The Emerging Roles of Exosomes as EMT Regulators in Cancer
by
 , ,
, ,
Hyunwoo Kim
1,†, 
Sungmin Lee
1,†,
Eunguk Shin
1,
Ki Moon Seong
2,
Young Woo Jin
2,
HyeSook Youn
3,* and
BuHyun Youn
1,4,* 1
Department of Integrated Biological Science, Pusan National University, Busan 46241, Korea
2
Laboratory of Low Dose Risk Assessment, National Radiation Emergency Medical Center, Korea Institute of Radiological & Medical Sciences, Seoul 01812, Korea
3
Department of Integrative Bioscience and Biotechnology, Sejong University, Seoul 05006, Korea
4
Department of Biological Sciences, Pusan National University, Busan 46241, Korea
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Cells 2020, 9(4), 861; https://doi.org/10.3390/cells9040861
Submission received: 14 March 2020
/
Revised: 28 March 2020
/
Accepted: 31 March 2020
/
Published: 2 April 2020
(This article belongs to the Special Issue The Multifaceted Nature of Epithelial to Mesenchymal Transition (EMT): From Embryonic Development to Cellular Plasticity in Normal Tissue and Tumors)
Abstract
:Epithelial–mesenchymal transition (EMT) causes epithelial cells to lose their polarity and adhesion property, and endows them with migratory and invasive properties to enable them to become mesenchymal stem cells. EMT occurs throughout embryonic development, during wound healing, and in various pathological processes, including tumor progression. Considerable research in the last few decades has revealed that EMT is invariably related to tumor aggressiveness and metastasis. Apart from the interactions between numerous intracellular signaling pathways known to regulate EMT, extracellular modulators in the tumor microenvironment also influence tumor cells to undergo EMT, with extracellular vesicles (EVs) receiving increasing attention as EMT inducers. EVs comprise exosomes and microvesicles that carry proteins, nucleic acids, lipids, and other small molecules to stimulate EMT in cells. Among EVs, exosomes have been investigated in many studies, and their role has been found to be significant with respect to regulating intercellular communications. In this review, we summarize recent studies on exosomes and their cargoes that induce cancer-associated EMT. Furthermore, we describe the possible applications of exosomes as promising therapeutic strategies.
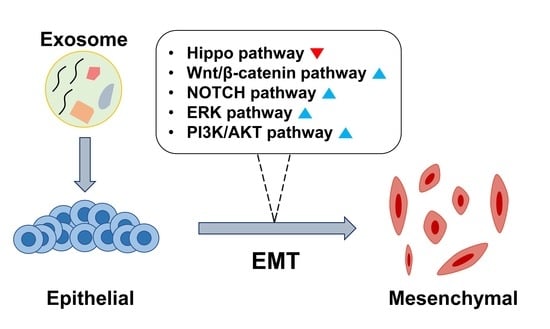
1. Introduction
Epithelial–mesenchymal transition (EMT) is a reversible cellular process that transforms epithelial cells into quasi-mesenchymal cell states. It is observed during normal development, embryonic development, wound healing, and certain pathological processes. During EMT, cells lose their adhesion property, apical–basal polarity, and basement anchoring, and gain migratory and invasive properties. In tumor cells, EMT induces invasiveness and metastatic properties that can lead to malignancy, resulting in poor prognosis for the patient [1]. Additionally, EMT often induces the development of therapeutic resistance through acquisition of stemness and inhibition of tumor cell death [2,3,4]. Although several studies have investigated the suppression of EMT in tumor cells, the underlying mechanisms have not yet been fully elucidated. Many studies have identified the intracellular signaling pathways involved in EMT and their corresponding regulators. Several tumor-cell-derived growth factors, such as transforming growth factor β (TGF-β), hepatocyte growth factor (HGF), epidermal growth factor (EGF), fibroblast growth factor (FGF), Wnt ligand, interleukin-6 (IL-6), and hedgehogs act as extracellular EMT inducers and activate downstream signaling, including the TGF-β/SMAD, PI3K/ protein kinase B (AKT), mitogen-activated protein kinases (MAPKs), and Wnt/β-catenin signaling pathways [5]. Although downstream transcription factors including β-catenin, Zinc finger protein SNAI1 (SNAIL)/Zinc Finger Protein SNAI2 (SLUG), Zinc finger E-box-binding homeobox 1/2 (ZEB1/2), and Twist-related protein 1 (TWIST) govern EMT-related gene expression, recent studies suggest that controlling the expression and function of these factors does not completely inhibit EMT induction in tumor cells [6,7,8].
Recently, extracellular vesicles (EVs) have been identified as important communicators between tumor cells and their microenvironment. EVs are lipid-bilayer-delimited particles that are released from cells, and are usually categorized into at least three classes: exosomes, microvesicles, and apoptotic bodies. They carry proteins, nucleic acids, lipids, metabolites, and even organelles from donor cells. EVs enter recipient cells via endocytosis and release their bioactive contents into the cytosolic space. Among the three types of EVs, previous studies have found that cancer-derived exosomes considerably induce EMT in recipient cells, and identified the major molecules contained in the exosomes [9,10,11]. Furthermore, it has been reported that regulation of exosomal bioactive molecules such as nucleotides, proteins, and organic intermediates can be employed as a strategy to suppress EMT.
In this review, we summarize the EMT-regulating molecules contained in exosomes and describe the associated molecular mechanisms. In addition, we suggest that exosomal contents can potentially be used for predicting tumor prognosis and in the formulation of personalized therapies to overcome tumor EMT induction.
2. Exosome-Induced EMT
Exosomes are secretory endosome-derived membranous vesicles with a size of ~40–100 nm, secreted from a various types of mammalian cells. The term “exosome” was first coined by Trams et al. in the early 1980s. Exosomes were initially assumed to be garbage disposal agents; however, recent data have revealed the direct role of exosomes in governing the physiological and pathological conditions of the surrounding cells by transferring information from donor cells to recipient cells (Figure 1). The generation of exosomes begins from endocytosis to form early endosomes and multivesicular endosomes (MVEs) are formed by inward budding of late endosomes. The MVEs merge with the cell membrane and release the intraluminal endosomal vesicles to become exosomes into the extracellular space [12,13]. Exosome can transport all the biomolecules, including lipids, proteins, DNAs, mRNAs, and miRNA [14]. Most of the contents in exosomes are proteins related with biogenesis and transportation ability of exosomes. Tetraspanins (CD9, CD63, CD81, CD82) take charge of cell penetration, invasion, and fusion events. MVB formation proteins (Alix, TSG101) are involved in exosome release. Annexins and Rab protein have roles in membrane transport and fusion. Alix, fotillin, and TSG101 are participants in exosome biogenesis [15,16]. These exosome abundant proteins (TSG101, HSP70, CD81, CD63) are commonly used as exosomal marker proteins [17]. Besides, other exosomal contents involved in the intercellular crosstalk in the tumor microenvironment plays a large role in cancer development through the formation of a mesenchymal niche. Recently, many studies have been conducted to investigate the significant involvement of exosomes in the induction of cancer-related EMT. We summarized these reports and proposed that the critical components in exosomes can serve as promising EMT regulators, based on the signaling pathways and exosome-derived materials involved.
