RNA-Guided Genomic Localization of H2A.L.2 Histone Variant

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Spermatogenic Cell Fraction Purification
2.2. Quantification of Expression of Major Satellite Sequences in Male Germ Cells
2.2.1. RNA Extraction from Male Germ Cells
2.2.2. Sequencing
2.2.3. Post Processing of Fastq Files
2.3. Cell Culture and Transfection
2.4. RNA Fluorescence In Situ Hybridization
2.4.1. Preparation of Probes
2.4.2. Hybridization
2.5. Immunofluorescence on Germ Cells
2.6. Microscope Analysis and Image Processing
2.7. Ethics
3. Results
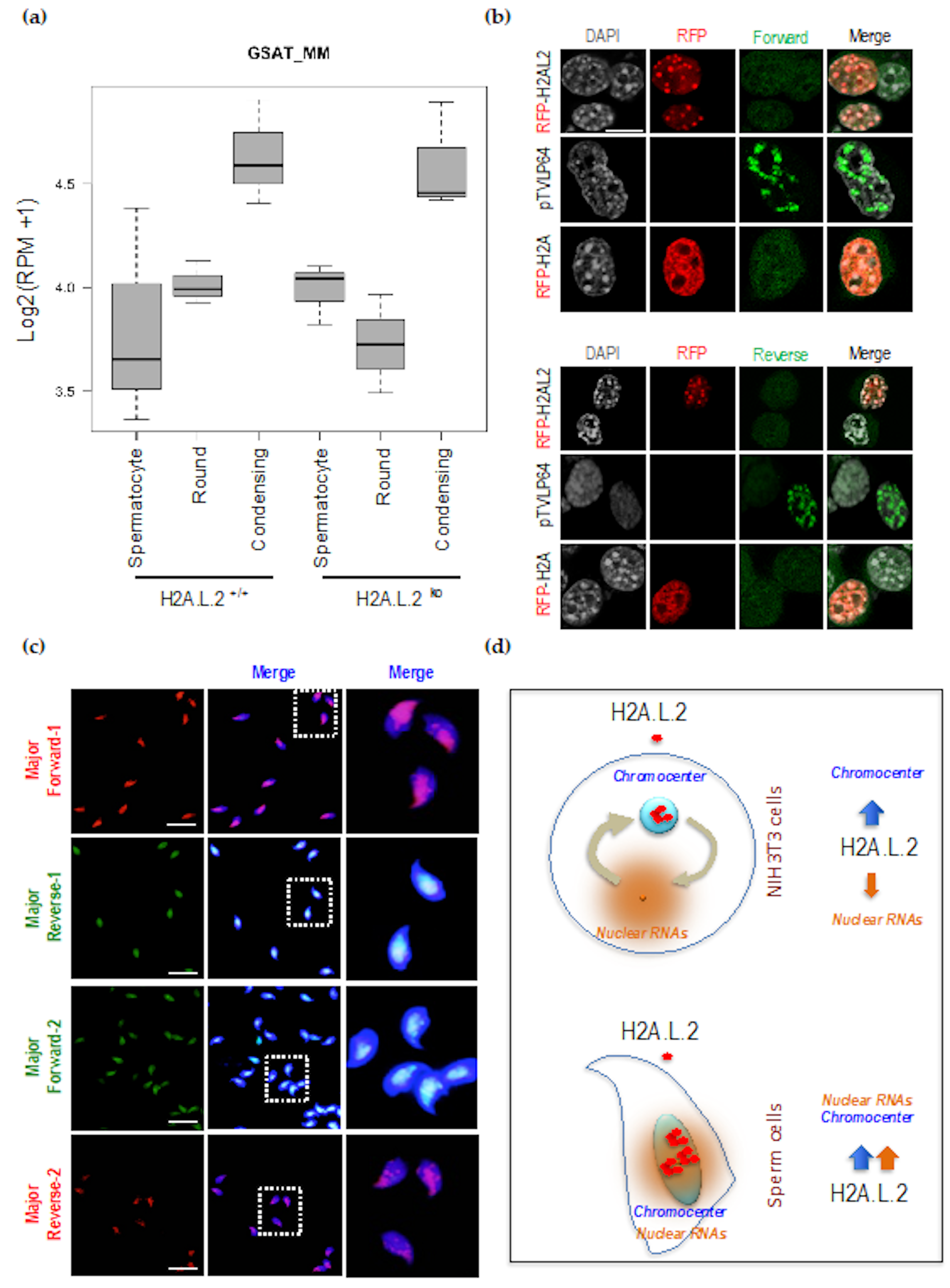
3.1. H2A.L.2 Has the Intrinsic Property to Target Pericentric Heterochromatin
3.2. Pericentric Localization of H2A.L.2 is Controlled by its N-terminal RNA-Binding Motif
3.3. The Dynamic Turnover of H2A.L.2 is Controlled by its N-terminal RNA Binding Domain
3.4. Enhanced Major Satellite RNA Expression and Localization during Late Spermatogenesis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shiota, H.; Barral, S.; Buchou, T.; Tan, M.; Couté, Y.; Charbonnier, G.; Reynoird, N.; Boussouar, F.; Gérard, M.; Zhu, M.; et al. Nut Directs p300-Dependent, Genome-Wide H4 Hyperacetylation in Male Germ Cells. Cell Rep. 2018, 24, 3477–3487.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaucher, J.; Boussouar, F.; Montellier, E.; Curtet, S.; Buchou, T.; Bertrand, S.; Hery, P.; Jounier, S.; Depaux, A.; Vitte, A.-L.; et al. Bromodomain-dependent stage-specific male genome programming by Brdt. EMBO J. 2012, 31, 3809–3820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goudarzi, A.; Zhang, D.; Huang, H.; Barral, S.; Kwon, O.K.; Qi, S.; Tang, Z.; Buchou, T.; Vitte, A.-L.; He, T.; et al. Dynamic Competing Histone H4 K5K8 Acetylation and Butyrylation Are Hallmarks of Highly Active Gene Promoters. Mol. Cell 2016, 62, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govin, J.; Escoffier, E.; Rousseaux, S.; Kuhn, L.; Ferro, M.; Thévenon, J.; Catena, R.; Davidson, I.; Garin, J.; Khochbin, S.; et al. Pericentric heterochromatin reprogramming by new histone variants during mouse spermiogenesis. J. Cell Biol. 2007, 176, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Montellier, E.; Boussouar, F.; Rousseaux, S.; Zhang, K.; Buchou, T.; Fenaille, F.; Shiota, H.; Debernardi, A.; Héry, P.; Curtet, S.; et al. Chromatin-to-nucleoprotamine transition is controlled by the histone H2B variant TH2B. Genes Dev. 2013, 27, 1680–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barral, S.; Morozumi, Y.; Tanaka, H.; Montellier, E.; Govin, J.; de Dieuleveult, M.; Charbonnier, G.; Couté, Y.; Puthier, D.; Buchou, T.; et al. Histone Variant H2A.L.2 Guides Transition Protein-Dependent Protamine Assembly in Male Germ Cells. Mol. Cell 2017, 66, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Brykczynska, U.; Hisano, M.; Erkek, S.; Ramos, L.; Oakeley, E.J.; Roloff, T.C.; Beisel, C.; Schübeler, D.; Stadler, M.B.; Peters, A.H.F.M. Repressive and active histone methylation mark distinct promoters in human and mouse spermatozoa. Nat. Struct. Mol. Biol. 2010, 17, 679–687. [Google Scholar] [CrossRef]
