CD38 Expression by Myeloma Cells and Its Role in the Context of Bone Marrow Microenvironment: Modulation by Therapeutic Agents


1
Department of Medicine and Surgery, University of Parma, 43126 Parma, Italy
2
Hematology, “Azienda Ospedaliero-Universitaria di Parma”, 43126 Parma, Italy
*
Author to whom correspondence should be addressed.
Cells 2019, 8(12), 1632; https://doi.org/10.3390/cells8121632
Submission received: 8 November 2019
/
Revised: 28 November 2019
/
Accepted: 11 December 2019
/
Published: 13 December 2019
(This article belongs to the Special Issue CD38 and Disease: A Bi-Directional Cross-Talk between Pathology and Physiology)
Abstract
:In the last decades CD38 has emerged as an attractive target for multiple myeloma (MM). CD38 is a novel multifunctional glycoprotein that acts as a receptor, adhesion molecule interacting with CD31 and as an ectoenzyme. As an ectoenzyme, CD38 functions as a metabolic sensor catalyzing the extracellular conversion of NAD+ to the immunosuppressive factor adenosine (ADO). Other ectoenzymes, CD73 and CD203a, together with CD38, are also involved in the alternative axis of extracellular production of ADO, bypassing the canonical pathway mediated by CD39. CD38 is ubiquitously expressed in the bone marrow microenvironment; however, only MM cells display a very high surface density, which lead to the development of several anti-CD38 monoclonal antibodies (mAbs). The efficacy of anti-CD38 mAbs depends from the presence of CD38 on the surface of MM and immune-microenvironment cells. Interestingly, it has been reported that several drugs like lenalidomide, panobinostat, the all-trans retinoic acid and the DNA methyltransferase inhibitors may increase the expression of CD38. Hence, the possibility to modulate CD38 by increasing its expression on MM cells is the pre-requisite to potentiate the clinical efficacy of the anti-CD38 mAbs and to design clinical trials with the combination of anti-CD38 mAbs and these drugs.
1. Introduction
Multiple myeloma (MM) cells are characterized by tight relationship with the bone marrow (BM) microenvironment which supports their growth, survival, and induces drug resistance. MM cells overexpress several surface adhesion molecules that are involved in the relationship with the micro-environmental cells like BM stromal cells (BMSCs), osteoblasts (OBs), and endothelial cells [1]. Among the adhesion molecules, CD38 is highly expressed by MM cells. CD38 has become one of the main targets for monoclonal antibody (mAb) therapy in MM patients [2]. Daratumumab (DARA) is the first anti- CD38 mAb approved for the treatment of relapsed and refractory MM based on the results of two phase I/II trials [3]. Other anti-CD38 mAb were also clinically developed as isatuximab and MOR202 [4]. The combination of anti-CD38 mAbs with main anti-MM drugs as proteasome inhibitors (PIs) and immunomodulatory drugs (IMiDs) demonstrated a high clinical efficacy in randomized phase III trials [5] leading to new therapeutic paradigms in relapsed/refractory MM patients. However, a considerable amount of MM patients does not respond or are refractory to the treatment with anti-CD38 mAbs either as single agents or in combination with PIs or IMiDs [6,7,8].
The clinical efficacy of anti-CD38 mAbs seems to be related, at least in part, to the intensity of CD38 expression by MM cells and other cells of the immune-microenvironment. The possibility to modulate CD38 through an increase of its expression by MM cells is the pre-requisite to potentiate the efficacy of anti-CD38 mAbs. Different pharmacological agents have demonstrated the capacity to increase the expression of CD38 by MM and BM microenvironment cells. This review summarizes the main experimental evidences on this topic giving the rational for drug combinations with anti-CD38 mAbs in the treatment of MM.
2. CD38 Expression by MM Cells and BM Microenvironment
MM is a hematological cancer characterized by the accumulation and proliferation of malignant plasma cells (PCs) in the BM [9]. The close interaction between PCs and BM microenvironment cells creates a permissive niche for tumor survival and disease progression, characterized by osteolytic bone disease and immune-suppression [10]. Both soluble factors and cell-to-cell contact mechanisms are involved in this cross-talk. Among the surface molecules, which allow the adhesion to the microenvironment, MM cells highly express CD38 [11], which made it an attractive therapeutic target for mAbs [12,13]. Several studies demonstrated that only PCs strongly express CD38 antigens in BM, and that no PCs are detectable in either CD38neg cell fraction or fraction of cells weakly expressing CD38 antigens (CD38low) [14,15]. However, activated B cell, T cells and NK cells up-regulate CD38 surface expression to levels similar to that found on PCs [16].
CD38 is a 45-kDa type II transmembrane glycoprotein, which plays a dual role as a receptor and ectoenzyme [17]. It is expressed on normal cell subsets, such as T cells, NK cells, B cells, and dendritic cells [18]. It is involved in T cell activation and proliferation, B cell differentiation, and neutrophils and monocytes chemotaxis, [17,19,20]. In addition, CD38 interacts with the non-substrate ligand CD31, which is constitutively expressed by endothelial cells [21]. Interestingly, a co-expression of CD38 and CD31 was also demonstrated in MM cells but not on PC leukemia [22]. Accordingly, we have recently reported that extra-medullary MM cells can also lose the expression of CD38 [23].
As ectoenzyme, CD38 acts like a metabolic sensor which catalyzes the extracellular conversion of NAD+ to regulators of calcium signaling, such as adenosine (ADO), according to pH status [24]. Other ectoenzymes, CD73 and CD203a, together with CD38, are involved in the alternative axis of extracellular production of the immunosuppressive factor adenosine ADO, bypassing the canonical pathway mediated by CD39 [25].
ADO levels are significantly higher in the BM plasma of MM patients than asymptomatic monoclonal gammopathies as Monoclonal Gammopathy of Uncertain Significance (MGUS) and smoldering MM (SMM), suggesting that ADO is produced in the MM niche by an ectoenzymatic CD38 network [26]. Recently, we have also investigated the expression and function of ectoenzymes on microvesicles (MVs) isolated from BM plasma samples of MM patients. Our results show that the percentage of MVs expressing high levels of ectoenzymes was higher when derived from MM patients compared to MGUS and SMM. Consistently, BM CD138+ PCs from MM patients expressed high levels of all ectoenzymes [27]. The MVs immunophenotype of MM patients indicated a high expression level of CD38, CD39, CD73, and CD203a ectoenzymes as shared by CD138+ PCs. Finally, we demonstrated that the ATP, NAD+, ADPR, and AMP to ADO catabolism was higher in MVs from MM patients than in those from controls. This indicates that the ectoenzymes expressed by MVs isolated from BM samples of MM patients were functionally active and involved in the higher ADO production as compared to MGUS and SMM [27].
