Cochlea. Front. Mol. Neurosci. 2021, 14, 729625. [Google Scholar] [CrossRef] [PubMed]
- Day, I.N.; Thompson, R.J. UCHL1 (PGP 9.5): Neuronal biomarker and ubiquitin system protein. Prog. Neurobiol. 2010, 90, 327–362. [Google Scholar] [CrossRef] [PubMed]
- Larsen, C.N.; Price, J.S.; Wilkinson, K.D. Substrate binding and catalysis by ubiquitin C-terminal hydrolases: Identification of two active site residues. Biochemistry 1996, 35, 6735–6744. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; He, J.; Yang, C.; Orange, M.; Ren, X.; Blair, N.; Tan, T.; Yang, J.M.; Zhu, H. UCH-L1 promotes invasion of breast cancer cells through activating Akt signaling pathway. J. Cell. Biochem. 2018, 119, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Diaz, P.C.; Chang, J.C.; Moses, E.S.; Dao, T.; Chen, Y.; Hung, J.Y. Ubiquitin carboxyl-terminal esterase L1 (UCHL1) is associated with stem-like cancer cell functions in pediatric high-grade glioma. PLoS ONE 2017, 12, e0176879. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kim, K.; Lee, Y.Y.; Choo, O.S.; Jang, J.H.; Choung, Y.H. Downregulated UCHL1 Accelerates Gentamicin-Induced Auditory Cell Death via Autophagy. Mol. Neurobiol. 2019, 56, 7433–7447. [Google Scholar] [CrossRef]
- Herget, M.; Scheibinger, M.; Guo, Z.; Jan, T.A.; Adams, C.M.; Cheng, A.G.; Heller, S. A simple method for purification of vestibular hair cells and non-sensory cells, and application for proteomic analysis. PLoS ONE 2013, 8, e66026. [Google Scholar] [CrossRef]
- Samarajeewa, A.; Lenz, D.R.; Xie, L.; Chiang, H.; Kirchner, R.; Mulvaney, J.F.; Edge, A.S.B.; Dabdoub, A. Transcriptional response to Wnt activation regulates the regenerative capacity of the mammalian cochlea. Development 2018, 145, dev166579. [Google Scholar] [CrossRef]
- Ni, W.; Zeng, S.; Li, W.; Chen, Y.; Zhang, S.; Tang, M.; Sun, S.; Chai, R.; Li, H. Wnt activation followed by Notch inhibition promotes mitotic hair cell regeneration in the postnatal mouse cochlea. Oncotarget 2016, 7, 66754–66768. [Google Scholar] [CrossRef]
- Gao, H.; Freeling, J.; Wu, P.; Liang, A.P.; Wang, X.; Li, Y. UCHL1 regulates muscle fibers and mTORC1 activity in skeletal muscle. Life Sci. 2019, 233, 116699. [Google Scholar] [CrossRef]
- Hussain, S.; Bedekovics, T.; Liu, Q.; Hu, W.; Jeon, H.; Johnson, S.H.; Vasmatzis, G.; May, D.G.; Roux, K.J.; Galardy, P.J. UCH-L1 bypasses mTOR to promote protein biosynthesis and is required for MYC-driven lymphomagenesis in mice. Blood 2018, 132, 2564–2574. [Google Scholar] [CrossRef]
- Naujokat, C.; Saric, T. Concise review: Role and function of the ubiquitin-proteasome system in mammalian stem and progenitor cells. Stem Cells 2007, 25, 2408–2418. [Google Scholar] [CrossRef]
- Buckley, S.M.; Aranda-Orgilles, B.; Strikoudis, A.; Apostolou, E.; Loizou, E.; Moran-Crusio, K.; Farnsworth, C.L.; Koller, A.A.; Dasgupta, R.; Silva, J.C.; et al. Regulation of pluripotency and cellular reprogramming by the ubiquitin-proteasome system. Cell Stem Cell 2012, 11, 783–798. [Google Scholar] [CrossRef]
- Rhie, B.H.; Antao, A.M.; Karapurkar, J.K.; Kim, M.S.; Jo, W.J.; Ramakrishna, S.; Kim, K.S. Ubiquitin-Specific Protease 3 Deubiquitinates and Stabilizes Oct4 Protein in Human Embryonic Stem Cells. Int. J. Mol. Sci. 2021, 22, 5584. [Google Scholar] [CrossRef]
- Cox, B.C.; Chai, R.; Lenoir, A.; Liu, Z.; Zhang, L.; Nguyen, D.H.; Chalasani, K.; Steigelman, K.A.; Fang, J.; Rubel, E.W.; et al. Spontaneous hair cell regeneration in the neonatal mouse cochlea in vivo. Development 2014, 141, 816–829. [Google Scholar] [CrossRef]
- Gao, H.; Hartnett, S.; Li, Y. Ubiquitin C-Terminal Hydrolase L1 regulates myoblast proliferation and differentiation. Biochem. Biophys. Res. Commun. 2017, 492, 96–102. [Google Scholar] [CrossRef]