- Carone, B.R.; Hung, J.-H.; Hainer, S.J.; Chou, M.-T.; Carone, D.M.; Weng, Z.; Fazzio, T.G.; Rando, O.J. High-resolution mapping of chromatin packaging in mouse embryonic stem cells and sperm. Dev. Cell 2014, 30, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Erkek, S.; Hisano, M.; Liang, C.-Y.; Gill, M.; Murr, R.; Dieker, J.; Schübeler, D.; van der Vlag, J.; Stadler, M.B.; Peters, A.H.F.M. Molecular determinants of nucleosome retention at CpG-rich sequences in mouse spermatozoa. Nat. Struct. Mol. Biol. 2013, 20, 868–875. [Google Scholar] [CrossRef] [Green Version]
- Hammoud, S.S.; Nix, D.A.; Zhang, H.; Purwar, J.; Carrell, D.T.; Cairns, B.R. Distinctive chromatin in human sperm packages genes for embryo development. Nature 2009, 460, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Samans, B.; Yang, Y.; Krebs, S.; Sarode, G.V.; Blum, H.; Reichenbach, M.; Wolf, E.; Steger, K.; Dansranjavin, T.; Schagdarsurengin, U. Uniformity of nucleosome preservation pattern in Mammalian sperm and its connection to repetitive DNA elements. Dev. Cell 2014, 30, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, K.; Hada, M.; Fukuda, Y.; Inoue, E.; Makino, Y.; Katou, Y.; Shirahige, K.; Okada, Y. Re-evaluating the Localization of Sperm-Retained Histones Revealed the Modification-Dependent Accumulation in Specific Genome Regions. Cell Rep. 2018, 23, 3920–3932. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Muratani, M.; Araki, H.; Miura, F.; Suzuki, T.; Dohmae, N.; Katou, Y.; Shirahige, K.; Ito, T.; Ishii, S. Mapping of histone-binding sites in histone replacement-completed spermatozoa. Nat. Commun. 2018, 9, 3885. [Google Scholar] [CrossRef]
- Royo, H.; Stadler, M.B.; Peters, A.H.F.M. Alternative Computational Analysis Shows No Evidence for Nucleosome Enrichment at Repetitive Sequences in Mammalian Spermatozoa. Dev. Cell 2016, 37, 98–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soboleva, T.A.; Nekrasov, M.; Pahwa, A.; Williams, R.; Huttley, G.A.; Tremethick, D.J. A unique H2A histone variant occupies the transcriptional start site of active genes. Nat. Struct. Mol. Biol. 2011, 19, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Miyanari, Y.; Ziegler-Birling, C.; Torres-Padilla, M.-E. Live visualization of chromatin dynamics with fluorescent TALEs. Nat. Struct. Mol. Biol. 2013, 20, 1321–1324. [Google Scholar] [CrossRef] [PubMed]
