
The Regulation of Cellular Responses to Mechanical Cues by Rho GTPases

{kind=link}
Abstract
:1. Introduction
2. First Contact with the ECM
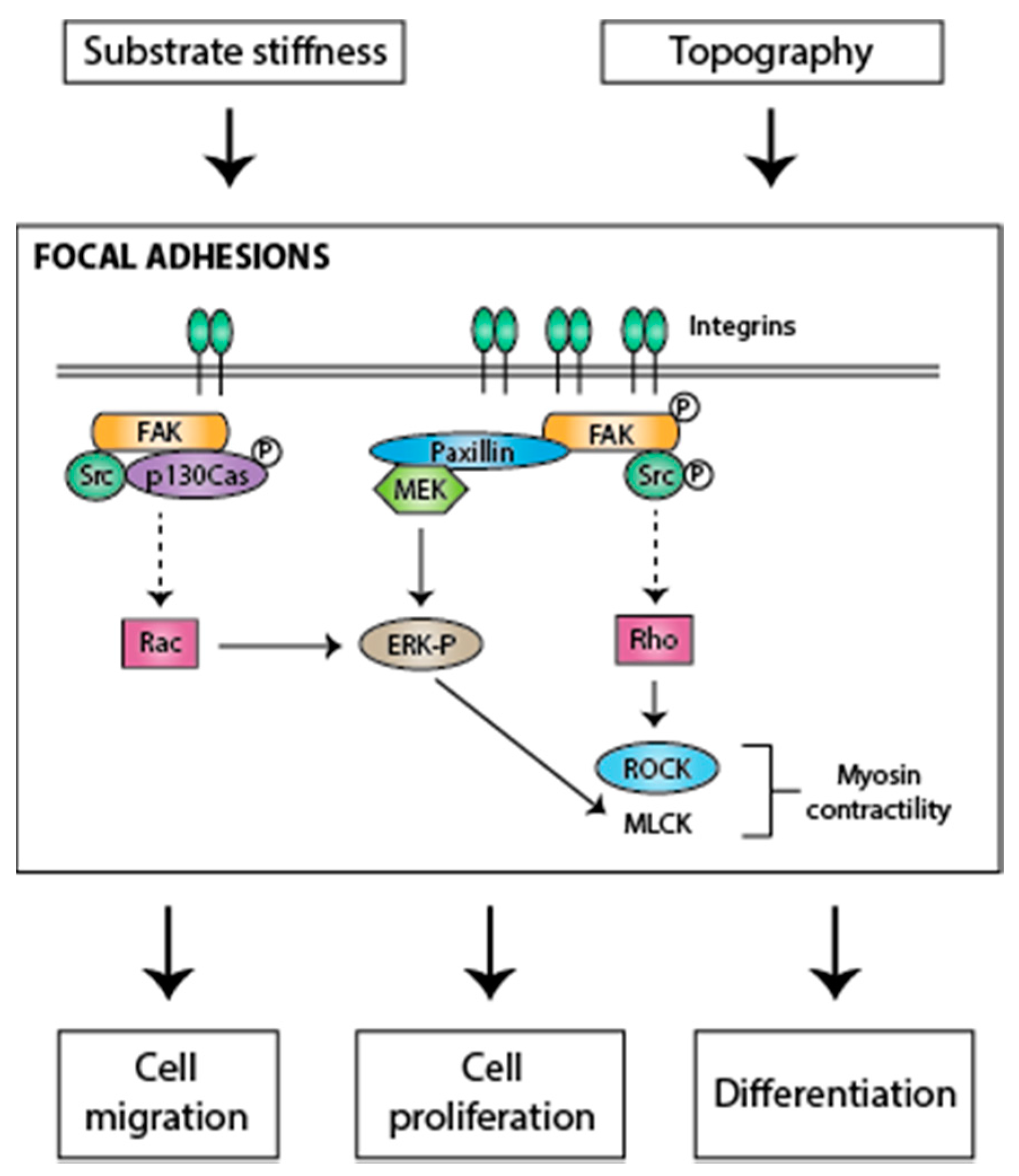
3. The Regulation of Cellular Responses to Mechanical Cues by Rho GTPases
| Stiffness |
| The stiffness of an object is the extent to which it is able to resist deformation by an applied force. It is a structural property and is influenced by the geometry of the material. |
| Elastic modulus |
| The elastic modulus (E) measures the resistance of a material to being deformed elastically when a force is applied to it. It is a material property and is not influenced by geometry. The elastic modulus is measured by E = σ/ε (stress/strain). A stiffer material has a higher elastic modulus. |
| Contractile force |
| The contractile force is the force generated by a cell within a three-dimensional (3D) environment. It is resisted by the stiffness of the environment. |
| Cell traction force |
| The cell traction force is the tangential tension exerted by cells on the substrate. It is generated by actomyosin interaction and actin polymerization. |
| Topography |
| Topography is the study of features or structures on a surface. |
| Isotropic topography |
| Isotropic topography refers to features that are the same in all directions, without any continuous surface. |
| Anisotropic topography |
| Anisotropic topography refers to features that are different in all directions, often observed with a continuous surface. |
| Contact guidance |
| Contact guidance is a phenomenon where the physical environment modulates the alignment and orientation of the cell and its cytoskeleton components. |
4. Cell Migration
4.1. Cell Migration in a 3D Matrix
4.2. Topography and Cell Migration
5. Cell Proliferation
Cell Shape Affects Proliferation
6. Differentiation
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Sit, S.T.; Manser, E. Rho gtpases and their role in organizing the actin cytoskeleton. J. Cell. Sci. 2011, 124, 679–683. [Google Scholar] [CrossRef] [PubMed]
- Heasman, S.J.; Ridley, A.J. Mammalian rho gtpases: New insights into their functions from in vivo studies. Nat. Rev. Mol. Cell. Biol. 2008, 9, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Koh, C.G. Rho gtpases and their regulators in neuronal functions and development. Neurosignals 2006, 15, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Heng, Y.W.; Koh, C.G. Actin cytoskeleton dynamics and the cell division cycle. Int. J. Biochem. Cell. Biol. 2010, 42, 1622–1633. [Google Scholar] [CrossRef] [PubMed]
- Etienne-Manneville, S.; Hall, A. Rho gtpases in cell biology. Nature 2002, 420, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.F.; Ashworth, A.; Hall, A. An essential role for rho, rac, and cdc42 gtpases in cell cycle progression through g1. Science 1995, 269, 1270–1272. [Google Scholar] [CrossRef] [PubMed]
- Narumiya, S.; Yasuda, S. Rho gtpases in animal cell mitosis. Curr. Opin. Cell. Biol. 2006, 18, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Gefen, A.; Margulies, S.S. Are in vivo and in situ brain tissues mechanically similar? J. Biomech. 2004, 37, 1339–1352. [Google Scholar] [CrossRef] [PubMed]
- Tilghman, R.W.; Cowan, C.R.; Mih, J.D.; Koryakina, Y.; Gioeli, D.; Slack-Davis, J.K.; Blackman, B.R.; Tschumperlin, D.J.; Parsons, J.T. Matrix rigidity regulates cancer cell growth and cellular phenotype. PLoS ONE 2010, 5, e12905. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Shea, B.S.; Mih, J.D.; Tager, A.M.; Tschumperlin, D.J. Lung parenchymal tissue stiffness in fibrosis and cellular responses to substrate stiffness. Biophys. J. 2009, 96, 395a. [Google Scholar] [CrossRef]
- Engler, A.J.; Griffin, M.A.; Sen, S.; Bönnemann, C.G.; Sweeney, H.L.; Discher, D.E. Myotubes differentiate optimally on substrates with tissue-like stiffness: Pathological implications for soft or stiff microenvironments. J. Cell Biol. 2004, 166, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, Y.; Yasuike, T.; Yagi, A.; Yamada, T. Transverse elasticity of myofibrils of rabbit skeletal muscle studied by atomic force microscopy. Biochem. Biophys. Res. Commun. 1999, 256, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Paszek, M.J.; Zahir, N.; Johnson, K.R.; Lakins, J.N.; Rozenberg, G.I.; Gefen, A.; Reinhart-King, C.A.; Margulies, S.S.; Dembo, M.; Boettiger, D.; et al. Tensional homeostasis and the malignant phenotype. Cancer Cell. 2005, 8, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Tonazzini, I.; Cecchini, A.; Elgersma, Y.; Cecchini, M. Interaction of sh-sy5y cells with nanogratings during neuronal differentiation: Comparison with primary neurons. Adv. Healthc. Mater. 2014, 3, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Chua, J.S.; Chng, C.P.; Moe, A.A.; Tann, J.Y.; Goh, E.L.; Chiam, K.H.; Yim, E.K. Extending neurites sense the depth of the underlying topography during neuronal differentiation and contact guidance. Biomaterials 2014, 35, 7750–7761. [Google Scholar] [CrossRef] [PubMed]
- Yim, E.K.; Pang, S.W.; Leong, K.W. Synthetic nanostructures inducing differentiation of human mesenchymal stem cells into neuronal lineage. Exp. Cell. Res. 2007, 313, 1820–1829. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; McBeath, R.; Chen, C.S. Stem cell shape regulates a chondrogenic versus myogenic fate through rac1 and n-cadherin. Stem Cells 2010, 28, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Lu, H.; Kawazoe, N.; Chen, G. Adipogenic differentiation of individual mesenchymal stem cell on different geometric micropatterns. Langmuir 2011, 27, 6155–6162. [Google Scholar] [CrossRef] [PubMed]
- Kilian, K.A.; Bugarija, B.; Lahn, B.T.; Mrksich, M. Geometric cues for directing the differentiation of mesenchymal stem cells. Proc. Natl. Acad. Sci. USA 2010, 107, 4872–4877. [Google Scholar] [CrossRef] [PubMed]
