Cell-Free Protein Synthesis: Chassis toward the Minimal Cell


Abstract
:1. Introduction
2. Regulation of the CFPS System
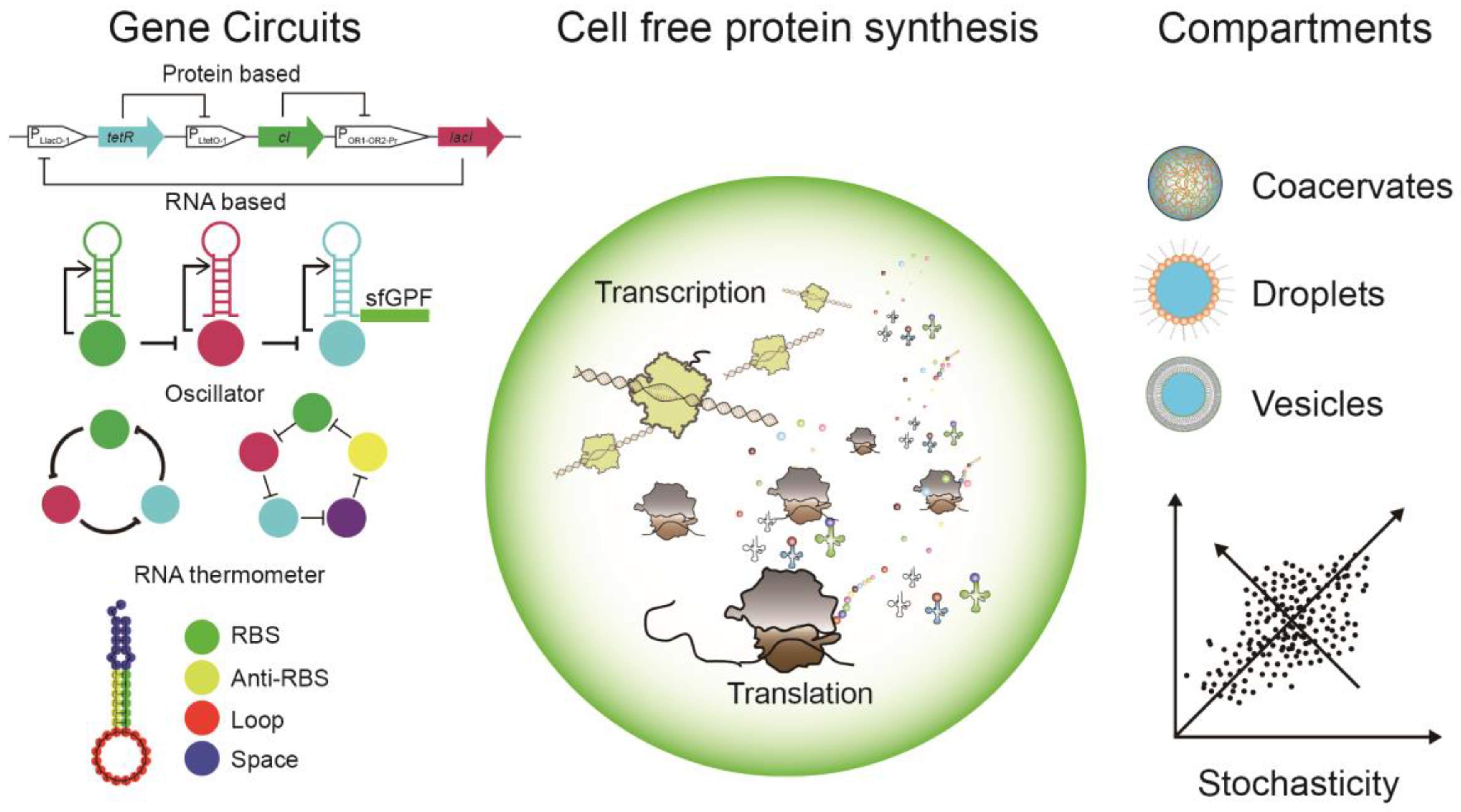
2.1. Genetic Circuits
2.2. Protein Based Gene Circuits
2.3. RNA-Based Gene Circuits
3. Programming Spatiotemporal Patterns—Toward the Minimal Cell Division System
4. Toward Self-Organization in CFPS Systems
5. Compartmentalization
6. Stochasticity
7. Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Schwille, P.; Spatz, J.; Landfester, K.; Bodenschatz, E.; Herminghaus, S.; Sourjik, V.; Erb, T.; Bastiaens, P.; Lipowsky, R.; Hyman, A.; et al. MaxSynBio - Avenues towards creating cells from the bottom up. Angew. Chem. Int. Ed. Engl. 2018, 13382–13392. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.Y.; Heymann, M.; Bernhard, F.; Schwille, P.; Kai, L. Cell-free protein synthesis in micro compartments: Building a minimal cell from biobricks. New Biotechnol. 2017, 39, 199–205. [Google Scholar] [CrossRef]
- Caschera, F.; Noireaux, V. Integration of biological parts toward the synthesis of a minimal cell. Curr. Opin. Chem. Biol. 2014, 22, 85–91. [Google Scholar] [CrossRef]
- Yewdall, N.A.; Mason, A.F.; van Hest, J.C.M. The hallmarks of living systems: Towards creating artificial cells. Interface Focus 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Luisi, P.L.; Stano, P. Synthetic Biology: Minimal cell mimicry. Nat. Chem. 2011, 3, 755–756. [Google Scholar] [CrossRef] [PubMed]
- Forster, A.C.; Church, G.M. Towards synthesis of a minimal cell. Mol. Syst. Biol. 2006, 2, 45. [Google Scholar] [CrossRef]
- Jewett, M.C.; Forster, A.C. Update on designing and building minimal cells. Curr. Opin. Biotechnol. 2010, 21, 697–703. [Google Scholar] [CrossRef]
- Nirenberg, M.W.; Matthaei, J.H. The dependence of cell-free protein synthesis in E. coli upon naturally occurring or synthetic polyribonucleotides. Proc. Natl. Acad. Sci. USA 1961, 47, 1588–1602. [Google Scholar] [CrossRef]
- Casteleijn, M.G.; Urtti, A.; Sarkhel, S. Expression without boundaries: Cell-free protein synthesis in pharmaceutical research. Int. J. Pharm. 2013, 440, 39–47. [Google Scholar] [CrossRef]
- Rues, R.B.; Dotsch, V.; Bernhard, F. Co-translational formation and pharmacological characterization of beta1-adrenergic receptor/nanodisc complexes with different lipid environments. Biochim. Biophys. Acta 2016, 1858, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Carlson, E.D.; Gan, R.; Hodgman, C.E.; Jewett, M.C. Cell-free protein synthesis: Applications come of age. Biotechnol. Adv. 2012, 30, 1185–1194. [Google Scholar] [CrossRef]
- Zubay, G. In-Vitro Synthesis of Protein in Microbial Systems. Annu. Rev. Genet. 1973, 7, 267–287. [Google Scholar] [CrossRef]