It is known that the interactions between MM PCs and other cells of the BM niche, such as osteoclasts (OCs), OBs, and BMSCs, induce the production of ADO, which promotes tumor survival and immune escape [28,29,30]. Several studies also reported that ADO concentrations in the BM MM niche correlates with disease progression and may be an useful prognostic marker related to ISS staging together with others [26]. In addition, it has been interestingly shown that the fully humanized anti-CD38 mAb, DARA, is able to modulate CD38 enzymatic activity in vitro, thus reducing ADO levels and reverting its immunosuppressive effect [28].
Analysis of CD38 and the other ectoenzyme distribution within MM bone niche revealed that only PCs express high levels of CD38 [31]. However, some studies showed that CD38 expression intensity is highly heterogeneous on MM cells, and its expression does not differ from newly diagnosed and relapsed/refractory MM patients [32]. MM PCs also express other ectoenzymes like CD39 and CD73, although their levels differ from patient to patient [31].
The expression profile of BMSCs and OBs show that both cell types are CD38−/CD39− while expressing CD73 and CD203a [26,30,31]. Indeed, CD38 decreases during OB differentiation with a concomitant increase of CD203a [30].
By using murine and rabbit models, studies from Sun L. et al. [33,34] described an involvement of CD38 in the remodeling of the adult skeleton [33,34]. Specifically, a reduced bone mineral density was detected in CD38−/− mice, along with a strong ability of the hematopoietic stem cells to differentiate into highly resorptive OCs [34]. CD38 was also detectable on rabbit OC plasma membrane where it exerts an ADP ribosyl cyclase activity [34]. Bone resorption was inhibited after the treatment with an anti-CD38 agonist antibody, further confirming CD38 involvement in skeleton remodeling [33]. These studies also supported the hypothesis of CD38 as a metabolic sensor able to couple OC bone resorption activity with its Ca2+ signaling pathway [35].
However, few data are currently available on CD38 expression on human monocytes and OCs in MM. A recent study from Costa F. et al. [31] shows that CD38 is expressed on the surface of early OC progenitors but it is lost during in vitro differentiation toward an osteoclastogenic phenotype [31]. Moreover, in vitro experiments demonstrated that the use of the fully humanized anti-CD38 mAb, DARA, inhibits OC formation and activity, confirming the involvement of CD38 in bone remodeling, even in MM patients [31].
Besides their major function in bone remodeling, OCs also display immunosuppressive properties on T cells [36] which in turn induce osteoclastogenesis [37], thus establishing a feedback loop mechanism. An G. et al. [38] recently demonstrated that anti-CD38 mAb, SAR650984, enhances the cytotoxic effect of T cells on OCs in vitro [38]. On the other hand, the presence of SAR650984 in T cells-OCs co-cultures restores T cell proliferation, reverting the OC immune-suppressive effects [38]. Overall, these data suggested that targeting CD38 may affect MM-induce bone disease, by restoring T cell function or inhibiting early OC formation.
Lastly, Krejcik J. et al. [16] suggested that CD38 expression may define a subset of T regulatory cells (Tregs) with enhanced immune suppressive effect [16]. Studies on MM revealed that Tregs expresses CD38 at higher level compared to T constitutive cells (Tcons). Moreover, the percentage of CD38high cells is higher in MM patients versus healthy donors [16,39]. In vitro co-culture also showed that MM cells significantly induces Tcons conversion toward a Treg phenotype, which is characterized by an increased expression of CD38, CD25, and FoxP3 compared with natural Tregs [39].
Together all these data indicate a pleiotropic role of CD38 in MM bone niche, supporting the use of therapeutic strategies targeting this molecule to inhibit MM cell growth, survival, and to revert immune-suppression and bone disease in MM patients.
3. CD38 Modulation by Anti-CD38 Monoclonal Antibodies in MM Cells
Several anti-CD38 mAbs have been developed and are under clinical development: Two fully human Abs (MOR202, TAK-079) and one chimeric (isatuximab); while DARA is the yet approved for the treatment of MM as single agent and in combination with standards of care in relapsed/refractory MM patients [40].
Anti-CD38 mAbs show different mechanisms of action: Fc-dependent immune effector mechanisms, direct effects, and immunomodulatory effects. The first mechanism includes complement-dependent cytotoxicity (CDC), antibody-dependent cell-mediated cytotoxicity (ADCC), antibody-dependent cellular phagocytosis (ADCP), and apoptosis upon secondary cross-linking [40]. The direct effect is mediated by caspase-dependent apoptotic pathway, as well as the lysosomal cell death pathway [41]. Finally, it has been recently reported that DARA treatment induces elimination of CD38-positive immune suppressor cells, such as Tregs, regulatory B cells, and myeloid-derived suppressor cells [16,42], together with activated NK cells [43], suggesting DARA ability to target cells with lower levels of CD38.
Despite the in vitro effects results and the well-established clinical efficacy of the anti-CD38Abs, some open issues may occur. Specifically, a study from Nijhof IS et al. [8] reported that CD38 expression is reduced in both BM and PB CD138+ cells, together with non-tumor immune cells (NK, T cells, B cells, and monocytes) following the first DARA infusion and it increases again following DARA discontinuation [8]. Different mechanisms have been proposed to explain this event. One is that the interaction of DARA with its target induces a polar aggregation, known as “capping”, of CD38 molecules, followed by exocytosis as MVs [28] or endocytosis of DARA-CD38 complex [44]. Another mechanism involves the degradation of the antigen–antibody complex or the rapid elimination of myeloma cells expressing high levels of CD38, as demonstrated by the fact that MM CD38 levels were only reduced in the presence of complement or effector cells. More recently, it has been suggested a mechanism called “trogocytosis”, where fragments of the plasma membrane carrying antigen–antibody complexes, together with other parts of cell membrane (CD49d, CD56, CD138, CD54, and CD44) are transferred by monocytes and granulocytes, in the absence of evident phagocytosis of tumor cells [44]. This process seems to be involved the reduction of CD38 not only on MM cells but also on NK cells. Interestingly, these in vitro studies were then confirmed in vivo in patients treated with DARA, both in the presence or absence of lenalidomide [44].
Since drug-response was maintained in many patients despite the decreased CD38 expression by MM cells [44], events other than those just described may be responsible for treatment failure. Nonetheless, repeated treatment with DARA is possible and efficacious [43].
Therapeutic combination with agents that increase CD38 expression, such as panobinostat an–histone deacetylase inhibitor [45] and trans retinoic acid [32], can overcome these limitations as demonstrated by the increased of anti-CD38 mAb-mediated ADCC in vitro [32,45]. More details are reported in the following section.
Recently, Moreno L et al. [46] described that in vitro treatment of several human myeloma cell lines with another anti-CD38 mAb, isatuximab, induces CD38 internalization without its release from MM cell surface [46]. The authors found that isatuximab-mediated ADCC, ADCP, and CDC are triggered only in the presence of a certain number of surface CD38 molecules [46].
