HLA-DQA1 and HLA-DQB1 Alleles, Conferring Susceptibility to Celiac Disease and Type 1 Diabetes, Are More Expressed Than Non-Predisposing Alleles and Are Coordinately Regulated

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients Enrollment and Selection of Antigen Presenting Cells
2.2. Monoclonal Antibodies and Flow Cytometry Analysis
2.3. cDNA Cloning and B-LCL Nucleofection
2.4. RNA Quantization
2.5. T Cell Functional Assay
2.6. Statistical Analysis
3. Results
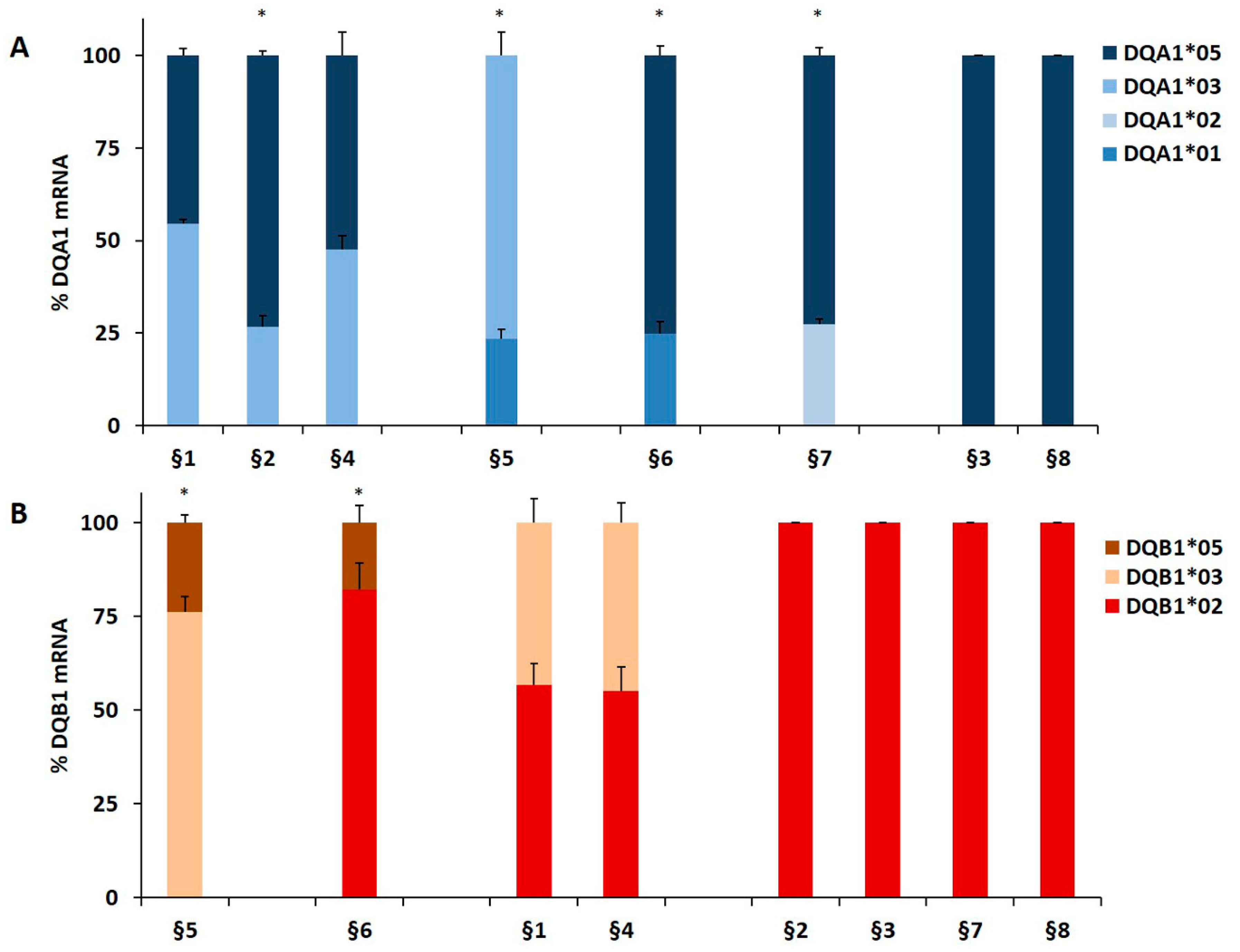
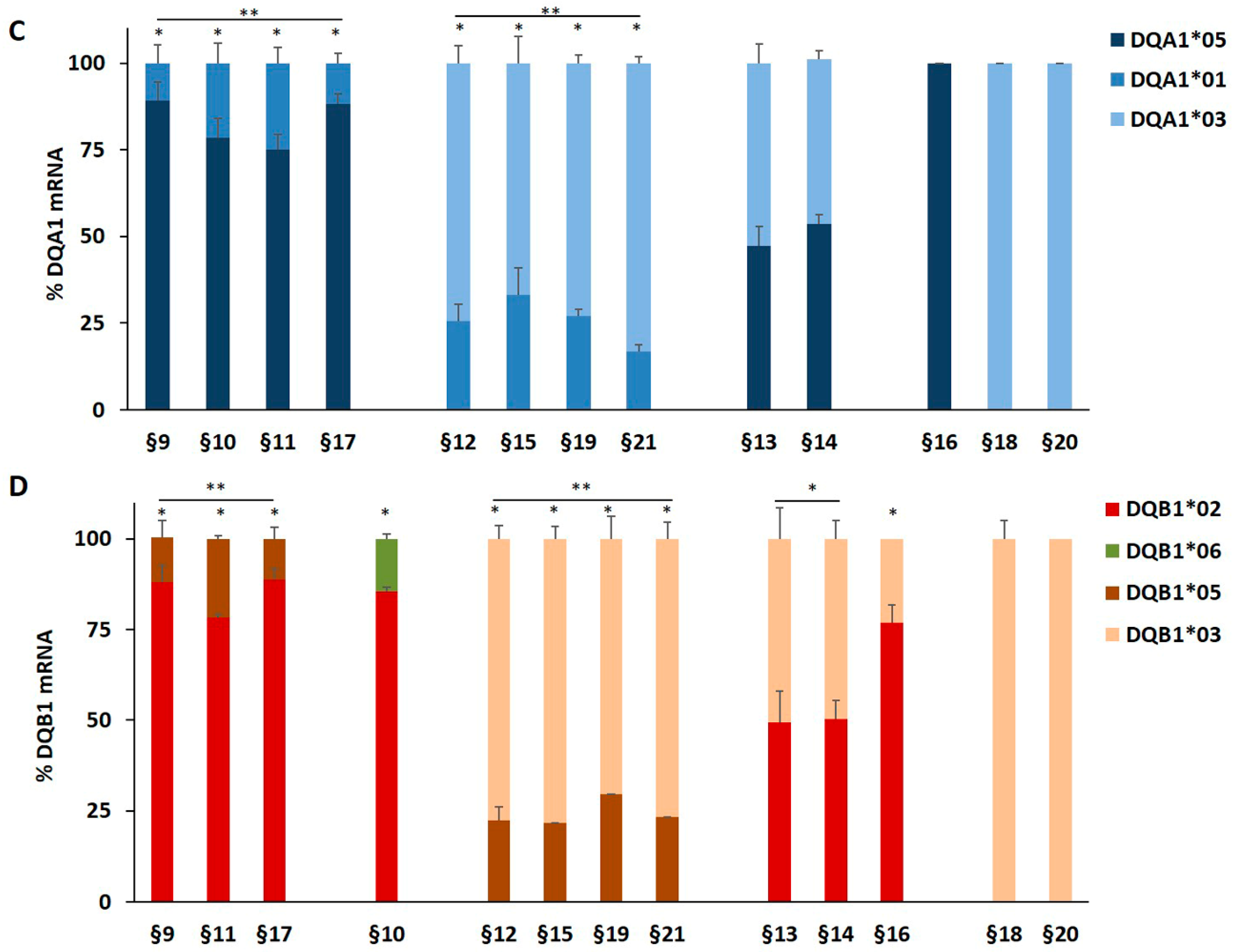
3.1. The DQA1*03 and DQB1*03 mRNA Associated with T1D Risk Were More Abundant than the mRNA of Non-T1D Related Alleles
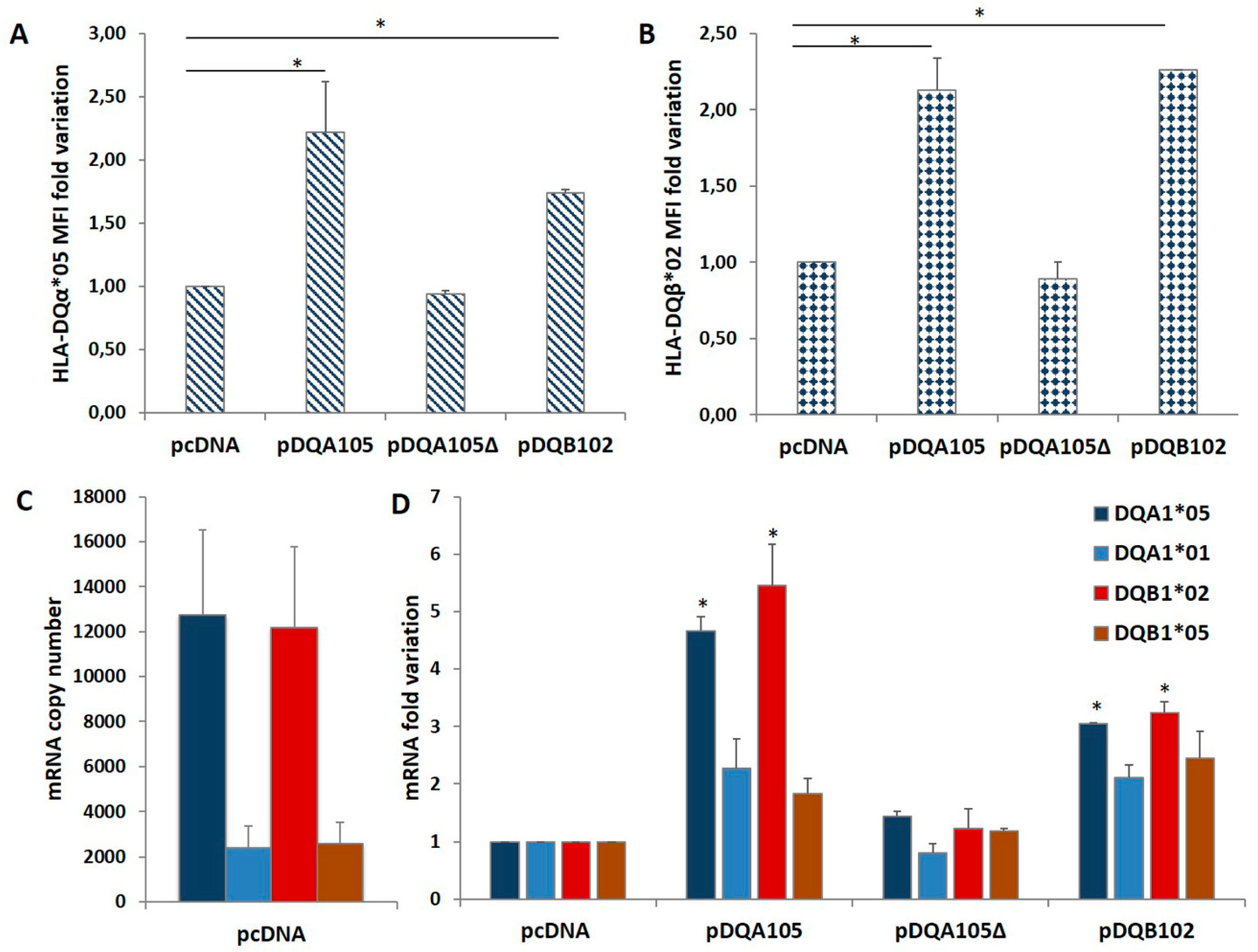
3.2. Analysis of Co-Regulated Expression of DQA1*05 and DQB1*02 mRNA
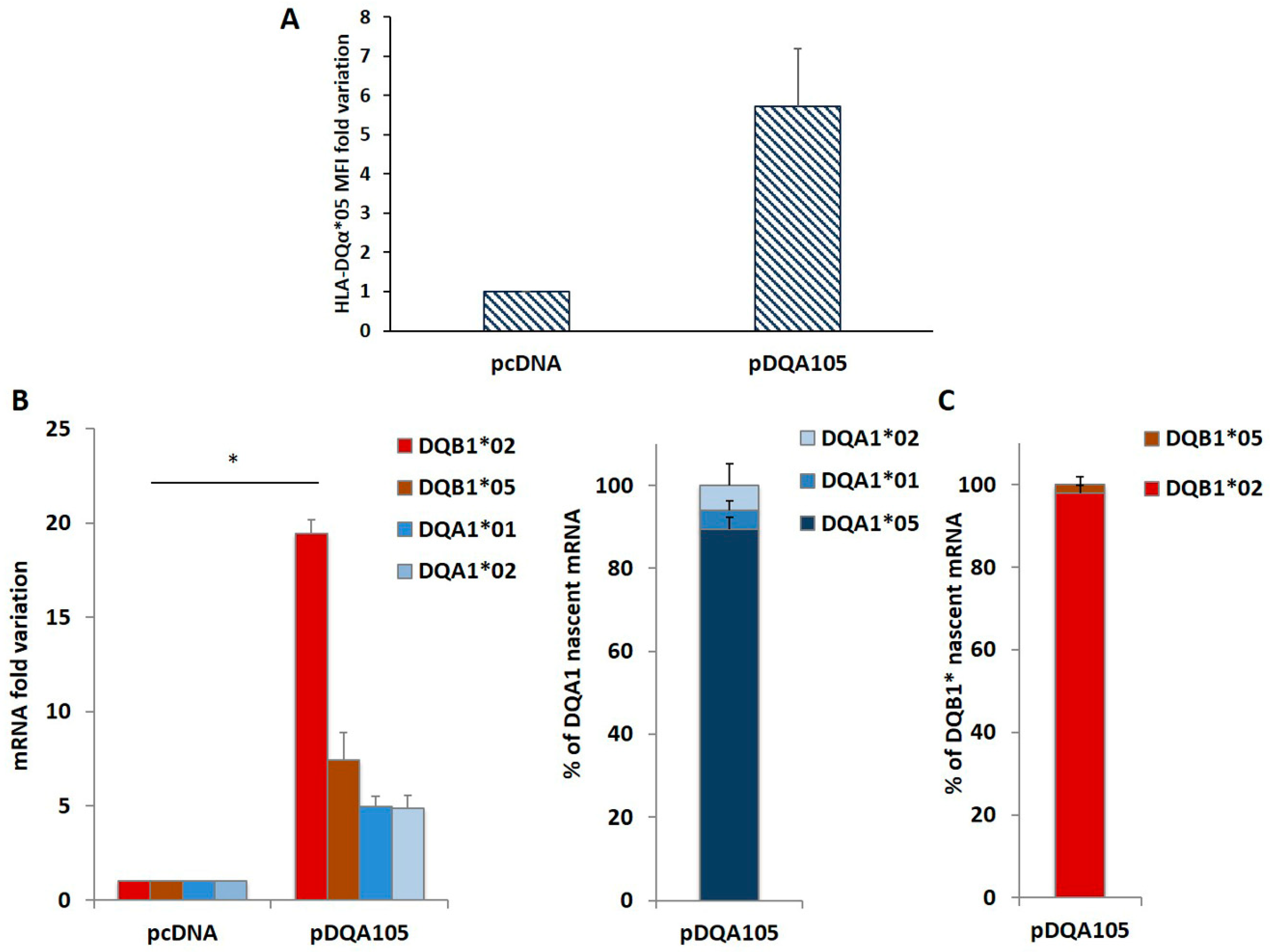
3.3. DR7/DR14 B-LCLs Transfected with pDQA105 Efficiently Stimulated Gluten-Specific CD4+ T Cells
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Romanos, J.; Bakker, S.F.; Onengut-gumuscu, S.; Simsek, S.; Rewers, M.; Mulder, C.J.; Liu, E.; Rich, S.S. Contrasting the Genetic Background of Type 1 Diabetes and Celiac Disease Autoimmunity. Diab. Care 2015, 38, 37–44. [Google Scholar]
- Van Lummel, M.; Duinkerken, G.; Van Veelen, P.A.; De Ru, A.; Cordfunke, R.; Zaldumbide, A.; Gomez-Touriño, I.; Arif, S.; Peakman, M.; Drijfhout, J.W.; et al. Posttranslational modification of HLA-DQ binding islet autoantigens in type 1 diabetes. Diabetes 