- Spirin, A.S.; Baranov, V.I.; Ryabova, L.A.; Ovodov, S.Y.; Alakhov, Y.B. A continuous cell-free translation system capable of producing polypeptides in high yield. Science 1988, 242, 1162–1164. [Google Scholar] [CrossRef]
- Schwarz, D.; Junge, F.; Durst, F.; Frolich, N.; Schneider, B.; Reckel, S.; Sobhanifar, S.; Dotsch, V.; Bernhard, F. Preparative scale expression of membrane proteins in Escherichia coli-based continuous exchange cell-free systems. Nat. Protoc. 2007, 2, 2945–2957. [Google Scholar] [CrossRef]
- Schwarz, D.; Dotsch, V.; Bernhard, F. Production of membrane proteins using cell-free expression systems. Proteomics 2008, 8, 3933–3946. [Google Scholar] [CrossRef]
- Bernhard, F.; Tozawa, Y. Cell-free expression--making a mark. Curr. Opin. Struct. Biol. 2013, 23, 374–380. [Google Scholar] [CrossRef]
- Jewett, M.C.; Calhoun, K.A.; Voloshin, A.; Wuu, J.J.; Swartz, J.R. An integrated cell-free metabolic platform for protein production and synthetic biology. Mol. Syst. Biol. 2008, 4, 220. [Google Scholar] [CrossRef]
- Jewett, M.C. Cell-free synthetic biology special issue. ACS Synth. Biol. 2014, 3, 332. [Google Scholar] [CrossRef]
- Mansy, S.S.; Szostak, J.W. Reconstructing the emergence of cellular life through the synthesis of model protocells. Cold Spring Harb. Symp. Quant. Biol. 2009, 74, 47–54. [Google Scholar] [CrossRef]
- Szostak, J.W.; Bartel, D.P.; Luisi, P.L. Synthesizing life. Nature 2001, 409, 387–390. [Google Scholar] [CrossRef]
- Trifonov, E.N. Vocabulary of definitions of life suggests a definition. J. Biomol. Struct. Dyn. 2011, 29, 259–266. [Google Scholar] [CrossRef]
- Sarma, R.H. A conversation on definition of life. J. Biomol. Struct. Dyn. 2012, 29, 597–598. [Google Scholar] [CrossRef]
- Takahashi, M.K.; Hayes, C.A.; Chappell, J.; Sun, Z.Z.; Murray, R.M.; Noireaux, V.; Lucks, J.B. Characterizing and prototyping genetic networks with cell-free transcription-translation reactions. Methods 2015, 86, 60–72. [Google Scholar] [CrossRef]
- Werner, E. An introduction to systems biology: Design principles of biological circuits. Nature 2007, 446, 493–494. [Google Scholar] [CrossRef]
- Mcadams, H.H.; Shapiro, L. Circuit Simulation of Genetic Networks. Science 1995, 269, 650–656. [Google Scholar] [CrossRef] [PubMed]
- McAdams, H.H.; Arkin, A. Gene regulation: Towards a circuit engineering discipline. Curr. Biol. 2000, 10, R318–R320. [Google Scholar] [CrossRef]
- Elowitz, M.B.; Leibler, S. A synthetic oscillatory network of transcriptional regulators. Nature 2000, 403, 335–338. [Google Scholar] [CrossRef]
- Lim, W.A. Designing customized cell signalling circuits. Nat. Rev. Mol. Cell Biol. 2010, 11, 393–403. [Google Scholar] [CrossRef]
- Nandagopal, N.; Elowitz, M.B. Synthetic Biology: Integrated Gene Circuits. Science 2011, 333, 1244–1248. [Google Scholar] [CrossRef]
