
Vimentin in Bacterial Infections


Abstract
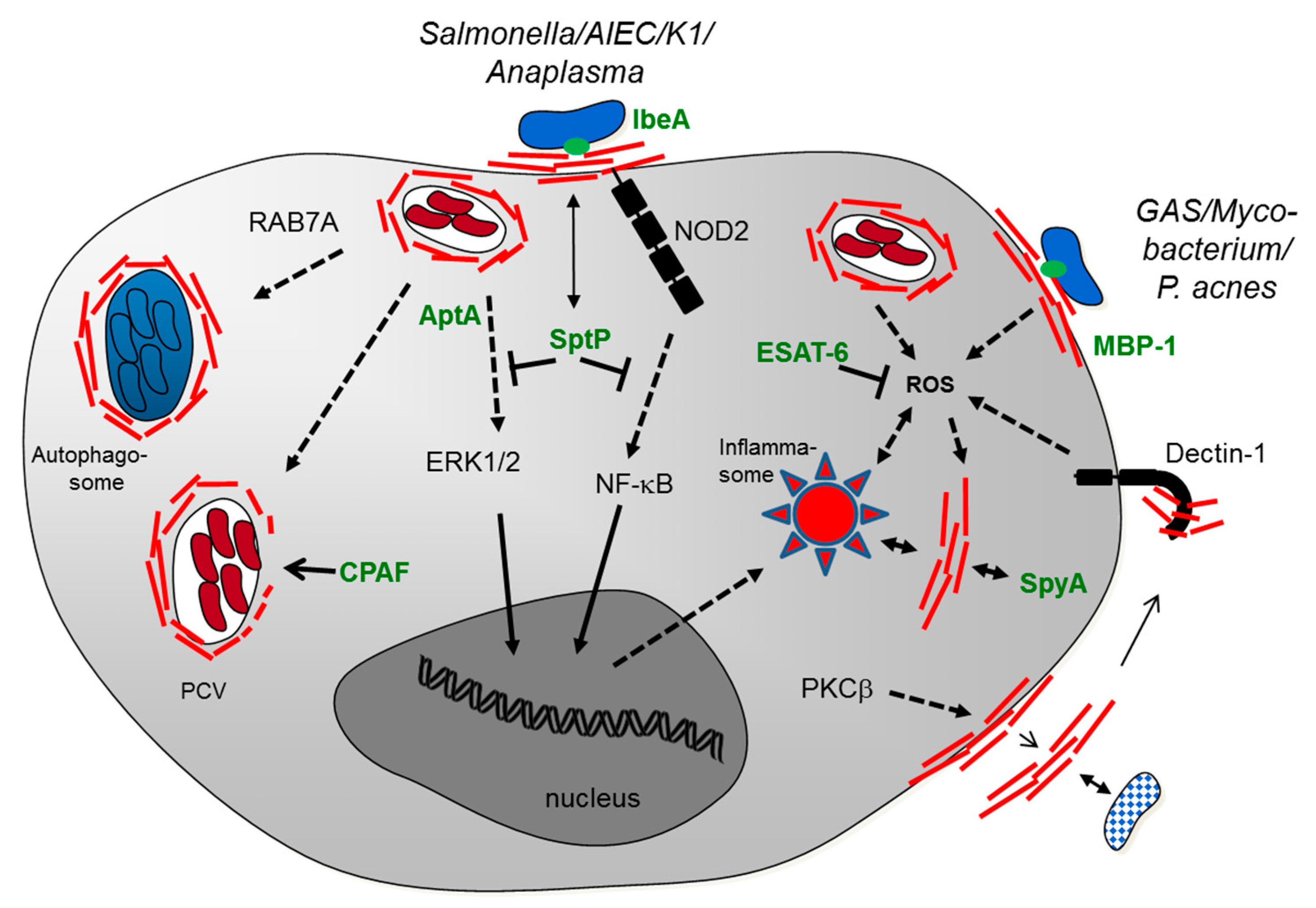
:1. Introduction
2. Interactions of Vimentin with Gram-Negative Bacterial Species
3. Interactions of Vimentin with Gram-Positive Bacterial Species
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Yoon, M.; Moir, R.D.; Prahlad, V.; Goldman, R.D. Motile properties of vimentin intermediate filament networks in living cells. J. Cell Biol. 1998, 143, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Mendez, M.G.; Kojima, S.; Goldman, R.D. Vimentin induces changes in cell shape, motility, and adhesion during the epithelial to mesenchymal transition. FASEB J. 2010, 24, 1838–1851. [Google Scholar] [CrossRef] [PubMed]
- Rogel, M.R.; Soni, P.N.; Troken, J.R.; Sitikov, A.; Trejo, H.E.; Ridge, K.M. Vimentin is sufficient and required for wound repair and remodeling in alveolar epithelial cells. FASEB J. 2011, 25, 3873–3883. [Google Scholar] [CrossRef] [PubMed]
- Menko, A.S.; Bleaken, B.M.; Libowitz, A.A.; Zhang, L.; Stepp, M.A.; Walker, J.L. A central role for vimentin in regulating repair function during healing of the lens epithelium. Mol. Biol. Cell. 2014, 25, 776–790. [Google Scholar] [CrossRef] [PubMed]
- Mor-Vaknin, N.; Punturieri, A.; Sitwala, K.; Markovitz, D.M. Vimentin is secreted by activated macrophages. Nat. Cell. Biol. 2003, 5, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; He, L.; Huang, S.H. Identification of a surface protein on human brain microvascular endothelial cells as vimentin interacting with Escherichia coli invasion protein IbeA. Biochem. Biophys. Res. Commun. 2006, 351, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Chi, F.; Jong, T.D.; Wang, L.; Ouyang, Y.; Wu, C.; Li, W.; Huang, S.H. Vimentin-mediated signalling is required for IbeA+ E. coli K1 invasion of human brain microvascular endothelial cells. Biochem. J. 2010, 427, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Chi, F.; Bo, T.; Wu, C.H.; Jong, A.; Huang, S.H. Vimentin and PSF act in concert to regulate IbeA+ E. coli K1 induced activation and nuclear translocation of NF-κB in human brain endothelial cells. PLoS ONE 2012, 7, e35862. [Google Scholar] [CrossRef] [PubMed]
- Mor-Vaknin, N.; Legendre, M.; Yu, Y.; Serezani, C.H.; Garg, S.K.; Jatzek, A.; Swanson, M.D.; Gonzalez-Hernandez, M.J.; Teitz-Tennenbaum, S.; Punturieri, A.; et al. Murine colitis is mediated by vimentin. Sci. Rep. 2013, 3, 1045. [Google Scholar] [CrossRef] [PubMed]
- Henderson, P.; Wilson, D.C.; Satsangi, J.; Stevens, C. A role for vimentin in Crohn disease. Autophagy 2012, 8, 1695–1696. [Google Scholar] [CrossRef] [PubMed]