2014, 63, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Abadie, V.; Sollid, L.M.; Barreiro, L.B.; Jabri, B. Integration of Genetic and Immunological Insights into a Model of Celiac Disease Pathogenesis. Annu. Rev. Immunol. 2011, 29, 493–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, J.A.; Valdes, A.M. Genetics of the HLA region in the prediction of type 1 diabetes. Curr. Diab. Rep. 2011, 11, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Meresse, B.; Malamut, G.; Cerf-Bensussan, N. Celiac Disease: An Immunological Jigsaw. Immunity 2012, 36, 907–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erlich, H.; Valdes, A.M.; Noble, J.; Carlson, J.A.; Varney, M.; Concannon, P.; Mychaleckyj, J.C.; Todd, J.A.; Bonella, P.; Fear, A.L.; et al. HLA DR-DQ Haplotypes and Genotypes and Type 1 Diabetes Risk. Diabetes 2008, 57, 1084–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, J.A. Immunogenetics of type 1 diabetes: A comprehensive review. J. Autoimmun. 2015, 64, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Pisapia, L.; Camarca, A.; Picascia, S.; Bassi, V.; Barba, P.; Del Pozzo, G.; Gianfrani, C. HLA-DQ2.5 genes associated with celiac disease risk are preferentially expressed with respect to non-predisposing HLA genes: Implication for anti-gluten T cell response. J. Autoimmun. 2016, 70, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Corso, C.; Pisapia, L.; Citro, A.; Cicatiello, V.; Barba, P.; Cigliano, L.; Abrescia, P.; Maffei, A.; Manco, G.; Del Pozzo, G. EBP1 and DRBP76/NF90 binding proteins are included in the major histocompatibility complex class II RNA operon. Nucleic Acids Res. 2011. [Google Scholar] [CrossRef] [PubMed]