- Noireaux, V.; Bar-Ziv, R.; Libchaber, A. Principles of cell-free genetic circuit assembly. Proc. Natl. Acad. Sci. USA 2003, 100, 12672–12677. [Google Scholar] [CrossRef]
- Shin, J.; Noireaux, V. Efficient cell-free expression with the endogenous E. Coli RNA polymerase and sigma factor 70. J. Biol. Eng. 2010, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Chappell, J.; Jensen, K.; Freemont, P.S. Validation of an entirely in vitro approach for rapid prototyping of DNA regulatory elements for synthetic biology. Nucleic Acids Res. 2013, 41, 3471–3481. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.Z.; Yeung, E.; Hayes, C.A.; Noireaux, V.; Murray, R.M. Linear DNA for Rapid Prototyping of Synthetic Biological Circuits in an Escherichia coli Based TX-TL Cell-Free System. ACS Synth. Biol. 2014, 3, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Niederholtmeyer, H.; Sun, Z.Z.; Hori, Y.; Yeung, E.; Verpoorte, A.; Murray, R.M.; Maerkl, S.J. Rapid cell-free forward engineering of novel genetic ring oscillators. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Hockenberry, A.J.; Jewett, M.C. Synthetic in vitro circuits. Curr. Opin. Chem. Biol. 2012, 16, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Karig, D.K.; Iyer, S.; Simpson, M.L.; Doktycz, M.J. Expression optimization and synthetic gene networks in cell-free systems. Nucleic Acids Res. 2012, 40, 3763–3774. [Google Scholar] [CrossRef]
- Shin, J.; Noireaux, V. An E. coli cell-free expression toolbox: Application to synthetic gene circuits and artificial cells. ACS Synth. Biol. 2012, 1, 29–41. [Google Scholar] [CrossRef]
- Garamella, J.; Marshall, R.; Rustad, M.; Noireaux, V. The All E. coli TX-TL Toolbox 2.0: A Platform for Cell-Free Synthetic Biology. ACS Synth. Biol. 2016, 5, 344–355. [Google Scholar] [CrossRef]
- Siegal-Gaskins, D.; Tuza, Z.A.; Kim, J.; Noireaux, V.; Murray, R.M. Gene Circuit Performance Characterization and Resource Usage in a Cell-Free “Breadboard”. ACS Synth. Biol. 2014, 3, 416–425. [Google Scholar] [CrossRef]
- Shin, J.; Noireaux, V. Study of messenger RNA inactivation and protein degradation in an Escherichia coli cell-free expression system. J. Biol. Eng. 2010, 4, 9. [Google Scholar] [CrossRef]
- Karzbrun, E.; Shin, J.; Bar-Ziv, R.H.; Noireaux, V. Coarse-Grained Dynamics of Protein Synthesis in a Cell-Free System. Phys. Rev. Lett. 2011, 106. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.K.; Chappell, J.; Hayes, C.A.; Sun, Z.Z.; Kim, J.; Singhal, V.; Spring, K.J.; Al-Khabouri, S.; Fall, C.P.; Noireaux, V.; et al. Rapidly Characterizing the Fast Dynamics of RNA Genetic Circuitry with Cell-Free Transcription Translation (TX-TL) Systems. ACS Synth. Biol. 2015, 4, 503–515. [Google Scholar] [CrossRef]
- Chappell, J.; Takahashi, M.K.; Lucks, J.B. Creating small transcription activating RNAs. Nat. Chem. Biol. 2015, 11, 214-U165. [Google Scholar] [CrossRef]