All together these data thus suggest that not all patients could benefit from the same anti-CD38 mAbs and combination with other therapeutic agents should be investigated.
4. Drug-Mediated Modulation of CD38 Expression by MM Cells
The expression of CD38 by MM cells is potentially modulated by different agents used in the treatment of MM or for other hematological malignancies.
Firstly, authors investigated the role of all-trans retinoic acid (ATRA) on the expression of CD38 by MM cells. Low doses of ATRA were able to increase up to 4.4 fold CD38 expression on MM cell lines. Accordingly, ATRA significantly increased CD38 expression in all the BM mononuclear cells of MM patients. Moreover, in a humanized mouse model ATRA enhances DARA mediated ADCC and CDC against MM cells either in vitro or in vivo [32]. Similarly, we recently observed that the inhibitory effect of DARA on OC formation is significantly enhanced by ATRA treatment. This treatment increased CD38 expression by monocytes and early OC progenitors [31].
There are different mechanisms involved in the modulation of CD38 by ATRA. Previous studies have demonstrated that the retinoic acid receptor has an important role in the induction of CD38 by ATRA [47]; indeed, the CD38 gene contains a retinoic acid-responsive element. Non-classical retinoic acid signaling is also involved in CD38 up-regulation, independently from the conventional retinoic acid receptor pathway [48]. Among these mechanisms, signaling mediated by protein kinase Cδ [48] and phosphatidylinositol 3-kinase [49] have been described.
IMiDs are agents able to act on both MM and microenvironment cells by targeting several adhesion molecules [50]. Specifically, it was reported that the integrin-signaling pathway is significantly modulated by lenalidomide and pomalidomide in MM cell lines. In particular, CD38 was up regulated after IMiDs treatment whereas ITGA8 and ICAM2 (CD102) were both down-regulated [51]. More recently, Fedele P. et al. [52] reported that lenalidomide treatment increases CD38 expression in several human myeloma cell lines, through Ikaros and Aiolos degradation. Thereafter, the authors tested the hypothesis that the additive effect observed by the combination of anti-CD38 mAbs and IMiDs in vitro could be due to the upregulation of CD38 by IMiDs [52]. They found that the additive effect of the treatment combination was directly correlated with the increased CD38 surface expression on MM cells [52]. However, in the presence of lenalidomide it was not observed any additional effect on DARA direct cytotoxicity [52]. On the other hand, lenalidomide potentiated DARA-induced ADCC against MM cells by directly stimulating NK cells without modifying CD38 expression on these cells. Overall these evidences indicate that the clinical synergistic effect of the combination of lenalidomide and DARA is likely due either to the increased CD38 expression on MM cells or to the stimulation of NK activity [52].
Panobinostat is a pan-histone deacetylase inhibitor (HDACi) approved for the treatment of MM relapsed patients in combination with bortezomib [53]. Recent data indicate that panobinostat is able to up-regulate the expression of CD38 by primary PCs either in newly diagnosed MM or in relapsed MM patients [45]. Interestingly, this effect seems to be specific for MM cells and does not occur in lymphoma cells [45]. Consistently with the effect of panobinostat on the expression of CD38 by MM cells, it has been demonstrated that the cytotoxic effect of DARA was increased by the treatment of panobinostat [45]. A significant increase of ADCC was observed against panobinostat pre-treated cells compared to untreated myeloma cells in all BM samples obtained from MM patients [45]. Interestingly, panobinostat treatment did not affect CD38 expression on other cell types as T cells [45].
The mechanism by which panobinostat increases CD38 expression may involve the interaction between Ikaros and the nucleosome remodeling deacetylase (NuRD) complex, known to regulate several transcriptional events involved in oncogenesis and cancer progression [54]. NuRD contains at least two subunits with an ATP-dependent chromatin remodeling, together with HDAC1/2 subunits, which catalyze protein deacetylation [54]. Co-immunoprecipitation experiments demonstrated that Ikaros interacts with HDAC1/2 in MM cells, by sharing similar binding patterns in the CD38 locus [52]. It is thus conceivable that Ikaros-induced CD38 repression is mediated by its interaction with HDAC1/2. These results were further supported by the synergistic upregulation of CD38 after combination treatment with lenalidomide and low doses of HDAC- inhibitor panobinostat [52].
Together with the pan- HDACi, panobinostat, the effect of the class I HDAC and HDAC6 inhibitors were investigated. The class I HDAC-specific inhibitor is able to up-regulate CD38 expression by MM cells and to increase the effect of interferon (IFN)-α and ATRA, in several MM cell lines tested. On the other hand, HDAC6 inhibitor attenuated the CD38 expression induced by IFN-α and ATRA. Similarly, these authors found that panobinostat attenuates IFN-α and ATRA up-regulated expression of CD38 [55]. It is known that the upstream sequence of CD38 gene contains an interferon regulatory factor 1 (IRF1)-binding site [56,57] suggesting the role of STAT-1-IRF1 pathway activation in the up-regulation of CD38 by IFN-α. A dual opposite effect was thus observed regarding the different type of HDAC inhibitors [55]. Indeed, IFN-α and ATRA up-regulation of CD38 expression was attenuated by HDAC6 inhibitor, ACY-1215, but not by the class I HDAC1 inhibitor, MS-275 [55]. These in vitro evidences suggest that HDAC class I inhibitors could be used in combination with ATRA to potentiate the effect of anti-CD38 mAbs.
CD38 was identified as a differential methylating region in MM patients with a negative correlation between DNA methylation and gene expression. Moreover, analysis of different independent MM datasets showed an inverse relationship between normal PCs and malignant MM cells in terms of CD38 methylation and gene expression status [58]. Accordingly, it has been recently demonstrated by flow-cytometry that DNA methyltrasnferase inhibitors (DNMTi), as azacytidine (AZA) and decitabine (DEC), increase CD38 expression on a panel of human MM cell lines, without affecting cell viability [58]. Furthermore, the combination of AZA or DEC with ATRA, significantly increased surface CD38 expression as compared with the treatment with the single drugs [58]. Next, by using immortalized NK transgenic cell line, the authors showed a significant increase in DARA-mediated-ADCC against MM cells pre-treated with DNMTi as compared to not-treated cells [58]. The mechanism behind the AZA-mediated upregulation of CD38 expression by MM cells involved TNF-α signaling as demonstrated by the abrogation of DNMTi effect in the presence of neutralizing TNF-α antibody [58].