In the next section, we describe the major exosomal cargo molecules (oligonucleotides, proteins, lipids, etc.) that cause the activation of EMT-related signaling pathways (Figure 2).
2.1. Hippo Pathway
Numerous studies have shown that the Hippo tumor suppressor signaling pathway has a crucial role in restricting organ size by inhibiting the oncogenic co-activators Yes-associated protein (YAP)/transcriptional coactivator with PDZ-binding motif (TAZ) [18,19,20]. Upon inactivation of the Hippo pathway, unphosphorylated YAP/TAZ translocates to the nucleus and bind to the TEA Domain Transcription Factor (TEAD) transcription factor [21,22]. TEAD-mediated target gene transcription increases the expression of mesenchymal markers such as Vimentin and N-cadherin, while simultaneously inhibiting the expression of epithelial markers including E-cadherin [23,24,25,26]. Additionally, YAP interacts with EMT-induced transcription factors ZEB1, SNAIL, and SLUG, and their complex promotes cancer stem cell (CSC) traits [27,28,29]. Several recent studies have revealed that exosomes could induce EMT through modulation of the Hippo pathway. Exosomes from mesenchymal-stem-cell-derived adipocytes promoted Hippo-mediated EMT in breast cancer cells, while their principal cargo was not identified [30]. In hepatocellular carcinoma cells, exosomal miR-665 has been suggested to serve as a novel invasive biomarker [31]. A previously conducted mechanistic study showed that miR-665 directly suppressed tyrosine phosphatase receptor type B (PTPRB) expression as well as Hippo-mediated EMT in tumor cells [32]. In addition to exosomal microRNAs, chronic-myelogenous-leukemia-derived exosomal amphiregulin (AREG) activated Epidermal growth factor receptor (EGFR) and SNAIL expression [33]. AREG has been reported to be involved in the formation of a feedforward cycle of EGFR/TAZ/AREG and in the induction of EMT [34]. These results suggest that exosomal miRNA and proteins targeting the Hippo pathway could induce EMT in recipient cells.
2.2. β-Catenin Signaling Pathway
The Wnt/β-catenin signaling pathway is one of the most widely studied of the pathways that trigger EMT, and β-catenin is a key molecule involved in cancer-promoting pathways [35,36]. This signaling pathway also activates several EMT-related transcription factors, such as SNAIL, SLUG, and TWIST [37,38]. In studies about exosomes, Wnt/β-catenin signaling pathway is one of the most common target pathways.
Exosomal miR-301a is secreted by hypoxic glioblastoma (GBM) cells and targets transcription elongation factor A like 7 (TCEAL7), which, in turn, results in the activation of the Wnt/β-catenin signaling pathway [39]. In a previous study, miR-301a was found to induce EMT in prostate cancer cells through inhibition of p63, which caused the release of ZEB1/2 in a miRNA-mediated inhibition and induction EMT [40]. Exosomal miR-1260b reportedly activates the Wnt/β-catenin signaling pathway in lung adenocarcinoma cells [41]. miR-1260b was suggested to directly suppress suppressor of cytokine singling 6 (SOCS6) expression and activate KIT proto-oncogene (KIT) signaling in non-small-cell lung cancer (NSCLC) [42]. KIT signaling is known to induce EMT via the KIT-MEK-ERK pathways, supporting the significant roles of miR-1260b [43]. In another study, it was reported that miR-92a in fibroblast-derived exosomes activates the Wnt/β-catenin signaling pathway and induces EMT in colorectal cancer cells [44]. Additionally, miR-92a induced EMT by regulating the PI3K/AKT signaling pathway by targeting PTEN in NSCLC [45]. Similarly, exosomal delivery of miR-155 promoted EMT in breast cancer cells [46]. Although the underlying molecular mechanisms were not investigated in the study, miR-155 has been suggested to be oncogenic and EMT-inducible [47,48,49]. In the context of EMT, miR-155 activated the PI3K/SGK3/β-catenin signaling pathways through direct regulation of p85α expression [50,51]. Lastly, overexpressed PRMT5 circular RNA (circPRMT5), secreted via exosomes was found to promote EMT in recipient cells both in vivo and in vitro via sponging miR-30c [52,53]. Another similar study found that miR-30c targeted BCL9, a coactivator of the Wnt/β-catenin signaling pathway [54]. In a similar study, circABCC1 present within colorectal-cancer-cell-derived exosomes reportedly induced Wnt/β-catenin signaling activation in recipient cells, but the underlying molecular mechanisms were not investigated [55].
In studies about exosome-mediated Wnt/β-catenin signaling pathway, many Wnt ligands have been shown to be delivered via exosomes for biological function. Exosomal delivery of Wnt1 induced migration of colorectal cancer cells through activation of Wnt signaling, which was suppressed by inhibition of exosome production [56]. Wnt3a in fibroblast-derived exosomes induced activation of Wnt signaling and the development of drug resistance in colorectal cancer cells [57]. Wnt5a in exosomes from endothelial cells was biologically active and increased pericyte recruitment [58]. Wnt10b delivered by exosomes from fibroblasts induced EMT in breast cancer cells [59]. Fibroblast-derived exosomes promoted autocrine Wnt11a stimulation through formation of the PCP complex that is essential for exosome-mediated EMT induction in cancer cells [60,61]. Hypoxic-colorectal-cancer-cell-derived exosomal Wnt4 activated the β-catenin signaling pathway in endothelial cells, which contributed to colorectal cancer progression [62]. In large B-cell lymphoma, exosome-mediated Wnt3a activated the Wnt signaling pathway and promoted the progression of lymphoma cells [63]. Colorectal-cancer-cell-derived exosomes containing mutant β-catenin activated Wnt signaling in recipient cells in an autocrine manner [64]. Macrophage-derived exosomes containing Wnt5a activated the Wnt/β-catenin signaling pathway in breast cancer cells; this was further supported by the fact that Wnt5a increased exosome production in melanoma cells [65,66]. Therefore, regulation of Wnt ligands present in exosomes could serve as a promising therapeutic strategy to control EMT induction in cancer cells.
Significant roles of exosome-derived proteins have also been reported in the context of Wnt/β-catenin pathway activation. A recent study found that the EMT regulator SNAIL moves directly to the exosomes. Exosomes released by cancer-associated fibroblasts (CAFs) transferred SNAIL to lung cancer cells, and this SNAIL induced EMT in recipient lung cancer cells [67]. Deficiency of p53 specifically increased the expression of ubiquitin regulatory protein ligase E3 component N-recognin 2 (UBR2) in bone marrow mesenchymal stem cells as well as in its exosomes, activating Wnt signaling and downstream gene expression in gastric cancer cells [68]. Recently, a large extracellular matrix glycoprotein, Tenascin-C (TNC), was identified as an exosomal protein capable of inducing EMT. TNC is transferred by autocrinal exosomes from pancreatic ductal adenocarcinoma cells, and was found to promote EMT in the pancreatic ductal adenocarcinoma (PDAC) cell line via regulation of the Wnt/β-catenin pathway [69]. In breast cancer cells, TNC induces EMT by activation of SRC proto-oncogene tyrosine kinase (SRC) through phosphorylation at Y418 and through phosphorylation of focal adhesion kinase (FAK) at SRC substrate sites Y861 and Y925 [70].