- Sadler, F.W.; Dodevski, I.; Sarkar, C.A. RNA Thermometers for the PURExpress System. ACS Synth. Biol. 2018, 7, 292–296. [Google Scholar] [CrossRef]
- Chizzolini, F.; Forlin, M.; Cecchi, D.; Mansy, S.S. Gene position more strongly influences cell-free protein expression from operons than T7 transcriptional promoter strength. ACS Synth. Biol. 2014, 3, 363–371. [Google Scholar] [CrossRef]
- Ogawa, A. Rational design of artificial riboswitches based on ligand-dependent modulation of internal ribosome entry in wheat germ extract and their applications as label-free biosensors. RNA 2011, 17, 478–488. [Google Scholar] [CrossRef]
- DebRoy, S.; Gebbie, M.; Ramesh, A.; Goodson, J.R.; Cruz, M.R.; van Hoof, A.; Winkler, W.C.; Garsin, D.A. A riboswitch-containing sRNA controls gene expression by sequestration of a response regulator. Science 2014, 345, 937–940. [Google Scholar] [CrossRef]
- Wieland, M.; Hartig, J.S. Artificial riboswitches: Synthetic mRNA-based regulators of gene expression. ChemBioChem 2008, 9, 1873–1878. [Google Scholar] [CrossRef]
- Weigand, J.E.; Suess, B. Aptamers and riboswitches: Perspectives in biotechnology. Appl. Microbiol. Biotechnol. 2009, 85, 229–236. [Google Scholar] [CrossRef]
- Winkler, W.C. Riboswitches and the role of noncoding RNAs in bacterial metabolic control. Curr. Opin. Chem. Biol. 2005, 9, 594–602. [Google Scholar] [CrossRef]
- Martini, L.; Mansy, S.S. Cell-like systems with riboswitch controlled gene expression. Chem. Commun. 2011, 47, 10734–10736. [Google Scholar] [CrossRef] [PubMed]
- Kondo, S.; Miura, T. Reaction-Diffusion Model as a Framework for Understanding Biological Pattern Formation. Science 2010, 329, 1616–1620. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, S.; Schwille, P. Pattern formation on membranes and its role in bacterial cell division. Curr. Opin. Cell Biol. 2016, 38, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, H.; de Boer, P.A.J. Pattern formation in Escherichia coli: A model for the pole-to-pole oscillations of Min proteins and the localization of the division site. Proc. Natl. Acad. Sci. USA 2001, 98, 14202–14207. [Google Scholar] [CrossRef] [PubMed]
- Loose, M.; Fischer-Friedrich, E.; Ries, J.; Kruse, K.; Schwille, P. Spatial regulators for bacterial cell division self-organize into surface waves in vitro. Science 2008, 320, 789–792. [Google Scholar] [CrossRef]
- Cross, M.C.; Hohenberg, P.C. Pattern-Formation Outside of Equilibrium. Rev. Mod. Phys. 1993, 65, 851–1112. [Google Scholar] [CrossRef]
- Wu, L.J.; Errington, J. Nucleoid occlusion and bacterial cell division. Nat. Rev. Microbiol. 2012, 10, 8–12. [Google Scholar] [CrossRef]