- Stevens, C.; Henderson, P.; Nimmo, E.R.; Soares, D.C.; Dogan, B.; Simpson, K.W.; Barrett, J.C.; International Inflammatory Bowel Disease Genetics Consortium; Wilson, D.C.; Satsangi, J. The intermediate filament protein, vimentin, is a regulator of NOD2 activity. Gut 2013, 62, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Murli, S.; Watson, R.O.; Galán, J.E. Role of tyrosine kinases and the tyrosine phosphatase SptP in the interaction of Salmonella with host cells. Cell. Microbiol. 2001, 3, 795–810. [Google Scholar] [CrossRef] [PubMed]
- Guignot, J.; Servin, A.L. Maintenance of the Salmonella-containing vacuole in the juxtanuclear area: A role for intermediate filaments. Microb. Pathog. 2008, 45, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, B.; Mastronunzio, J.E.; Narasimhan, S.; Fankhauser, S.; Uchil, P.D.; Levy, R.; Graham, M.; Colpitts, T.M.; Lesser, C.F.; Fikrig, E. Anaplasma phagocytophilum AptA modulates Erk1/2 signalling. Cell. Microbiol. 2011, 13, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Kumar, Y.; Valdivia, R.H. Actin and intermediate filaments stabilize the Chlamydia trachomatis vacuole by forming dynamic structural scaffolds. Cell Host Microbe 2008, 4, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, I.; Bednar, M.M.; Amin, V.; Davis, B.K.; Ting, J.P.; McCafferty, D.G.; Valdivia, R.H. The Chlamydia protease CPAF regulates host and bacterial proteins to maintain pathogen vacuole integrity and promote virulence. Cell Host Microbe 2011, 10, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Snavely, E.A.; Kokes, M.; Dunn, J.D.; Saka, H.A.; Nguyen, B.D.; Bastidas, R.J.; McCafferty, D.G.; Valdivia, R.H. Reassessing the role of the secreted protease CPAF in Chlamydia trachomatis infection through genetic approaches. Pathog. Dis. 2014, 71, 336–351. [Google Scholar] [CrossRef] [PubMed]
- Bryant, A.E.; Bayer, C.R.; Huntington, J.D.; Stevens, D.L. Group A streptococcal myonecrosis: Increased vimentin expression after skeletal-muscle injury mediates the binding of Streptococcus pyogenes. J. Infect. Dis. 2006, 193, 1685–1692. [Google Scholar] [CrossRef] [PubMed]
- Icenogle, L.M.; Hengel, S.M.; Coye, L.H.; Streifel, A.; Collins, C.M.; Goodlett, D.R.; Moseley, S.L. Molecular and biological characterization of Streptococcal SpyA-mediated ADP-ribosylation of intermediate filament protein vimentin. J. Biol. Chem. 2012, 287, 21481–21491. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.E.; Beasley, F.C.; Keller, N.; Hollands, A.; Urbano, R.; Troemel, E.R.; Hoffman, H.M.; Nizet, V. A group A Streptococcus ADP-ribosyltransferase toxin stimulates a protective interleukin 1β-dependent macrophage immune response. MBio 2015, 6, e00133. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Barnes, P.F.; Porgador, A.; Roy, S.; Wu, S.; Nanda, J.S.; Griffith, D.E.; Girard, W.M.; Rawal, N.; Shetty, S.; et al. Vimentin expressed on Mycobacterium tuberculosis-infected human monocytes is involved in binding to the NKp46 receptor. J. Immunol. 2006, 177, 6192–6198. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, P.P.; Retnakumar, R.J.; Mundayoor, S. Downregulation of vimentin in macrophages infected with live Mycobacterium tuberculosis is mediated by Reactive Oxygen Species. Sci. Rep. 2016, 6, 21526. [Google Scholar] [CrossRef] [PubMed]