- McBeath, R.; Pirone, D.M.; Nelson, C.M.; Bhadriraju, K.; Chen, C.S. Cell shape, cytoskeletal tension, and rhoa regulate stem cell lineage commitment. Dev. Cell. 2004, 6, 483–495. [Google Scholar] [CrossRef]
- Guilluy, C.; Swaminathan, V.; Garcia-Mata, R.; O’Brien, E.T.; Superfine, R.; Burridge, K. The rho gefs larg and gef-h1 regulate the mechanical response to force on integrins. Nat. Cell. Biol. 2011, 13, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Chen, Q.K.; Lui, C.; Cichon, M.A.; Radisky, D.C.; Nelson, C.M. Matrix compliance regulates rac1b localization, nadph oxidase assembly, and epithelial–mesenchymal transition. Mol. Biol. Cell 2012, 23, 4097–4108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Landmann, F.; Zahreddine, H.; Rodriguez, D.; Koch, M.; Labouesse, M. A tension-induced mechanotransduction pathway promotes epithelial morphogenesis. Nature 2011, 471, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Katsumi, A.; Milanini, J.; Kiosses, W.B.; del Pozo, M.A.; Kaunas, R.; Chien, S.; Hahn, K.M.; Schwartz, M.A. Effects of cell tension on the small gtpase rac. J. Cell. Biol. 2002, 158, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Burridge, K.; Chrzanowska-Wodnicka, M. Focal adhesions, contractility, and signaling. Annu. Rev. Cell Dev. Biol. 1996, 12, 463–519. [Google Scholar] [CrossRef] [PubMed]
- Zaidel-Bar, R.; Cohen, M.; Addadi, L.; Geiger, B. Hierarchical assembly of cell–matrix adhesion complexes. Biochem. Soc. Trans. 2004, 32, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; Pan, Z.; Guan, E.; Ge, S.; Liu, Y.; Nakamura, T.; Ren, X.-D.; Rafailovich, M.; Clark, R.A.F. Cell adaptation to a physiologically relevant ecm mimic with different viscoelastic properties. Biomaterials 2007, 28, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Mih, J.D.; Marinkovic, A.; Liu, F.; Sharif, A.S.; Tschumperlin, D.J. Matrix stiffness reverses the effect of actomyosin tension on cell proliferation. J. Cell. Sci. 2012, 125, 5974–5983. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.; Jiang, H.; Ho, C.-H.; Grinnell, F. Microtubule function in fibroblast spreading is modulated according to the tension state of cell–matrix interactions. Proc. Natl. Acad. Sci. USA 2007, 104, 5425–5430. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J.; Hall, A. The small gtp-binding protein rho regulates the assembly of focal adhesions and actin stress fibers in response to growth factors. Cell 1992, 70, 389–399. [Google Scholar] [CrossRef]
- Hotulainen, P.; Lappalainen, P. Stress fibers are generated by two distinct actin assembly mechanisms in motile cells. J. Cell. Biol. 2006, 173, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Burnette, D.T.; Shao, L.; Ott, C.; Pasapera, A.M.; Fischer, R.S.; Baird, M.A.; Der Loughian, C.; Delanoe-Ayari, H.; Paszek, M.J.; Davidson, M.W.; et al. A contractile and counterbalancing adhesion system controls the 3d shape of crawling cells. J. Cell. Biol. 2014, 205, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Khatau, S.B.; Hale, C.M.; Stewart-Hutchinson, P.J.; Patel, M.S.; Stewart, C.L.; Searson, P.C.; Hodzic, D.; Wirtz, D. A perinuclear actin cap regulates nuclear shape. Proc. Natl. Acad. Sci. USA 2009, 106, 19017–19022. [Google Scholar] [CrossRef] [PubMed]
- Legate, K.R.; Wickström, S.A.; Fässler, R. Genetic and cell biological analysis of integrin outside-in signaling. Genes Dev. 2009, 23, 397–418. [Google Scholar] [CrossRef] [PubMed]
- Mason, B.; Califano, J.; Reinhart-King, C. Matrix stiffness: A regulator of cellular behavior and tissue formation. In Engineering biomaterials for regenerative medicine; Bhatia, S.K., Ed.; Springer: New York, NY, USA, 2012; pp. 19–37. [Google Scholar]
- Discher, D.E.; Janmey, P.; Wang, Y.-L. Tissue cells feel and respond to the stiffness of their substrate. Science 2005, 310, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Zaidel-Bar, R.; Itzkovitz, S.; Ma'ayan, A.; Iyengar, R.; Geiger, B. Functional atlas of the integrin adhesome. Nat. Cell. Biol. 2007, 9, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Zaidel-Bar, R.; Geiger, B. The switchable integrin adhesome. J. Cell. Sci. 2010, 123, 1385–1388. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.G.; Miyazu, M.; Matsushita, E.; Sokabe, M.; Naruse, K. Uniaxial cyclic stretch induces focal adhesion kinase (fak) tyrosine phosphorylation followed by mitogen-activated protein kinase (mapk) activation. Biochem. Biophys. Res. Commun. 2001, 288, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Moalli, M.R.; Wang, S.; Caldwell, N.J.; Patil, P.V.; Maynard, C.R. Mechanical stimulation induces pp125(fak) and pp60(src) activity in an in vivo model of trabecular bone formation. J. Appl. Physiol. 2001, 91, 912–918. [Google Scholar] [PubMed]
- Molina, T.; Kabsch, K.; Alonso, A.; Kohl, A.; Komposch, G.; Tomakidi, P. Topographic changes of focal adhesion components and modulation of p125fak activation in stretched human periodontal ligament fibroblasts. J. Dent. Res. 2001, 80, 1984–1989. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Millward-Sadler, S.J.; Wright, M.O.; Nuki, G.; Salter, D.M. Integrin and mechanosensitive ion channel-dependent tyrosine phosphorylation of focal adhesion proteins and beta-catenin in human articular chondrocytes after mechanical stimulation. J. Bone Miner. Res. 2000, 15, 1501–1509. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Guo, W.Y.; Li, W.J.; Zhang, D.X.; Lv, A.L.; Luan, R.H.; Liu, B.; Wang, H.C. Cyclic stretch upregulates sdf-1alpha/cxcr4 axis in human saphenous vein smooth muscle cells. Biochem. Biophys. Res. Commun. 2009, 386, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Fluck, M.; Carson, J.A.; Gordon, S.E.; Ziemiecki, A.; Booth, F.W. Focal adhesion proteins fak and paxillin increase in hypertrophied skeletal muscle. Am. J. Physiol. 1999, 277, C152–C162. [Google Scholar] [PubMed]