5. Clinical Results of Anti-CD38 mAbs in Combination with Agents Able to Upregulate CD38 Expression in MM Patients
Overall the evidences indicate that different pharmacological agents can modulate the expression of CD38 with a possible translational effect. Published data suggests that the use of IMiDs, ATRA, and panobinostat may increase the clinical efficacy of anti-CD38 as DARA, isatuximab and MOR202. The combination of DARA with IMIDs have been extensively tested in several studies on MM patients at different stages of disease. The outstanding efficacy of the combination of DARA with lenalidomide have led to a rapid approval of the regimen in relapsed/refractory MM patients, demonstrating an overall response rate (ORR) of 92.5% and a progression free survival (PFS) at 12 months of 83.2% [7]. The same regimen has been tried in the first-line setting, showing a complete response (CR) or better in 47.6% of patients, with a 70.6% of patients without disease progression at 30 months [59]. DARA has also been tested in combination with pomalidomide, showing promising results in a phase Ib trial, even in heavily pretreated patients and in patients exhibiting high-risk cytogenetic features (ORR: 60% in the whole population, 59% in high-risk patients, 55% in patients with >3 previous lines of therapy; median PFS: 8.8 months) [60]. Accordingly, it was reported the significant clinical effect of the combination anti-CD38 antibody isatuximab and lenalidomide. A phase Ib trial reported an ORR of 56% with a median PFS of 8.5 months, with no safety concerns [61]. The phase Ib trial of isatuximab in combination with pomalidomide showed even more promising results, with an ORR of 62% and a median PFS of 17.6 months in patients heavily pretreated [62]. The phase III randomized trial is currently ongoing. However, to our knowledge, none of these studies have evaluated the expression in vivo of CD38 on MM cells.
Based on the results of pre-clinical studies, a phase I/II clinical trial of DARA in combination with ATRA has been designed, even if no results have been published yet. Conversely, there is no evidence of possible clinical use of the combination of anti-CD38 mAbs and panobinostat. Results of main clinical trial of anti-CD38 antibodies in combination with drugs that potentially modulated the expression of CD38 by MM cells are reported in Table 1.
6. Conclusions
CD38 is a suitable target for immunotherapy in MM patients due to its expression profile in the BM microenvironment. MM cells expressed CD38 at high levels. On the other hand, among the cells of the BM microenvironment it has been demonstrated that NK, T cells, and monocyte express CD38 with different levels of expression. Growing evidence indicate that the efficacy of anti-CD38 mAbs is related, at least in part, to the CD38 intensity of expression by MM cells and those of the immune-microenvironment. The possibility to modulate CD38 increasing its expression by MM cells is the pre-requisite to potentiate the efficacy of anti-CD38 mAbs. Moreover, it has been shown that anti-CD38 mAbs may modulate the CD38 expression on the surface of MM cells by its internalization or capping.
Different pharmacological agents have demonstrated the capacity to increase the expression of CD38 by MM cells and their BM microenvironment. Particularly different experimental data indicate that ATRA is able to increase the expression of CD38. Among the anti-MM drugs, it has been shown that the HDAC inhibitor panobinostat increased CD38 expression by MM cells. The same effect has been found with lenalidomide and pomalidomide. More recently, it has been reported that DNMTi as AZA or DEC also increase CD38 expression by MM cells [58].
Figure 1 summarizes the main mechanisms involved in the modulation of CD38 expression in MM cells and in the BM microenvironment by different molecules with a possible therapeutic impact.
These observations provide the rational to design clinical trials using anti-CD38 mAbs such as DARA and isatuximab in combination with IMiDs, HDACi, and DNMTi. Clinical trial showed that the combination of DARA with IMiDs is highly clinical efficient to induce a profound response in relapsed/refractory MM patients.
Author Contributions
F.C., N.G. wrote the manuscript; F.C., N.G. and B.D.P. reviewed the manuscript.
Funding
F.C. is supported by a fellowship from Società Italiana di Ematologia Sperimentale (SIES); N.G. is supported by a grant from the Associazione Italiana per la Ricerca sul Cancro IG2017 n. 20299, the International Myeloma Foundation under 2018 Brian D. Novis Senior Research Grant and a grant from the Ministero della Salute Italiana PE-2016-02361261.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Anderson, K.C. Multiple myeloma. Hematol. Oncol. Clin. North. Am. 2014, 28, xi–xii. [Google Scholar] [CrossRef] [PubMed]