- Molaro, A.; Young, J.M.; Malik, H.S. Evolutionary origins and diversification of testis-specific short histone H2A variants in mammals. Genome Res. 2018, 28, 460–473. [Google Scholar] [CrossRef] [Green Version]
- Syed, S.H.; Boulard, M.; Shukla, M.S.; Gautier, T.; Travers, A.; Bednar, J.; Faivre-Moskalenko, C.; Dimitrov, S.; Angelov, D. The incorporation of the novel histone variant H2AL2 confers unusual structural and functional properties of the nucleosome. Nucleic Acids Res. 2009, 37, 4684–4695. [Google Scholar] [CrossRef] [Green Version]
- Soboleva, T.A.; Parker, B.J.; Nekrasov, M.; Hart-Smith, G.; Tay, Y.J.; Tng, W.-Q.; Wilkins, M.; Ryan, D.; Tremethick, D.J. A new link between transcriptional initiation and pre-mRNA splicing: The RNA binding histone variant H2A.B. PLoS Genet. 2017, 13, e1006633. [Google Scholar] [CrossRef]
- Audas, T.E.; Jacob, M.D.; Lee, S. Immobilization of proteins in the nucleolus by ribosomal intergenic spacer noncoding RNA. Mol. Cell 2012, 45, 147–157. [Google Scholar] [CrossRef]
- Dueva, R.; Akopyan, K.; Pederiva, C.; Trevisan, D.; Dhanjal, S.; Lindqvist, A.; Farnebo, M. Neutralization of the Positive Charges on Histone Tails by RNA Promotes an Open Chromatin Structure. Cell Chem. Biol. 2019, 26, 1436–1449.e5. [Google Scholar] [CrossRef] [PubMed]
- Arimura, Y.; Kimura, H.; Oda, T.; Sato, K.; Osakabe, A.; Tachiwana, H.; Sato, Y.; Kinugasa, Y.; Ikura, T.; Sugiyama, M.; et al. Structural basis of a nucleosome containing histone H2A.B/H2A.Bbd that transiently associates with reorganized chromatin. Sci. Rep. 2013, 3, 3510. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Konesky, K.; Park, Y.-J.; Rosu, S.; Dyer, P.N.; Rangasamy, D.; Tremethick, D.J.; Laybourn, P.J.; Luger, K. Nucleosomes containing the histone variant H2A.Bbd organize only 118 base pairs of DNA. EMBO J. 2004, 23, 3314–3324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyen, C.-M.; An, W.; Angelov, D.; Bondarenko, V.; Mietton, F.; Studitsky, V.M.; Hamiche, A.; Roeder, R.G.; Bouvet, P.; Dimitrov, S. Mechanism of polymerase II transcription repression by the histone variant macroH2A. Mol. Cell. Biol. 2006, 26, 1156–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nekrasov, M.; Amrichova, J.; Parker, B.J.; Soboleva, T.A.; Jack, C.; Williams, R.; Huttley, G.A.; Tremethick, D.J. Histone H2A.Z inheritance during the cell cycle and its impact on promoter organization and dynamics. Nat. Struct. Mol. Biol. 2012, 19, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
- Saitou, M.; Kurimoto, K. Paternal nucleosomes: are they retained in developmental promoters or gene deserts? Dev. Cell 2014, 30, 6–8. [Google Scholar] [CrossRef] [Green Version]