Analysis of the expression of SRY-Box transcription factor 2 (SOX2) overlapping transcript (Sox2ot)—a long non-coding RNA (lncRNA)—derived from exosomes of highly invasive PDAC cells in plasma samples revealed that the plasma exosomal Sox2ot expression was high and correlated with TNM classification of malignant tumor stage and overall survival rate of PDAC patients. Further research showed that Sox2ot promotes EMT and stem-cell-like properties by regulating Sox2 expression. Sox2ot competitively binds to the miR-200 family to regulate the expression of Sox2, thus promoting invasion and metastasis of PDAC [71]. Previously, Sox2 inhibits Wnt/β-catenin signaling pathway through phosphorylation of β-catenin and glycogen synthase kinase 3 beta (GSK3β), leading to inhibition of the downstream gene expression [72]. The transfer and expression of long intergenic non-coding RNA regulator of reprogramming (linc-ROR) by CSC exosomes is responsible for the induction of EMT and the significant increase in the invasive potential of normal thyroid cells. Exosomal transfer of linc-ROR, a long non-coding RNA, was reported to induce EMT through the formation of a distant metastatic niche [73]. The significant role of linc-ROR through the Wnt/β-catenin pathway has been previously suggested in various types of cancers [74,75]. Furthermore, linc-ROR sponged miR-205 and increased the stability of the EMT inducer, ZEB2 [74]. These studies suggest that exosomal delivery of lncRNA may cause EMT induction.
2.3. ERK Pathway
miR-21 shows features of an oncogenic miRNA by targeting many tumor suppressor genes related to cell proliferation, apoptosis, and invasion in multiple histological cancer types, including OSCC. Hypoxia enhanced miR-21 levels in both OSCC cells and OSCC-derived exosomes. This induction of miR-21 by hypoxia was directly regulated by HIF-1a and HIF-2a in OSCC cells [76]. Additionally, inhibition of miR-21 reversed EMT and CSC phenotype, accompanied by phosphatase and tensin homolog (PTEN) upregulation and AKT/extracellular Signal Regulated Kinase (ERK)1/2 inactivation. Interestingly, downregulation of PTEN by siPTEN suppressed the effects of miR-21 antagomir on EMT and CSC phenotype, confirming that PTEN is a target of miR-21 during reversal of EMT and CSC phenotype in breast cancer cells [77].
Studies on liver cancer suggest that controlling exosome production and secretion could regulate exosomal EMT induction. Rab27a, a small GTPase, is required for efficient secretion of exosomes via mediation of multivesicular endosome docking at the plasma membrane [78]. In HLE (hepatoma cell line), the MAPK/ERK-mediated EMT was induced by MHCC97H hepatoma-cell-derived exosomes, and Rab27a knockdown reduced exosome secretion as well as EMT induction. Exosomes released by gastric cancer cells enhanced tumor proliferation by activating the MAPK/ERK pathway [79]. A similar result was obtained in a study that demonstrated that exosomes from hepatocellular carcinoma (HCC) cells increased ERK activity in normal hepatocytes [80].
An exosomal lncRNA, HOTAIR (Hox antisense intergenic RNA), was found to be highly expressed in bladder cancer patients and induced the invasiveness of the tumor through induction of EMT [81,82,83,84]. HOTAIR bound to miR-29b and rescued expression of DNMT3b, leading to PTEN expression, activation of the ERK and AKT signaling, and activation of EMT signaling pathway [85].
3. Therapeutic Candidates for Preventing Exosomal EMT Induction
Based on the roles of exosomes with respect to transferring bioactive molecules from donor cells to recipient cells, some investigators noticed that delivery of tumor-suppressive molecules via exosomes may regulate tumor progression. This was in accordance with some reports suggesting the significant involvement of exosomal delivery of nucleic acids in the inhibition of EMT in tumor cells, and their potential for therapeutic application. In this section, we summarize promising exosomal contents that prevent EMT in recipient tumor cells and their molecular relevance, based on previous studies.
In a study performed using cholangiocarcinoma cells, it was confirmed that exosome-encapsulated miR-30e delivery could suppress EMT in tumor cells through inhibition of SNAIL expression [86]. miR-30e has been widely studied and suggested to be a tumor suppressive miRNA that regulates tumor growth, invasion, and metastasis [87,88]. In the context of EMT induction, miR-30e is an important EMT inhibitor that works by targeting ITGB1, TUSC3, USP22, and SOX9 mRNAs, which reportedly induce EMT by activating transcription factors including SNAIL, SLUG, TWIST, and ZEB1/2 [89,90,91,92]. The preclinical significance of miR-30e treatment for EMT regulation was validated in tumor xenograft models, making exosomal miR-30e delivery a promising therapeutic strategy.
Recently, the EMT-suppressive effects of exosomal miR-26a were demonstrated in prostate cancer cells [93]. Although specific molecular events governed by miR-26 were not fully described, miR-26a was reportedly downregulated in tumor tissues and prevented EMT through direct inhibition of connective tissue growth factor (CTGF), rho-associated, coiled-coil-containing protein kinase 1 (ROCK1), enhancer of zeste homolog 2 (EZH2), and Jagged-1, which led to inactivation of the Jagged-1/Notch and TGFβ1/SMAD pathways [94,95,96,97,98]. Among the target genes, CTGF and ROCK1 indicated the involvement of miR-26a in YAP/TAZ signaling and further emphasized the importance of the miRNA [99].
miR-34a was supposed to be one of the foremost tumor-suppressive miRNAs, and the molecular mechanisms underlying its function were extensively studied [100]. A study on fibroblast-derived exosomes revealed that the level of miR-34a was significantly reduced in exosomes from CAFs, and that overexpression of miR-34a through exosomal transfer could suppress EMT in oral squamous cancer cells [101]. As miR-34a also reportedly targets signal transducer and activator of transcription 3 (STAT3), lymphoid Enhancer Binding Factor 1 (LEF1), ZEB1, and SNAIL, direct regulation of EMT could be achieved by treatment with miR-34a [102,103,104,105].
Exosomal miR-34c, a functionally similar family member of miR-34a, was reported to inhibit EMT by targeting β-catenin in nasopharyngeal cancer cells. Tumor-suppressive miR-34c was often downregulated in various types of tumor cells and regulated EMT through direct binding with SOX9, SATB2, MAP3K2, and DANCR mRNA [106,107,108,109,110]. The clinical prospects of miR-34c were substantiated by a study which showed that miR-34c inhibited exosome shedding by directly targeting RAB27B, thereby terminating the vicious cycle of oncogenic exosome-mediated intercellular communication [111].