- Hu, Z.L.; Mukherjee, A.; Pichoff, S.; Lutkenhaus, J. The MinC component of the division site selection system in Escherichia coli interacts with FtsZ to prevent polymerization. Proc. Natl. Acad. Sci. USA 1999, 96, 14819–14824. [Google Scholar] [CrossRef]
- Osawa, M.; Anderson, D.E.; Erickson, H.P. Reconstitution of contractile FtsZ rings in liposomes. Science 2008, 320, 792–794. [Google Scholar] [CrossRef]
- Litschel, T.; Ramm, B.; Maas, R.; Heymann, M.; Schwille, P. Beating Vesicles: Encapsulated Protein Oscillations Cause Dynamic Membrane Deformations. Angew. Chem. Int. Edit. 2018, 57, 16286–16290. [Google Scholar] [CrossRef]
- Keener, J.P.; Sneyd, J. Mathematical Physiology; Springer: New York, NY, USA, 1998; Volume 1. [Google Scholar]
- Tayar, A.M.; Daube, S.S.; Bar-Ziv, R.H. Progress in programming spatiotemporal patterns and machine-assembly in cell-free protein expression systems. Curr. Opin. Chem. Biol. 2017, 40, 37–46. [Google Scholar] [CrossRef]
- Niederholtmeyer, H.; Stepanova, V.; Maerkl, S.J. Implementation of cell-free biological networks at steady state. Proc. Natl. Acad. Sci. USA 2013, 110, 15985–15990. [Google Scholar] [CrossRef]
- Karzbrun, E.; Tayar, A.M.; Noireaux, V.; Bar-Ziv, R.H. Programmable on-chip DNA compartments as artificial cells. Science 2014, 345, 829–832. [Google Scholar] [CrossRef]
- Tayar, A.M.; Karzbrun, E.; Noireaux, V.; Bar-Ziv, R.H. Propagating gene expression fronts in a one-dimensional coupled system of artificial cells. Nat. Phys. 2015, 11, 1037–1041. [Google Scholar] [CrossRef]
- Tayar, A.M.; Karzbrun, E.; Noireaux, V.; Bar-Ziv, R.H. Synchrony and pattern formation of coupled genetic oscillators on a chip of artificial cells. Proc. Natl. Acad. Sci. USA 2017, 114, 11609–11614. [Google Scholar] [CrossRef]
- Alon, U. An Introduction to Systems Biology: Design Principles of Biological Circuits, 1st ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2006. [Google Scholar]
- Huang, X.; Patil, A.J.; Li, M.; Mann, S. Design and Construction of Higher-Order Structure and Function in Proteinosome-Based Protocells. J. Am. Chem. Soc. 2014, 136, 9225–9234. [Google Scholar] [CrossRef]
- Lentini, R.; Santero, S.P.; Chizzolini, F.; Cecchi, D.; Fontana, J.; Marchioretto, M.; Del Bianco, C.; Terrell, J.L.; Spencer, A.C.; Martini, L.; et al. Integrating artificial with natural cells to translate chemical messages that direct E-coli behaviour. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef]
- Weitz, M.; Muckl, A.; Kapsner, K.; Berg, R.; Meyer, A.; Simmel, F.C. Communication and Computation by Bacteria Compartmentalized within Microemulsion Droplets. J. Am. Chem. Soc. 2014, 136, 72–75. [Google Scholar] [CrossRef]
- Shapiro, R. A simpler origin for life. Sci. Am. 2007, 296, 46–53. [Google Scholar] [CrossRef]
- Tawfik, D.S.; Griffiths, A.D. Man-made cell-like compartments for molecular evolution. Nat. Biotechnol. 1998, 16, 652–656. [Google Scholar] [CrossRef]