- Pisapia, L.; Cicatiello, V.; Barba, P.; Malanga, D.; Maffei, A.; Hamilton, R.S.; Del Pozzo, G. Co-regulated expression of alpha and beta mRNAs encoding HLA-DR surface heterodimers is mediated by the MHCII RNA operon. Nucleic Acids Res. 2013, 41, 3772–3786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amar, A.; Radka, S.F.; Holbeck, S.L.; Kim, S.J.; Nepom, B.S.; Nelson, K.; Nepom, G.T. Characterization of specific HLA-DQ alpha allospecificities by genomic, biochemical, and serologic analysis. J. Immunol. 1987, 138, 3986–3990. [Google Scholar] [PubMed]
- Viken, H.D.; Paulsen, G.; Sollid, L.M.; Lundin, K.E.A.; Tjønnfjord, G.E.; Thorsby, E.; Gaudernack, G. Characterization of an HLA-DQ2-specific monoclonal antibody. Influence of amino acid substitutions in DQβ 1*0202. Hum. Immunol. 1995, 42, 319–327. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Camarca, A.; Anderson, R.P.; Mamone, G.; Fierro, O.; Facchiano, A.; Costantini, S.; Zanzi, D.; Sidney, J.; Auricchio, S.; Sette, A.; et al. Intestinal T cell responses to gluten peptides are largely heterogeneous: Implications for a peptide-based therapy in celiac disease. J.Imunol. 2009, 182, 4158–4166. [Google Scholar] [CrossRef] [PubMed]
