The Adenosine A2B Receptor Drives Osteoclast-Mediated Bone Resorption in Hypoxic Microenvironments

Botnar Research Centre, Nuffield Department of Orthopaedics Rheumatology and Musculoskeletal Sciences, University of Oxford, Headington, Oxford OX3 7LD, UK
Cells 2019, 8(6), 624; https://doi.org/10.3390/cells8060624
Submission received: 8 May 2019
/
Revised: 12 June 2019
/
Accepted: 20 June 2019
/
Published: 21 June 2019
(This article belongs to the Special Issue Pathways Contributing to Cartilage and Bone Destruction in Arthritis)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Osteoclast-mediated bone destruction is amplified in the hypoxic synovial microenvironment of rheumatoid arthritis (RA). This increased bone resorption is driven by the hypoxia-inducible transcription factor HIF. We identified hypoxic induction of the HIF-regulated adenosine A2B receptor in primary human osteoclasts (mRNA, 3.8-fold increase, p < 0.01) and sought to identify the role(s) of purinergic signaling via this receptor in the bone resorption process. Primary human osteoclasts were differentiated from CD14+ monocytes and exposed to hypoxia (2% O2) and A2B receptor inhibitors (MRS1754, PSB603). The hypoxic increase in bone resorption was prevented by the inhibition of the A2B receptor, at least partly by the attenuation of glycolytic and mitochondrial metabolism via inhibition of HIF. A2B receptor inhibition also reduced osteoclastogenesis in hypoxia by inhibiting early cell fusion (day 3–4, p < 0.05). The A2B receptor is only functional in hypoxic or inflammatory environments when the extracellular concentrations of adenosine (1.6-fold increase, p < 0.05) are sufficient to activate the receptor. Inhibition of the A2B receptor under normoxic conditions therefore did not affect any parameter tested. Reciprocal positive regulation of HIF and the A2B receptor in a hypoxic microenvironment thus enhances glycolytic and mitochondrial metabolism in osteoclasts to drive increased bone resorption. A2B receptor inhibition could potentially prevent the pathological osteolysis associated with hypoxic diseases such as rheumatoid arthritis.
1. Introduction
Rheumatoid arthritis (RA) is a chronic inflammatory disease characterised by the progressive destruction of bone and articular cartilage. Bone resorption in RA is mediated by multi-nucleated osteoclasts and contributes significantly to disability and poor prognosis [1,2,3]. RA osteoclasts form by the fusion of circulating CD14+ mononuclear precursors or synovial macrophages in the presence of macrophage colony-stimulating factor (M-CSF) and receptor activator for nuclear factor κB ligand (RANKL) [4,5]. Their bone resorption capacity is increased in the hypoxic microenvironment of the rheumatoid joint in a manner dependent on the hypoxia-inducible factor (HIF) transcription factor [6,7].
Increasing attention has been paid to the role of purinergic signaling, a form of extracellular signaling mediated by purine nucleotides and nucleosides, in osteoclast differentiation and function. This has mainly focused on the effect of extracellular ATP to increase the formation and activity of osteoclasts, acting predominantly via the P2Y1, P2Y12 and P2X7 receptors [8]. There is more uncertainty regarding the role of extracellular adenosine, which has variously been reported to stimulate [9,10,11,12], inhibit [13,14] or have no effect on [15,16] osteoclastogenesis and bone resorption. Adenosine stimulates the P1 receptor family (A1, A2A, A2B, A3), and the majority of studies have focused on the A1 and A2A receptors.
Extracellular adenosine is formed by hydrolysis of extracellular ATP and ADP by ectonucleotidases located in the plasma membrane. ATP release into the extracellular space is increased under conditions of hypoxia or inflammation, via increased vesicular exocytosis from osteoblasts [17] for example, resulting in greatly increased extracellular concentrations of adenosine in hypoxia [18]. Our previous study identified the adenosine A2B receptor (A2BR, ADORA2b) as hypoxia-inducible in primary human osteoclasts [19]. The A2B receptor is hypoxia-inducible in many tissues but is also only activated in hypoxia; because of its low affinity for adenosine (EC50 = 24 μM), it is only under hypoxic or inflammatory conditions that sufficient extracellular adenosine is available to activate the A2B receptor [20].
RA is both inflammatory and hypoxic, and therefore adenosine levels would be expected to be elevated in RA patients. Adenosine is very unstable, however, and is hydrolysed by the adenosine deaminase present in synovial fluid, making its actual levels hard to measure clinically, although elevation of adenosine has been observed in murine models of RA [21,22]. High levels of adenosine abolish methotrexate (MTX)-induced suppression of osteoclastogenesis and inflammatory bone destruction in adjuvant-induced arthritis, via a mechanism mediated specifically by the A2B receptor [21]. Clinically, A2B receptor expression is reported within both rheumatoid synovium [23] and rheumatoid nodules [24]. This suggests that elevated concentrations of adenosine in RA may drive resistance to treatments such as MTX via stimulation of osteoclast formation and function.
This work aims to determine whether the A2B receptor mediates the hypoxia-induced increase of osteoclast-mediated bone destruction. It describes how A2B receptor inhibitors specifically reduce hypoxic bone resorption in osteoclasts, which is driven by a hypoxia-induced increase in A2B receptor expression as well as its activation by accumulation of extracellular adenosine. In turn, A2B receptor activation further stabilises HIF-1α to promote bone resorption via an increase in cellular metabolic rate.
2. Materials and Methods
2.1. Materials and Ethics
Reagents were obtained as follows: human M-CSF (R&D Systems, Abingdon, UK), human soluble RANKL (Peprotech, London UK), MRS1754 and PSB603 (Tocris / BioTechne, Abingdon, UK). Elephant dentine was obtained from HM Revenue & Customs, Heathrow Airport, UK. Unless stated, other reagents were from Sigma-Aldrich (Gillingham, UK). The study was conducted in accordance with the Declaration of Helsinki; the use of leucocyte cones for osteoclast differentiation was approved by the London–Fulham Research Ethics Committee (11/H0711/7).
2.2. Osteoclast Differentiation and Cell Culture
CD14+ monocytes were positively selected from the peripheral blood mononuclear cell component of leucocyte cones (NHS Blood and Transplant, UK) using magnetic CD14+ microbeads (Miltenyi Biotech, Surrey, UK). Monocytes were seeded at 1–1.25 × 106 cells/ml onto dentine discs or plastic dishes in α-MEM (without ribonucleosides/deoxyribonucleosides) containing 10% heat-inactivated foetal bovine serum (FBS), 2 mM L-glutamine, 50 IU/ml penicillin and 50 μg/ml streptomycin sulphate. Osteoclastogenesis was induced by treatment with 25 ng/ml M-CSF and 30 ng/ml RANKL every 3–4 days for 9 days. Monocytes were maintained in 25 ng/ml M-CSF. Primary human osteoblasts were purchased from Sigma-Aldrich. Hypoxic exposure was conducted at 2% O2, 5% CO2, balance N2 in a MiniGalaxy incubator (RS Biotech, Irvine, UK).
2.3. Osteoclast Formation and Activity Assays
Tartrate-resistant acid phosphatase (TRAP) staining of formalin-fixed cells used naphthol AS-BI phosphate as a substrate, with reaction of the product with Fast Violet B salt. Multi-nucleated cells containing three or more nuclei were considered osteoclasts. Vitronectin receptor (VNR) was detected by immunocytochemistry for CD51/61 (clone 23C6, 1:400; Bio-Rad, Oxford, UK). Resorption tracks produced by mature osteoclasts on dentine discs were visualised by staining with 0.5% toluidine blue following the removal of cells by sonication. Dentines were photographed, resorption tracks highlighted, and the resorbed area quantified using ImageJ (LOCI, Madison, Wisconsin, USA).
2.4. Transfection with siRNA and Luciferase Assays
Osteoclasts were transfected with 50 nM siRNA targeting HIF1α, HIF2A or a HIF1α scrambled control using RNAiMAX (Invitrogen, Paisley, UK). Duplexes were removed after 4 h, and osteoclasts were cultured for a further 48 h prior to assay. Cells were transfected with a phosphoglycerate kinase (PGK) hypoxia response element (HRE)–firefly luciferase plasmid ([25]; gifted by Professor AL Harris, University of Oxford, UK) or a pGL4-Per2–luciferase plasmid (containing a 419 bp region of the Per2 promoter around the E-box; gifted by Professor QJ Meng, University of Manchester, UK) as well as a pHRG–TK Renilla luciferase control plasmid (Promega, Southampton, UK) using Lipofectamine 2000 (Invitrogen). Luminescence was assayed after 24 h using the Dual-Luciferase Reporter Assay System (Promega), with firefly luciferase normalized to the Renilla transfection control. To account for potential cyclicity, Per2-luciferase experiments were conducted at the same times of day as the resorption experiments to which they corresponded, cumulative luciferase indicating the overall Per2 promoter activity during this 24 h period.