- Basson, M.D.; Yu, C.F.; Herden-Kirchoff, O.; Ellermeier, M.; Sanders, M.A.; Merrell, R.C.; Sumpio, B.E. Effects of increased ambient pressure on colon cancer cell adhesion. J. Cell. Biochem. 2000, 78, 47–61. [Google Scholar] [CrossRef]
- Provenzano, P.P.; Inman, D.R.; Eliceiri, K.W.; Keely, P.J. Matrix density-induced mechanoregulation of breast cell phenotype, signaling and gene expression through a fak-erk linkage. Oncogene 2009, 28, 4326–4343. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, P.P.; Keely, P.J. Mechanical signaling through the cytoskeleton regulates cell proliferation by coordinated focal adhesion and rho gtpase signaling. J. Cell. Sci. 2011, 124, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Cary, L.A.; Chang, J.F.; Guan, J.L. Stimulation of cell migration by overexpression of focal adhesion kinase and its association with Src and Fyn. J. Cell. Sci. 1996, 109, 1787–1794. [Google Scholar] [PubMed]
- Owen, J.D.; Ruest, P.J.; Fry, D.W.; Hanks, S.K. Induced focal adhesion kinase (fak) expression in fak-null cells enhances cell spreading and migration requiring both auto- and activation loop phosphorylation sites and inhibits adhesion-dependent tyrosine phosphorylation of pyk2. Mol. Cell. Biol. 1999, 19, 4806–4818. [Google Scholar] [CrossRef] [PubMed]
- Cary, L.A.; Han, D.C.; Polte, T.R.; Hanks, S.K.; Guan, J.L. Identification of p130cas as a mediator of focal adhesion kinase-promoted cell migration. J. Cell. Biol. 1998, 140, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Han, D.C.; Shen, T.L.; Guan, J.L. Role of grb7 targeting to focal contacts and its phosphorylation by focal adhesion kinase in regulation of cell migration. J. Biol. Chem. 2000, 275, 28911–28917. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.L.; Han, D.C.; Guan, J.L. Association of grb7 with phosphoinositides and its role in the regulation of cell migration. J. Biol. Chem. 2002, 277, 29069–29077. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.D.; Kiosses, W.B.; Sieg, D.J.; Otey, C.A.; Schlaepfer, D.D.; Schwartz, M.A. Focal adhesion kinase suppresses rho activity to promote focal adhesion turnover. J. Cell. Sci. 2000, 113, 3673–3678. [Google Scholar] [PubMed]
- Chen, B.H.; Tzen, J.T.; Bresnick, A.R.; Chen, H.C. Roles of rho-associated kinase and myosin light chain kinase in morphological and migratory defects of focal adhesion kinase-null cells. J. Biol. Chem. 2002, 277, 33857–33863. [Google Scholar] [CrossRef] [PubMed]
- Hsia, D.A.; Mitra, S.K.; Hauck, C.R.; Streblow, D.N.; Nelson, J.A.; Ilic, D.; Huang, S.; Li, E.; Nemerow, G.R.; Leng, J.; et al. Differential regulation of cell motility and invasion by fak. J. Cell. Biol. 2003, 160, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Suetsugu, S.; Cooper, L.A.; Takenawa, T.; Guan, J.L. Focal adhesion kinase regulation of n-wasp subcellular localization and function. J. Biol. Chem. 2004, 279, 9565–9576. [Google Scholar] [CrossRef] [PubMed]
- Clemente, C.F.M.Z.; Corat, M.A.F.; Saad, S.T.O.; Franchini, K.G. Differentiation of c2c12 myoblasts is critically regulated by fak signaling. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R862–R870. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Song, G.; Ju, Y.; Li, X.; Song, Y.; Watanabe, S. Rhoa/rock, cytoskeletal dynamics, and focal adhesion kinase are required for mechanical stretch-induced tenogenic differentiation of human mesenchymal stem cells. J. Cell. Physiol. 2012, 227, 2722–2729. [Google Scholar] [CrossRef] [PubMed]
- Salasznyk, R.M.; Klees, R.F.; Williams, W.A.; Boskey, A.; Plopper, G.E. Focal adhesion kinase signaling pathways regulate the osteogenic differentiation of human mesenchymal stem cells. Exp. Cell. Res. 2007, 313, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Shih, Y.R.; Tseng, K.F.; Lai, H.Y.; Lin, C.H.; Lee, O.K. Matrix stiffness regulation of integrin-mediated mechanotransduction during osteogenic differentiation of human mesenchymal stem cells. J. Bone Miner. Res. 2011, 26, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; He, J.; Li, Y.-S.; Wang, X.-M.; Ai, H.-J.; Cui, F.-Z. Activation of the erk1/2 signaling pathway during the osteogenic differentiation of mesenchymal stem cells cultured on substrates modified with various chemical groups. BioMed Res. Inter. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Chrzanowska-Wodnicka, M.; Burridge, K. Rho-stimulated contractility drives the formation of stress fibers and focal adhesions. J. Cell. Biol. 1996, 133, 1403–1415. [Google Scholar] [CrossRef] [PubMed]
- Riveline, D.; Zamir, E.; Balaban, N.Q.; Schwarz, U.S.; Ishizaki, T.; Narumiya, S.; Kam, Z.; Geiger, B.; Bershadsky, A.D. Focal contacts as mechanosensors: Externally applied local mechanical force induces growth of focal contacts by an mdia1-dependent and rock-independent mechanism. J. Cell. Biol. 2001, 153, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- del Rio, A.; Perez-Jimenez, R.; Liu, R.; Roca-Cusachs, P.; Fernandez, J.M.; Sheetz, M.P. Stretching single talin rod molecules activates vinculin binding. Science 2009, 323, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Sawada, Y.; Tamada, M.; Dubin-Thaler, B.J.; Cherniavskaya, O.; Sakai, R.; Tanaka, S.; Sheetz, M.P. Force sensing by mechanical extension of the src family kinase substrate p130cas. Cell 2006, 127, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Albuschies, J.; Vogel, V. The role of filopodia in the recognition of nanotopographies. Sci. Rep. 2013, 3, 1658. [Google Scholar] [CrossRef] [PubMed]