A recent study about exosomes from bone-marrow-derived mesenchymal stem cells found that exosomal miR-101 inhibited EMT in oral cancer cells [112]. Although the molecular mechanisms were only elucidated to regulate COL10A1 expression, miR-101 reportedly played a role in EMT inhibition through direct inhibition of ZEB1/2, SOX9, TRIM44, ROCK2, and DUSP1, which supports its functional relevance [113,114,115,116,117].
Exosomes from miR-143 overexpressing bone-marrow-derived mesenchymal stem cells suppressed EMT in prostate cancer cells by directly targeting TFF3 mRNA [118]. Although not much is known about the role of TFF3 in EMT regulation, its significant correlation with TWIST1 has been reported in colorectal cancer [119]. The relationship between miR-143/TFF/EMT was also supported by previous studies which showed that miR-143 directly targets MYO6, HMB1, ERK5, Activin A, and LAMP3 mRNA, and eventually inactivates TWIST1, SNAIL, and SLUG [120,121,122,123,124].
Although the roles of miR-148b in regulation of tumor repression have been widely studied, only a few studies suggested ROCK1 and BRG1 as molecular targets related to EMT regulation [125,126]. Nevertheless, two studies have suggested that miR-148b in exosomes derived from mesenchymal stem cells and fibroblasts suppress EMT in recipient tumor cells [127,128]. Further studies are warranted to elucidate the molecular mechanisms underlying the function of exosomal miR-148b in EMT regulation.
In a previous study, exosomal miR-200 family reduced the invasiveness of colon cancer through inhibition of ZEB1 and SLUG [129]. With respect to the molecular mechanisms, the miR-200 family is known to be significantly involved in the induction of EMT via targeting of ZEB1, NANOG, and FOXF2 mRNA [130,131,132]. Among them, the effects of ZEB1 expression were clearly evident in a couple of studies. However, another study reported that extracellular vesicles containing miR-200 promoted cancer metastasis in xenograft mouse models [133]. As the EMT-suppressive roles of miR-200 have been studied widely, further studies could clarify the precise roles of the exosomal miR-200 in EMT regulation.
The application of EMT-suppressive miR-375 was ascertained in a study that found that treatment with Emblica officinalis extracts causes exosomal miR-375 to increasingly target SNAIL and inhibit invasiveness of ovarian cancer [134]. Similarly, exosomal miR-375 suppressed EMT in glioblastoma cells through direct targeting of SLC31A1 mRNA [135]. miR-375 is often downregulated in various types of cancers and shows tumor-suppressive effects by directly targeting various oncogenes at the mRNA level [136]. With respect to EMT induction, the regulation of YAP1 expression by direct binding of miR-375 was controlled by ZEB1 in prostate cancer, thereby forming a feedforward cycle [137]. Additionally, miR-375 inhibited the expression of 14-3-3ζ, which led to inhibition of the Wnt/β-catenin signaling pathway and EMT in gastric cancer [138]. These studies demonstrated the significant involvement of miR-375 in regulation of EMT in cancer cells.
These studies cumulatively proved that miRNAs previously known to regulate EMT also showed the same effects upon exosomal delivery, the exosomes being obtained from mesenchymal stem cells with transfection of the desired miRNAs. It can be theorized that a universal recipe for exosomal encapsulation of miRNAs may be developed, wherein these constructs serve as adjuvants for anti-cancer therapy based on these investigations. Although many studies have established the preclinical significance of co-treatment of miRNAs with anti-cancer therapies including chemotherapy and radiotherapy, the therapeutic effects of combining multiple miRNAs have not been widely investigated. For better therapeutic significance, a combination of several miRNAs mentioned above could be formulated to achieve synergistic effects on the regulation of tumor EMT and progression in future studies.
4. Clinical Application
To date, many studies have been conducted to understand the biological functions of exosomes, and some have identified clinical utility. Most clinical studies have used exosomes as biomarkers for diagnosis or prognosis of disorders including cancers. In the same context, exosomes containing EMT inducers have been utilized for prediction of metastatic prognosis of cancers in clinical trials. For example, profiling exosomal RNA in primary high-grade osteosarcoma metastasizing to the lung could provide diagnostic and prognostic biomarkers [139] (NCT03108677). In addition, one investigation analyzed circulating exosomes and miRNAs in cancer patients to predict the possibility of bone metastasis [140] (NCT03895216). Additionally, exosomes were recently utilized as a treatment. A vaccination of tumor antigen was mediated by dendritic-cell-derived exosomes [141] (NCT01159288), and a transfer of curcumin to normal and cancerous colon cells was mediated by plant exosomes [142] (NCT01294072). In clinical trials with pancreatic cancer patients, attempts were made to target the mutant form of the GTPase KRAS (KRASG12D) by transfer of siRNA mediated by mesenchymal-stromal-cell-derived exosomes (iExosomes) [143,144] (NCT03608631). Although there have not been many clinical investigations performed to date, we present in this section studies that confirmed therapeutic effects and show a marked potential for clinical applications.
As described above, miRNAs and proteins delivered via exosomes have exhibited potential as EMT regulators in a variety of cancers. As most studies were performed using cell models, data regarding studies performed on in vivo models are insufficient. There are two representative types of therapeutic strategy for inhibiting exosome-induced EMT. One is the transfer of tumor-suppressive factors through exosomes, and the other is the inhibition of EV production. This section discusses previous studies covering the in vivo effects of exosomes in regulating EMT in tumors.
There have been many attempts to incorporate various compounds into exosome therapy. Exosomes from the plasma of head and neck squamous cell carcinoma (HNSCC) patients treated with photodynamic therapy (PDT) downregulated the expression of SLUG and ZEB1 [145]. Studies on cancer EMT revealed that transcriptional activation of ZEB1 by SLUG promoted EMT progression that decreased adhesion and increased migration of cancer cells [146]. Exosomes from patient plasma could be used for management of EMT as part of a novel therapeutic approach. In a previous study, exosomes packed with an miR-21-sponge were successfully employed in a GBM rat model, suggesting that this construct could have EMT-inhibitory effects [147]. miR-21 targets several EMT pathway-related genes, such as PDCD4, TIMP3, and RECK [148]. This strategy of inhibition using exosomes containing an miR-21-sponge construct has therapeutic potential for blocking GBM and GBM-induced EMT. Another study suggested that suppressing miR-21 also enhanced the therapeutic effects of sunitinib-based chemotherapy [149].
Another approach is therapy based on the use of exosomes that repress tumor metastasis by targeting EMT inducers. The findings of studies on this therapy have been difficult to apply in clinical trials, but these studies might contribute toward developing a therapeutic strategy against cancer-induced EMT.
In a study on mouse- or rat-derived exosomes, artificial exosomes loaded with miRNAs targeting Claudin 7 and EpCAM through electroporation were intravenously injected, resulting in significant inhibition of cancer EMT and metastasis [150]. Claudin 7 promotes EMT in human colorectal cancer and is required for EpCAM expression [151]. For therapeutic applications, the use of exosomes has beneficial effects in the management of EMT. In a recent study, EVs obtained from bone marrow mesenchymal stem cells overexpressing miR-200b prevented EMT by targeting ZEB1 and ZEB2 mRNA. Although the therapeutic target was not cancer cells, the result suggested a significant role of miR-200b in clinical applications for regulation of EMT [152].