2.5. Realtime PCR
RNA was extracted in TRI reagent (Direct-Zol RNA Miniprep kit; Zymo Research, Irvine, CA, USA) and reverse-transcribed, and quantitative PCR was performed using Fast SYBR Green Master Mix in a Viia7 Real-Time PCR system (Applied Biosystems, Warrington, UK). Human primers were either pre-validated Quantitect primers (Qiagen, Manchester, UK) against ACTB (Hs_ACTB_2_SG), A2B receptor-coding sequence (Hs_ADORA2B_1_SG), HIF1A (Hs_HIF1A_1_SG) and HIF2A (Hs_EPAS1_1_SG) or designed in-house against CD39 (S: 5’-TTCTCTCCCTCCTTCTGCAA-3’, AS 5’-ATGGCCACTGTGAAAAGGAC-3’), CD73 (S: 5’-CGCAACAATGGCACAATTAC-3’, AS 5’-CTCGACACTTGGTGCAAAGA) and PER2 (S: 5’-CGCCCTTTCATCCACATCCT-3’, AS: 5’-AATCCGCTACCACCCCTTCC-3’). Relative quantification (comparative CT) of target gene mRNA was normalised to β-actin (ACTB) mRNA.
2.6. Western Blotting
Cells were homogenized in HIF lysis buffer (6.2 M urea, 10% glycerol, 5 mM dithiothreitol, 1% sodium dodecyl sulphate, protease inhibitors) or phospho-lysis buffer (1 mM EDTA, 1 mM phenylmethylsulphonyl fluoride (PMSF), 1 mM Na3VO4, 1 mM NaF in PBS). Gels were evenly loaded with 40–60 μg protein/lane, and molecular weights were compared against the full-range Rainbow molecular weight marker (Fisher Scientific, Loughborough, UK). Primary antibodies were against the A2B receptor (ab1589P, Abcam, Cambridge, UK), HIF-1α (clone 54, 1:1000; BD Biosciences, Oxford, UK), HIF-2α (ep190b, Novus Biologicals, Abingdon, UK), GLUT1 (ab14683, 1:2500; Abcam, Cambridge, UK), NFκB p65 and phospho NFκB p65 (C22B4 and 93H1, Cell Signalling Technology, Leiden, Netherlands) and β-tubulin (clone TUB2.1, 1:2500). Densitometric quantification of Western blots was performed in ImageJ (LOCI, Madison, Wisconsin, USA), normalizing experimental bands to the corresponding β-tubulin control.
2.7. Metabolic Assays
Glucose, adenosine and ATP were measured in the media using the Glucose (GO) Assay Kit, the Adenosine Assay Kit (Abcam, Cambridge, UK) and the ATP Determination Kit (ThermoFisher, Loughborough, UK). Lactate was assayed in heat-inactivated medium by the increase in absorbance (340 nm) as NAD+ was converted to NADH in the presence of 0.32 M glycine, 0.32 M hydrazine, 9.6 mM NAD+ and 3 U/ml lactate dehydrogenase. Mitochondrial dehydrogenase activity within the electron transport chain was assessed by adding Alamar blue (BioRad, Kidlington, UK) to cells in culture for 4 h. The results were normalized to the number of osteoclasts.
2.8. ELISAs
The concentrations of intracellular cyclic AMP (cAMP) and secreted interleukin-6 (IL-6) were measured using the cAMP Parameter and IL-6 Quantikine ELISA kits (BioTechne, Abingdon, UK), according to the manufacturer’s instructions.
2.9. Immunostaining
Osteoclasts were fixed in 4% formalin. The A2B receptor was visualized using the rabbit polyclonal H-200 antibody (Santa Cruz Biotechnology, Heidelberg, Germany) and a PBS control and either the VECTASTAIN Elite Universal ABC Kit and DAB (Vector Laboratories, Peterborough, UK) or an AlexaFluor594 fluorescent secondary antibody. For F-actin ring staining, the cells were permeabilised with 0.5% Triton X-100 and stained with FITC-conjugated phalloidin (0.5 μg/mL).
2.10. Statistical Methods
The results were derived from three or more independent experiments. Data are presented as mean ± standard error of mean (SEM) of data transformed to the mean of the control group. Data were analysed using Prism (Graphpad Software, San Diego, CA, USA). Statistical analysis comprised one-way or two-way ANOVA using Dunnett’s or Tukey’s multiple comparison as a post-hoc test. For experiments with only two conditions, a Students’ t-test was applied. The results were considered significant at p < 0.05.
3. Results
3.1. Hypoxic Induction of the A2B Receptor is HIF-Regulated in Osteoclasts
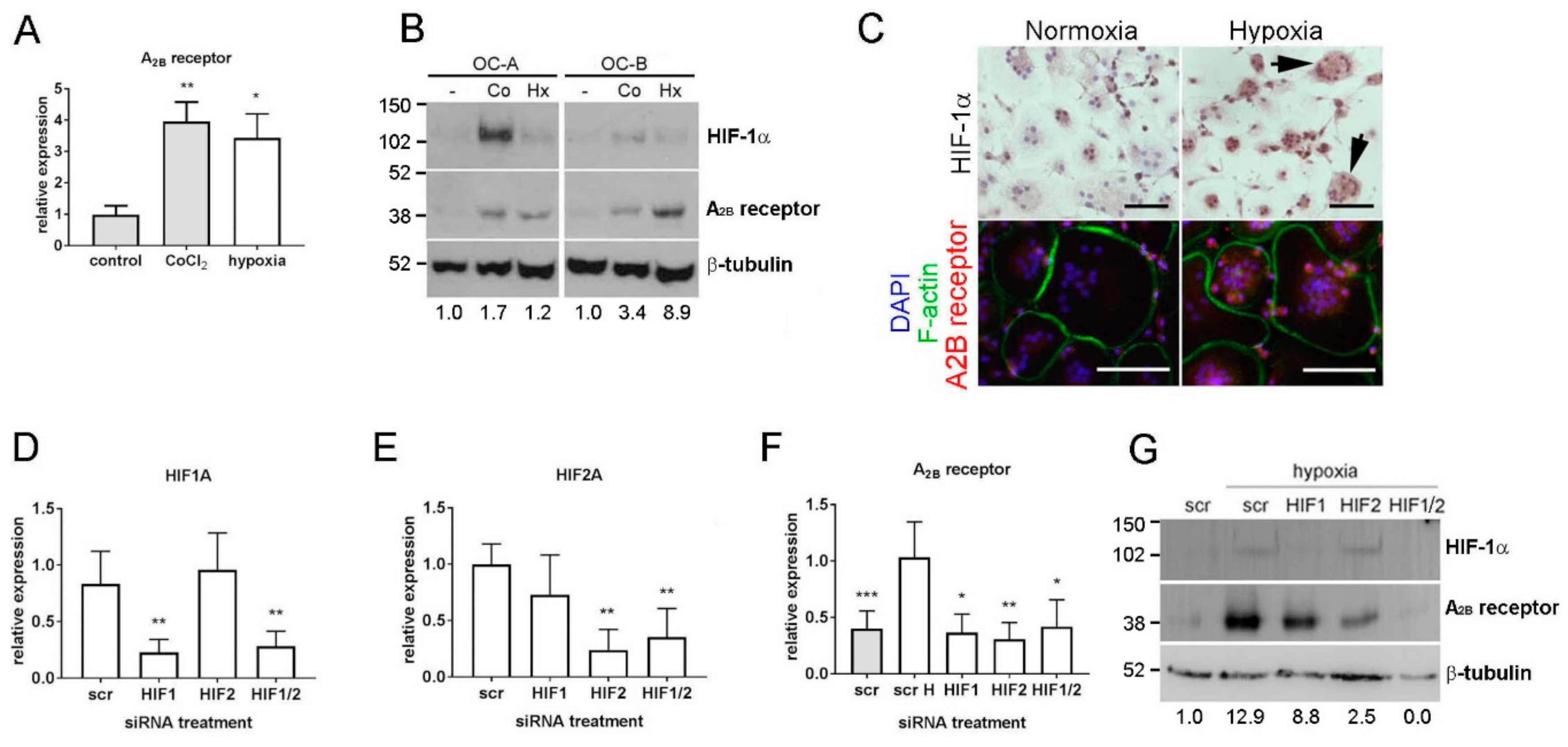
Our previous microarray analysis of hypoxia-induced genes in human monocyte-derived osteoclasts identified a 2.7-fold hypoxic induction of A2B receptor mRNA (p < 0.002) [19] that we confirmed in this study by real-time PCR (Figure 1A; 3.8-fold induction, p < 0.01). A2B receptor protein was also induced by hypoxia, with hypoxic conditions verified by stabilisation of HIF-1α (Figure 1B,C). CoCl2 induced A2B receptor mRNA and protein, suggesting a role for HIF in its transcriptional regulation. The variation in the magnitude of response to both stimuli is representative of the inter-individual variation characteristic of primary osteoclast preparations. HIF siRNA in osteoclasts achieved 79.7% and 77.1% knock-down of HIF1A and HIF2A mRNAs, respectively (Figure 1D,E) and almost complete ablation of hypoxia-induced HIF-1α protein (Figure 1G; HIF-2α protein is generally undetectable in osteoclasts by Western blot). In hypoxic osteoclasts, both HIF1A and HIF2A siRNA reduced the hypoxic induction of A2B receptor mRNA (Figure 1F) and protein (Figure 1G), indicating that the A2B receptor is regulated by both HIF-1α and HIF-2α in osteoclasts.
