A Positive Control for Detection of Functional CD4 T Cells in PBMC: The CPI Pool


 and
and
Abstract
:
1. Introduction
2. Materials and Methods
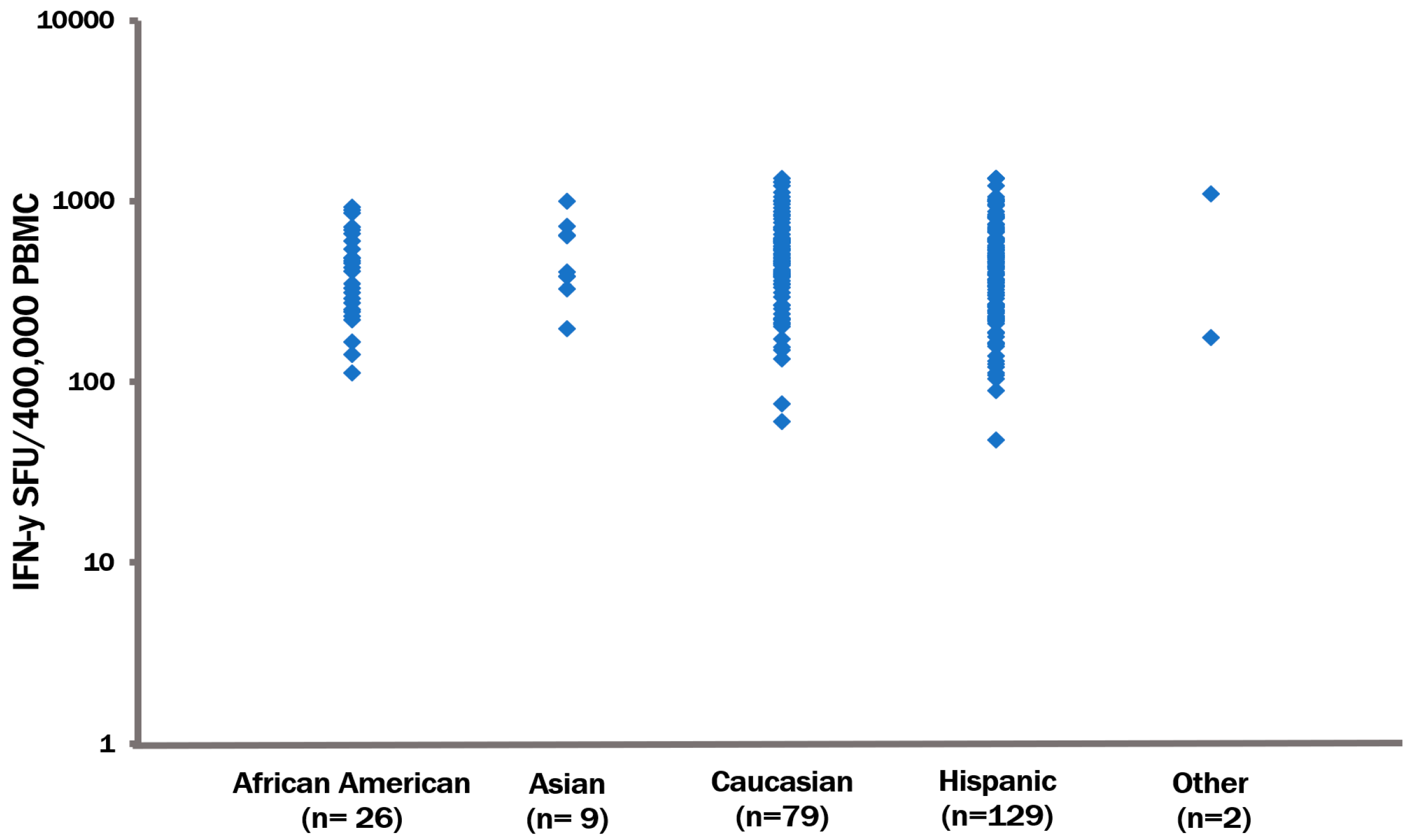
2.1. PBMC Donors
2.2. Antigens
2.3. Human Cytokine ELISPOT Assays
3. Results
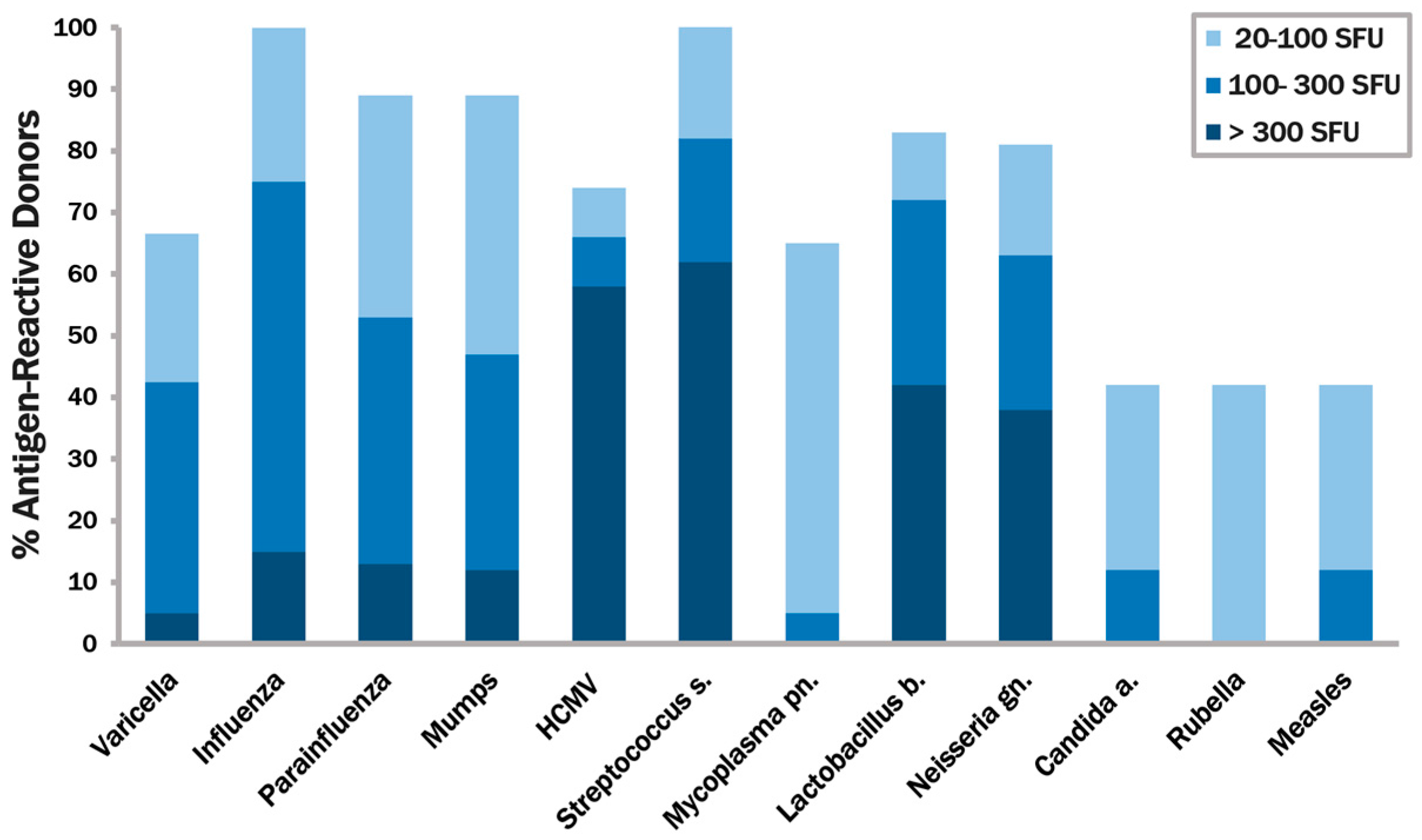
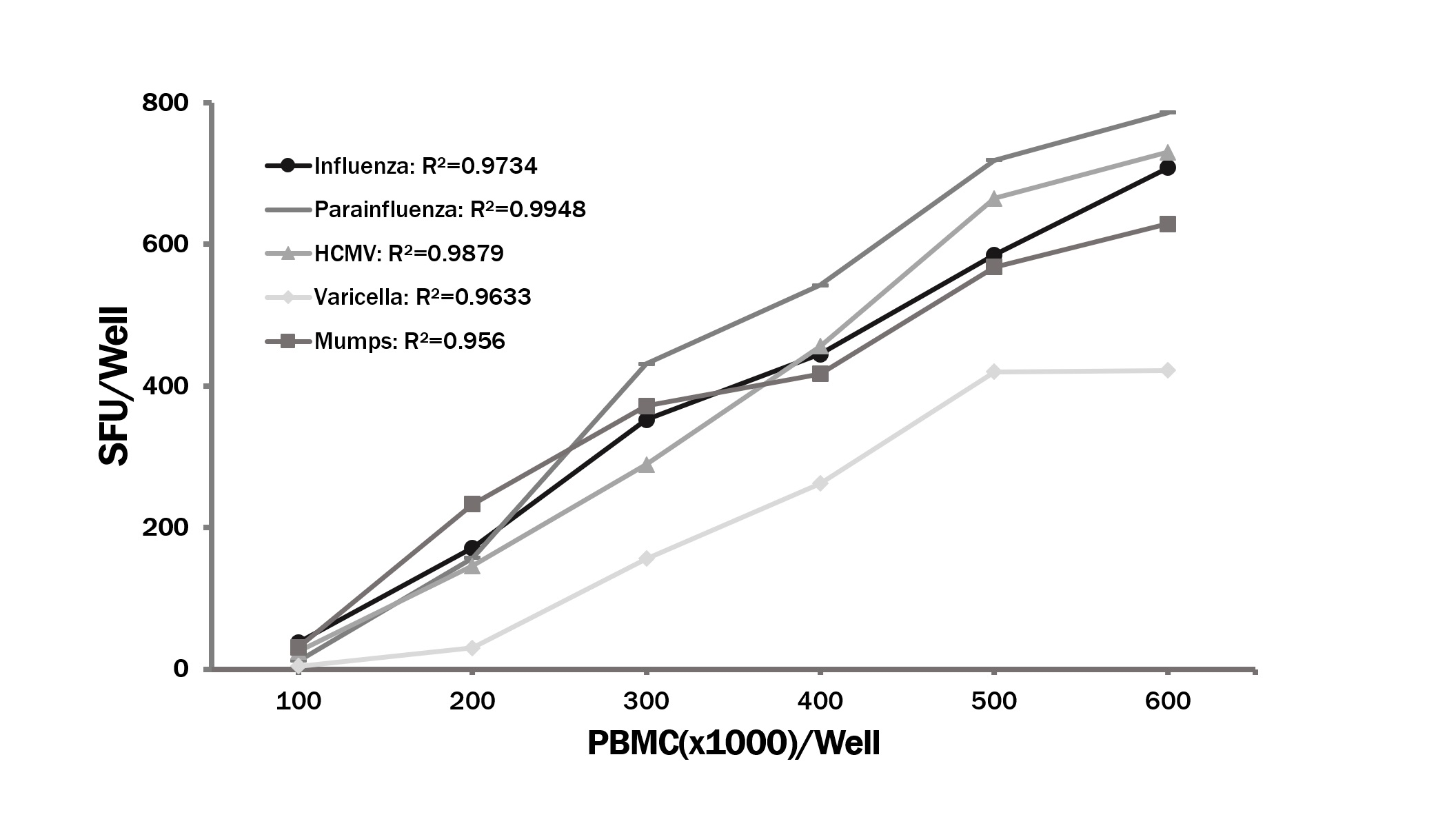
3.1. Identifying Recall Antigens that Elicit IFN-γ Production in the Majority of Healthy Human Donors
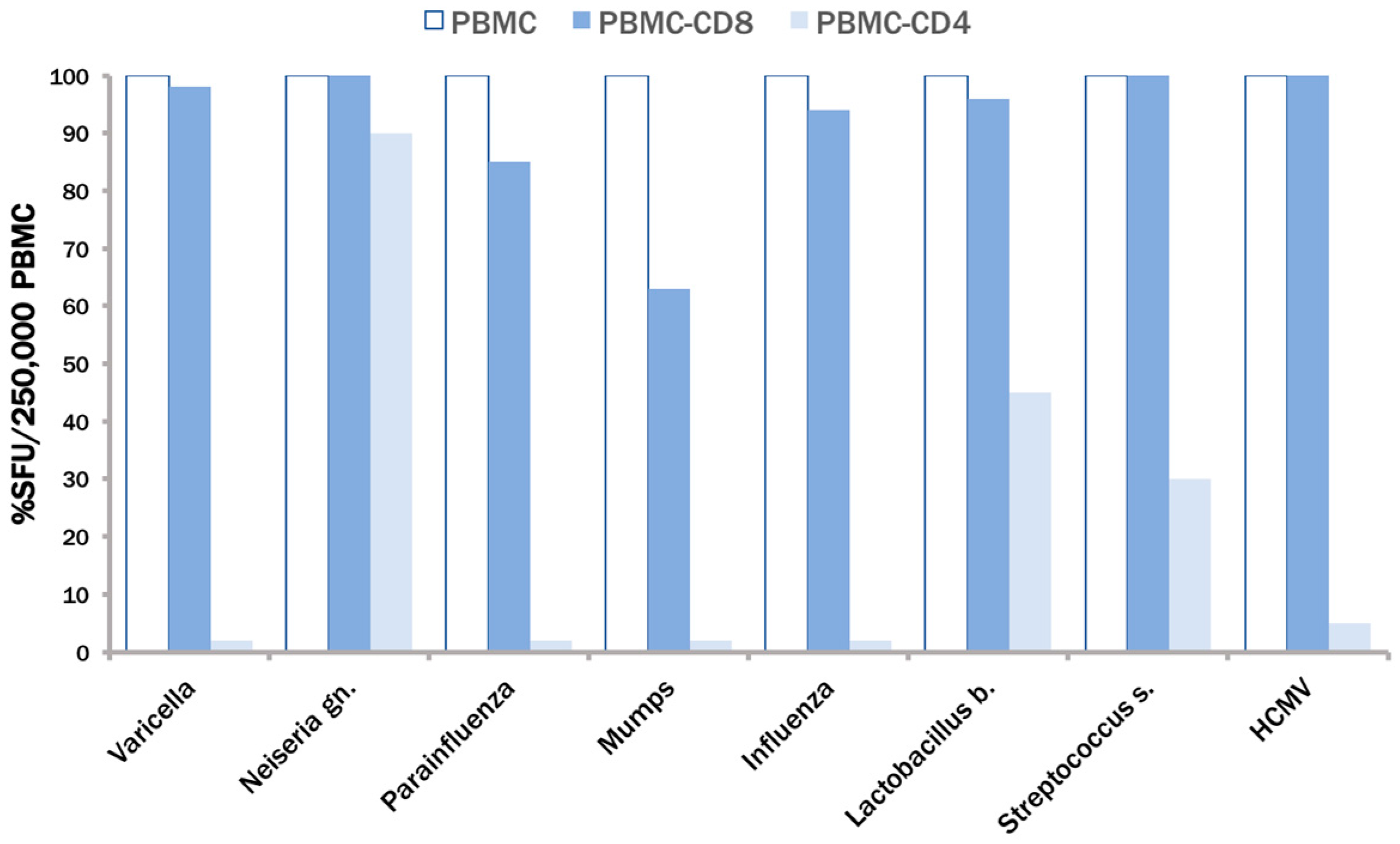
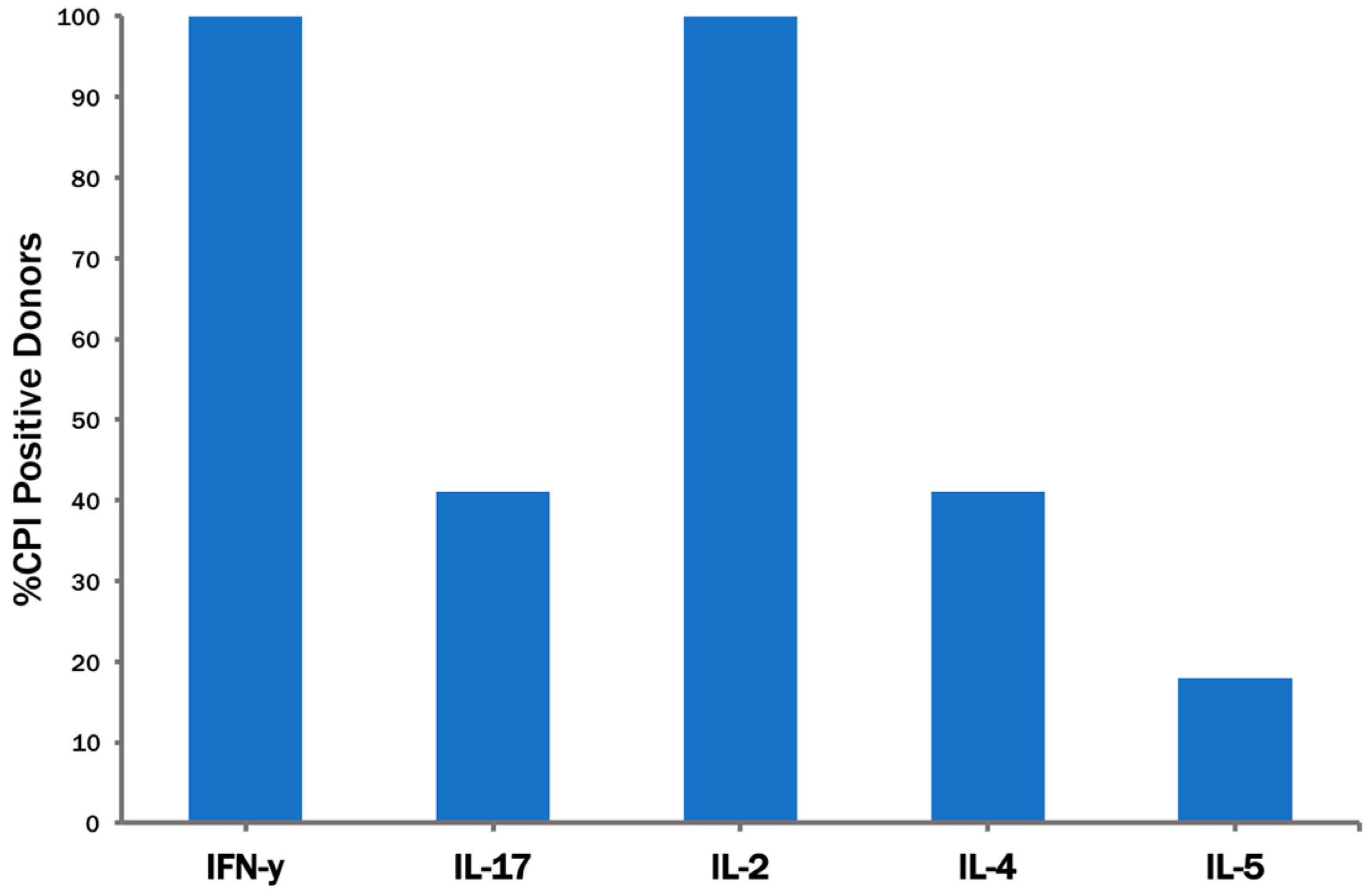
3.2. None of the Recall Antigens Tested Individually Suffices to Trigger CD4 Recall Responses in All Subjects
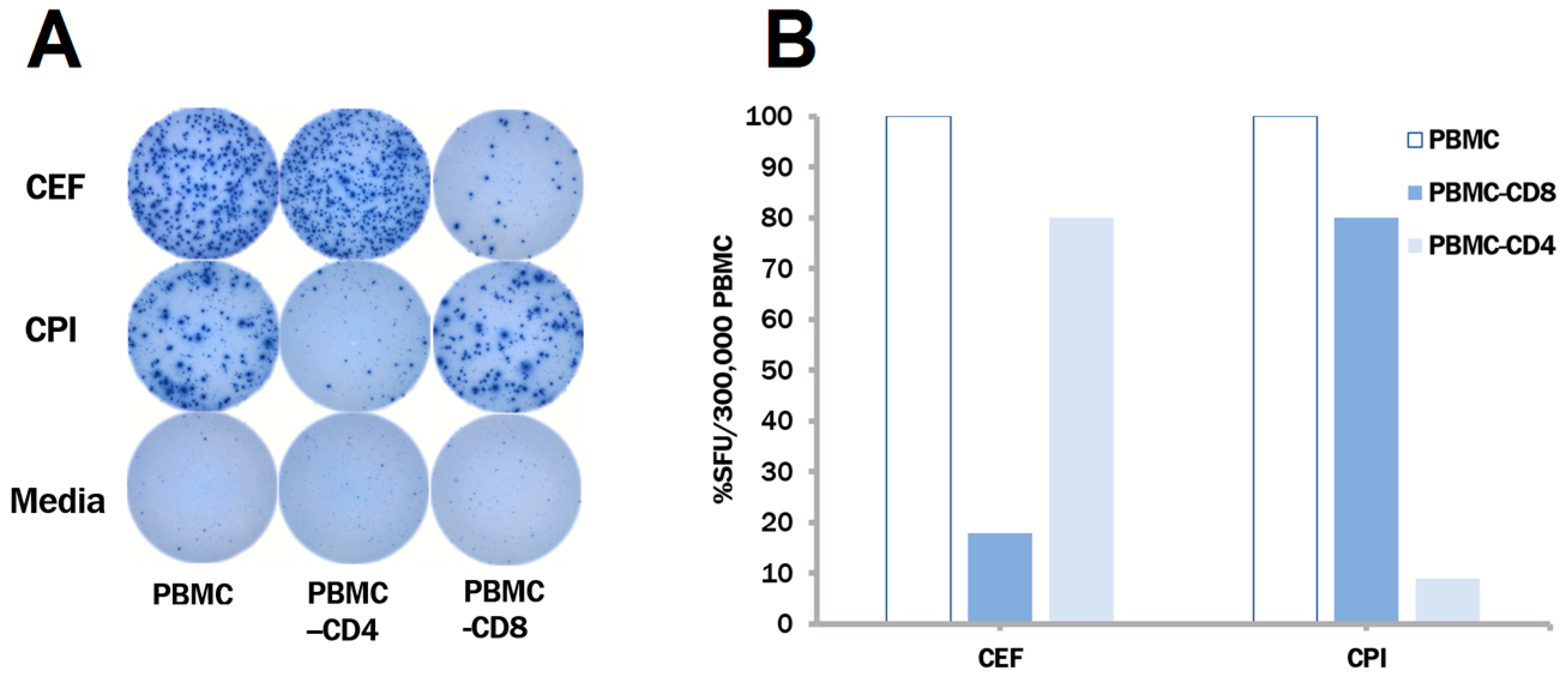
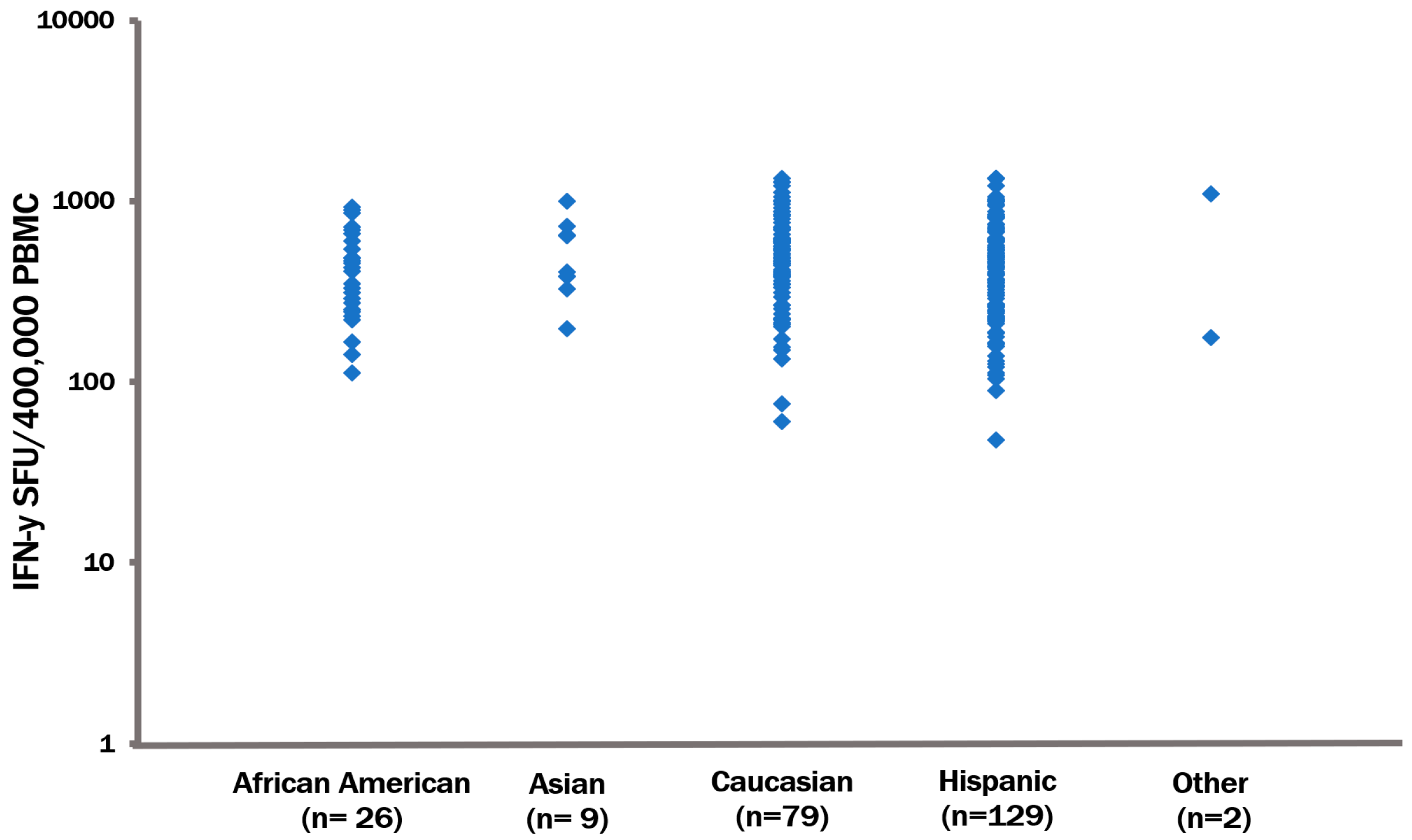
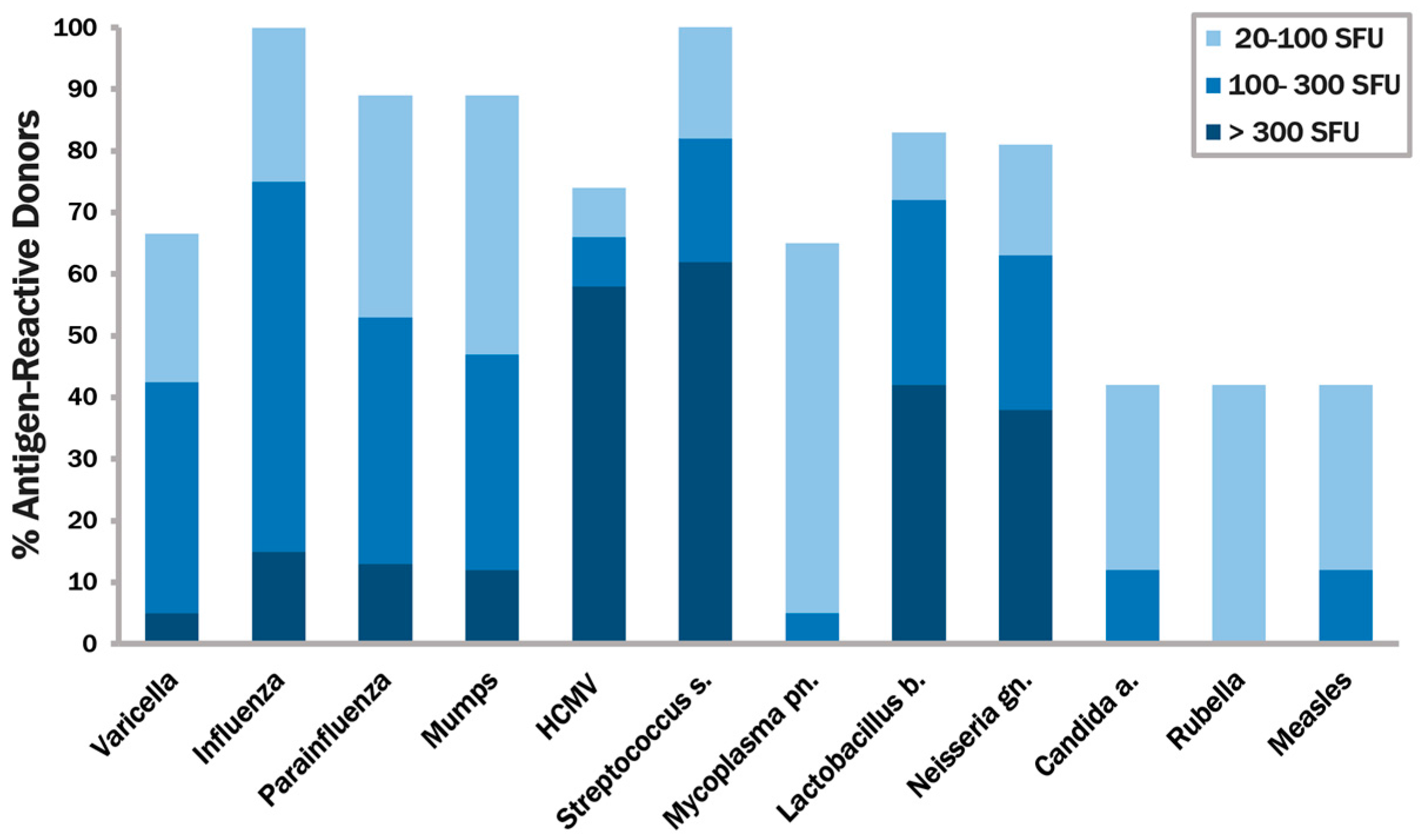
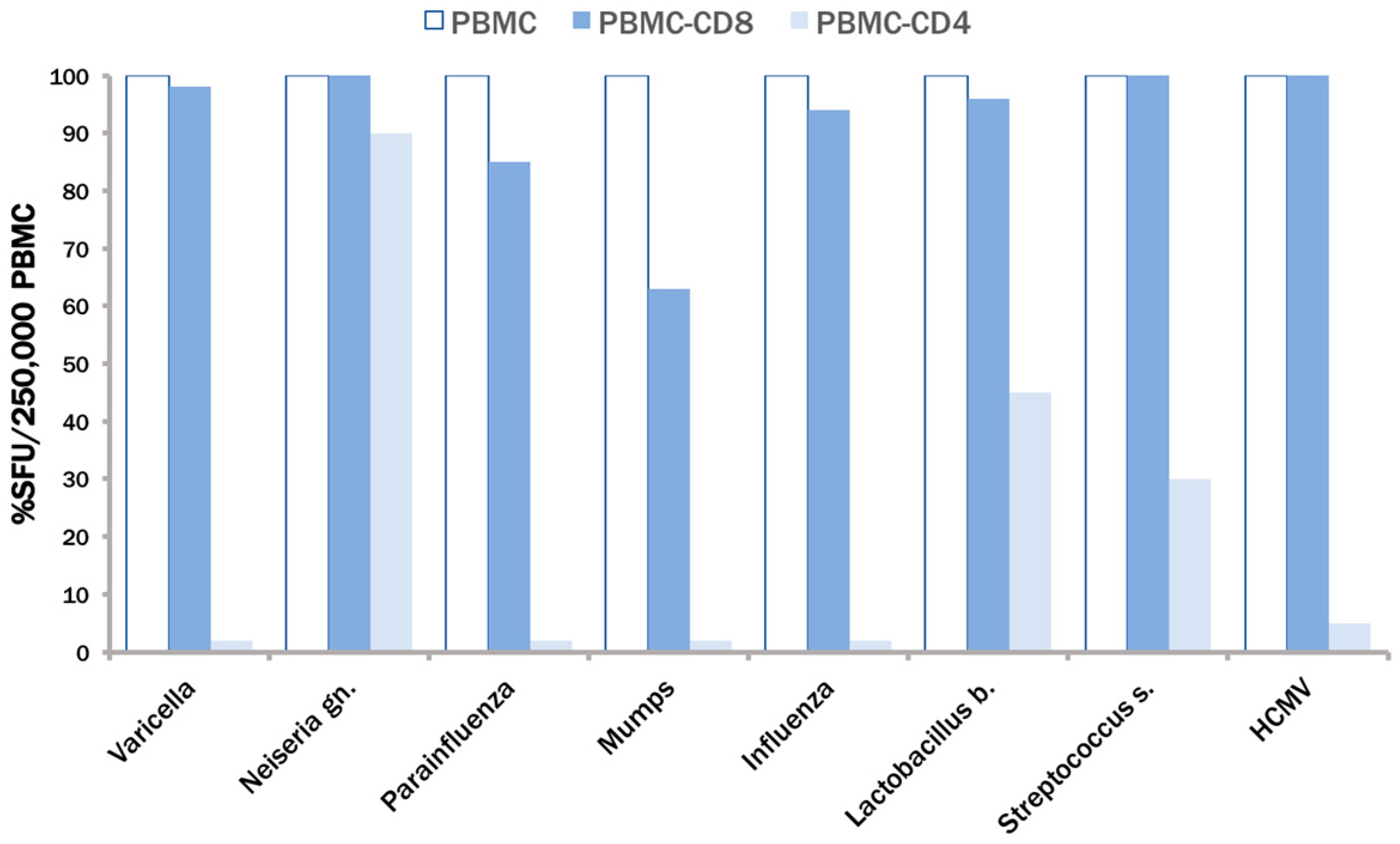
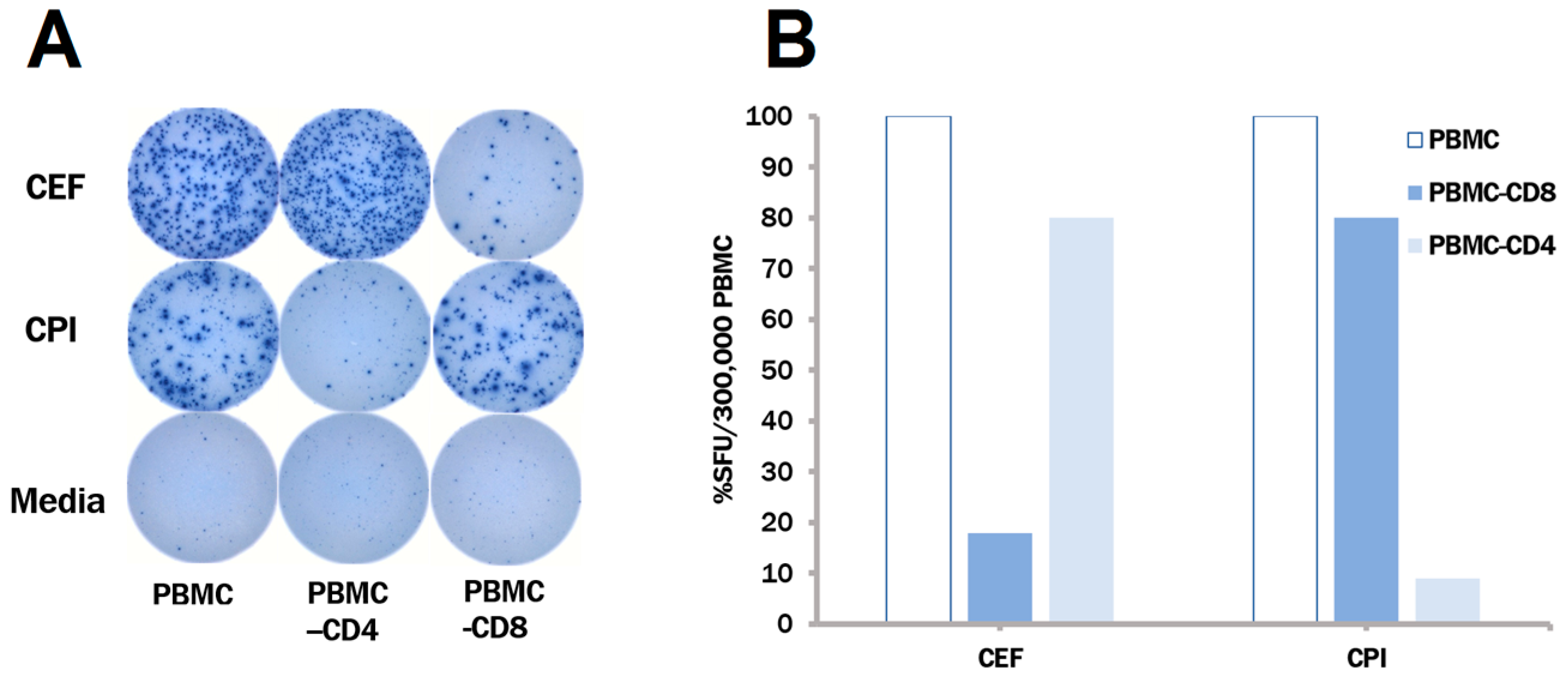
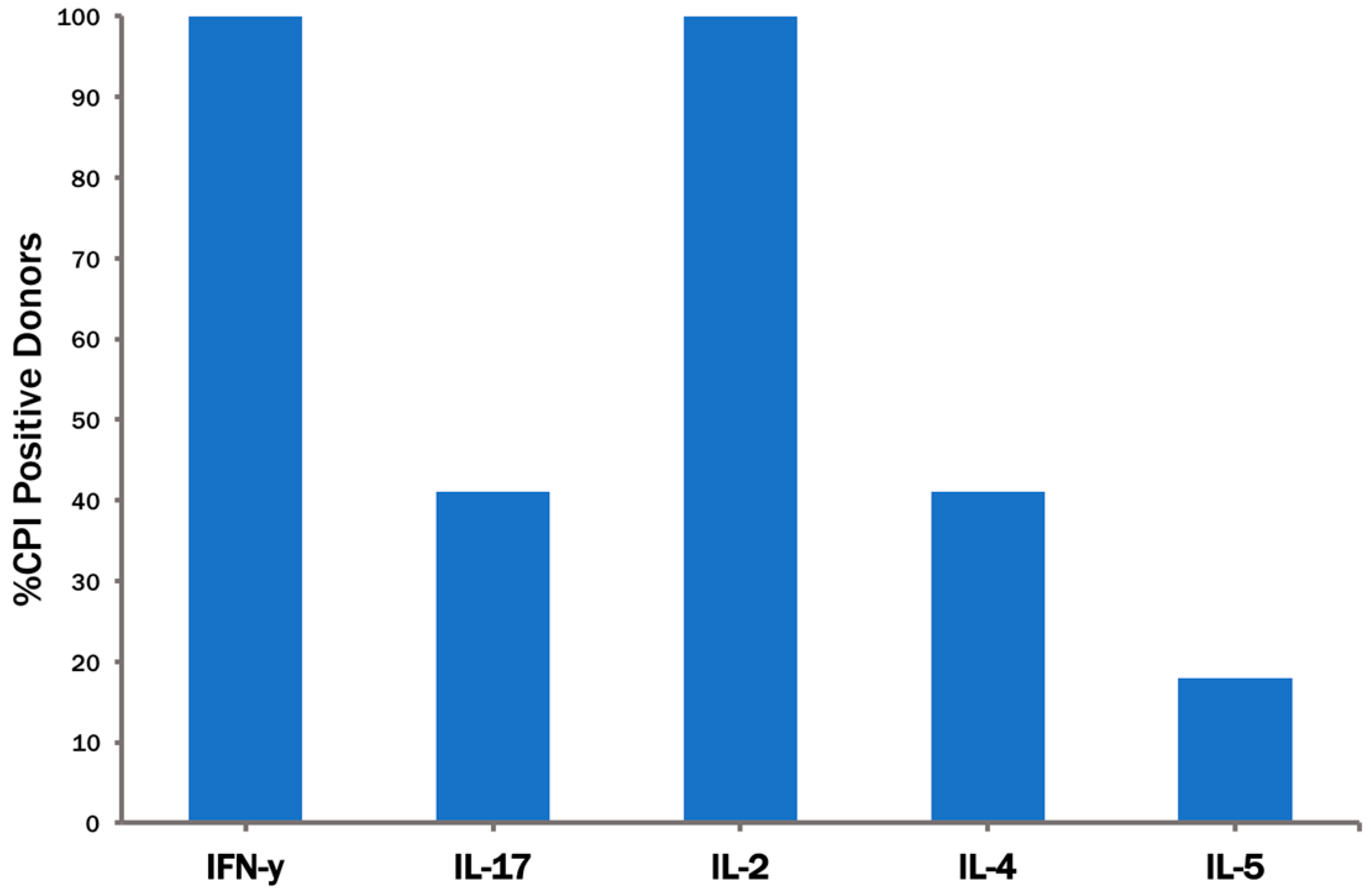
3.3. The CPI Pool Triggers CD4 Cell Recall Responses in 100% of Healthy Donors
3.4. In Contrast to CEF, CPI Pool Provides Comprehensive Determinant Coverage for the Detection of T Cell Memory