- Babrak, L.; Danelishvili, L.; Rose, S.J.; Kornberg, T.; Bermudez, L.E. The environment of “Mycobacterium avium subsp. hominissuis” microaggregates induces synthesis of small proteins associated with efficient infection of respiratory epithelial cells. Infect. Immun. 2015, 83, 625–636. [Google Scholar] [PubMed]
- Mak, T.N.; Fischer, N.; Laube, B.; Brinkmann, V.; Metruccio, M.M.; Sfanos, K.S.; Mollenkopf, H.J.; Meyer, T.F.; Brüggemann, H. Propionibacterium acnes host cell tropism contributes to vimentin-mediated invasion and induction of inflammation. Cell. Microbiol. 2012, 14, 1720–1733. [Google Scholar] [CrossRef] [PubMed]
- Bek-Thomsen, M.; Lomholt, H.B.; Scavenius, C.; Enghild, J.J.; Brüggemann, H. Proteome analysis of human sebaceous follicle infundibula extracted from healthy and acne-affected skin. PLoS ONE 2014, 9, e107908. [Google Scholar]
- Meriane, M.; Mary, S.; Comunale, F.; Vignal, E.; Fort, P.; Gauthier-Rouviére, C. Cdc42Hs and Rac1 GTPases induce the collapse of the vimentin intermediate filament network. J. Biol. Chem. 2000, 275, 33046–33052. [Google Scholar] [CrossRef] [PubMed]
- Perlson, E.; Hanz, S.; Ben-Yaakov, K.; Segal-Ruder, Y.; Seger, R.; Fainzilber, M. Vimentin-dependent spatial translocation of an activated MAP kinase in injured nerve. Neuron 2005, 45, 715–726. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, G.; Rogel, M.R.; Baker, M.A.; Troken, J.R.; Urich, D.; Morales-Nebreda, L.; Sennello, J.A.; Kutuzov, M.A.; Sitikov, A.; Davis, J.M.; et al. Vimentin regulates activation of the NLRP3 inflammasome. Nat. Commun. 2015, 6, 6574. [Google Scholar] [CrossRef] [PubMed]
- Thiagarajan, P.S.; Akbasli, A.C.; Kinter, M.T.; Willard, B.; Cathcart, M.K. Vimentin is a target of PKCβ phosphorylation in MCP-1-activated primary human monocytes. Inflamm. Res. 2013, 62, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wei, J.; Chang, M.; Liu, Z.; Li, D.; Hu, S.; Hu, L. Proteomic analysis of endothelial progenitor cells exposed to oxidative stress. Int. J. Mol. Med. 2013, 32, 607–614. [Google Scholar] [PubMed]
- Thiagarajan, P.S.; Yakubenko, V.P.; Elsori, D.H.; Yadav, S.P.; Willard, B.; Tan, C.D.; Rodriguez, E.R.; Febbraio, M.; Cathcart, M.K. Vimentin is an endogenous ligand for the pattern recognition receptor Dectin-1. Cardiovasc. Res. 2013, 99, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Du, N.; Cong, H.; Tian, H.; Zhang, H.; Zhang, W.; Song, L.; Tien, P. Cell surface vimentin is an attachment receptor for enterovirus 71. J. Virol. 2014, 88, 5816–5833. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.T.; Chien, S.C.; Chen, I.Y.; Lai, C.T.; Tsay, Y.G.; Chang, S.C.; Chang, M.F. Surface vimentin is critical for the cell entry of SARS-CoV. J. Biomed. Sci. 2016, 23, 14. [Google Scholar] [CrossRef] [PubMed]