- Griffiths, A.D.; Tawfik, D.S. Directed evolution of an extremely fast phosphotriesterase by in vitro compartmentalization. EMBO J. 2003, 22, 24–35. [Google Scholar] [CrossRef]
- Schmidli, P.K.; Schurtenberger, P.; Luisi, P.L. Liposome-Mediated Enzymatic-Synthesis of Phosphatidylcholine as an Approach to Self-Replicating Liposomes. J. Am. Chem. Soc. 1991, 113, 8127–8130. [Google Scholar] [CrossRef]
- Luisi, P.L.; Walde, P.; Oberholzer, T. Lipid vesicles as possible intermediates in the origin of life. Curr. Opin. Colloid Interface Sci. 1999, 4, 33–39. [Google Scholar] [CrossRef]
- Chen, I.A.; Walde, P. From self-assembled vesicles to protocells. Cold Spring Harb. Perspect. Biol. 2010, 2, a002170. [Google Scholar] [CrossRef]
- Discher, D.E.; Eisenberg, A. Polymer vesicles. Science 2002, 297, 967–973. [Google Scholar] [CrossRef]
- Otrin, L.; Marusic, N.; Bednarz, C.; Vidakovic-Koch, T.; Lieberwirt, I.; Landfester, K.; Sundmacher, K. Toward Artificial Mitochondrion: Mimicking Oxidative Phosphorylation in Polymer and Hybrid Membranes. Nano Lett. 2017, 17, 6816–6821. [Google Scholar] [CrossRef]
- Liu, X.M.; Zhou, P.; Huang, Y.D.; Li, M.; Huang, X.; Mann, S. Hierarchical Proteinosomes for Programmed Release of Multiple Components. Angew. Chem. Int. Edit. 2016, 55, 7095–7100. [Google Scholar] [CrossRef]
- Oberholzer, T.; Wick, R.; Luisi, P.L.; Biebricher, C.K. Enzymatic RNA replication in self-reproducing vesicles: An approach to a minimal cell. Biochem. Biophys. Res. Commun. 1995, 207, 250–257. [Google Scholar] [CrossRef]
- Wick, R.; Luisi, P.L. Enzyme-containing liposomes can endogenously produce membrane-constituting lipids. Chem. Biol. 1996, 3, 277–285. [Google Scholar] [CrossRef]
- Ota, S.; Yoshizawa, S.; Takeuchi, S. Microfluidic formation of monodisperse, cell-sized, and unilamellar vesicles. Angew. Chem. Int. Ed. Engl. 2009, 48, 6533–6537. [Google Scholar] [CrossRef]
- Glavas-Dodov, M.; Fredro-Kumbaradzi, E.; Goracinova, K.; Simonoska, M.; Calis, S.; Trajkovic-Jolevska, S.; Hincal, A.A. The effects of lyophilization on the stability of liposomes containing 5-FU. Int. J. Pharm. 2005, 291, 79–86. [Google Scholar] [CrossRef]
- Sun, B.; Chiu, D.T. Determination of the encapsulation efficiency of individual vesicles using single-vesicle photolysis and confocal single-molecule detection. Anal. Chem. 2005, 77, 2770–2776. [Google Scholar] [CrossRef]
- Tan, Y.C.; Hettiarachchi, K.; Siu, M.; Pan, Y.R.; Lee, A.P. Controlled microfluidic encapsulation of cells, proteins, and microbeads in lipid vesicles. J. Am. Chem. Soc. 2006, 128, 5656–5658. [Google Scholar] [CrossRef]
- Zhu, P.A.; Wang, L.Q. Passive and active droplet generation with microfluidics: A review. Lab. Chip 2017, 17, 34–75. [Google Scholar] [CrossRef]