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Currier, J.R.; Kuta, E.G.; Turk, E.; Earhart, L.B.; Loomis-Price, L.; Janetzki, S.; Ferrari, G.; Birx, D.L.; Cox, J.H. A panel of MHC class I restricted viral peptides for use as a quality control for vaccine trial ELISPOT assays. J. Immunol. Methods 2002, 260, 157–172. [Google Scholar] [CrossRef]
- Kreher, C.R.; Dittrich, M.T.; Guerkov, R.; Boehm, B.O.; Tary-Lehmann, M. CD4+ and CD8+ cells in cryopreserved human PBMC maintain full functionality in cytokine ELISPOT assays. J. Immunol. Methods 2003, 278, 79–93. [Google Scholar] [CrossRef]
- Ramachandran, H.; Laux, J.; Moldovan, I.; Caspell, R.; Lehmann, P.V.; Subbramanian, R.A. Optimal thawing of cryopreserved peripheral blood mononuclear cells for use in high-throughput human immune monitoring studies. Cells 2012, 1, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Caspell, R.; Karulin, A.Y.; Ahmad, M.; Haicheur, N.; Abdelsalam, A.; Johannesen, K.; Vignard, V.; Dudzik, P.; Georgakopoulou, K.; et al. ELISPOT assays provide reproducible results among different laboratories for T-cell immune monitoring--even in hands of ELISPOT-inexperienced investigators. J. Immunotoxicol. 2009, 6, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Gill, D.K.; Huang, Y.; Levine, G.L.; Sambor, A.; Carter, D.K.; Sato, A.; Kopycinski, J.; Hayes, P.; Hahn, B.; Birungi, J.; et al. Equivalence of ELISpot assays demonstrated between major HIV network laboratories. PLoS ONE 2010, 5, e14330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.G.; Liu, X.; Kaufhold, R.M.; Clair, J.; Caulfield, M.J. Development and Validation of a Gamma Interferon ELISPOT Assay for Quantitation of Cellular Immune Responses to Varicella-Zoster Virus. Clin. Diagn. Lab. Immunol. 2001, 8, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.C.; Shields, M.; DiBrino, M.; Brooks, A.; Coligan, J.E. Peptide binding to MHC class I molecules: Implications for antigenic peptide prediction. Immunol. Res. 1995, 14, 34–57. [Google Scholar] [CrossRef] [PubMed]