- Chillemi, A.; Zaccarello, G.; Quarona, V.; Ferracin, M.; Ghimenti, C.; Massaia, M.; Horenstein, A.L.; Malavasi, F. Anti-CD38 antibody therapy: Windows of opportunity yielded by the functional characteristics of the target molecule. Mol. Med. 2013, 19, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Lonial, S.; Weiss, B.M.; Usmani, S.Z.; Singhal, S.; Chari, A.; Bahlis, N.J.; Belch, A.; Krishnan, A.; Vescio, R.A.; Mateos, M.V.; et al. Daratumumab monotherapy in patients with treatment-refractory multiple myeloma (SIRIUS): An open-label, randomised, phase 2 trial. Lancet 2016, 387, 1551–1560. [Google Scholar] [CrossRef]
- Frerichs, K.A.; Nagy, N.A.; Lindenbergh, P.L.; Bosman, P.; Marin Soto, J.; Broekmans, M.; Groen, R.W.J.; Themeli, M.; Nieuwenhuis, L.; Stege, C.; et al. CD38-targeting antibodies in multiple myeloma: Mechanisms of action and clinical experience. Expert Rev. Clin. Immunol. 2018, 14, 197–206. [Google Scholar] [CrossRef]
- Oostvogels, R.; Jak, M.; Raymakers, R.; Mous, R.; Minnema, M.C. Efficacy of retreatment with immunomodulatory drugs and proteasome inhibitors following daratumumab monotherapy in relapsed and refractory multiple myeloma patients. Br. J. Haematol. 2018, 183, 60–67. [Google Scholar] [CrossRef]
- Usmani, S.Z.; Weiss, B.M.; Plesner, T.; Bahlis, N.J.; Belch, A.; Lonial, S.; Lokhorst, H.M.; Voorhees, P.M.; Richardson, P.G.; Chari, A.; et al. Clinical efficacy of daratumumab monotherapy in patients with heavily pretreated relapsed or refractory multiple myeloma. Blood 2016, 128, 37–44. [Google Scholar] [CrossRef]
- Dimopoulos, M.A.; Oriol, A.; Nahi, H.; San-Miguel, J.; Bahlis, N.J.; Usmani, S.Z.; Rabin, N.; Orlowski, R.Z.; Komarnicki, M.; Suzuki, K.; et al. Daratumumab, Lenalidomide, and Dexamethasone for Multiple Myeloma. New Engl. J. Med. 2016, 375, 1319–1331. [Google Scholar] [CrossRef] [Green Version]
- Nijhof, I.S.; Casneuf, T.; van Velzen, J.; van Kessel, B.; Axel, A.E.; Syed, K.; Groen, R.W.; van Duin, M.; Sonneveld, P.; Minnema, M.C.; et al. CD38 expression and complement inhibitors affect response and resistance to daratumumab therapy in myeloma. Blood 2016, 128, 959–970. [Google Scholar] [CrossRef] [Green Version]
- Kyle, R.A.; Rajkumar, S.V. Multiple myeloma. New Engl. J. Med. 2004, 351, 1860–1873. [Google Scholar] [CrossRef]
- Palumbo, A.; Anderson, K. Multiple myeloma. New Engl. J. Med. 2011, 364, 1046–1060. [Google Scholar] [CrossRef] [Green Version]
- Santonocito, A.M.; Consoli, U.; Bagnato, S.; Milone, G.; Palumbo, G.A.; Di Raimondo, F.; Stagno, F.; Guglielmo, P.; Giustolisi, R. Flow cytometric detection of aneuploid CD38(++) plasmacells and CD19(+) B-lymphocytes in bone marrow, peripheral blood and PBSC harvest in multiple myeloma patients. Leuk. Res. 2004, 28, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Atanackovic, D.; Steinbach, M.; Radhakrishnan, S.V.; Luetkens, T. Immunotherapies targeting CD38 in Multiple Myeloma. Oncoimmunology 2016, 5, e1217374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokhorst, H.M.; Plesner, T.; Laubach, J.P.; Nahi, H.; Gimsing, P.; Hansson, M.; Minnema, M.C.; Lassen, U.; Krejcik, J.; Palumbo, A.; et al. Targeting CD38 with Daratumumab Monotherapy in Multiple Myeloma. New Engl. J. Med. 2015, 373, 1207–1219. [Google Scholar] [CrossRef] [PubMed]
- Harada, H.; Kawano, M.M.; Huang, N.; Harada, Y.; Iwato, K.; Tanabe, O.; Tanaka, H.; Sakai, A.; Asaoku, H.; Kuramoto, A. Phenotypic difference of normal plasma cells from mature myeloma cells. Blood 1993, 81, 2658–2663. [Google Scholar] [CrossRef] [PubMed]
- Bataille, R.; Jego, G.; Robillard, N.; Barille-Nion, S.; Harousseau, J.L.; Moreau, P.; Amiot, M.; Pellat-Deceunynck, C. The phenotype of normal, reactive and malignant plasma cells. Identification of “many and multiple myelomas” and of new targets for myeloma therapy. Haematologica 2006, 91, 1234–1240. [Google Scholar]
- Krejcik, J.; Casneuf, T.; Nijhof, I.S.; Verbist, B.; Bald, J.; Plesner, T.; Syed, K.; Liu, K.; van de Donk, N.W.; Weiss, B.M.; et al. Daratumumab depletes CD38+ immune regulatory cells, promotes T-cell expansion, and skews T-cell repertoire in multiple myeloma. Blood 2016, 128, 384–394. [Google Scholar] [CrossRef] [Green Version]
- Malavasi, F.; Deaglio, S.; Funaro, A.; Ferrero, E.; Horenstein, A.L.; Ortolan, E.; Vaisitti, T.; Aydin, S. Evolution and function of the ADP ribosyl cyclase/CD38 gene family in physiology and pathology. Physiol. Rev. 2008, 88, 841–886. [Google Scholar] [CrossRef] [Green Version]
- Deaglio, S.; Aydin, S.; Vaisitti, T.; Bergui, L.; Malavasi, F. CD38 at the junction between prognostic marker and therapeutic target. Trends Mol. Med. 2008, 14, 210–218. [Google Scholar] [CrossRef]
- Deaglio, S.; Mallone, R.; Baj, G.; Arnulfo, A.; Surico, N.; Dianzani, U.; Mehta, K.; Malavasi, F. CD38/CD31, a receptor/ligand system ruling adhesion and signaling in human leukocytes. Chem. Immunol. 2000, 75, 99–120. [Google Scholar]
- Deaglio, S.; Morra, M.; Mallone, R.; Ausiello, C.M.; Prager, E.; Garbarino, G.; Dianzani, U.; Stockinger, H.; Malavasi, F. Human CD38 (ADP-ribosyl cyclase) is a counter-receptor of CD31, an Ig superfamily member. J. Immunol. 1998, 160, 395–402. [Google Scholar]