- Maamar, H.; Raj, A.; Dubnau, D. Noise in gene expression determines cell fate in Bacillus subtilis. Science 2007, 317, 526–529. [Google Scholar] [CrossRef]
- Chang, H.H.; Hemberg, M.; Barahona, M.; Ingber, D.E.; Huang, S. Transcriptome-wide noise controls lineage choice in mammalian progenitor cells. Nature 2008, 453, 544–547. [Google Scholar] [CrossRef]
- Graf, T.; Stadtfeld, M. Heterogeneity of Embryonic and Adult Stem Cells. Cell Stem Cell 2008, 3, 480–483. [Google Scholar] [CrossRef]
- Delgado, R.N.; Lim, D.A. Embryonic Nkx2.1-expressing neural precursor cells contribute to the regional heterogeneity of adult V-SVZ neural stem cells. Dev. Biol. 2015, 407, 265–274. [Google Scholar] [CrossRef]
- Luisi, P.L.; Allegretti, M.; de Souza, T.P.; Steiniger, F.; Fahr, A.; Stano, P. Spontaneous Protein Crowding in Liposomes: A New Vista for the Origin of Cellular Metabolism. ChemBioChem 2010, 11, 1989–1992. [Google Scholar] [CrossRef]
- De Souza, C.A.; Teixeira, P.C.; Faria, R.X.; Krylova, O.; Pohl, P.; Alves, L.A. A consensus segment in the M2 domain of the hP2X(7) receptor shows ion channel activity in planar lipid bilayers and in biological membranes. Biochim. Biophys. Acta 2012, 1818, 64–71. [Google Scholar] [CrossRef]
- Hansen, M.M.; Meijer, L.H.; Spruijt, E.; Maas, R.J.; Rosquelles, M.V.; Groen, J.; Heus, H.A.; Huck, W.T. Macromolecular crowding creates heterogeneous environments of gene expression in picolitre droplets. Nat. Nanotechnol. 2016, 11, 191–197. [Google Scholar] [CrossRef]
- Matsuura, T.; Tanimura, N.; Hosoda, K.; Yomo, T.; Shimizu, Y. Reaction dynamics analysis of a reconstituted Escherichia coli protein translation system by computational modeling. Proc. Natl. Acad. Sci. USA 2017, 114, E1336–E1344. [Google Scholar] [CrossRef]
- Chizzolini, F.; Forlin, M.; Martin, N.Y.; Berloffa, G.; Cecchi, D.; Mansy, S.S. Cell-Free Translation Is More Variable than Transcription. ACS Synth. Biol. 2017, 6, 638–647. [Google Scholar] [CrossRef]
- Elowitz, M.B.; Levine, A.J.; Siggia, E.D.; Swain, P.S. Stochastic gene expression in a single cell. Science 2002, 297, 1183–1186. [Google Scholar] [CrossRef]
- Karig, D.K.; Jung, S.Y.; Srijanto, B.; Collier, C.P.; Simpson, M.L. Probing Cell-Free Gene Expression Noise in Femtoliter Volumes. ACS Synth. Biol. 2013, 2, 497–505. [Google Scholar] [CrossRef]
- Nishimura, K.; Tsuru, S.; Suzuki, H.; Yomo, T. Stochasticity in Gene Expression in a Cell-Sized Compartment. ACS Synth. Biol. 2015, 4, 566–576. [Google Scholar] [CrossRef]
- Stogbauer, T.; Windhager, L.; Zimmer, R.; Radler, J.O. Experiment and mathematical modeling of gene expression dynamics in a cell-free system. Integr. Biol. 2012, 4, 494–501. [Google Scholar] [CrossRef]