- Vader, W.; Stepniak, D.; Kooy, Y.; Mearin, L.; Thompson, A.; van Rood, J.J.; Spaenij, L.; Koning, F. The HLA-DQ2 gene dose effect in celiac disease is directly related to the magnitude and breadth of gluten-specific T cell responses. Proc. Natl. Acad. Sci. USA 2003, 100, 12390–12395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalli, G.; Hayashi, M.; Jin, Y.; Yorgov, D.; Santorico, S.A.; Holcomb, C.; Spritz, R.A.; Dinarello, C.A. MHC class II super-enhancer increases surface expression of HLA-DR and HLA-DQ and affects cytokine production in autoimmune vitiligo. Proc. Natl. Acad. Sci. USA 2015. [Google Scholar] [CrossRef] [PubMed]
- Raj, P.; Rai, E.; Song, R.; Khan, S.; Wakeland, B.E.; Viswanathan, K.; Arana, C.; Liang, C.; Zhang, B.; Dozmorov, I.; et al. Regulatory polymorphisms modulate the expression of HLA class II molecules and promote autoimmunity. eLife 2016, 5, e12089. [Google Scholar] [CrossRef] [PubMed]
- Gianfrani, C.; Pisapia, L.; Picascia, S.; Strazzullo, M.; Del Pozzo, G. Expression level of risk genes of MHC class II is a susceptibility factor for autoimmunity: New insights. J. Autoimmun. 2018, 89, 1–10. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Cells a | Diagnosis | DQA1 Genotype | DQB1 Genotype | DQ Phenotype | DR Phenotype |
|---|---|---|---|---|---|---|
| §1 | B-LCL | T1D | *03/*05 | *02/*03 | DQ2.5/DQ8 | DR3/DR4 |
| §2 | B-LCL | T1D | *03/*05 | *02/*02 | DQ2.5/DQ2.3 | DR3/DR9 |
| §3 | B-LCL | T1D | *05/*05 | *02/*02 | DQ2.5/DQ2.5 | DR3/DR3 |