- Germain, R.N. The biochemistry and cell biology of antigen presentation by MHC class I and class II molecules. Implications for development of combination vaccines. Ann. N. Y. Acad. Sci. 1995, 754, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Cresswell, P. Antigen processing and presentation. Immunol. Rev. 2005, 207, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.G.; Joseph, H.R.; Green, T.; Field, J.A.; Wooters, M.; Kaufhold, R.M.; Antonello, J.; Caulfield, M.J. Establishing Acceptance Criteria for Cell-Mediated-Immunity Assays Using Frozen Peripheral Blood Mononuclear Cells Stored under Optimal and Suboptimal Conditions. Clin. Vaccine Immunol. 2007, 14, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Kuerten, S.; Batoulis, H.; Recks, M.S.; Karacsony, E.; Zhang, W.; Subbramanian, R.A.; Lehmann, P.V. Resting of Cryopreserved PBMC Does Not Generally Benefit the Performance of Antigen-Specific T Cell ELISPOT Assays. Cells 2012, 1, 409–427. [Google Scholar] [CrossRef] [PubMed]
- Wunsch, M.; Zhang, W.; Hanson, J.; Caspell, R.; Karulin, A.Y.; Recks, M.S.; Kuerten, S.; Sundararaman, S.; Lehmann, P.V. Characterization of the HCMV-Specific CD4 T Cell Responses that Are Associated with Protective Immunity. Viruses 2015, 7, 4414–4437. [Google Scholar] [CrossRef] [PubMed]
- Wunsch, M.; Caspell, R.; Kuerten, S.; Lehmann, P.V.; Sundararaman, S. Serial Measurements of Apoptotic Cell Numbers Provide Better Acceptance Criterion for PBMC Quality than a Single Measurement Prior to the T Cell Assay. Cells 2015, 4, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Duechting, A.; Przybyla, A.; Kuerten, S.; Lehmann, P.V. Delayed Activation Kinetics of Th2- and Th17 Cells Compared to Th1 Cells. Cells 2017, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Karulin, A.Y.; Karacsony, K.; Zhang, W.; Targoni, O.S.; Moldovan, I.; Dittrich, M.; Sundararaman, S.; Lehmann, P.V. ELISPOTs Produced by CD8 and CD4 Cells Follow Log Normal Size Distribution Permitting Objective Counting. Cells 2015, 4, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Karulin, A.Y.; Caspell, R.; Dittrich, M.; Lehmann, P.V. Normal Distribution of CD8+ T-Cell-Derived ELISPOT Counts within Replicates Justifies the Reliance on Parametric Statistics for Identifying Positive Responses. Cells 2015, 4, 96–111. [Google Scholar] [CrossRef] [PubMed]
- Steinman, L. A brief history of T(H)17, the first major revision in the T(H)1/T(H)2 hypothesis of T cell-mediated tissue damage. Nat. Med. 2007, 13, 139–145. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor-ID | CPI | CEF | Donor-ID | CPI | CEF | Donor-ID | CPI | CEF | Donor-ID | CPI | CEF |
|---|---|---|---|---|---|---|---|---|---|---|---|
| LP_235 | 810 | 112 | LP_260 | 462 | 362 | LP_285 | 268 | 83 | LP_313 | 333 | 29 |
| LP_236 | 1013 | 2 | LP_261 | 725 | 600 | LP_286 | 210 | 375 | LP_314 | 600 | 161 |
| LP_237 | 746 | 84 | LP_262 | 155 | 25 | LP_288 | 167 | 51 | LP_315 | 163 | 22 |
| LP_238 | 491 | 1 | LP_263 | 1220 | 323 | LP_290 | 223 | 7 | LP_316 | 393 | 116 |