- Feng, Z.; Tao, S.; Huang, Z.; Zheng, B.; Kong, X.; Xiang, Y.; Zhang, Q.; Song, H.; Xu, Z.; Wei, X.; et al. The deubiquitinase UCHL1 negatively controls osteoclastogenesis by regulating TAZ/NFATC1 signalling. Int. J. Biol. Sci. 2023, 19, 2319–2332. [Google Scholar] [CrossRef]
- Graham, S.H.; Liu, H. Life and death in the trash heap: The ubiquitin proteasome pathway and UCHL1 in brain aging, neurodegenerative disease and cerebral Ischemia. Ageing Res. Rev. 2017, 34, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Luo, L.; Chen, J. Roles of mTOR Signaling in Tissue Regeneration. Cells 2019, 8, 1075. [Google Scholar] [CrossRef]
- Shu, Y.; Li, W.; Huang, M.; Quan, Y.Z.; Scheffer, D.; Tian, C.; Tao, Y.; Liu, X.; Hochedlinger, K.; Indzhykulian, A.A.; et al. Renewed proliferation in adult mouse cochlea and regeneration of hair cells. Nat. Commun. 2019, 10, 5530. [Google Scholar] [CrossRef]
- Li, X.J.; Doetzlhofer, A. LIN28B/let-7 control the ability of neonatal murine auditory supporting cells to generate hair cells through mTOR signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 22225–22236. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Feldman, A.L.; Das, C.; Ziesmer, S.C.; Ansell, S.M.; Galardy, P.J. Ubiquitin hydrolase UCH-L1 destabilizes mTOR complex 1 by antagonizing DDB1-CUL4-mediated ubiquitination of raptor. Mol. Cell. Biol. 2013, 33, 1188–1197. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Hu, L.; Edge, A.S. Generation of hair cells in neonatal mice by beta-catenin overexpression in Lgr5-positive cochlear progenitors. Proc. Natl. Acad. Sci. USA 2013, 110, 13851–13856. [Google Scholar] [CrossRef]
- Bheda, A.; Yue, W.; Gullapalli, A.; Whitehurst, C.; Liu, R.; Pagano, J.S.; Shackelford, J. Positive reciprocal regulation of ubiquitin C-terminal hydrolase L1 and beta-catenin/TCF signaling. PLoS ONE 2009, 4, e5955. [Google Scholar] [CrossRef]
- Kabuta, T.; Mitsui, T.; Takahashi, M.; Fujiwara, Y.; Kabuta, C.; Konya, C.; Tsuchiya, Y.; Hatanaka, Y.; Uchida, K.; Hohjoh, H.; et al. Ubiquitin C-terminal hydrolase L1 (UCH-L1) acts as a novel potentiator of cyclin-dependent kinases to enhance cell proliferation independently of its hydrolase activity. J. Biol. Chem. 2013, 288, 12615–12626. [Google Scholar] [CrossRef]
- Kwan, S.Y.; Au-Yeung, C.L.; Yeung, T.L.; Rynne-Vidal, A.; Wong, K.K.; Risinger, J.I.; Lin, H.K.; Schmandt, R.E.; Yates, M.S.; Mok, S.C.; et al. Ubiquitin Carboxyl-Terminal Hydrolase L1 (UCHL1) Promotes Uterine Serous Cancer Cell Proliferation and Cell Cycle Progression. Cancers 2020, 12, 118. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.J.; Jeong, I.H.; Ha, J.H.; Kim, Y.S.; Sung, S.; Jang, J.H.; Choung, Y.-H. The Suppression of Ubiquitin C-Terminal Hydrolase L1 Promotes the Transdifferentiation of Auditory Supporting Cells into Hair Cells by Regulating the mTOR Pathway. Cells 2024, 13, 737. https://doi.org/10.3390/cells13090737
Kim YJ, Jeong IH, Ha JH, Kim YS, Sung S, Jang JH, Choung Y-H. The Suppression of Ubiquitin C-Terminal Hydrolase L1 Promotes the Transdifferentiation of Auditory Supporting Cells into Hair Cells by Regulating the mTOR Pathway. Cells. 2024; 13(9):737. https://doi.org/10.3390/cells13090737
Chicago/Turabian StyleKim, Yeon Ju, In Hye Jeong, Jung Ho Ha, Young Sun Kim, Siung Sung, Jeong Hun Jang, and Yun-Hoon Choung. 2024. "The Suppression of Ubiquitin C-Terminal Hydrolase L1 Promotes the Transdifferentiation of Auditory Supporting Cells into Hair Cells by Regulating the mTOR Pathway" Cells 13, no. 9: 737. https://doi.org/10.3390/cells13090737