- Watt, S.M.; Gschmeissner, S.E.; Bates, P.A. PECAM-1: Its expression and function as a cell adhesion molecule on hemopoietic and endothelial cells. Leuk. Lymphoma 1995, 17, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Vallario, A.; Chilosi, M.; Adami, F.; Montagna, L.; Deaglio, S.; Malavasi, F.; Caligaris-Cappio, F. Human myeloma cells express the CD38 ligand CD31. Br. J. Haematol. 1999, 105, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Accardi, F.; Notarfranchi, L.; Palma, B.D.; Manfra, I.; De Luca, F.; Mancini, C.; Martella, E.; Marchica, V.; Storti, P.; Bolzoni, M.; et al. The Loss of Cd38 Expression by Myeloma Plasma Cells May Occur in the Extramedullary Disease. Haematologica 2018, 103, S54. [Google Scholar]
- Aarhus, R.; Graeff, R.M.; Dickey, D.M.; Walseth, T.F.; Lee, H.C. ADP-ribosyl cyclase and CD38 catalyze the synthesis of a calcium-mobilizing metabolite from NADP. J. Biol. Chem. 1995, 270, 30327–30333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horenstein, A.L.; Chillemi, A.; Zaccarello, G.; Bruzzone, S.; Quarona, V.; Zito, A.; Serra, S.; Malavasi, F. A CD38/CD203a/CD73 ectoenzymatic pathway independent of CD39 drives a novel adenosinergic loop in human T lymphocytes. Oncoimmunology 2013, 2, e26246. [Google Scholar] [CrossRef] [Green Version]
- Horenstein, A.L.; Quarona, V.; Toscani, D.; Costa, F.; Chillemi, A.; Pistoia, V.; Giuliani, N.; Malavasi, F. Adenosine Generated in the Bone Marrow Niche Through a CD38-Mediated Pathway Correlates with Progression of Human Myeloma. Mol. Med. 2016, 22. [Google Scholar] [CrossRef] [Green Version]
- Morandi, F.; Marimpietri, D.; Horenstein, A.L.; Bolzoni, M.; Toscani, D.; Costa, F.; Castella, B.; Faini, A.C.; Massaia, M.; Pistoia, V.; et al. Microvesicles released from multiple myeloma cells are equipped with ectoenzymes belonging to canonical and non-canonical adenosinergic pathways and produce adenosine from ATP and NAD+. Oncoimmunology 2018, 7, e1458809. [Google Scholar] [CrossRef] [Green Version]
- Horenstein, A.L.; Chillemi, A.; Quarona, V.; Zito, A.; Roato, I.; Morandi, F.; Marimpietri, D.; Bolzoni, M.; Toscani, D.; Oldham, R.J.; et al. NAD+-Metabolizing Ectoenzymes in Remodeling Tumor-Host Interactions: The Human Myeloma Model. Cells 2015, 4, 520–537. [Google Scholar] [CrossRef] [Green Version]
- Abe, M.; Hiura, K.; Wilde, J.; Shioyasono, A.; Moriyama, K.; Hashimoto, T.; Kido, S.; Oshima, T.; Shibata, H.; Ozaki, S.; et al. Osteoclasts enhance myeloma cell growth and survival via cell-cell contact: A vicious cycle between bone destruction and myeloma expansion. Blood 2004, 104, 2484–2491. [Google Scholar] [CrossRef]
- Quarona, V.; Ferri, V.; Chillemi, A.; Bolzoni, M.; Mancini, C.; Zaccarello, G.; Roato, I.; Morandi, F.; Marimpietri, D.; Faccani, G.; et al. Unraveling the contribution of ectoenzymes to myeloma life and survival in the bone marrow niche. Ann. New York Acad. Sci. 2015, 1335, 10–22. [Google Scholar] [CrossRef]
- Costa, F.; Toscani, D.; Chillemi, A.; Quarona, V.; Bolzoni, M.; Marchica, V.; Vescovini, R.; Mancini, C.; Martella, E.; Campanini, N.; et al. Expression of CD38 in myeloma bone niche: A rational basis for the use of anti-CD38 immunotherapy to inhibit osteoclast formation. Oncotarget 2017, 8, 56598–56611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijhof, I.S.; Groen, R.W.; Lokhorst, H.M.; van Kessel, B.; Bloem, A.C.; van Velzen, J.; de Jong-Korlaar, R.; Yuan, H.; Noort, W.A.; Klein, S.K.; et al. Upregulation of CD38 expression on multiple myeloma cells by all-trans retinoic acid improves the efficacy of daratumumab. Leukemia 2015, 29, 2039–2049. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Adebanjo, O.A.; Moonga, B.S.; Corisdeo, S.; Anandatheerthavarada, H.K.; Biswas, G.; Arakawa, T.; Hakeda, Y.; Koval, A.; Sodam, B.; et al. CD38/ADP-ribosyl cyclase: A new role in the regulation of osteoclastic bone resorption. J. Cell Biol. 1999, 146, 1161–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Iqbal, J.; Dolgilevich, S.; Yuen, T.; Wu, X.B.; Moonga, B.S.; Adebanjo, O.A.; Bevis, P.J.; Lund, F.; Huang, C.L.; et al. Disordered osteoclast formation and function in a CD38 (ADP-ribosyl cyclase)-deficient mouse establishes an essential role for CD38 in bone resorption. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Adebanjo, O.A.; Koval, A.; Anandatheerthavarada, H.K.; Iqbal, J.; Wu, X.Y.; Moonga, B.S.; Wu, X.B.; Biswas, G.; Bevis, P.J.; et al. A novel mechanism for coupling cellular intermediary metabolism to cytosolic Ca2+ signaling via CD38/ADP-ribosyl cyclase, a putative intracellular NAD+ sensor. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2002, 16, 302–314. [Google Scholar] [CrossRef] [Green Version]
- Grassi, F.; Manferdini, C.; Cattini, L.; Piacentini, A.; Gabusi, E.; Facchini, A.; Lisignoli, G. T cell suppression by osteoclasts in vitro. J. Cell. Physiol. 2011, 226, 982–990. [Google Scholar] [CrossRef]
- Takayanagi, H.; Ogasawara, K.; Hida, S.; Chiba, T.; Murata, S.; Sato, K.; Takaoka, A.; Yokochi, T.; Oda, H.; Tanaka, K.; et al. T-cell-mediated regulation of osteoclastogenesis by signalling cross-talk between RANKL and IFN-gamma. Nature 2000, 408, 600–605. [Google Scholar] [CrossRef]