- Weiss, P. Experiments on cell and axon orientation in vitro; the role of colloidal exudates in tissue organization. J. Exp. Zool. 1945, 100, 353–386. [Google Scholar] [CrossRef] [PubMed]
- Ventre, M.; Natale, C.F.; Rianna, C.; Netti, P.A. Topographic cell instructive patterns to control cell adhesion, polarization and migration. J. R. Soc. Interface 2014, 11, 20140687. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.I.; Abrams, G.A.; Bertics, P.J.; Murphy, C.J.; Nealey, P.F. Epithelial contact guidance on well-defined micro- and nanostructured substrates. J. Cell. Sci. 2003, 116, 1881–1892. [Google Scholar] [CrossRef] [PubMed]
- Nemethova, M.; Auinger, S.; Small, J.V. Building the actin cytoskeleton: Filopodia contribute to the construction of contractile bundles in the lamella. J. Cell. Biol. 2008, 180, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Yim, E.K.F.; Darling, E.M.; Kulangara, K.; Guilak, F.; Leong, K.W. Nanotopography-induced changes in focal adhesions, cytoskeletal organization, and mechanical properties of human mesenchymal stem cells. Biomaterials 2010, 31, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Galbraith, C.G.; Yamada, K.M.; Galbraith, J.A. Polymerizing actin fibers position integrins primed to probe for adhesion sites. Science 2007, 315, 992–995. [Google Scholar] [CrossRef] [PubMed]
- Oakley, C.; Brunette, D.M. The sequence of alignment of microtubules, focal contacts and actin filaments in fibroblasts spreading on smooth and grooved titanium substrata. J. Cell. Sci. 1993, 106, 343–354. [Google Scholar] [PubMed]
- You, R.; Li, X.; Liu, Y.; Liu, G.; Lu, S.; Li, M. Response of filopodia and lamellipodia to surface topography on micropatterned silk fibroin films. J. Biomed. Mater. Res. A 2014, 102, 4206–4212. [Google Scholar] [CrossRef] [PubMed]
- You, R.; Li, X.; Luo, Z.; Qu, J.; Li, M. Directional cell elongation through filopodia-steered lamellipodial extension on patterned silk fibroin films. Biointerphases 2015, 10, 011005. [Google Scholar] [CrossRef] [PubMed]
- Geiger, B.; Spatz, J.P.; Bershadsky, A.D. Environmental sensing through focal adhesions. Nat. Rev. Mol. Cell. Biol. 2009, 10, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Kato, T.; Fujita, A.; Ishizaki, T.; Narumiya, S. Cooperation between mdia1 and rock in rho-induced actin reorganization. Nat. Cell. Biol. 1999, 1, 136–143. [Google Scholar] [PubMed]
- Burridge, K.; Wennerberg, K. Rho and rac take center stage. Cell 2004, 116, 167–179. [Google Scholar] [CrossRef]
- Pelham, R.J.; Wang, Y.-L. Cell locomotion and focal adhesions are regulated by substrate flexibility. Proc. Natl. Acad. Sci. USA 1997, 94, 13661–13665. [Google Scholar] [CrossRef] [PubMed]
- Califano, J.P.; Reinhart-King, C.A. Substrate stiffness and cell area predict cellular traction stresses in single cells and cells in contact. Cell. Mol. Bioeng. 2010, 3, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Ghibaudo, M.; Saez, A.; Trichet, L.; Xayaphoummine, A.; Browaeys, J.; Silberzan, P.; Buguin, A.; Ladoux, B. Traction forces and rigidity sensing regulate cell functions. Soft Matter 2008, 4, 1836–1843. [Google Scholar] [CrossRef]
- Lo, C.-M.; Wang, H.-B.; Dembo, M.; Wang, Y.-L. Cell movement is guided by the rigidity of the substrate. Biophys. J. 2000, 79, 144–152. [Google Scholar] [CrossRef]
- Isenberg, B.C.; DiMilla, P.A.; Walker, M.; Kim, S.; Wong, J.Y. Vascular smooth muscle cell durotaxis depends on substrate stiffness gradient strength. Biophys. J. 2009, 97, 1313–1322. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Lin, J.S. Cell traction force and measurement methods. Biomech. Model. Mechanobiol. 2007, 6, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Dembo, M.; Wang, Y.L. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophys. J. 1999, 76, 2307–2316. [Google Scholar] [CrossRef]
- Pelham, R.J.; Wang, Y.-L. High resolution detection of mechanical forces exerted by locomoting fibroblasts on the substrate. Mol. Biol. Cell. 1999, 10, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.H.; Mui, K.L.; Hsu, B.Y.; Liu, S.-L.; Cretu, A.; Razinia, Z.; Xu, T.; Puré, E.; Assoian, R.K. A fak-cas-rac-lamellipodin signaling module transduces extracellular matrix stiffness into mechanosensitive cell cycling. Sci. Signal. 2014, 7, ra57. [Google Scholar] [CrossRef] [PubMed]
- Pasapera, A.M.; Plotnikov, S.V.; Fischer, R.S.; Case, L.B.; Egelhoff, T.T.; Waterman, C.M. Rac1-dependent phosphorylation and focal adhesion recruitment of myosin iia regulates migration and mechanosensing. Curr. Biol. 2015, 25, 175–186. [Google Scholar] [CrossRef] [PubMed]
- LaLonde, D.P.; Grubinger, M.; Lamarche-Vane, N.; Turner, C.E. Cdgap associates with actopaxin to regulate integrin-dependent changes in cell morphology and motility. Curr. Biol. 2006, 16, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Nikolopoulos, S.N.; Turner, C.E. Molecular dissection of actopaxin-integrin-linked kinase-paxillin interactions and their role in subcellular localization. J. Biol. Chem. 2002, 277, 1568–1575. [Google Scholar] [CrossRef] [PubMed]
- Nikolopoulos, S.N.; Turner, C.E. Actopaxin, a new focal adhesion protein that binds paxillin ld motifs and actin and regulates cell adhesion. J. Cell. Biol. 2000, 151, 1435–1448. [Google Scholar] [CrossRef] [PubMed]
- Wormer, D.B.; Davis, K.A.; Henderson, J.H.; Turner, C.E. The focal adhesion-localized cdgap regulates matrix rigidity sensing and durotaxis. PLoS ONE 2014, 9, e91815. [Google Scholar]
- Peyton, S.R.; Putnam, A.J. Extracellular matrix rigidity governs smooth muscle cell motility in a biphasic fashion. J. Cell. Physiol. 2005, 204, 198–209. [Google Scholar] [CrossRef] [PubMed]