3.2. The A2B Receptor Drives the Hypoxic Increase in Osteoclast Bone Resorption
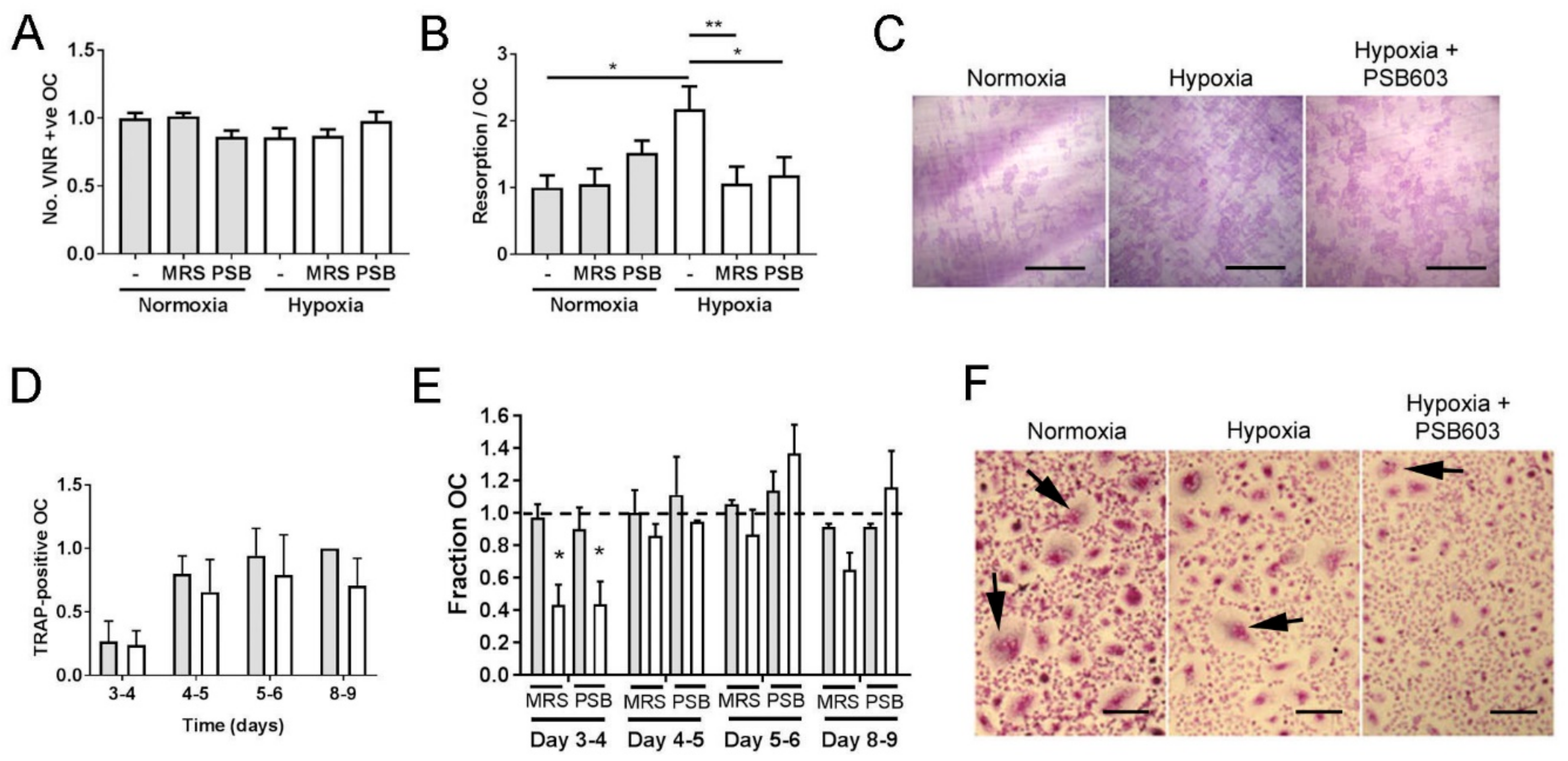
Hypoxia and HIF moderately inhibit osteoclastogenesis and greatly enhance bone resorption by osteoclasts [6,7]. We therefore investigated whether the A2B receptor might be a HIF-regulated mediator of these processes. The A2B receptor inhibitors MRS1754 and PSB603 were first tested for their effects on the survival and bone resorption capacity of mature osteoclasts. No concentration tested affected osteoclast number (Figure 2A), but both inhibitors were able to prevent the hypoxic increase in osteoclast-mediated bone resorption without affecting the normoxic levels of resorption (Figure 2B,C). Data shown correspond to the lowest concentration of each inhibitor able to prevent the hypoxic increase in bone resorption (2.5 μM MRS1754, 10 nM PSB603); these concentrations were selected for further experiments.
Continuous long-term hypoxia prevents osteoclastogenesis and causes apoptosis of mature osteoclasts [26,27]. We therefore exposed differentiating osteoclasts at different stages of differentiation to a limited 24 h period of hypoxia (Figure 2D), which overall had a moderate suppressive effect on the number of multi-nucleated osteoclasts formed at each stage of differentiation (p < 0.05). Exposure to MRS1754 or PSB603 during this 24 h period had no effect under normoxic culture conditions but delayed fusion under hypoxia at the 3–4-day timepoint (Figure 2E,F).