| LP_239 | 843 | 4 | LP_264 | 999 | 557 | LP_292 | 240 | 352 | LP_317 | 188 | 4 |
| LP_240 | 619 | 509 | LP_265 | 585 | 155 | LP_293 | 224 | 138 | LP_318 | 248 | 18 |
| LP_241 | 563 | 287 | LP_266 | 369 | 91 | LP_294 | 268 | 58 | LP_319 | 273 | 32 |
| LP_242 | 957 | 41 | LP_267 | 550 | 123 | LP_295 | 500 | 101 | LP_320 | 89 | 2 |
| LP_243 | 125 | 55 | LP_268 | 433 | 591 | LP_297 | 260 | 11 | LP_321 | 600 | 99 |
| LP_244 | 536 | 358 | LP_269 | 439 | 346 | LP_298 | 215 | 208 | LP_322 | 231 | 0 |
| Donor-ID | Media | CPI | CEF | Gr.2 | HCMV Peptide Pools | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pp65 | IE-1 | IE-2 | UL28 | UL32 | UL36 | UL55 | UL82 | UL94 | UL103 | US3 | |||||
| LP_099 | 2 | 829 | 5 | 661 | 565 | 46 | 29 | 91 | 928 | 77 | 42 | 53 | 0 | 38 | 587 |
| LP_193 | 0 | 720 | 3 | 250 | 437 | 278 | 98 | 13 | 152 | 139 | 651 | 2 | 290 | 2 | 269 |
| LP_194 | 1 | 262 | 3 | 27 | 136 | 395 | 10 | 208 | 40 | 50 | 19 | 2 | 8 | 10 | 37 |
| LP_141 | 3 | 322 | 2 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 2 | 2 | 0 | 0 | 0 |
| LP_151 | 1 | 221 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 |
| LP_218 | 0 | 266 | 0 | 0 | 2 | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 2 | 0 | 0 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiller, A.; Zhang, T.; Li, R.; Duechting, A.; Sundararaman, S.; Przybyla, A.; Kuerten, S.; Lehmann, P.V. A Positive Control for Detection of Functional CD4 T Cells in PBMC: The CPI Pool. Cells 2017, 6, 47. https://doi.org/10.3390/cells6040047
Schiller A, Zhang T, Li R, Duechting A, Sundararaman S, Przybyla A, Kuerten S, Lehmann PV. A Positive Control for Detection of Functional CD4 T Cells in PBMC: The CPI Pool. Cells. 2017; 6(4):47. https://doi.org/10.3390/cells6040047
Chicago/Turabian StyleSchiller, Annemarie, Ting Zhang, Ruliang Li, Andrea Duechting, Srividya Sundararaman, Anna Przybyla, Stefanie Kuerten, and Paul V. Lehmann. 2017. "A Positive Control for Detection of Functional CD4 T Cells in PBMC: The CPI Pool" Cells 6, no. 4: 47. https://doi.org/10.3390/cells6040047