- Mietton, F.; Sengupta, A.K.; Molla, A.; Picchi, G.; Barral, S.; Heliot, L.; Grange, T.; Wutz, A.; Dimitrov, S. Weak but uniform enrichment of the histone variant macroH2A1 along the inactive X chromosome. Mol. Cell. Biol. 2009, 29, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Perche, P.Y.; Vourc’h, C.; Konecny, L.; Souchier, C.; Robert-Nicoud, M.; Dimitrov, S.; Khochbin, S. Higher concentrations of histone macroH2A in the Barr body are correlated with higher nucleosome density. Curr. Biol. 2000, 10, 1531–1534. [Google Scholar] [CrossRef] [Green Version]
- Yelagandula, R.; Stroud, H.; Holec, S.; Zhou, K.; Feng, S.; Zhong, X.; Muthurajan, U.M.; Nie, X.; Kawashima, T.; Groth, M.; et al. The histone variant H2A.W defines heterochromatin and promotes chromatin condensation in Arabidopsis. Cell 2014, 158, 98–109. [Google Scholar] [CrossRef] [Green Version]
- Borsos, M.; Torres-Padilla, M.-E. Building up the nucleus: nuclear organization in the establishment of totipotency and pluripotency during mammalian development. Genes Dev. 2016, 30, 611–621. [Google Scholar] [CrossRef]
- Probst, A.V.; Okamoto, I.; Casanova, M.; El Marjou, F.; Le Baccon, P.; Almouzni, G. A strand-specific burst in transcription of pericentric satellites is required for chromocenter formation and early mouse development. Dev. Cell 2010, 19, 625–638. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Plasmid | Source |
|---|---|
| pTag-RFP-H2A.L.2 | This paper |
| pTag-RFP-H2A | This paper |
| pTag-RFP-H2A.L.2-H2Antere | This paper |
| pTag-RFP-H2A.B.3 | This paper |
| pTVLP64-ms-Ca B15 | Gift from Maria Elena Torres-Padilla |
| pLKO.1 major satellite tandem repeats | This Paper |
| Oligonucleotide | Sequence (5′ to 3′) |
|---|---|
| Forward 1 | FITC-TCTTGCCATATTCCACGTCC |
| Forward 2 | Cy3-GATTTCGTCATTTTTCAAGT |
| Reverse 1 | Cy3-GCGAGGAAAACTGAAAAAGG |
| Reverse 2 | FITC-GCGAGAAAACTGAAAATCAC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoghoughi, N.; Barral, S.; Curtet, S.; Chuffart, F.; Charbonnier, G.; Puthier, D.; Buchou, T.; Rousseaux, S.; Khochbin, S. RNA-Guided Genomic Localization of H2A.L.2 Histone Variant. Cells 2020, 9, 474. https://doi.org/10.3390/cells9020474
Hoghoughi N, Barral S, Curtet S, Chuffart F, Charbonnier G, Puthier D, Buchou T, Rousseaux S, Khochbin S. RNA-Guided Genomic Localization of H2A.L.2 Histone Variant. Cells. 2020; 9(2):474. https://doi.org/10.3390/cells9020474
Chicago/Turabian StyleHoghoughi, Naghmeh, Sophie Barral, Sandrine Curtet, Florent Chuffart, Guillaume Charbonnier, Denis Puthier, Thierry Buchou, Sophie Rousseaux, and Saadi Khochbin. 2020. "RNA-Guided Genomic Localization of H2A.L.2 Histone Variant" Cells 9, no. 2: 474. https://doi.org/10.3390/cells9020474