- An, G.; Acharya, C.; Feng, X.; Wen, K.; Zhong, M.; Zhang, L.; Munshi, N.C.; Qiu, L.; Tai, Y.T.; Anderson, K.C. Osteoclasts promote immune suppressive microenvironment in multiple myeloma: Therapeutic implication. Blood 2016, 128, 1590–1603. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Zhang, L.; Acharya, C.; An, G.; Wen, K.; Qiu, L.; Munshi, N.C.; Tai, Y.T.; Anderson, K.C. Targeting CD38 Suppresses Induction and Function of T Regulatory Cells to Mitigate Immunosuppression in Multiple Myeloma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 4290–4300. [Google Scholar] [CrossRef] [Green Version]
- Van de Donk, N.W.; Janmaat, M.L.; Mutis, T.; Lammerts van Bueren, J.J.; Ahmadi, T.; Sasser, A.K.; Lokhorst, H.M.; Parren, P.W. Monoclonal antibodies targeting CD38 in hematological malignancies and beyond. Immunol. Rev. 2016, 270, 95–112. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Acharya, C.; An, G.; Zhong, M.; Feng, X.; Wang, L.; Dasilva, N.; Song, Z.; Yang, G.; Adrian, F.; et al. SAR650984 directly induces multiple myeloma cell death via lysosomal-associated and apoptotic pathways, which is further enhanced by pomalidomide. Leukemia 2016, 30, 399–408. [Google Scholar] [CrossRef]
- Van de Donk, N. Immunomodulatory effects of CD38-targeting antibodies. Immunol. Lett. 2018, 199, 16–22. [Google Scholar] [CrossRef]
- Alici, E.; Chrobok, M.; Lund, J.; Ahmadi, T.; Khan, I.; Duru, A.D.; Nahi, H. Re-challenging with anti-CD38 monotherapy in triple-refractory multiple myeloma patients is a feasible and safe approach. Br. J. Haematol. 2016, 174, 473–477. [Google Scholar] [CrossRef] [Green Version]
- Krejcik, J.; Frerichs, K.A.; Nijhof, I.S.; van Kessel, B.; van Velzen, J.F.; Bloem, A.C.; Broekmans, M.E.C.; Zweegman, S.; van Meerloo, J.; Musters, R.J.P.; et al. Monocytes and Granulocytes Reduce CD38 Expression Levels on Myeloma Cells in Patients Treated with Daratumumab. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 7498–7511. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Guerrero, E.; Gogishvili, T.; Danhof, S.; Schreder, M.; Pallaud, C.; Perez-Simon, J.A.; Einsele, H.; Hudecek, M. Panobinostat induces CD38 upregulation and augments the antimyeloma efficacy of daratumumab. Blood 2017, 129, 3386–3388. [Google Scholar] [CrossRef]
- Moreno, L.; Perez, C.; Zabaleta, A.; Manrique, I.; Alignani, D.; Ajona, D.; Blanco, L.; Lasa, M.; Maiso, P.; Rodriguez, I.; et al. The Mechanism of Action of the Anti-CD38 Monoclonal Antibody Isatuximab in Multiple Myeloma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2019, 25, 3176–3187. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, H.; Hoshino, S.; Ohori, M.; Kontani, K.; Nishina, H.; Suzawa, M.; Kato, S.; Katada, T. Molecular mechanism of human CD38 gene expression by retinoic acid. Identification of retinoic acid response element in the first intron. J. Biol. Chem. 1998, 273, 15429–15434. [Google Scholar] [CrossRef] [Green Version]
- Uruno, A.; Noguchi, N.; Matsuda, K.; Nata, K.; Yoshikawa, T.; Chikamatsu, Y.; Kagechika, H.; Harigae, H.; Ito, S.; Okamoto, H.; et al. All-trans retinoic acid and a novel synthetic retinoid tamibarotene (Am80) differentially regulate CD38 expression in human leukemia HL-60 cells: Possible involvement of protein kinase C-delta. J. Leukoc. Biol. 2011, 90, 235–247. [Google Scholar] [CrossRef]
- Lewandowski, D.; Linassier, C.; Iochmann, S.; Degenne, M.; Domenech, J.; Colombat, P.; Binet, C.; Herault, O. Phosphatidylinositol 3-kinases are involved in the all-trans retinoic acid-induced upregulation of CD38 antigen on human haematopoietic cells. Br. J. Haematol. 2002, 118, 535–544. [Google Scholar] [CrossRef]
- Quach, H.; Ritchie, D.; Stewart, A.K.; Neeson, P.; Harrison, S.; Smyth, M.J.; Prince, H.M. Mechanism of action of immunomodulatory drugs (IMiDS) in multiple myeloma. Leukemia 2010, 24, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Bolzoni, M.; Storti, P.; Bonomini, S.; Todoerti, K.; Guasco, D.; Toscani, D.; Agnelli, L.; Neri, A.; Rizzoli, V.; Giuliani, N. Immunomodulatory drugs lenalidomide and pomalidomide inhibit multiple myeloma-induced osteoclast formation and the RANKL/OPG ratio in the myeloma microenvironment targeting the expression of adhesion molecules. Exp. Hematol. 2013, 41, 387–397 e381. [Google Scholar] [CrossRef] [PubMed]
- Fedele, P.L.; Willis, S.N.; Liao, Y.; Low, M.S.; Rautela, J.; Segal, D.H.; Gong, J.N.; Huntington, N.D.; Shi, W.; Huang, D.C.S.; et al. IMiDs prime myeloma cells for daratumumab-mediated cytotoxicity through loss of Ikaros and Aiolos. Blood 2018, 132, 2166–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratt, G. Panobinostat plus bortezomib and dexamethasone for relapsed myeloma. Lancet Haematol. 2016, 3, e498–e499. [Google Scholar] [CrossRef]
- Lai, A.Y.; Wade, P.A. Cancer biology and NuRD: A multifaceted chromatin remodelling complex. Nat. Rev. Cancer 2011, 11, 588–596. [Google Scholar] [CrossRef]
- Bat-Erdene, A.; Nakamura, S.; Oda, A.; Iwasa, M.; Teramachi, J.; Ashtar, M.; Harada, T.; Miki, H.; Tenshin, H.; Hiasa, M.; et al. Class 1 HDAC and HDAC6 inhibition inversely regulates CD38 induction in myeloma cells via interferon-alpha and ATRA. Br. J. Haematol. 2019, 185, 969–974. [Google Scholar] [CrossRef]