3.3. Hypoxic Osteoclasts Secrete ATP to Drive Increased Adenosine Concentrations
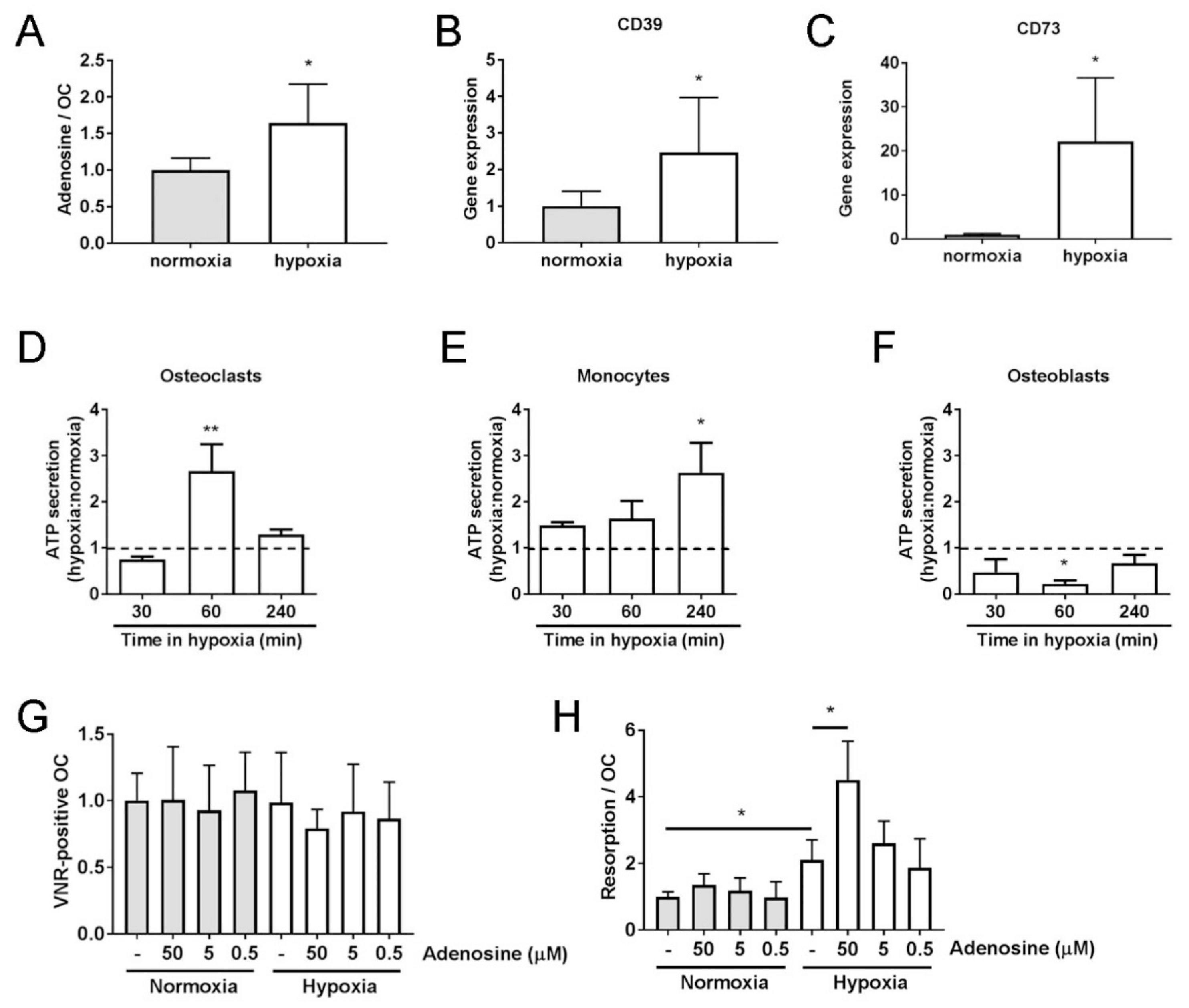
Despite A2B receptor protein being present in both normoxic and hypoxic osteoclasts, inhibition of its activity only affected bone resorption and osteoclastogenesis under hypoxia. This suggests that the concentration of extracellular adenosine, the substrate for the A2B receptor, is sufficiently elevated in in vitro cultured hypoxic osteoclasts to enable activation of the A2B receptor in the absence of any other cell type or stimulus. As hypothesised, extracellular adenosine was elevated 1.6-fold after 24 h in hypoxia (Figure 3A, p < 0.05). Extracellular adenosine is formed by the hydrolysis of extracellular ATP by the ectonucleotidases CD39 and CD73, restricting its actions to cells close to the release site. Expression of CD39 and CD73 was also increased in hypoxic osteoclasts (Figure 3B,C). Osteoblasts and osteoclasts constitutively secrete ATP [11,17,28], but only in osteoblasts is this function known to be affected by hypoxia, where acute (<3 min) hypoxic exposure increases ATP release [17]. Acute hypoxic exposure (≤30 min) did not affect osteoclast secretion of ATP; however, longer exposure did elicit this response in osteoclasts (60 min, p > 0.01) and their CD14+ mononuclear precursors (Figure 3D–F).
We therefore investigated the effects of adding A2B receptor-activating concentrations of adenosine to osteoclast cultures. Exogenous addition of adenosine only increased osteoclast-mediated bone resorption under hypoxic culture conditions (Figure 3G,H). This suggests that if the effects of exogenously added adenosine are also mediated by the A2B receptor, intracellular signaling after receptor activation must require an additional hypoxia-activated process or pathway.
3.4. The A2B Receptor Activates Glycolytic Pathways
Several intracellular signaling pathways downstream of the A2B receptor have potential relevance to osteoclast-mediated bone resorption. These include the interaction of the A2B receptor with the actin cytoskeleton post-stimulation, via its recruitment to the plasma membrane, which initiates the interaction with actin-associated ezrin and actinin-1 [29,30]. Rearrangement of the actin cytoskeleton is vital for osteoclast bone resorption, and although we observed increased F-actin ring formation under hypoxia, inhibition of the A2B receptor did not affect this (Supplementary Figure S1A,B). Activation of the A2B receptor in myeloid cells by exposure to adenosine causes the release of interleukin-6 (IL-6) [31]. IL-6 stimulates both osteoclast formation and activity; however, its production by osteoclasts was not affected by either hypoxia or A2B receptor inhibition (Supplementary Figure S1C).
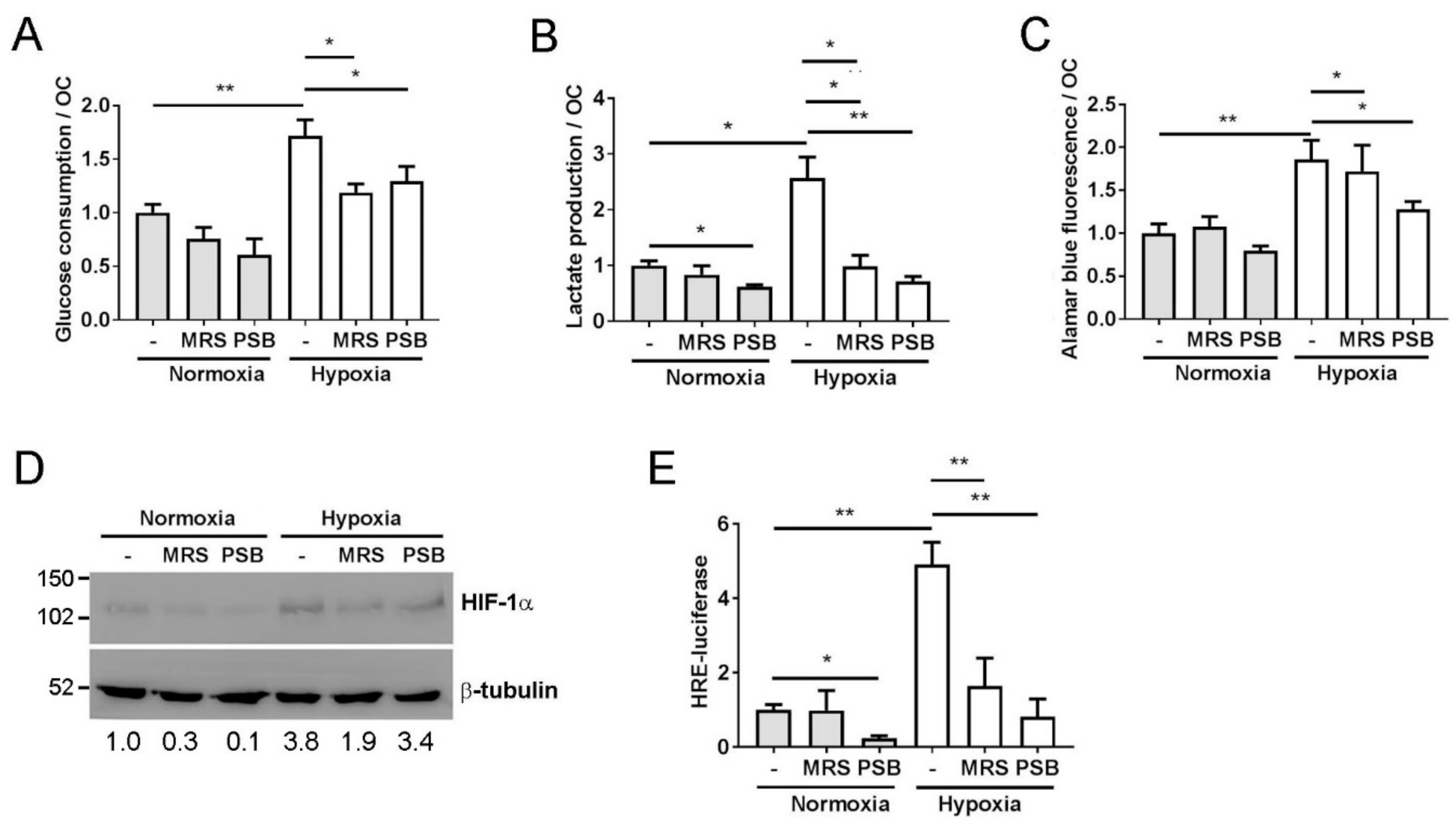
Activation of the A2B receptor also increases the glycolytic rate via an intracellular feedback loop whereby hypoxic activation of the A2B receptor further activates HIF-1α signaling, which in turn stimulates glycolysis as part of the cellular metabolic adaptation to hypoxia [32,33]. Glycolysis plays a central role in the hypoxic increase in osteoclast-mediated bone resorption [34,35]. Both MRS1754 and PSB603 prevented the hypoxic increase in glucose consumption (Figure 4A) and lactate production (Figure 4B), as well as the osteoclast-specific hypoxic increase in mitochondrial reductase activity (Figure 4C) [34]. Inhibition of the A2B receptor reduced the stabilisation of HIF-1α (Figure 4D) as well as reducing HRE-driven transcription under hypoxia (Figure 4E). One proposed mechanism by which the A2B receptor activates HIF signaling is via the circadian rhythm protein period 2 (Per2). A2B receptor signaling commonly involves adenylyl cyclase and leads to an increase in intracellular cAMP [20], which, in turn, stabilises Per2 and, sequentially, HIF-1α, with consequent effects on glycolysis [33]. However, cAMP and PER2 mRNA levels were reduced in hypoxic osteoclasts, while PER2-driven transcription was increased, although this could potentially be due to the presence of HIF-driven hypoxia-response elements in the Per2 promoter E-box [36]. No effect of A2B receptor inhibition under hypoxia was evident in either case (Supplementary Figure S2).