- De Rivero Vaccari, J.P.; Minkiewicz, J.; Wang, X.; de Rivero Vaccari, J.C.; German, R.; Marcillo, A.E.; Dietrich, W.D.; Keane, R.W. Astrogliosis involves activation of retinoic acid-inducible gene-like signaling in the innate immune response after spinal cord injury. Glia 2012, 60, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Mostowy, S.; Shenoy, A.R. The cytoskeleton in cell-autonomous immunity: Structural determinants of host defence. Nat. Rev. Immunol. 2015, 15, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.C.; Wei, Y.; An, Z.; Zou, Z.; Xiao, G.; Bhagat, G.; White, M.; Reichelt, J.; Levine, B. Akt-mediated regulation of autophagy and tumorigenesis through Beclin 1 phosphorylation. Science 2012, 338, 956–959. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Bacterial Species | Host Cell | Interaction and Outcome | Virulence Factor | Reference |
|---|---|---|---|---|
| Gram negative | ||||
| Escherichia coli | endothelial cells | bacterial binding and invasion; vimentin interaction with NOD2; activation of NF-kB and ERK1/2 | IbeA | [6,7,8,9,10,11] |
| Salmonella enterica | epithelial cells macrophages | vimentin recruitment to membrane ruffles; bacterial invasion; fixation of vacuole in juxtanuclear area | SptP | [12,13] |
| Anaplasma phagocytophilum | neutrophils endothelial cells | modulation of vimentin network; activation of ERK1/2 signaling | AptA | [14] |
| Chlamydia trachomatis | epithelial cells | vimentin remodeling and cleavage; de/stabilization of bacterial vacuole | CPAF | [15,16,17] |
| Gram positive | ||||
| Streptococcus pyogenes | epithelial cells macrophages muscle cells | bacterial binding; ADP ribosylation of vimentin leading to inhibition of vimentin filament formation; altered immune signaling; inflammasome (?) | SpyA | [18,19,20] |
| Mycobacterium tuberculosis | monocytes macrophages | modulation of vimentin expression; altered ROS production; bacterial persistence | ESAT-6? | [21,22] |
| Mycobacterium avium | epithelial cells | bacterial binding; vimentin polymerization | MBP-1 | [23] |
| Propionibacterium acnes | epithelial cells | bacterial binding and invasion; modulation of immune signaling | ? | [24,25] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mak, T.N.; Brüggemann, H. Vimentin in Bacterial Infections. Cells 2016, 5, 18. https://doi.org/10.3390/cells5020018
Mak TN, Brüggemann H. Vimentin in Bacterial Infections. Cells. 2016; 5(2):18. https://doi.org/10.3390/cells5020018
Chicago/Turabian StyleMak, Tim N., and Holger Brüggemann. 2016. "Vimentin in Bacterial Infections" Cells 5, no. 2: 18. https://doi.org/10.3390/cells5020018
APA StyleMak, T. N., & Brüggemann, H. (2016). Vimentin in Bacterial Infections. Cells, 5(2), 18. https://doi.org/10.3390/cells5020018