- DiMilla, P.A.; Stone, J.A.; Quinn, J.A.; Albelda, S.M.; Lauffenburger, D.A. Maximal migration of human smooth muscle cells on fibronectin and type iv collagen occurs at an intermediate attachment strength. J. Cell. Biol. 1993, 122, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Calve, S.; Simon, H.-G. Biochemical and mechanical environment cooperatively regulate skeletal muscle regeneration. FASEB J. 2012, 26, 2538–2545. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-Y.; Wu, C.-H.; Wang, M.-H.; Chen, B.-S.; Chiou, L.-L.; Lee, H.-S. Cooperative regulation of substrate stiffness and extracellular matrix proteins in skin wound healing of axolotls. BioMed Res. Inter. 2015, 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Lautscham, L.A.; Kammerer, C.; Lange, J.R.; Kolb, T.; Mark, C.; Schilling, A.; Strissel, P.L.; Strick, R.; Gluth, C.; Rowat, A.C.; et al. Migration in confined 3d environments is determined by a combination of adhesiveness, nuclear volume, contractility, and cell stiffness. Biophys. J. 2015, 109, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Pathak, A.; Kumar, S. Independent regulation of tumor cell migration by matrix stiffness and confinement. Proc. Natl. Acad. Sci. USA 2012, 109, 10334–10339. [Google Scholar] [CrossRef] [PubMed]
- Riching, K.M.; Keely, P.J. Rho family gtpases: Making it to the third dimension. Int. J. Biochem. Cell. Biol. 2015, 59, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Worthylake, R.A.; Burridge, K. Rhoa and rock promote migration by limiting membrane protrusions. J. Biol. Chem. 2003, 278, 13578–13584. [Google Scholar] [CrossRef] [PubMed]
- Pankov, R.; Endo, Y.; Even-Ram, S.; Araki, M.; Clark, K.; Cukierman, E.; Matsumoto, K.; Yamada, K.M. A rac switch regulates random versus directionally persistent cell migration. J. Cell. Biol. 2005, 170, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Diehl, K.A.; Foley, J.D.; Nealey, P.F.; Murphy, C.J. Nanoscale topography modulates corneal epithelial cell migration. J. Biomed. Mater. Res. A 2005, 75, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Yim, E.K.; Reano, R.M.; Pang, S.W.; Yee, A.F.; Chen, C.S.; Leong, K.W. Nanopattern-induced changes in morphology and motility of smooth muscle cells. Biomaterials 2005, 26, 5405–5413. [Google Scholar] [CrossRef] [PubMed]
- Biela, S.A.; Su, Y.; Spatz, J.P.; Kemkemer, R. Different sensitivity of human endothelial cells, smooth muscle cells and fibroblasts to topography in the nano-micro range. Acta. Biomater. 2009, 5, 2460–2466. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Driscoll, M.K.; Guven, C.; Das, S.; Parent, C.A.; Fourkas, J.T.; Losert, W. Asymmetric nanotopography biases cytoskeletal dynamics and promotes unidirectional cell guidance. Proc. Natl. Acad. Sci. USA 2015, 112, 12557–12562. [Google Scholar] [CrossRef] [PubMed]
- Tozluoglu, M.; Tournier, A.L.; Jenkins, R.P.; Hooper, S.; Bates, P.A.; Sahai, E. Matrix geometry determines optimal cancer cell migration strategy and modulates response to interventions. Nat. Cell. Biol. 2013, 15, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.; Te Lindert, M.; Krause, M.; Alexander, S.; Te Riet, J.; Willis, A.L.; Hoffman, R.M.; Figdor, C.G.; Weiss, S.J.; Friedl, P. Physical limits of cell migration: Control by ecm space and nuclear deformation and tuning by proteolysis and traction force. J. Cell. Biol. 2013, 201, 1069–1084. [Google Scholar] [CrossRef] [PubMed]
- Legant, W.R.; Choi, C.K.; Miller, J.S.; Shao, L.; Gao, L.; Betzig, E.; Chen, C.S. Multidimensional traction force microscopy reveals out-of-plane rotational moments about focal adhesions. Proc. Natl. Acad. Sci. USA 2013, 110, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Style, R.W.; Boltyanskiy, R.; German, G.K.; Hyland, C.; MacMinn, C.W.; Mertz, A.F.; Wilen, L.A.; Xu, Y.; Dufresne, E.R. Traction force microscopy in physics and biology. Soft Matter 2014, 10, 4047–4055. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ohlin, C.A.; Lu, Q.; Hu, J. Cell directional migration and oriented division on three-dimensional laser-induced periodic surface structures on polystyrene. Biomaterials 2008, 29, 2049–2059. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Provenzano, P.P.; Smith, C.L.; Levchenko, A. Matrix nanotopography as a regulator of cell function. J. Cell. Biol. 2012, 197, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Han, K.; Gupta, K.; Kwon, K.W.; Suh, K.Y.; Levchenko, A. Mechanosensitivity of fibroblast cell shape and movement to anisotropic substratum topography gradients. Biomaterials 2009, 30, 5433–5444. [Google Scholar] [CrossRef] [PubMed]
- Rajnicek, A.; McCaig, C. Guidance of cns growth cones by substratum grooves and ridges: Effects of inhibitors of the cytoskeleton, calcium channels and signal transduction pathways. J. Cell. Sci. 1997, 110, 2915–2924. [Google Scholar] [PubMed]
- Karuri, N.W.; Liliensiek, S.; Teixeira, A.I.; Abrams, G.; Campbell, S.; Nealey, P.F.; Murphy, C.J. Biological length scale topography enhances cell-substratum adhesion of human corneal epithelial cells. J. Cell. Sci. 2004, 117, 3153–3164. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.I.; Nealey, P.F.; Murphy, C.J. Responses of human keratocytes to micro- and nanostructured substrates. J. Biomed. Mater. Res. A 2004, 71, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Ranucci, C.S.; Moghe, P.V. Substrate microtopography can enhance cell adhesive and migratory responsiveness to matrix ligand density. J. Biomed. Mater. Res. 2001, 54, 149–161. [Google Scholar] [CrossRef]
- Mello, A.P.; Volkov, Y.; Kelleher, D.; Prendergast, P.J. Comparative locomotory behavior of t lymphocytes versus t lymphoma cells on flat and grooved surfaces. Ann. Biomed. Eng. 2003, 31, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Brammer, K.S.; Oh, S.; Gallagher, J.O.; Jin, S. Enhanced cellular mobility guided by tio2 nanotube surfaces. Nano Lett. 2008, 8, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Saltzman, W.M. Topographical control of human neutrophil motility on micropatterned materials with various surface chemistry. Biomaterials 2002, 23, 3215–3225. [Google Scholar] [CrossRef]