| §4 | B-LCL | T1D with CD | *03/*05 | *02/*03 | DQ2.5/DQ8 | DR3/DR4 |
| §5 | B-LCL | T1D with CD | *01/*03 | *05/*03 | DQ8/DQ5 | DR4/DR16 |
| §6 | B-LCL | T1D with CD | *01/*05 | *02/*05 | DQ2.5/DQ5 | DR1/DR3 |
| §7 | B-LCL | T1D with CD | *02/*05 | *02/*02 | DQ2.5/DQ2.2 | DR3/DR7 |
| §8 | B-LCL | T1D with CD | *05/*05 | *02/*02 | DQ2.5/DQ2.5 | DR3/DR3 |
| §9 | PBMC | T1D | *01/*05 | *02/*05 | DQ2.5/DQ5 | DR1/DR3 |
| §10 | PBMC | T1D | *01/*05 | *02/*06 | DQ2.5/DQ6 | DR3/DR15 |
| §11 | PBMC | T1D | *01/*05 | *02/*05 | DQ2.5/DQ5 | DR1/DR3 |
| §12 | PBMC | T1D | *01/*03 | *03/*05 | DQ8/DQ5 | DR4/DR1 |
| §13 | PBMC | T1D | *03/*05 | *02/*03 | DQ2.5/DQ8 | DR3/DR4 |
| §14 | PBMC | T1D | *03/*05 | *02/*03 | DQ2.5/DQ8 | DR3/DR4 |
| §15 | PBMC | T1D | *01/*03 | *03/*05 | DQ8/DQ5 | DR4/DR1 |
| §16 | PBMC | T1D | *05/*05 | *02/*03 | DQ2.5/DQ7 | DR3/DR5 |
| §17 | PBMC | T1D | *01/*05 | *02/*05 | DQ2.5/DQ5 | DR3/DR1 |
| §18 | PBMC | T1D | *03/*03 | *03/*03 | DQ8/DQ8 | DR4/DR4 |
| §19 | PBMC | T1D | *01/*03 | *03/*05 | DQ8/DQ5 | DR4/DR1 |
| §20 | PBMC | T1D | *03/*03 | *03/*03 | DQ8/DQ8 | DR4/DR4 |
| §21 | PBMC | T1D | *01/*03 | *03/*05 | DQ8/DQ5 | DR4/DR1 |
| Gene | Primers | Sequences 5′ → 3′ |
|---|---|---|
| β-Actin | ACT-F ACT-R | TCATGAAGTGTGACGTTGACA CCTAGAAGCATTTGCGGTGCAC |
| HLA-DQA1*01 | DQA1*01-F DQA1*R | CGGTGGCCTGAGTTCAGCAA GGAGACTTGGAAAACACTGTGACC |
| HLA-DQA1*02 | DQA1*02-F DQA1*R | AAGTTGCCTCTGTTCCACAGAC GGAGACTTGGAAAACACTGTGACC |