4. Discussion
Within the literature, there is considerable uncertainty regarding the effect of extracellular adenosine on osteoclast formation and activity. Actions of adenosine via the A1 receptor generally stimulate osteoclasts [9,10,12], whereas reports of effects via the A2A receptor are divergent [11,13,14]. Exogenous addition of adenosine could potentially affect all four adenosine receptors and result in a combined lack of effect on osteoclast biology [15,16]. Indeed, we also saw no effect of the exogenous addition of adenosine on osteoclast-mediated bone resorption under the normoxic culture conditions in which the above referenced publications were performed.
Exogenous adenosine did increase osteoclast-mediated bone resorption in hypoxic culture, however, suggestive of the requirement for a hypoxia-inducible mediator(s) of this process. The A2B receptor is hypoxia-responsive and is regulated by HIF-1α in human endothelial cells, intestinal and alveolar epithelial cells, dendritic cells and breast cancer cells [37,38,39,40]. There are no previous reports of HIF-2α regulating the A2B receptor, suggesting that its induction by both HIF-1α and HIF-2α in osteoclasts might be cell-type specific. It is unclear why HIF isoform-specific siRNAs comparably reduced A2B receptor mRNA expression, while HIF-2α siRNA more strongly inhibited A2B receptor protein levels. It is possible that the intracellular A2B receptor–HIF feedback loop [32,33] is stronger for HIF-2α than HIF-1α in osteoclasts and could account for this discrepancy; a hypothesis that would be of interest to follow up in the future.
A2B receptor expression in osteoclasts increases during their differentiation from mononuclear precursors [41] and subsequently during hypoxic exposure. The observed intracellular distribution of the A2B receptor in both normoxic and hypoxic osteoclasts was predominantly cytoplasmic as described (www.proteinatlas.org), with a peri-nuclear bias and some nuclear involvement similar to that observed in human sclera fibroblasts and retinal pigment epithelial cells [42,43]. Due to its low affinity for adenosine, the A2B receptor is only activated at the elevated concentrations of extracellular adenosine achieved in hypoxic conditions, despite the protein being present in normoxia. Osteoclast cultures exhibited increased concentrations of ATP and adenosine in the media following acute and extended periods of hypoxic exposure, respectively. Extracellular adenosine is formed by hydrolysis of extracellular ATP and ADP by the ectonucleotidases CD39 and CD73 located in the plasma membrane, whose mRNA levels also increased following hypoxic exposure. The controlled release of ATP from osteoclasts via the P2X7 receptor results in the accumulation of extracellular adenosine [11,28], although this process has not been described as hypoxia-regulated. Elevated ATP release by hypoxic osteoblasts via vesicular exocytosis [17] could also contribute to A2B receptor activation in adjacent hypoxic osteoclasts in vivo.
For the reasons above, A2B receptor inhibition with either MRS1754 or PSB603 only suppressed osteoclast fusion and bone resorption in hypoxic culture conditions, when elevated extracellular concentrations of adenosine were sufficient to activate the A2B receptor in the hypoxic cells. Fusogenic actions of extracellular adenosine have been described in both murine and human osteoclast cultures and attributed to the A1 or A2A receptors [9,11,12,44]. We did not look at the effects of adenosine on osteoclastogenesis in normoxic culture but found that early fusion in hypoxia was suppressed by the inhibition of the A2B receptor, adding an additional fusogenic role for this receptor to the list. There are potential A2B receptor-independent effects of both inhibitors. MRS1754 is less selective than PSB603 (200-fold versus >17000-fold selectivity) and therefore might partially inhibit other adenosine receptors, although the fact that the effects were only observed in hypoxia would indicate predominant inhibition of the A2B receptor. Despite high selectivity for the A2B receptor, PSB603 increased the rate of oxygen consumption of colorectal cancer cells in an A2B receptor-independent manner [45]. However, this effect was not seen at the low 10 nM concentration used in our study, which also induced hypoxia-dependent and inhibitory effects, again suggestive of a predominantly A2B receptor-driven pathway.
Prevention of the hypoxic increase in osteoclast-mediated bone resorption by A2B receptor inhibitors suggests that adenosine also stimulates processes involved in bone resorption via the A2B receptor. No effect of A2B receptor inhibitors on bone resorption was seen under normoxic conditions. Previous reports regarding the effects of the A2B receptor on osteoclast activity are opposing. Bone marrow-derived osteoclast formation and bone resorption ex vivo was enhanced in cells from A2B receptor knockout mice [46]. However, these results were obtained under normoxic culture conditions in which the A2B receptor should be functionally inactive, suggestive of potential off-target effects in the knock-out animals. The A2B receptor partial agonist BAY60-6583 inhibited precursor fusion and the expression of osteoclast marker genes during differentiation of murine and human bone marrow-derived mononuclear cells into osteoclasts, as well as reduced F-actin ring formation and bone resorption [41,47]. This is harder to reconcile with our data. It is possible that artificial stimulation of the A2B receptor activates alternative intracellular signaling pathways to those activated by physiological stimulation of the receptor, resulting in cellular effects that are not related to the normal cellular functions of the A2B receptor. It has, for example, been noted that in the presence of high levels of adenosine, BAY60-6583 can instead act as an antagonist and block the effects of adenosine at A2B receptors [48].
The adenosine A2B receptor–HIF signaling pathway is one intracellular pathway that would likely not be activated by A2B receptor agonists administered in normoxic culture, due to the requirement for hypoxia to cause inactivation of the HIF-regulating prolyl hydroxylase (PHD) enzymes and subsequent stabilization of HIF-α proteins [7,49]. Adenosine and the A2B receptor amplify HIF signaling under hypoxic conditions, contributing to further stabilisation of HIF protein and activation of HIF-mediated transcription in neuroendocrine enterochromaffin cells of the gut [50], smooth muscle cells [32], oral squamous cell carcinoma cells [51], cardiomyocytes [33] and microglial cells [52], as well as in adenosine deaminase-deficient mice and in vivo models of sickle cell disease [32]. Activation of this signaling pathway leads to an A2B receptor- and HIF-dependent increase in glycolytic metabolism [33,52]. Increased glucose uptake and glycolysis in hypoxic conditions is vital to maintain mitochondrial metabolism and enable osteoclast-mediated bone resorption in these highly energy-dependent cells [34,35]. The fundamental requirement for increased glycolytic and mitochondrial metabolism to enable increased bone resorption in hypoxic osteoclasts explains why adenosine only increases bone resorption activity under hypoxia, when other essential intracellular components of this pathway are also active.
There is evidence that similar pathways are active in osteoclasts in vivo. Plasma adenosine levels are markedly elevated in rats with adjuvant-induced arthritis, a murine model of RA. The time of elevation of adenosine corresponds to the time-point at which the administration of MTX fails to suppress inflammatory bone destruction [21]. In vitro, adenosine abrogates MTX-induced suppression of osteoclastogenesis by suppressing the expression of osteoprotegerin in supporting stromal cells in an A2B receptor-dependent manner [21]. Clinically, the A2B receptor is expressed within both the rheumatoid synovium [23] and rheumatoid nodules [24], suggesting that elevated concentrations of adenosine in RA may drive resistance to treatments such as MTX via multiple mechanisms that stimulate osteoclast formation and function. It is possible that adenosine also drives bone destruction in other diseases. For example, multiple myeloma is a haematological malignancy of terminally differentiated plasma cells that arises within the bone marrow and gives rise to osteolytic bone disease. Elevated extracellular adenosine concentrations in multiple myeloma have been proposed to be indicative of aggressive disease [53], which might in part cause further activation of bone destruction by osteoclasts.