- Liliensiek, S.J.; Wood, J.A.; Yong, J.; Auerbach, R.; Nealey, P.F.; Murphy, C.J. Modulation of human vascular endothelial cell behaviors by nanotopographic cues. Biomaterials 2010, 31, 5418–5426. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Cavalcanti-Adam, E.A.; Glass, R.; Blummel, J.; Eck, W.; Kantlehner, M.; Kessler, H.; Spatz, J.P. Activation of integrin function by nanopatterned adhesive interfaces. Chemphyschem 2004, 5, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti-Adam, E.A.; Volberg, T.; Micoulet, A.; Kessler, H.; Geiger, B.; Spatz, J.P. Cell spreading and focal adhesion dynamics are regulated by spacing of integrin ligands. Biophys. J. 2007, 92, 2964–2974. [Google Scholar] [CrossRef] [PubMed]
- Frey, M.T.; Tsai, I.Y.; Russell, T.P.; Hanks, S.K.; Wang, Y.-L. Cellular responses to substrate topography: Role of myosin ii and focal adhesion kinase. Biophys. J. 2006, 90, 3774–3782. [Google Scholar] [CrossRef] [PubMed]
- Ghibaudo, M.; Trichet, L.; Le Digabel, J.; Richert, A.; Hersen, P.; Ladoux, B. Substrate topography induces a crossover from 2d to 3d behavior in fibroblast migration. Biophys. J. 2009, 97, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Assoian, R.K.; Klein, E.A. Growth control by intracellular tension and extracellular stiffness. Trends Cell Biol. 2008, 18, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Reverte, C.G.; Benware, A.; Jones, C.W.; LaFlamme, S.E. Perturbing integrin function inhibits microtubule growth from centrosomes, spindle assembly, and cytokinesis. J. Cell Biol. 2006, 174, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Klein, E.A.; Yin, L.; Kothapalli, D.; Castagnino, P.; Byfield, F.J.; Xu, T.; Levental, I.; Hawthorne, E.; Janmey, P.A.; Assoian, R.K. Cell-cycle control by physiological matrix elasticity and in vivo tissue stiffening. Curr. Biol. 2009, 19, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, T.A.; de Juan Pardo, E.M.; Kumar, S. The mechanical rigidity of the extracellular matrix regulates the structure, motility, and proliferation of glioma cells. Cancer Res. 2009, 69, 4167–4174. [Google Scholar] [CrossRef] [PubMed]
- Oktay, M.; Wary, K.K.; Dans, M.; Birge, R.B.; Giancotti, F.G. Integrin-mediated activation of focal adhesion kinase is required for signaling to jun nh2-terminal kinase and progression through the g1 phase of the cell cycle. J. Cell Biol. 1999, 145, 1461–1470. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Bergami, P.; Huang, C.; Goydos, J.S.; Yip, D.; Bar-Eli, M.; Herlyn, M.; Smalley, K.S.M.; Mahale, A.; Eroshkin, A.; Aaronson, S.; et al. Rewired erk-jnk signaling pathways in melanoma. Cancer Cell. 2007, 11, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Bartek, J.; Bartkova, J.; Lukas, J. The retinoblastoma protein pathway and the restriction point. Curr. Opin. Cell. Biol. 1996, 8, 805–814. [Google Scholar] [CrossRef]
- Riento, K.; Ridley, A.J. Rocks: Multifunctional kinases in cell behaviour. Nat. Rev. Mol. Cell. Biol. 2003, 4, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, M.; Ishizaki, T.; Boku, S.; Watanabe, N.; Fujita, A.; Iwamatsu, A.; Obinata, T.; Ohashi, K.; Mizuno, K.; Narumiya, S. Signaling from rho to the actin cytoskeleton through protein kinases rock and lim-kinase. Science 1999, 285, 895–898. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, X.R.; Riddick, N.; Bryden, M.; Baur, W.; Zhang, X.; Surks, H.K. Rock isoform regulation of myosin phosphatase and contractility in vascular smooth muscle cells. Circ. Res. 2009, 104, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.S.; Wynne, J.; Treisman, R. The rho family gtpases rhoa, rac1, and cdc42hs regulate transcriptional activation by srf. Cell 1995, 81, 1159–1170. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, D.Z.; Hockemeyer, D.; McAnally, J.; Nordheim, A.; Olson, E.N. Myocardin and ternary complex factors compete for srf to control smooth muscle gene expression. Nature 2004, 428, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.N.; Nordheim, A. Linking actin dynamics and gene transcription to drive cellular motile functions. Nat. Rev. Mol. Cell. Biol. 2010, 11, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J.; Moscona, A. Role of cell shape in growth control. Nature 1978, 273, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.S.; Mrksich, M.; Huang, S.; Whitesides, G.M.; Ingber, D.E. Geometric control of cell life and death. Science 1997, 276, 1425–1428. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Chen, C.S.; Ingber, D.E. Control of cyclin d1, p27kip1, and cell cycle progression in human capillary endothelial cells by cell shape and cytoskeletal tension. Mol. Biol. Cell 1998, 9, 3179–3193. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of yap/taz in mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Itoga, K.; Okano, T.; Yonemura, S.; Sasaki, H. Hippo pathway regulation by cell morphology and stress fibers. Development 2011, 138, 3907–3914. [Google Scholar] [CrossRef] [PubMed]