| HLA-DQA1*03 | DQA1*03-F DQA1*R | CTCTGTTCCGCAGATTTAGAAGA GGAGACTTGGAAAACACTGTGACC |
| HLA-DQA1*05 | DQA1*05-F DQA1*R | CTCTGTTCCGCAGATTTAGAAGA GGAGACTTGGAAAACACTGTGACC |
| HLA-DQB1*02 | DQB1*02-F DQB1*R | TCTTGTGAGCAGAAGCATCT CAGGATCTGGAAGGTCCAGT |
| HLA-DQB1*03 | DQB1*03-F DQB1*R | CGGAGTTGGACACGGTGTGC CAGGATCTGGAAGGTCCAGT |
| HLA-DQB1*05 | DQB1*05 DQB1*R | ACAACTACGAGGTGGCGTACC CAGGATCTGGAAGGTCCAGT |
| HLA-DQB1*06 | DQB1*06-F DQB1*R | CAGATCAAAGTCCGGTGGTTTC CAGGATCTGGAAGGTCCAGT |
| Cells | Patients | mRNA Alleles | Fold Change | Cells | Patients | mRNA Alleles | Fold Change |
|---|---|---|---|---|---|---|---|
| B-LCL | §1 | DQA1*05 = DQA1*03 | 1 | B-LCL | §5 | DQB1*03 > DQB1*05 | 3.2 |
| §4 | |||||||
| B-LCL | §2 | DQA1*05 > DQA1*03 | 2.7 | B-LCL | §6 | DQB1*02 > DQB1*05 | 4.6 |
| B-LCL | §5 | DQA1*03 > DQA1*01 | 3.2 | B-LCL | §1 | DQB1*02 = DQB1*03 | 1 |
| §4 | |||||||
| B-LCL | §6 | DQA1*05 > DQA1*01 | 3.0 | PBMC | §9 | DQB1*02 > DQB1*05 | 5.6 |
| §11 | |||||||
| §17 | |||||||
| B-LCL | §7 | DQA1*05 > DQA1*02 | 2.7 | PBMC | §10 | DQB1*02 > DQB1*06 | 5.9 |
| PBMC | §9 | DQA1*05 > DQA1*01 | 4.8 | PBMC | §19 | DQB1*03 > DQB1*05 | 3.1 |
| §10 | §21 | ||||||
| §11 | §12 | ||||||
| §17 | §15 | ||||||
| PBMC | §12 | DQA1*03 > DQA1*01 | 2.9 | PBMC | §13 | DQB1*02 = DQB1*03 | 1 |
| §15 | |||||||
| §19 | §14 | ||||||
| §21 | |||||||
| PBMC | §13 | DQA1*05 = DQA1*03 | 1 | PBMC | §16 | DQB1*02 > DQB1*03 | 3.3 |