Another study demonstrated that the tumor suppressive miR-335 could be produced in B cells by plasmid DNA induction (iEVs) which is suggested a new therapeutic method utilizing exosomes and showed anticancer effects. miR-335 has been reported to inhibit tumor re-initiation, and is known to be widely silenced in genetic and epigenetic manners during tumorigenesis. Induced exosomal miR-355 can overcome this limitation. One of the targets of miR-335 is SOX4, a transcription factor involved in embryonic development, cell fate determination, and EMT [153].
Although there have been few clinical attempts to inhibit exosome production in order to regulate the cancer EMT, there has been a report supporting its anti-tumorigenic roles. A study prevented EV production using cetuximab, a monoclonal antibody targeting EGFR, and was successful in reducing the production of EMT-inducing exosomes from oral cancer cells [154].
In addition to the two major therapeutic strategies described above, some studies have suggested new approaches that need to be improved for their clinical application. One group posited exosomes or exosome-like platforms for anticancer drug delivery. Exosomes derived from brain endothelial cells overexpressing CD63 tetraspanin transmembrane proteins successfully penetrated the blood-brain barrier (BBB) and delivered containing anticancer drugs in the zebrafish brain cancer model. Exosome-delivered drugs significantly decreased the growth of xenografted cancer cells [155]. Another group devised exosome-mimetic nanosystems (EMNs) that simulated natural tumor-derived exosomes with respect to their structure and functionality, but with a controlled composition, for targeted delivery of therapeutic oligonucleotides to lung adenocarcinoma cells. These EMNs delivered miRNA-145 to lung adenocarcinoma cells [156]. These studies offer new strategies for the delivery of anti-cancer, anti-EMT drugs, and therapeutic oligonucleotides via artificially exosomes or exosome mimics. Further research could open avenues to new anticancer therapies by delivering drugs and miRNAs that are effective against a variety of cancers or a variety of tumorigenic processes, including EMT.
5. Conclusions
In this review, we summarized the major exosomal molecules that induce EMT and their underlying molecular mechanisms (Figure 3, Table 1). Previously known EMT-related signaling pathways are involved in exosome-induced EMT, miRNAs being the important mediators. Furthermore, we reviewed studies on therapeutic candidates that can be delivered via exosomes and their clinical applications as anti-cancer therapies. Although there is more to be discovered about the roles of exosomes, it is undeniable that exosomes play a critical role in the regulation of EMT in recipient tumor cells. This research review should be helpful in providing a deeper understanding of exosome-induced EMT, and could also guide researchers pursuing novel investigations in this area.
Author Contributions
Conceptualization, S.L., H.K. and B.Y.; writing—original draft preparation, S.L., H.K., E.S., K.M.S., Y.W.J., H.Y. and B.Y.; writing—review and editing, S.L., H.K., E.S., K.M.S., Y.W.J., H.Y. and B.Y.; supervision, B.Y.; project administration, B.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by a grant of the Korea Institute of Radiological and Medical Sciences (KIRAMS), funded by Nuclear Safety and Security Commission (NSSC) of the Republic of Korea (No. 50091-2020), Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2017R1D1A1B03028769), the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (NRF-2019R1A2C1008051 to H Youn), and PNU-RENovation (2018–2019 to H Kim).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ABCC1 | ATP binding cassette subfamily C member 1 |
| AKT | protein kinase B |
| AREG | amphiregulin |
| BCL9 | B-cell CLL/lymphoma 9 protein |
| BRG1 | brahma-related gene 1 |
| CAF | cancer associated fibroblast |
| COL10A1 | collagen type X alpha 1 chain |
| CSC | cancer stem cell |
| CTGF | connective tissue growth factor |
| DANCR | differentiation antagonizing non-protein coding RNA |
| DUSP1 | dual specificity protein phosphatase 1 |
| EGF | epidermal growth factor |
| EGFR | epidermal growth factor receptor |
| EMN | exosome-mimetic nanosystem |
| EMT | epithelial-mesenchymal-transition |
| EpCAM | epithelial cell adhesion molecule |
| ERK | extracellular signal regulated kinase |
| EV | extracellular vesicle |
| EZH2 | enhancer of zeste homolog 2 |
| FAK | focal adhesion kinase |
| FBXW7 | F-Box and WD repeat domain containing 7 |
| FGF | fibroblast growth factor |
| FOXF2 | forkhead box F2 |
| FOXO3a | forkhead box O3 |
| GBM | glioblastoma multiform |
| HCC | hepatocellular carcinoma |
| HGF | hepatocyte growth factor |
| HIF-1α | hypoxia-inducible factor 1-alpha |
| HLE | human lens epithelial cell |
| HMB1 | high mobility group box 1 |
| HNSCC | head and neck squamous cell carcinoma |
| ITGB1 | integrin subunit beta 1 |
| KIT | KIT proto-oncogene |
| LAMP3 | lysosome-associated membrane protein 3 |
| LEF1 | lymphoid enhancer-binding factor 1 |
| MAP3K2 | mitogen-activated protein kinase kinase kinase 2 |
| MAPK | mitogen-activated protein kinase |
| MEK | MAP kinase-ERK kinase |
| MOAP1 | modulator of apoptosis 1 |
| MYO6 | myosine 6 |
| NSCLC | non-small-cell lung carcinoma |
| OSCC | oral squamous cell carcinoma |
| PCP | planar cell polarity |
| PDAC | pancreatic ductal adenocarcinoma |
| PDCD4 | programmed cell death protein 4 |
| PDT | photodynamic therapy |
| PI3K | phosphoinositide 3-kinase |
| PRMT5 | protein arginine methyltransferase 5 |
| PTEN | phosphatase and tensin homolog |
| PTPRB | protein tyrosine phosphatase receptor type B |
| RAB27B | RAB27B, member RAS oncogene family |
| RECK | reversion-inducing-cysteine-rich protein with kazal motifs |
| ROCK1 | rho-associated, coiled-coil-containing protein kinase 1 |
| ROR | RNA regulator of reprogramming |
| SATB2 | special AT-rich sequence-binding protein 2 |