- Ghrebi, S.; Hamilton, D.W.; Douglas Waterfield, J.; Brunette, D.M. The effect of surface topography on cell shape and early erk1/2 signaling in macrophages; linkage with fak and src. J. Biomed. Mater. Res. A 2013, 101, 2118–2128. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, M.A.; Chen, C.S. Mechanotransduction in development: A growing role for contractility. Nat. Rev. Mol. Cell. Biol. 2009, 10, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Bettinger, C.J.; Zhang, Z.; Gerecht, S.; Borenstein, J.T.; Langer, R. Enhancement of in vitro capillary tube formation by substrate nanotopography. Adv. Mater. 2008, 20, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Gerecht, S.; Bettinger, C.J.; Zhang, Z.; Borenstein, J.T.; Vunjak-Novakovic, G.; Langer, R. The effect of actin disrupting agents on contact guidance of human embryonic stem cells. Biomaterials 2007, 28, 4068–4077. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, S.; Masako, N.; Lee, D.J.; Cooper, L.F.; Ko, C.C. Proliferation of preosteoblasts on tio nanotubes is fak/rhoa related. RSC Adv. 2015, 5, 38117–38124. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tuft, B.W.; Xu, L.; Polacco, M.A.; Clarke, J.C.; Guymon, C.A.; Hansen, M.R. Microtopographical features generated by photopolymerization recruit rhoa/rock through trpv1 to direct cell and neurite growth. Biomaterials 2015, 53, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Milner, K.R.; Siedlecki, C.A. Submicron poly(l-lactic acid) pillars affect fibroblast adhesion and proliferation. J. Biomed. Mater. Res. A 2007, 82, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Washburn, N.R.; Yamada, K.M.; Simon, C.G., Jr.; Kennedy, S.B.; Amis, E.J. High-throughput investigation of osteoblast response to polymer crystallinity: Influence of nanometer-scale roughness on proliferation. Biomaterials 2004, 25, 1215–1224. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, V.; Maiorano, G.; Rizzello, L.; Sorce, B.; Sabella, S.; Cingolani, R.; Pompa, P.P. Neurons sense nanoscale roughness with nanometer sensitivity. Proc. Natl. Acad. Sci. USA 2010, 107, 6264–6269. [Google Scholar] [CrossRef] [PubMed]
- Ohlstein, B.; Kai, T.; Decotto, E.; Spradling, A. The stem cell niche: Theme and variations. Curr. Opin. Cell. Biol. 2004, 16, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Rowlands, A.S.; George, P.A.; Cooper-White, J.J. Directing osteogenic and myogenic differentiation of mscs: Interplay of stiffness and adhesive ligand presentation. Am. J. Physiol. Cell Physiol. 2008, 295, C1037–C1044. [Google Scholar] [CrossRef] [PubMed]
- Colley, H.E.; Mishra, G.; Scutt, A.M.; McArthur, S.L. Plasma polymer coatings to support mesenchymal stem cell adhesion, growth and differentiation on variable stiffness silicone elastomers. Plasma Processes Polym. 2009, 6, 831–839. [Google Scholar] [CrossRef]
- Lanniel, M.; Huq, E.; Allen, S.; Buttery, L.; Williams, P.M.; Alexander, M.R. Substrate induced differentiation of human mesenchymal stem cells on hydrogels with modified surface chemistry and controlled modulus. Soft Matter 2011, 7, 6501–6514. [Google Scholar] [CrossRef]
- Wang, L.S.; Boulaire, J.; Chan, P.P.; Chung, J.E.; Kurisawa, M. The role of stiffness of gelatin-hydroxyphenylpropionic acid hydrogels formed by enzyme-mediated crosslinking on the differentiation of human mesenchymal stem cell. Biomaterials 2010, 31, 8608–8616. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Vincent, L.G.; Lee, A.R.; Dobke, M.K.; Engler, A.J. Mechanical derivation of functional myotubes from adipose-derived stem cells. Biomaterials 2012, 33, 2482–2491. [Google Scholar] [CrossRef] [PubMed]
- Young, D.A.; Choi, Y.S.; Engler, A.J.; Christman, K.L. Stimulation of adipogenesis of adult adipose-derived stem cells using substrates that mimic the stiffness of adipose tissue. Biomaterials 2013, 34, 8581–8588. [Google Scholar] [CrossRef] [PubMed]
- Lui, C.; Lee, K.; Nelson, C.M. Matrix compliance and rhoa direct the differentiation of mammary progenitor cells. Biomech. Model. Mechanobiol. 2012, 11, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Saha, K.; Keung, A.J.; Irwin, E.F.; Li, Y.; Little, L.; Schaffer, D.V.; Healy, K.E. Substrate modulus directs neural stem cell behavior. Biophys. J. 2008, 95, 4426–4438. [Google Scholar] [CrossRef] [PubMed]
- Keung, A.J.; de Juan-Pardo, E.M.; Schaffer, D.V.; Kumar, S. Rho gtpases mediate the mechanosensitive lineage commitment of neural stem cells. Stem Cells 2011, 29, 1886–1897. [Google Scholar] [CrossRef] [PubMed]
- Leipzig, N.D.; Shoichet, M.S. The effect of substrate stiffness on adult neural stem cell behavior. Biomaterials 2009, 30, 6867–6878. [Google Scholar] [CrossRef] [PubMed]
- Evans, N.D.; Minelli, C.; Gentleman, E.; LaPointe, V.; Patankar, S.N.; Kallivretaki, M.; Chen, X.; Roberts, C.J.; Stevens, M.M. Substrate stiffness affects early differentiation events in embryonic stem cells. Eur. Cell. Mater. 2009, 18, e13. [Google Scholar] [CrossRef]
- Kimura, K.; Ito, M.; Amano, M.; Chihara, K.; Fukata, Y.; Nakafuku, M.; Yamamori, B.; Feng, J.; Nakano, T.; Okawa, K.; et al. Regulation of myosin phosphatase by rho and rho-associated kinase (rho-kinase). Science 1996, 273, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.A.; Even-Ram, S.; Liu, C.; Yamada, K.M.; Adelstein, R.S. Defects in cell adhesion and the visceral endoderm following ablation of nonmuscle myosin heavy chain ii-a in mice. J. Biol. Chem. 2004, 279, 41263–41266. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-F.; Cheng, S.-L. Αvβ integrins play an essential role in bmp-2 induction of osteoblast differentiation. J. Bone Miner. Res. 2005, 20, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-K.; Yu, X.; Cohen, D.M.; Wozniak, M.A.; Yang, M.T.; Gao, L.; Eyckmans, J.; Chen, C.S. Bone morphogenetic protein-2-induced signaling and osteogenesis is regulated by cell shape, rhoa/rock, and cytoskeletal tension. Stem Cells Dev. 2012, 21, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Chen, X.; Liang, X.; Zhang, G.; Xu, J.; He, L.; Zhan, Q.; Feng, X.-Q.; Chien, S.; Yang, C. Integrin activation and internalization on soft ecm as a mechanism of induction of stem cell differentiation by ecm elasticity. Proc. Natl. Acad. Sci. USA 2011, 108, 9466–9471. [Google Scholar] [CrossRef] [PubMed]