5. Conclusions
This study demonstrates that the reciprocal positive regulation of HIF and the A2B receptor in a hypoxic microenvironment enhances glycolytic and mitochondrial metabolism in osteoclasts to promote osteoclast differentiation and drive increased bone resorption. The adenosine A2B receptor–HIF signaling pathway is only active in hypoxic microenvironments, meaning that the inhibition of the A2B receptor only affects these cellular processes under hypoxia. As tissue hypoxia is a disease-specific environment, the A2B receptor could represent a potential therapeutic target, with specificity for hypoxic diseased tissue, which could prevent the pathological osteolysis associated with diseases such as RA.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4409/8/6/624/s1, Figure S1: ADORA2b does not modulate F-actin ring formation or IL-6 secretion in hypoxic osteoclasts. Figure S2: ADORA2b inhibitors do not affect cAMP levels or PER2 expression.
Funding
This research was funded by Arthritis Research UK, grant number MP/19200. Work in the Nuffield Department of Orthopaedics, Rheumatology and Musculoskeletal Sciences (NDORMS) is additionally supported by the Oxford National Institute of Health Research (NIHR) Musculoskeletal Biomedical Research Unit (BRU).
Acknowledgments
Thanks to AL Harris (University of Oxford, UK) for the PGK HRE–firefly luciferase plasmid and to Q.J. Meng (University of Manchester, UK) for the pGL4-Per2–luciferase plasmid. Thanks to Afsie Sabokbar and Emma Morris for critical reading of the manuscript.
Conflicts of Interest
The author declares no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Kaarela, K. Prognostic Factors and Diagnostic Criteria in Early Rheumatoid Arthritis. Scand. J. Rheumatol. Suppl. 1985, 57, 1–54. [Google Scholar] [CrossRef] [PubMed]
- Harrison, B.J.; Hutchinson, C.E.; Adams, J.; Bruce, I.N.; Herrick, A.L. Assessing Periarticular Bone Mineral Density in Patients with Early Psoriatic Arthritis or Rheumatoid Arthritis. Ann. Rheum. Dis. 2002, 61, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Gough, A.; Sambrook, P.; Devlin, J.; Huissoon, A.; Njeh, C.; Robbins, S.; Nguyen, T.; Emery, P. Osteoclastic Activation Is the Principal Mechanism Leading to Secondary Osteoporosis in Rheumatoid Arthritis. J. Rheumatol. 1998, 25, 1282–1289. [Google Scholar] [PubMed]
- Fujikawa, Y.; Sabokbar, A.; Neale, S.; Athanasou, N.A. Human Osteoclast Formation and Bone Resorption by Monocytes and Synovial Macrophages in Rheumatoid Arthritis. Ann. Rheum. Dis. 1996, 55, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Mochizuki, S.; Tomoyasu, A.; Yano, K.; Goto, M.; et al. Osteoclast Differentiation Factor Is a Ligand for Osteoprotegerin/Osteoclastogenesis-Inhibitory Factor and Is Identical to Trance/Rankl. Proc. Natl. Acad. Sci. USA 1998, 95, 3597–3602. [Google Scholar] [CrossRef] [PubMed]
- Knowles, H.J. Hypoxic Regulation of Osteoclast Differentiation and Bone Resorption Activity. Hypoxia 2015, 3, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Hulley, P.A.; Bishop, T.; Vernet, A.; Schneider, J.E.; Edwards, J.R.; Athanasou, N.A.; Knowles, H.J. Hypoxia-Inducible Factor 1-Alpha Does Not Regulate Osteoclastogenesis but Enhances Bone Resorption Activity Via Prolyl-4-Hydroxylase 2. J. Pathol. 2017, 242, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Orriss, I.R. The Role of Purinergic Signalling in the Musculoskeletal System. Auton. Neurosci. 2015, 191, 124–134. [Google Scholar] [CrossRef]
- Kara, F.M.; Chitu, V.; Sloane, J.; Axelrod, M.; Fredholm, B.B.; Stanley, E.R.; Cronstei, B.N. Adenosine A1 Receptors (A1rs) Play a Critical Role in Osteoclast Formation and Function. FASEB J. 2010, 24, 2325–2333. [Google Scholar] [CrossRef]
- Kara, F.M.; Doty, S.B.; Boskey, A.; Goldring, S.; Zaidi, M.; Fredholm, B.B.; Cronstein, B.N. Adenosine a(1) Receptors Regulate Bone Resorption in Mice: Adenosine a(1) Receptor Blockade or Deletion Increases Bone Density and Prevents Ovariectomy-Induced Bone Loss in Adenosine a(1) Receptor-Knockout Mice. Arthritis Rheum. 2010, 62, 534–541. [Google Scholar] [CrossRef]
- Pellegatti, P.; Falzoni, S.; Donvito, G.; Lemaire, I.; Di Virgilio, F. P2x7 Receptor Drives Osteoclast Fusion by Increasing the Extracellular Adenosine Concentration. FASEB J. 2011, 25, 1264–1274. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Cronstein, B.N. Adenosine A1 Receptor Regulates Osteoclast Formation by Altering Traf6/Tak1 Signaling. Purinergic Signal. 2012, 8, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Mediero, A.; Frenkel, S.R.; Wilder, T.; He, W.; Mazumder, A.; Cronstein, B.N. Adenosine A2a Receptor Activation Prevents Wear Particle-Induced Osteolysis. Sci. Transl. Med. 2012. [Google Scholar] [CrossRef] [PubMed]
- Mediero, A.; Kara, F.M.; Wilder, T.B.; Cronstein, N. Adenosine a(2a) Receptor Ligation Inhibits Osteoclast Formation. Am. J. Pathol. 2012, 180, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Hajjawi, M.O.; Patel, J.J.; Corcelli, M.; Arnett, T.R.; Orriss, I.R. Lack of Effect of Adenosine on the Function of Rodent Osteoblasts and Osteoclasts in Vitro. Purinergic Signal. 2016, 12, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Morrison, L.; Turin, M.S.; King, B.F.; Burnstock, G.; Arnett, T.R. Atp Is a Potent Stimulator of the Activation and Formation of Rodent Osteoclasts. J. Physiol. 1998, 511, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Orriss, I.R.; Knight, G.E.; Utting, J.C.; Taylor, S.E.; Burnstock, G.; Arnett, T.R. Hypoxia Stimulates Vesicular Atp Release from Rat Osteoblasts. J. Cell Physiol. 2009, 220, 155–162. [Google Scholar] [CrossRef]
- Strazzulla, L.C.; Cronstein, B.N. Regulation of Bone and Cartilage by Adenosine Signaling. Purinergic Signal. 2016, 12, 583–593. [Google Scholar] [CrossRef]
- Knowles, H.J.; Cleton-Jansen, A.M.; Korsching, E.; Athanasou, N.A. Hypoxia-Inducible Factor Regulates Osteoclast-Mediated Bone Resorption: Role of Angiopoietin-Like 4. FASEB J. 2010, 24, 4648–4659. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, P. Huang. Adenosine A2b Receptor: From Cell Biology to Human Diseases. Front. Chem. 2016, 4, 37. [Google Scholar] [CrossRef]
- Teramachi, J.; Kukita, A.; Li, Y.J.; Ushijima, Y.; Ohkuma, H.; Wada, N.; Watanabe, T.; Nakamura, S.; Kukita, T. Adenosine Abolishes Mtx-Induced Suppression of Osteoclastogenesis and Inflammatory Bone Destruction in Adjuvant-Induced Arthritis. Lab. Invest. 2011, 91, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.S.; Du, Z.H.; Li, Z.H.; Xie, W.X.; Huang, K.T.; Chen, Y.; Chen, Z.Y.; Hu, H.; Wang, J.L.; Fang, J.Q. Repeated Electroacupuncture Persistently Elevates Adenosine and Ameliorates Collagen-Induced Arthritis in Rats. Evid. Based Complement. Alternat. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Stamp, L.K.; Hazlett, J.; Roberts, R.L.; Frampton, C.; Highton, J.; Hessian, P.A. Adenosine Receptor Expression in Rheumatoid Synovium: A Basis for Methotrexate Action. Arthritis Res. Ther. 2012, 14, R138. [Google Scholar] [CrossRef]
- Houlder, E.L.; Millier, M.J.; Highton, J.; Gwynne-Jones, D.; Stamp, L.K.; Hessian, P.A. Expression of the Genes Facilitating Methotrexate Action within Subcutaneous Rheumatoid Nodules. Clin. Exp. Rheumatol. 2017, 35, 943–947. [Google Scholar] [PubMed]
- Jones, D.T.; Harris, A.L. Identification of Novel Small-Molecule Inhibitors of Hypoxia-Inducible Factor-1 Transactivation and DNA Binding. Mol. Cancer Ther. 2006, 5, 2193–2202. [Google Scholar] [CrossRef] [PubMed]
- Knowles, H.J.; Athanasou, N.A. Acute Hypoxia and Osteoclast Activity: A Balance between Enhanced Resorption and Increased Apoptosis. J. Pathol. 2009, 218, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Yu, R.; Zhao, J.; Sun, L.; Jian, L.; Li, C.; Liu, X. Constant Hypoxia Inhibits Osteoclast Differentiation and Bone Resorption by Regulating Phosphorylation of Jnk and Ikappabalpha. Inflamm. Res. 2019, 68, 157–166. [Google Scholar] [CrossRef]