| SGK3 | serum and glucocorticoid kinase 3 |
| SLAI1 | snail family transcriptional repressor 1 |
| SLC31A1 | solute carrier family 31 member 1 |
| SOCS6 | suppressor of cytokine signaling 6 |
| Sox2ot | SOX2 overlapping transcript |
| SOX9 | SRY-Box transcription factor 9 |
| SRC | SRC proto-oncogene tyrosine kinase |
| STAT3 | signal transducer and activator of transcription 3 |
| TAZ | transcriptional coactivator with PDZ-binding motif |
| TCEAL7 | transcription elongation factor A like 7 |
| TEAD | transcriptional enhanced associate domain |
| TFF3 | trefoil factor 3 |
| TGF-β | transforming growth factor β |
| TIMP3 | metalloproteinase inhibitor 3 |
| TNC | tenascin C |
| TRIM44 | tripartite motif-containing protein 44 |
| TUSC3 | tumor suppressor candidate 3 |
| UBR2 | ubiquitin regulatory protein ligase E3 component n-recognin 2 |
| USP22 | ubiquitin specific peptidase 22 |
| YAP | yes-associated protein |
| YY1 | YY1 transcription factor |
| ZEB1/2 | zinc finger E-box binding homeobox 1 |
References
- Pastushenko, I.; Blanpain, C. EMT Transition States during Tumor Progression and Metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Weinberg, R.A. Epithelial-to-mesenchymal transition in cancer: Complexity and opportunities. Front. Med. 2018, 12, 361–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Li, B.; Liu, F.; Zhang, M.; Wang, Q.; Liu, Y.; Yao, Y.; Li, D. The epithelial to mesenchymal transition (EMT) and cancer stem cells: Implication for treatment resistance in pancreatic cancer. Mol. Cancer 2017, 16, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Settleman, J. EMT, cancer stem cells and drug resistance: An emerging axis of evil in the war on cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azmi, A.S. Unveiling the role of nuclear transport in epithelial-to-mesenchymal transition. Curr. Cancer Drug Targets 2013, 13, 906–914. [Google Scholar] [CrossRef]
- Xu, R.; Won, J.Y.; Kim, C.H.; Kim, D.E.; Yim, H. Roles of the Phosphorylation of Transcriptional Factors in Epithelial-Mesenchymal Transition. J. Oncol. 2019, 2019, 5810465. [Google Scholar] [CrossRef]
- Dave, N.; Guaita-Esteruelas, S.; Gutarra, S.; Frias, À.; Beltran, M.; Peiró, S.; de Herreros, A.G. Functional cooperation between Snail1 and twist in the regulation of ZEB1 expression during epithelial to mesenchymal transition. J. Biol. Chem. 2011, 286, 12024–12032. [Google Scholar] [CrossRef] [Green Version]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, Zeb and bHLH factors in tumour progression: An alliance against the epithelial phenotype? Nat. Rev. Cancer 2007, 7, 415–428. [Google Scholar] [CrossRef]
- Blackwell, R.H.; Foreman, K.E.; Gupta, G.N. The Role of Cancer-Derived Exosomes in Tumorigenicity & Epithelial-to-Mesenchymal Transition. Cancers 2017, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.M.; Molaei, F.; Alahari, S.K. Exosomes: Composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol. Cancer 2019, 18, 75. [Google Scholar] [CrossRef]
- Huang, T.; Song, C.; Zheng, L.; Xia, L.; Li, Y.; Zhou, Y. The roles of extracellular vesicles in gastric cancer development, microenvironment, anti-cancer drug resistance, and therapy. Mol. Cancer 2019, 18, 62. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and exosomal microRNA: Trafficking, sorting, and function. Genom. Proteom. Bioinf. 2015, 13, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruenberg, J.; van der Goot, F.G. Mechanisms of pathogen entry through the endosomal compartments. Nat. Rev. Mol. Cell Biol. 2006, 7, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Maia, J.; Caja, S.; Strano Moraes, M.C.; Couto, N.; Costa-Silva, B. Exosome-Based Cell-Cell Communication in the Tumor Microenvironment. Front. Cell Dev. Biol. 2018, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Biophys. Acta 2012, 1820, 940–948. [Google Scholar] [CrossRef]
- Yoshioka, Y.; Konishi, Y.; Kosaka, N.; Katsuda, T.; Kato, T.; Ochiya, T. Comparative marker analysis of extracellular vesicles in different human cancer types. J. Extracell. Vesicles 2013, 2, 20424. [Google Scholar] [CrossRef]
- Yu, F.-X.; Zhao, B.; Guan, K.-L. Hippo Pathway in Organ Size Control, Tissue Homeostasis, and Cancer. Cell 2015, 163, 811–828. [Google Scholar] [CrossRef] [Green Version]
- Camargo, F.D.; Gokhale, S.; Johnnidis, J.B.; Fu, D.; Bell, G.W.; Jaenisch, R.; Brummelkamp, T.R. YAP1 increases organ size and expands undifferentiated progenitor cells. Curr. Biol. 2007, 17, 2054–2060. [Google Scholar] [CrossRef] [Green Version]
- Boopathy, G.T.K.; Hong, W. Role of Hippo Pathway-YAP/TAZ Signaling in Angiogenesis. Front. Cell Dev. Biol. 2019, 7, 49. [Google Scholar] [CrossRef]
- Liu, H.; Du, S.; Lei, T.; Wang, H.; He, X.; Tong, R.; Wang, Y. Multifaceted regulation and functions of YAP/TAZ in tumors (Review). Oncol. Rep. 2018, 40, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Huh, H.D.; Kim, D.H.; Jeong, H.-S.; Park, H.W. Regulation of TEAD Transcription Factors in Cancer Biology. Cells 2019, 8, 600. [Google Scholar] [CrossRef] [Green Version]
- Adhikary, A.; Chakraborty, S.; Mazumdar, M.; Ghosh, S.; Mukherjee, S.; Manna, A.; Mohanty, S.; Nakka, K.K.; Joshi, S.; De, A.; et al. Inhibition of epithelial to mesenchymal transition by E-cadherin up-regulation via repression of slug transcription and inhibition of E-cadherin degradation: Dual role of scaffold/matrix attachment region-binding protein 1 (SMAR1) in breast cancer cells. J. Biol. Chem. 2014, 289, 25431–25444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.T.; Zhong, H.T.; Li, G.W.; Shen, J.X.; Ye, Q.Q.; Zhang, M.L.; Liu, J. Oncogenic functions of the EMT-related transcription factor ZEB1 in breast cancer. J. Transl. Med. 2020, 18, 51. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, J.; Ying, X.; Lin, P.C.; Zhou, B.P. Twist-mediated Epithelial-mesenchymal Transition Promotes Breast Tumor Cell Invasion via Inhibition of Hippo Pathway. Sci. Rep. 2016, 6, 24606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, C.Y.; Chai, J.Y.; Tang, T.F.; Wong, W.F.; Sethi, G.; Shanmugam, M.K.; Chong, P.P.; Looi, C.Y. The E-Cadherin and N-Cadherin Switch in Epithelial-to-Mesenchymal Transition: Signaling, Therapeutic Implications, and Challenges. Cells 2019, 8, 1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Shin, J.E.; Park, H.W. The Role of Hippo Pathway in Cancer Stem Cell Biology. Mol. Cells 2018, 41, 83–92. [Google Scholar] [CrossRef]