| §14 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farina, F.; Picascia, S.; Pisapia, L.; Barba, P.; Vitale, S.; Franzese, A.; Mozzillo, E.; Gianfrani, C.; Del Pozzo G, G. HLA-DQA1 and HLA-DQB1 Alleles, Conferring Susceptibility to Celiac Disease and Type 1 Diabetes, Are More Expressed Than Non-Predisposing Alleles and Are Coordinately Regulated. Cells 2019, 8, 751. https://doi.org/10.3390/cells8070751
Farina F, Picascia S, Pisapia L, Barba P, Vitale S, Franzese A, Mozzillo E, Gianfrani C, Del Pozzo G G. HLA-DQA1 and HLA-DQB1 Alleles, Conferring Susceptibility to Celiac Disease and Type 1 Diabetes, Are More Expressed Than Non-Predisposing Alleles and Are Coordinately Regulated. Cells. 2019; 8(7):751. https://doi.org/10.3390/cells8070751
Chicago/Turabian StyleFarina, Federica, Stefania Picascia, Laura Pisapia, Pasquale Barba, Serena Vitale, Adriana Franzese, Enza Mozzillo, Carmen Gianfrani, and Giovanna Del Pozzo G. 2019. "HLA-DQA1 and HLA-DQB1 Alleles, Conferring Susceptibility to Celiac Disease and Type 1 Diabetes, Are More Expressed Than Non-Predisposing Alleles and Are Coordinately Regulated" Cells 8, no. 7: 751. https://doi.org/10.3390/cells8070751
APA StyleFarina, F., Picascia, S., Pisapia, L., Barba, P., Vitale, S., Franzese, A., Mozzillo, E., Gianfrani, C., & Del Pozzo G, G. (2019). HLA-DQA1 and HLA-DQB1 Alleles, Conferring Susceptibility to Celiac Disease and Type 1 Diabetes, Are More Expressed Than Non-Predisposing Alleles and Are Coordinately Regulated. Cells, 8(7), 751. https://doi.org/10.3390/cells8070751