- Kempf, N.; Remes, C.; Ledesch, R.; Zuchner, T.; Hofig, H.; Ritter, I.; Katranidis, A.; Fitter, J. A Novel Method to Evaluate Ribosomal Performance in Cell-Free Protein Synthesis Systems. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Niess, A.; Failmezger, J.; Kuschel, M.; Siemann-Herzberg, M.; Takors, R. Experimentally Validated Model Enables Debottlenecking of in Vitro Protein Synthesis and Identifies a Control Shift under in Vivo Conditions. ACS Synth. Biol. 2017, 6, 1913–1921. [Google Scholar] [CrossRef]
- Schwander, T.; von Borzyskowski, L.S.; Burgener, S.; Cortina, N.S.; Erb, T.J. A synthetic pathway for the fixation of carbon dioxide in vitro. Science 2016, 354, 900–904. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Brea, R.J.; Niederholtmeyer, H.; Devaraj, N.K. A minimal biochemical route towards de novo formation of synthetic phospholipid membranes. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Van Nies, P.; Westerlaken, I.; Blanken, D.; Salas, M.; Mencia, M.; Danelon, C. Self-replication of DNA by its encoded proteins in liposome-based synthetic cells. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Regulate Elements | Regulation Description | Control Level | References |
|---|---|---|---|
| E. coli sigma factors | Transcriptional activation units | Transcription | [37] |
| SsrA-ClpXP | Positive degradation of reporter protein | [31,40,41] | |
| TetR, LacI, AraC, and lambda repressors Cl and Cro | Inducible transcriptional repression and activation; bistable switches; genetic oscillators | [23,33,37] | |
| Pr, Pr1, and Pr2 | Provides constant transcription | [39] | |
| pT181 and its derivatives | RNA transcriptional attenuator; antisense RNA mediated transcriptional repressor; three level RNA transcription cascades | [42] | |
| small transcription activating RNAs | Small RNAs that activate the transcription of a specific gene regulated by a terminator (T181, AD1) | [43] | |
| BetI, PhIF and SrpR, TetR, LacI, and QacR | 3,4 and 5-node oscillators | [34] | |
| RNA thermometers as well as other rational designed sequences | control the secondary structure of mRNA leading to control of the ribosomal binding rate | Translation | [44] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, K.; Zhu, Y.; Kai, L. Cell-Free Protein Synthesis: Chassis toward the Minimal Cell. Cells 2019, 8, 315. https://doi.org/10.3390/cells8040315
Yue K, Zhu Y, Kai L. Cell-Free Protein Synthesis: Chassis toward the Minimal Cell. Cells. 2019; 8(4):315. https://doi.org/10.3390/cells8040315
Chicago/Turabian StyleYue, Ke, Yiyong Zhu, and Lei Kai. 2019. "Cell-Free Protein Synthesis: Chassis toward the Minimal Cell" Cells 8, no. 4: 315. https://doi.org/10.3390/cells8040315
APA StyleYue, K., Zhu, Y., & Kai, L. (2019). Cell-Free Protein Synthesis: Chassis toward the Minimal Cell. Cells, 8(4), 315. https://doi.org/10.3390/cells8040315