- Noguchi, S.; Saito, A.; Nagase, T. YAP/TAZ Signaling as a Molecular Link between Fibrosis and Cancer. Int. J. Mol. Sci. 2018, 19, 3674. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, W.; Mossmann, D.; Kleemann, J.; Mock, K.; Meisinger, C.; Brummer, T.; Herr, R.; Brabletz, S.; Stemmler, M.P.; Brabletz, T. ZEB1 turns into a transcriptional activator by interacting with YAP1 in aggressive cancer types. Nat. Commun. 2016, 7, 10498. [Google Scholar] [CrossRef]
- Wang, S.; Su, X.; Xu, M.; Xiao, X.; Li, X.; Li, H.; Keating, A.; Zhao, R.C. Exosomes secreted by mesenchymal stromal/stem cell-derived adipocytes promote breast cancer cell growth via activation of Hippo signaling pathway. Stem Cell Res. Ther. 2019, 10, 117. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.; Wu, J.; Wu, J.; Ji, A.; Qiang, G.; Jiang, Y.; Jiang, C.; Ding, Y. Exosomal miR-665 as a novel minimally invasive biomarker for hepatocellular carcinoma diagnosis and prognosis. Oncotarget 2017, 8, 80666–80678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Yang, C.; Yang, S.; Cheng, F.; Rao, J.; Wang, X. miR-665 promotes hepatocellular carcinoma cell migration, invasion, and proliferation by decreasing Hippo signaling through targeting PTPRB. Cell Death Dis. 2018, 9, 954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrado, C.; Saieva, L.; Raimondo, S.; Santoro, A.; De Leo, G.; Alessandro, R. Chronic myelogenous leukaemia exosomes modulate bone marrow microenvironment through activation of epidermal growth factor receptor. J. Cell Mol. Med. 2016, 20, 1829–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N.; Morrison, C.D.; Liu, P.; Miecznikowski, J.; Bshara, W.; Han, S.; Zhu, Q.; Omilian, A.R.; Li, X.; Zhang, J. TAZ induces growth factor-independent proliferation through activation of EGFR ligand amphiregulin. Cell Cycle 2012, 11, 2922–2930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Yun, F.; Shi, L.; Li, Z.-H.; Luo, N.-R.; Jia, Y.-F. Roles of Signaling Pathways in the Epithelial-Mesenchymal Transition in Cancer. Asian Pac. J. Cancer Prev. 2015, 16, 6201–6206. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.-Y.; Zhu, M.-X.; Yang, Y.-W.; Zhang, P.-F.; Yang, X.; Peng, R.; Gao, C.; Lu, J.-C.; Wang, L.; Deng, X.-Y.; et al. Downregulation of RNF128 activates Wnt/β-catenin signaling to induce cellular EMT and stemness via CD44 and CTTN ubiquitination in melanoma. J. Hematol. Oncol. 2019, 12, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Li, S.-N.; Anjum, K.M.; Gui, L.-X.; Zhu, S.-S.; Liu, J.; Chen, J.-K.; Liu, Q.-F.; Ye, G.-D.; Wang, W.-J.; et al. A double-negative feedback loop between Wnt-β-catenin signaling and HNF4α regulates epithelial-mesenchymal transition in hepatocellular carcinoma. J. Cell Sci. 2013, 126, 5692–5703. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Weinberg, R.A. Epithelial-mesenchymal transition: At the crossroads of development and tumor metastasis. Dev. Cell 2008, 14, 818–829. [Google Scholar] [CrossRef] [Green Version]
- Yue, X.; Lan, F.; Xia, T. Hypoxic Glioma Cell-Secreted Exosomal miR-301a Activates Wnt/β-catenin Signaling and Promotes Radiation Resistance by Targeting TCEAL7. Mol. Ther. 2019, 27, 1939–1949. [Google Scholar] [CrossRef]
- Nam, R.K.; Benatar, T.; Wallis, C.J.D.; Amemiya, Y.; Yang, W.; Garbens, A.; Naeim, M.; Sherman, C.; Sugar, L.; Seth, A. MiR-301a regulates E-cadherin expression and is predictive of prostate cancer recurrence. Prostate 2016, 76, 869–884. [Google Scholar] [CrossRef]
- Xia, Y.; Wei, K.; Hu, L.-Q.; Zhou, C.-R.; Lu, Z.-B.; Zhan, G.-S.; Pan, X.-L.; Pan, C.-F.; Wang, J.; Wen, W.; et al. Exosome-mediated transfer of miR-1260b promotes cell invasion through Wnt/β-catenin signaling pathway in lung adenocarcinoma. J. Cell Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wei, K.; Yang, F.-M.; Hu, L.-Q.; Pan, C.-F.; Pan, X.-L.; Wu, W.-B.; Wang, J.; Wen, W.; He, Z.-C.; et al. miR-1260b, mediated by YY1, activates KIT signaling by targeting SOCS6 to regulate cell proliferation and apoptosis in NSCLC. Cell Death Dis. 2019, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-L.; Fan, Y.-L.; Jiang, J.; Li, K.-D.; Zheng, M.; Chen, W.; Ma, X.-R.; Geng, N.; Chen, Q.-M.; Chen, Y.; et al. C-kit induces epithelial-mesenchymal transition and contributes to salivary adenoid cystic cancer progression. Oncotarget 2014, 5, 1491–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.L.; Wang, W.; Lan, X.L.; Zeng, Z.C.; Liang, Y.S.; Yan, Y.R.; Song, F.Y.; Wang, F.F.; Zhu, X.H.; Liao, W.J.; et al. CAFs secreted exosomes promote metastasis and chemotherapy resistance by enhancing cell stemness and epithelial-mesenchymal transition in colorectal cancer. Mol. Cancer 2019, 18, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Shan, Z.; Hong, J.; Yang, L. MicroRNA-92a promotes epithelial-mesenchymal transition through activation of PTEN/PI3K/AKT signaling pathway in non-small cell lung cancer metastasis. Int. J. Oncol. 2017, 51, 235–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, J.C.; Lima, N.d.S.; Sarian, L.O.; Matheu, A.; Ribeiro, M.L.; Derchain, S.F.M. Exosome-mediated breast cancer chemoresistance via miR-155 transfer. Sci. Rep. 2018, 8, 829. [Google Scholar] [CrossRef] [Green Version]
- Shen, R.; Wang, Y.; Wang, C.-X.; Yin, M.; Liu, H.-L.; Chen, J.-P.; Han, J.-Q.; Wang, W.-B. MiRNA-155 mediates TAM resistance by modulating SOCS6-STAT3 signalling pathway in breast cancer. Am. J. Transl. Res. 2015, 7, 2115–2126. [Google Scholar]