- Brandao-Burch, A.; Key, J.; Patel, M.L.; Arnett, T.R.; Orriss, I.R. The P2x7 Receptor Is an Important Regulator of Extracellular Atp Levels. Front. Endocrinol. 2012. [Google Scholar] [CrossRef]
- Sitaraman, S.V.; Wang, L.; Wong, M.; Bruewer, M.; Hobert, M.; Yun, C.H.; Merlin, D.; Madara, J.L. The Adenosine 2b Receptor Is Recruited to the Plasma Membrane and Associates with E3karp and Ezrin Upon Agonist Stimulation. J. Biol. Chem. 2002, 277, 33188–33195. [Google Scholar] [CrossRef]
- Sun, Y.; Hu, W.; Yu, X.; Liu, Z.; Tarran, R.; Ravid, K.; Huang, P. Actinin-1 Binds to the C-Terminus of A2b Adenosine Receptor (A2bar) and Enhances A2bar Cell-Surface Expression. Biochem. J. 2016, 473, 2179–2186. [Google Scholar] [CrossRef]
- Hu, X.; Adebiyi, M.G.; Luo, J.; Sun, K.; Le, T.T.; Zhang, Y.; Wu, H.; Zhao, S.; Karmouty-Quintana, H.; et al. Sustained Elevated Adenosine Via Adora2b Promotes Chronic Pain through Neuro-Immune Interaction. Cell Rep. 2016, 16, 106–119. [Google Scholar] [CrossRef]
- Ning, C.; Wen, Y.; Zhang, Y.; Dai, W.; Wang, W.; Zhang, L.; Qi, A.; Grenz, H.K.; Eltzschig, M.R.; et al. Excess Adenosine A2b Receptor Signaling Contributes to Priapism through Hif-1alpha Mediated Reduction of Pde5 Gene Expression. FASEB J. 2014, 28, 2725–2735. [Google Scholar] [CrossRef]
- Eckle, T.; Hartmann, K.; Bonney, S.; Reithel, S.; Mittelbronn, M.; Walker, L.A.; Lowes, B.D.; Han, J.; Borchers, C.H.; Buttrick, P.M.; et al. Adora2b-Elicited Per2 Stabilization Promotes a Hif-Dependent Metabolic Switch Crucial for Myocardial Adaptation to Ischemia. Nat. Med. 2012, 18, 774–782. [Google Scholar] [CrossRef] [PubMed]
- Morten, K.J.; Badder, L.; Knowles, H.J. Differential Regulation of Hif-Mediated Pathways Increases Mitochondrial Metabolism and Atp Production in Hypoxic Osteoclasts. J. Pathol. 2013, 229, 755–764. [Google Scholar] [CrossRef]
- Arnett, T.R.; Orriss, I.R. Metabolic Properties of the Osteoclast. Bone 2018, 115, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Okabe, T.M.; Kumagai, Y.; Nakajima, S.; Shirotake, K.; Kodaira, M.; Oyama, M.U.; Ikeda, M. The Impact of Hif1alpha on the Per2 Circadian Rhythm in Renal Cancer Cell Lines. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed]
- Poth, J.M.; Brodsky, K.; Ehrentraut, H.; Grenz, A.; Eltzschig, H.K. Transcriptional Control of Adenosine Signaling by Hypoxia-Inducible Transcription Factors During Ischemic or Inflammatory Disease. J. Mol. Med. 2013, 91, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Eckle, T.; Kewley, E.M.; Brodsky, K.; Tak, E.; Bonney, S.; Gobel, M.; Anderson, D.; Glover, L.E.; Riegel, A.K.; Colgan, S.P.; et al. Eltzschig. Identification of Hypoxia-Inducible Factor Hif-1a as Transcriptional Regulator of the A2b Adenosine Receptor During Acute Lung Injury. J. Immunol. 2014, 192, 1249–1256. [Google Scholar] [CrossRef]
- Lan, J.; Lu, H.; Samanta, D.; Salman, S.; Lu, Y.; Semenza, G.L. Hypoxia-Inducible Factor 1-Dependent Expression of Adenosine Receptor 2b Promotes Breast Cancer Stem Cell Enrichment. Proc. Natl. Acad. Sci. USA 2018. [Google Scholar] [CrossRef]
- Yang, M.; Ma, C.; Liu, S.; Shao, Q.; Gao, W.; Song, B.; Sun, J.; Xie, Q.; Zhang, Y.; Feng, A.; et al. Hif-Dependent Induction of Adenosine Receptor A2b Skews Human Dendritic Cells to a Th2-Stimulating Phenotype under Hypoxia. Immunol. Cell Biol. 2010, 88, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Oh, J.H.; Lee, N.K. The Inactivation of Erk1/2, P38 and Nf-Kb Is Involved in the Down-Regulation of Osteoclastogenesis and Function by A2b Adenosine Receptor Stimulation. Mol. Cells 2017, 40, 752–760. [Google Scholar]
- Cui, D.; Trier, K.; Chen, X.; Zeng, J.; Yang, X.; Hu, J.; Ge, J. Distribution of Adenosine Receptors in Human Sclera Fibroblasts. Mol. Vis. 2008, 14, 523–529. [Google Scholar] [PubMed]
- Wan, W.J.; Cui, D.M.; Yang, X.; Hu, J.M.; Li, C.X.; Hu, S.L.; Trier, K.; Zeng, J.W. Expression of Adenosine Receptors in Human Retinal Pigment Epithelium Cells in Vitro. Chin. Med. J. 2011, 124, 1139–1144. [Google Scholar] [PubMed]
- Merrill, J.T.; Shen, C.; Schreibman, D.; Coffey, D.; Zakharenko, O.; Fisher, R.; Lahita, R.G.; Salmon, J.; Cronstein, B.N. Adenosine A1 Receptor Promotion of Multinucleated Giant Cell Formation by Human Monocytes: A Mechanism for Methotrexate-Induced Nodulosis in Rheumatoid Arthritis. Arthritis Rheum. 1997, 40, 1308–1315. [Google Scholar] [PubMed]
- Molck, C.; Ryall, J.; Failla, L.M.; Coates, J.L.; Pascussi, J.M.; Heath, J.K.; Stewart, G.; Hollande, F. The A2b Adenosine Receptor Antagonist Psb-603 Promotes Oxidative Phosphorylation and Ros Production in Colorectal Cancer Cells Via Adenosine Receptor-Independent Mechanism. Cancer Lett. 2016, 383, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Corciulo, C.; Wilder, T.; Cronstein, B.N. Adenosine A2b Receptors Play an Important Role in Bone Homeostasis. Purinergic Signal. 2016, 12, 537–547. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Mazumder, A.; Wilder, T.; Cronstein, B.N. Adenosine Regulates Bone Metabolism Via A1, A2a, and A2b Receptors in Bone Marrow Cells from Normal Humans and Patients with Multiple Myeloma. FASEB J. 2013, 27, 3446–3454. [Google Scholar] [CrossRef] [PubMed]
- Hinz, S.; Lacher, S.K.; Seibt, B.F.; Muller, C.F. Bay60-6583 Acts as a Partial Agonist at Adenosine A2b Receptors. J. Pharmacol. Exp. Ther. 2014, 349, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Majmundar, A.J.; Wong, W.J.; Simon, M.C. Hypoxia-Inducible Factors and the Response to Hypoxic Stress. Mol. Cell 2010, 40, 294–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dammen, R.; Haugen, M.; Svejda, B.; Alaimo, D.; Brenna, O.; Pfragner, R.; Gustafsson, B.I.; Kidd, M. The Stimulatory Adenosine Receptor Adora2b Regulates Serotonin (5-Ht) Synthesis and Release in Oxygen-Depleted Ec Cells in Inflammatory Bowel Disease. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Kasama, H.; Sakamoto, Y.; Kasamatsu, A.; Okamoto, A.; Koyama, T.; Minakawa, Y.; Ogawara, K.; Yokoe, H.; Shiiba, M.; Tanzawa, H.; et al. Adenosine A2b Receptor Promotes Progression of Human Oral Cancer. BMC Cancer 2015. [Google Scholar] [CrossRef] [PubMed]
- Merighi, S.; Borea, P.A.; Stefanelli, A.; Bencivenni, S.; Castillo, C.A.; Varani, K.; Gessi, S. A2a and A2b Adenosine Receptors Affect Hif-1alpha Signaling in Activated Primary Microglial Cells. Glia 2015, 63, 1933–1952. [Google Scholar] [CrossRef] [PubMed]
- Quarona, V.; Ferri, V.; Chillemi, A.; Bolzoni, M.; Mancini, C.; Zaccarello, G.; Roato, I.; Morandi, F.; Marimpietri, D.; Faccani, G.; et al. Unraveling the Contribution of Ectoenzymes to Myeloma Life and Survival in the Bone Marrow Niche. Ann. N. Y. Acad. Sci. 2015, 1335, 10–22. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The A2B receptor is regulated by HIF-1α and HIF-2α in osteoclasts. (A) A2B receptor mRNA expression is increased by 24 h exposure to CoCl2 (100 μM) or hypoxia (2% O2). (B) Expression of HIF-1α and A2B receptor protein in two independent osteoclast populations (OC-A, OC-B) after 24 h exposure to 100 μM CoCl2 (Co) or hypoxia (Hx). Densitometric quantification of the A2B receptor is presented below. (C) Immunohistochemistry visualises HIF-1α stabilisation in hypoxic osteoclasts (arrows) and the concomitant increase in A2B receptor expression (red) in multi-nucleated osteoclasts (F-actin ring, green; DAPI, blue). Scale bars = 100 μM. (D–G) Expression of (D) HIF1A, (E) HIF-2A and (F) A2B receptor mRNA and (G) HIF-1α and A2B receptor protein in response to hypoxia (2% O2, 24 h; white bars) after HIF-1α, HIF-2α or HIF1α + HIF-2α siRNA, relative to the scrambled (scr) siRNA control. Densitometric quantification of the A2B receptor is presented below; * p < 0.05, ** p < 0.01, *** p < 0.001; n = 6.