- Arnsdorf, E.J.; Tummala, P.; Kwon, R.Y.; Jacobs, C.R. Mechanically induced osteogenic differentiation--the role of rhoa, rockii and cytoskeletal dynamics. J. Cell. Sci. 2009, 122, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Bryan, B.A.; Mitchell, D.C.; Zhao, L.; Ma, W.; Stafford, L.J.; Teng, B.-B.; Liu, M. Modulation of muscle regeneration, myogenesis, and adipogenesis by the rho family guanine nucleotide exchange factor geft. Mol. Cell. Biol. 2005, 25, 11089–11101. [Google Scholar] [CrossRef] [PubMed]
- Horii, T.; Morita, S.; Kimura, M.; Hatada, I. Epigenetic regulation of adipocyte differentiation by a rho guanine nucleotide exchange factor, wgef. PLoS ONE 2009, 4, e5809. [Google Scholar] [CrossRef] [PubMed]
- Haudenschild, D.R.; Chen, J.; Pang, N.; Lotz, M.K.; D’Lima, D.D. Rho kinase-dependent sox9 activation in chondrocytes. Arthritis Rheum. 2010, 62, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.; Wang, G.; Beier, F. Rhoa/rock signaling regulates sox9 expression and actin organization during chondrogenesis. J. Biol. Chem. 2005, 280, 11626–11634. [Google Scholar] [CrossRef] [PubMed]
- Jungmann, P.M.; Mehlhorn, A.T.; Schmal, H.; Schillers, H.; Oberleithner, H.; Sudkamp, N.P. Nanomechanics of human adipose-derived stem cells: Small gtpases impact chondrogenic differentiation. Tissue Eng. Part A 2012, 18, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Biggs, M.J.; Richards, R.G.; Gadegaard, N.; Wilkinson, C.D.; Dalby, M.J. The effects of nanoscale pits on primary human osteoblast adhesion formation and cellular spreading. J. Mater. Sci. Mater. Med. 2007, 18, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Biggs, M.J.; Richards, R.G.; McFarlane, S.; Wilkinson, C.D.; Oreffo, R.O.; Dalby, M.J. Adhesion formation of primary human osteoblasts and the functional response of mesenchymal stem cells to 330nm deep microgrooves. J. R Soc. Interface 2008, 5, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Dalby, M.J.; Gadegaard, N.; Tare, R.; Andar, A.; Riehle, M.O.; Herzyk, P.; Wilkinson, C.D.; Oreffo, R.O. The control of human mesenchymal cell differentiation using nanoscale symmetry and disorder. Nat. Mater. 2007, 6, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Galli, C.; Piemontese, M.; Lumetti, S.; Ravanetti, F.; Macaluso, G.M.; Passeri, G. Actin cytoskeleton controls activation of wnt/beta-catenin signaling in mesenchymal cells on implant surfaces with different topographies. Acta Biomater. 2012, 8, 2963–2968. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wen, F.; Wong, Y.S.; Boey, F.Y.; Subbu, V.S.; Leong, D.T.; Ng, K.W.; Ng, G.K.; Tan, L.P. Direct laser machining-induced topographic pattern promotes up-regulation of myogenic markers in human mesenchymal stem cells. Acta Biomater. 2012, 8, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Lumetti, S.; Mazzotta, S.; Ferrillo, S.; Piergianni, M.; Piemontese, M.; Passeri, G.; Macaluso, G.M.; Galli, C. Rhoa controls wnt upregulation on microstructured titanium surfaces. Biomed. Res. Int. 2014, 2014, 401859. [Google Scholar] [CrossRef] [PubMed]
- Seo, C.H.; Furukawa, K.; Montagne, K.; Jeong, H.; Ushida, T. The effect of substrate microtopography on focal adhesion maturation and actin organization via the rhoa/rock pathway. Biomaterials 2011, 32, 9568–9575. [Google Scholar] [CrossRef] [PubMed]
- Chakravorty, N.; Hamlet, S.; Jaiprakash, A.; Crawford, R.; Oloyede, A.; Alfarsi, M.; Xiao, Y.; Ivanovski, S. Pro-osteogenic topographical cues promote early activation of osteoprogenitor differentiation via enhanced tgfβ, wnt, and notch signaling. Clin. Oral Implants Res. 2014, 25, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Navarrete, R.; Hyzy, S.; Wieland, M.; Boyan, B.D.; Schwartz, Z. The roles of wnt signaling modulators dickkopf-1 (dkk1) and dickkopf-2 (dkk2) and cell maturation state in osteogenesis on microstructured titanium surfaces. Biomaterials 2010, 31, 2015–2024. [Google Scholar] [CrossRef] [PubMed]
- Galli, C.; Passeri, G.; Ravanetti, F.; Elezi, E.; Pedrazzoni, M.; Macaluso, G.M. Rough surface topography enhances the activation of wnt/beta-catenin signaling in mesenchymal cells. J. Biomed. Mater. Res. A 2010, 95, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Ogino, Y.; Liang, R.; Mendonca, D.B.; Mendonca, G.; Nagasawa, M.; Koyano, K.; Cooper, L.F. Rhoa-mediated functions in c3h10t1/2 osteoprogenitors are substrate topography-dependent. J. Cell. Physiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Lyon, C.A.; Koutsouki, E.; Aguilera, C.M.; Blaschuk, O.W.; George, S.J. Inhibition of n-cadherin retards smooth muscle cell migration and intimal thickening via induction of apoptosis. J. Vasc. Surg. 2010, 52, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Prowse, P.D.H.; Elliott, C.G.; Hutter, J.; Hamilton, D.W. Inhibition of rac and rock signalling influence osteoblast adhesion, differentiation and mineralization on titanium topographies. PLoS ONE 2013, 8, e58898. [Google Scholar]
- Yamazoe, T.; Shiraki, N.; Toyoda, M.; Kiyokawa, N.; Okita, H.; Miyagawa, Y.; Akutsu, H.; Umezawa, A.; Sasaki, Y.; Kume, K.; et al. A synthetic nanofibrillar matrix promotes in vitro hepatic differentiation of embryonic stem cells and induced pluripotent stem cells. J. Cell. Sci. 2013, 126, 5391–5399. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoon, J.L.; Tan, M.H.; Koh, C.-G. The Regulation of Cellular Responses to Mechanical Cues by Rho GTPases. Cells 2016, 5, 17. https://doi.org/10.3390/cells5020017
Hoon JL, Tan MH, Koh C-G. The Regulation of Cellular Responses to Mechanical Cues by Rho GTPases. Cells. 2016; 5(2):17. https://doi.org/10.3390/cells5020017
Chicago/Turabian StyleHoon, Jing Ling, Mei Hua Tan, and Cheng-Gee Koh. 2016. "The Regulation of Cellular Responses to Mechanical Cues by Rho GTPases" Cells 5, no. 2: 17. https://doi.org/10.3390/cells5020017