- Ferrero, E.; Malavasi, F. Human CD38, a leukocyte receptor and ectoenzyme, is a member of a novel eukaryotic gene family of nicotinamide adenine dinucleotide+-converting enzymes: Extensive structural homology with the genes for murine bone marrow stromal cell antigen 1 and aplysian ADP-ribosyl cyclase. J. Immunol. 1997, 159, 3858–3865. [Google Scholar]
- Bauvois, B.; Durant, L.; Laboureau, J.; Barthelemy, E.; Rouillard, D.; Boulla, G.; Deterre, P. Upregulation of CD38 gene expression in leukemic B cells by interferon types I and II. J. Interferon Cytokine Res. Off. J. Int. Soc. Interferon Cytokine Res. 1999, 19, 1059–1066. [Google Scholar] [CrossRef]
- Choudhry, P.; Mariano, M.C.; Geng, H.; Martin, T.G., 3rd; Wolf, J.L.; Wong, S.W.; Shah, N.; Wiita, A.P. DNA methyltransferase inhibitors upregulate CD38 protein expression and enhance daratumumab efficacy in multiple myeloma. Leukemia 2019. [Google Scholar] [CrossRef]
- Facon, T.; Kumar, S.; Plesner, T.; Orlowski, R.Z.; Moreau, P.; Bahlis, N.; Basu, S.; Nahi, H.; Hulin, C.; Quach, H.; et al. Daratumumab plus Lenalidomide and Dexamethasone for Untreated Myeloma. New Engl. J. Med. 2019, 380, 2104–2115. [Google Scholar] [CrossRef]
- Chari, A.; Suvannasankha, A.; Fay, J.W.; Arnulf, B.; Kaufman, J.L.; Ifthikharuddin, J.J.; Weiss, B.M.; Krishnan, A.; Lentzsch, S.; Comenzo, R.; et al. Daratumumab plus pomalidomide and dexamethasone in relapsed and/or refractory multiple myeloma. Blood 2017, 130, 974–981. [Google Scholar] [CrossRef]
- Martin, T.; Baz, R.; Benson, D.M.; Lendvai, N.; Wolf, J.; Munster, P.; Lesokhin, A.M.; Wack, C.; Charpentier, E.; Campana, F.; et al. A phase 1b study of isatuximab plus lenalidomide and dexamethasone for relapsed/refractory multiple myeloma. Blood 2017, 129, 3294–3303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikhael, J.; Richardson, P.; Usmani, S.Z.; Raje, N.; Bensinger, W.; Karanes, C.; Campana, F.; Kanagavel, D.; Dubin, F.; Liu, Q.; et al. A phase 1b study of isatuximab plus pomalidomide/dexamethasone in relapsed/refractory multiple myeloma. Blood 2019, 134, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
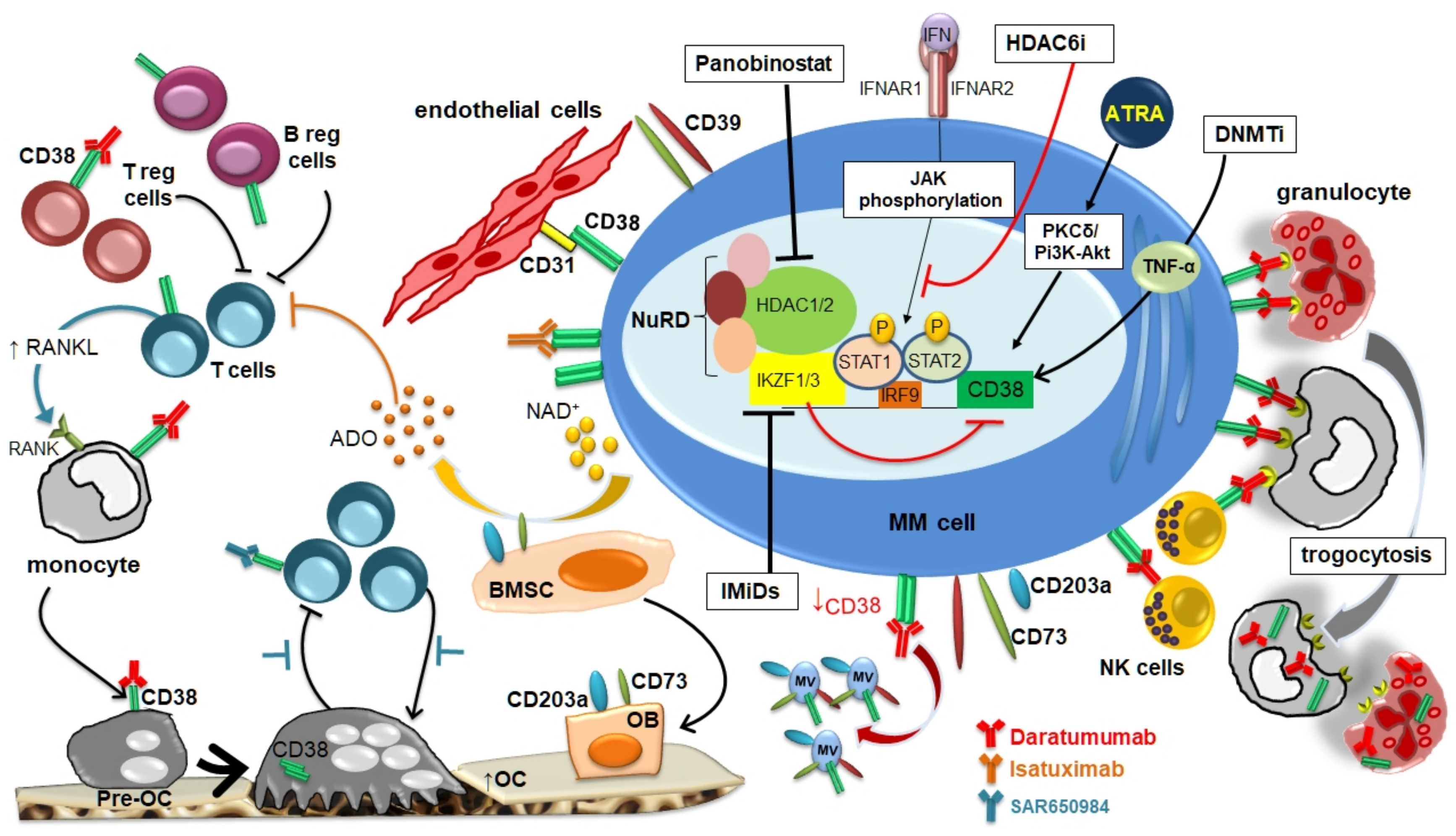
Figure 1.
CD38 expression in multiple myeloma (MM) microenvironment and its modulation by different agents.
Figure 1.
CD38 expression in multiple myeloma (MM) microenvironment and its modulation by different agents.


{kind=link}
Table 1.
Clinical trial of combination of anti-CD38 mAbs with drugs able to modulate CD38 expression.
Table 1.
Clinical trial of combination of anti-CD38 mAbs with drugs able to modulate CD38 expression.
| Drug Combination | Trial Name (NCT number) [Ref.] | Phase | Number of Patients | Primary Outcome | Results Reported | AEs (Grade 3–4) |
|---|---|---|---|---|---|---|
| DARA/ Lenalidomide | 54767414MMY3008 (NCT02252172) [59] | III | 737 | PFS | PFS 70.6% at 30 mos | Neutropenia 50%, lymphopenia 15% |
| DARA/ Lenalidomide | Pollux Study (NCT02076009) [7] | III | 569 | PFS | ORR 92.9% vs 76.4% Median PFS nr vs 17.5 months (HR 0.44) | Neutropenia (54%), anemia (15.5%), pneumonia (12%) |
| Isatuximab/ Lenalidomide | A Phase 1b Study of SAR650984 (Anti-CD38 mAb) in Combination with Len and Dex for the Treatment of RRMM (NCT01749969) [61] | I | 57 | MTD of the combination | ORR 56% Median PFS 8.5 months | Neutropenia (60%), lymphopenia (58%) |
| DARA/Pomalidomide | 54767414MMY1001 (NCT01998971) [60] | Ib | 103 | MTD of the combination | ORR 60% Median PFS 8.8 months | Neutropenia (77%), anemia (28%), thrombocytopenia (19%) |
| Isatuximab/ Pomalidomide | TCD14079 (NCT02283775) [62] | Ib | 45 | MTD of the combination | ORR 62% Median PFS 17.6 months | (AEs all grade) Fatigue (62%), upper respiratory tract infection (42%) |
| DARA/ ATRA | A Phase 1 and Phase 2 Study of DARA in Combination with ATRA in RRMM (NCT02751255) | I/II | 60 | 1) MTD 2) ORR 3) RDL | No result posted | No result posted |
Abbreviations: DARA, Daratumumab; ATRA, All Trans-Retinoic Acid; RRMM, Relapsed/Refractory Multiple Myeloma; MTD, Maximum Tolerated Dose; ORR, Overall Response Rate; RDL, Recommended phase 2 dose level; PFS, Progression Free Survival; HR, Hazard Ratio; CBR, Clinical Benefit Rate; AEs, Adverse events.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Costa, F.; Dalla Palma, B.; Giuliani, N. CD38 Expression by Myeloma Cells and Its Role in the Context of Bone Marrow Microenvironment: Modulation by Therapeutic Agents. Cells 2019, 8, 1632. https://doi.org/10.3390/cells8121632
AMA Style
Costa F, Dalla Palma B, Giuliani N. CD38 Expression by Myeloma Cells and Its Role in the Context of Bone Marrow Microenvironment: Modulation by Therapeutic Agents. Cells. 2019; 8(12):1632. https://doi.org/10.3390/cells8121632
Chicago/Turabian StyleCosta, Federica, Benedetta Dalla Palma, and Nicola Giuliani. 2019. "CD38 Expression by Myeloma Cells and Its Role in the Context of Bone Marrow Microenvironment: Modulation by Therapeutic Agents" Cells 8, no. 12: 1632. https://doi.org/10.3390/cells8121632
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.