Figure 1.
The A2B receptor is regulated by HIF-1α and HIF-2α in osteoclasts. (A) A2B receptor mRNA expression is increased by 24 h exposure to CoCl2 (100 μM) or hypoxia (2% O2). (B) Expression of HIF-1α and A2B receptor protein in two independent osteoclast populations (OC-A, OC-B) after 24 h exposure to 100 μM CoCl2 (Co) or hypoxia (Hx). Densitometric quantification of the A2B receptor is presented below. (C) Immunohistochemistry visualises HIF-1α stabilisation in hypoxic osteoclasts (arrows) and the concomitant increase in A2B receptor expression (red) in multi-nucleated osteoclasts (F-actin ring, green; DAPI, blue). Scale bars = 100 μM. (D–G) Expression of (D) HIF1A, (E) HIF-2A and (F) A2B receptor mRNA and (G) HIF-1α and A2B receptor protein in response to hypoxia (2% O2, 24 h; white bars) after HIF-1α, HIF-2α or HIF1α + HIF-2α siRNA, relative to the scrambled (scr) siRNA control. Densitometric quantification of the A2B receptor is presented below; * p < 0.05, ** p < 0.01, *** p < 0.001; n = 6.

Figure 2.
A2B receptor inhibitors reduce osteoclast formation and activity in hypoxic culture. Quantified effect of 24 h exposure of mature human osteoclasts cultured on dentine to MRS1754 (2.5 μM) or PSB603 (10 nM) with respect to (A) osteoclast survival (no. VNR-positive osteoclasts present) and (B, C) the area of dentine resorbed per osteoclast. (n = 6). Scale bars = 700 μm. (D) Effect of 24 h acute hypoxic exposure (2% O2, white bars) on the number of TRAP-positive multi-nucleated osteoclasts formed during differentiation (n = 6) and (E, F) the additional effect of MRS1754 or PSB603 on osteoclastogenesis. Grey bars represent normoxia. (n = 3), * p < 0.05, ** p < 0.01. Arrows indicate multi-nucleated osteoclasts, scale bars = 100 μm..
Figure 2.
A2B receptor inhibitors reduce osteoclast formation and activity in hypoxic culture. Quantified effect of 24 h exposure of mature human osteoclasts cultured on dentine to MRS1754 (2.5 μM) or PSB603 (10 nM) with respect to (A) osteoclast survival (no. VNR-positive osteoclasts present) and (B, C) the area of dentine resorbed per osteoclast. (n = 6). Scale bars = 700 μm. (D) Effect of 24 h acute hypoxic exposure (2% O2, white bars) on the number of TRAP-positive multi-nucleated osteoclasts formed during differentiation (n = 6) and (E, F) the additional effect of MRS1754 or PSB603 on osteoclastogenesis. Grey bars represent normoxia. (n = 3), * p < 0.05, ** p < 0.01. Arrows indicate multi-nucleated osteoclasts, scale bars = 100 μm..

Figure 3.
Hypoxic osteoclasts secrete ATP to increase adenosine concentrations. (A) A 24 h exposure of mature human osteoclasts to hypoxia (2% O2, white bars) increases the concentration of extracellular adenosine (n = 4) and (B) increases the expression of ectonucleotidase CD39 and (C) ectonucleotidase CD73 mRNAs. (n = 5). (D–F) Effect of 30–240 min hypoxia on ATP secretion by (D) mature human osteoclasts, (E) CD14+ monocytes and (F) primary human osteoblasts (n = 4). (G) Quantified effect of 24 h exposure of mature human osteoclasts to adenosine (0.5–50 μM) with respect to osteoclast survival (number of vitronectin (VNR)-positive osteoclasts present) and (H) the area of dentine resorbed per osteoclast. (n = 6); * p < 0.05, ** p < 0.01.
Figure 3.
Hypoxic osteoclasts secrete ATP to increase adenosine concentrations. (A) A 24 h exposure of mature human osteoclasts to hypoxia (2% O2, white bars) increases the concentration of extracellular adenosine (n = 4) and (B) increases the expression of ectonucleotidase CD39 and (C) ectonucleotidase CD73 mRNAs. (n = 5). (D–F) Effect of 30–240 min hypoxia on ATP secretion by (D) mature human osteoclasts, (E) CD14+ monocytes and (F) primary human osteoblasts (n = 4). (G) Quantified effect of 24 h exposure of mature human osteoclasts to adenosine (0.5–50 μM) with respect to osteoclast survival (number of vitronectin (VNR)-positive osteoclasts present) and (H) the area of dentine resorbed per osteoclast. (n = 6); * p < 0.05, ** p < 0.01.

Figure 4.
A2B receptor inhibition prevents the HIF-mediated increase in hypoxic glycolysis. Effect of 24 h exposure of mature human osteoclasts to hypoxia (2% O2, white bars) and MRS1754 or PSB603 with respect to (A) glucose consumption, (B) lactate production or (C) mitochondrial reductase activity (Alamar blue fluorescence), all per osteoclast (n = 8). (D) Expression of HIF-1α protein and (E) HRE–luciferase activity (n = 5) after 24 h exposure of mature human osteoclasts to MRS1754 or PSB603 in either normoxic or hypoxic culture conditions. Densitometry for HIF-1α is presented below the blot; * p < 0.05, ** p < 0.01.
Figure 4.
A2B receptor inhibition prevents the HIF-mediated increase in hypoxic glycolysis. Effect of 24 h exposure of mature human osteoclasts to hypoxia (2% O2, white bars) and MRS1754 or PSB603 with respect to (A) glucose consumption, (B) lactate production or (C) mitochondrial reductase activity (Alamar blue fluorescence), all per osteoclast (n = 8). (D) Expression of HIF-1α protein and (E) HRE–luciferase activity (n = 5) after 24 h exposure of mature human osteoclasts to MRS1754 or PSB603 in either normoxic or hypoxic culture conditions. Densitometry for HIF-1α is presented below the blot; * p < 0.05, ** p < 0.01.

© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Knowles, H.J. The Adenosine A2B Receptor Drives Osteoclast-Mediated Bone Resorption in Hypoxic Microenvironments. Cells 2019, 8, 624. https://doi.org/10.3390/cells8060624
AMA Style
Knowles HJ. The Adenosine A2B Receptor Drives Osteoclast-Mediated Bone Resorption in Hypoxic Microenvironments. Cells. 2019; 8(6):624. https://doi.org/10.3390/cells8060624
Chicago/Turabian StyleKnowles, Helen J. 2019. "The Adenosine A2B Receptor Drives Osteoclast-Mediated Bone Resorption in Hypoxic Microenvironments" Cells 8, no. 6: 624. https://doi.org/10.3390/cells8060624
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.