- Ouyang, M.; Li, Y.; Ye, S.; Ma, J.; Lu, L.; Lv, W.; Chang, G.; Li, X.; Li, Q.; Wang, S.; et al. MicroRNA profiling implies new markers of chemoresistance of triple-negative breast cancer. PLoS ONE 2014, 9, e96228. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Kong, X.; Lv, L.; Gao, J. TGF-β1 acts through miR-155 to down-regulate TP53INP1 in promoting epithelial-mesenchymal transition and cancer stem cell phenotypes. Cancer Lett. 2015, 359, 288–298. [Google Scholar] [CrossRef]
- Kong, X.; Liu, F.; Gao, J. MiR-155 promotes epithelial-mesenchymal transition in hepatocellular carcinoma cells through the activation of PI3K/SGK3/β-catenin signaling pathways. Oncotarget 2016, 7, 66051–66060. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, E.; Jung, J.; Lee, J.W.; Kim, H.J.; Kim, J.; Yoo, H.J.; Lee, H.J.; Chae, S.Y.; Jeon, S.M.; et al. microRNA-155 positively regulates glucose metabolism via PIK3R1-FOXO3a-cMYC axis in breast cancer. Oncogene 2018, 37, 2982–2991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, J.; Ma, J.; Sun, T.; Zhou, Q.; Wang, W.; Wang, G.; Wu, P.; Wang, H.; Jiang, L.; et al. Exosomal circRNAs: Biogenesis, effect and application in human diseases. Mol. Cancer 2019, 18, 116. [Google Scholar] [CrossRef] [PubMed]
- Su, L.-L.; Chang, X.-J.; Zhou, H.-D.; Hou, L.-B.; Xue, X.-Y. Exosomes in esophageal cancer: A review on tumorigenesis, diagnosis and therapeutic potential. World J. Clin. Cases 2019, 7, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Ling, X.-H.; Chen, Z.-Y.; Luo, H.-W.; Liu, Z.-Z.; Liang, Y.-K.; Chen, G.-X.; Jiang, F.-N.; Zhong, W.-D.E. BCL9, a coactivator for Wnt/β-catenin transcription, is targeted by miR-30c and is associated with prostate cancer progression. Oncol. Lett. 2016, 11, 2001–2008. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Chen, S.; Fu, Q. Exosomes from CD133(+) cells carrying circ-ABCC1 mediate cell stemness and metastasis in colorectal cancer. J. Cell Biochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.-W.; Cao, C.-H.; Han, K.; Zhao, Y.-X.; Cai, M.-Y.; Xiang, Z.-C.; Zhang, J.-X.; Chen, J.-W.; Zhong, L.-P.; Huang, Y.; et al. APC-activated long noncoding RNA inhibits colorectal carcinoma pathogenesis through reduction of exosome production. J. Clin. Investig. 2019, 129, 727–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.B.; Yan, C.; Mu, L.; Mi, Y.L.; Zhao, H.; Hu, H.; Li, X.L.; Tao, D.D.; Wu, Y.Q.; Gong, J.P.; et al. Exosomal Wnt-induced dedifferentiation of colorectal cancer cells contributes to chemotherapy resistance. Oncogene 2019, 38, 1951–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, K.; Shamskhou, E.A.; Orcholski, M.E.; Nathan, A.; Reddy, S.; Honda, H.; Mani, V.; Zeng, Y.; Ozen, M.O.; Wang, L.; et al. Loss of Endothelium-Derived Wnt5a Is Associated With Reduced Pericyte Recruitment and Small Vessel Loss in Pulmonary Arterial Hypertension. Circulation 2019, 139, 1710–1724. [Google Scholar] [CrossRef]
- Chen, Y.; Zeng, C.; Zhan, Y.; Wang, H.; Jiang, X.; Li, W. Aberrant low expression of p85α in stromal fibroblasts promotes breast cancer cell metastasis through exosome-mediated paracrine Wnt10b. Oncogene 2017, 36, 4692–4705. [Google Scholar] [CrossRef] [Green Version]
- Luga, V.; Zhang, L.; Viloria-Petit, A.M.; Ogunjimi, A.A.; Inanlou, M.R.; Chiu, E.; Buchanan, M.; Hosein, A.N.; Basik, M.; Wrana, J.L. Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell 2012, 151, 1542–1556. [Google Scholar] [CrossRef] [Green Version]
- Luga, V.; Wrana, J.L. Tumor-stroma interaction: Revealing fibroblast-secreted exosomes as potent regulators of Wnt-planar cell polarity signaling in cancer metastasis. Cancer Res. 2013, 73, 6843–6847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Feng, Y. Exosomes Derived From Hypoxic Colorectal Cancer Cells Promote Angiogenesis Through Wnt4-Induced β-Catenin Signaling in Endothelial Cells. Oncol. Res. 2017, 25, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Koch, R.; Demant, M.; Aung, T.; Diering, N.; Cicholas, A.; Chapuy, B.; Wenzel, D.; Lahmann, M.; Güntsch, A.; Kiecke, C.; et al. Populational equilibrium through exosome-mediated Wnt signaling in tumor progression of diffuse large B-cell lymphoma. Blood 2014, 123, 2189–2198. [Google Scholar] [CrossRef] [PubMed]
- Kalra, H.; Gangoda, L.; Fonseka, P.; Chitti, S.V.; Liem, M.; Keerthikumar, S.; Samuel, M.; Boukouris, S.; Al Saffar, H.; Collins, C.; et al. Extracellular vesicles containing oncogenic mutant β-catenin activate Wnt signalling pathway in the recipient cells. J. Extracell. Vesicles 2019, 8, 1690217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menck, K.; Klemm, F.; Gross, J.C.; Pukrop, T.; Wenzel, D.; Binder, C. Induction and transport of Wnt 5a during macrophage-induced malignant invasion is mediated by two types of extracellular vesicles. Oncotarget 2013, 4, 2057–2066. [Google Scholar] [CrossRef] [Green Version]
- Ekström, E.J.; Bergenfelz, C.; von Bülow, V.; Serifler, F.; Carlemalm, E.; Jönsson, G.; Andersson, T.; Leandersson, K. WNT5A induces release of exosomes containing pro-angiogenic and immunosuppressive factors from malignant melanoma cells. Mol. Cancer 2014, 13, 88. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Li, M.; Cao, L.M.; Gu, Q.H.; Deng, P.B.; Tan, Y.; Hu, C.P. Snail1-dependent cancer-associated fibroblasts induce epithelial-mesenchymal transition in lung cancer cells via exosomes. QJM 2019, 112, 581–590. [Google Scholar] [CrossRef]
- Mao, J.; Liang, Z.; Zhang, B.; Yang, H.; Li, X.; Fu, H.; Zhang, X.; Yan, Y.; Xu, W.; Qian, H. UBR2 Enriched in p53 Deficient Mouse Bone Marrow Mesenchymal Stem Cell-Exosome Promoted Gastric Cancer Progression via Wnt/β-Catenin Pathway. Stem Cells 2017, 35, 2267–2279. [Google Scholar] [CrossRef] [Green Version]
- Qian, S.; Tan, X.; Liu, X.; Liu, P.; Wu, Y. Exosomal Tenascin-c induces proliferation and invasion of pancreatic cancer cells by WNT signaling. Onco Targets Ther. 2019, 12, 3197–3205. [Google Scholar] [CrossRef] [Green Version]
- Nagaharu, K.; Zhang, X.; Yoshida, T.; Katoh, D.; Hanamura, N.; Kozuka, Y.; Ogawa, T.; Shiraishi, T.; Imanaka-Yoshida, K. Tenascin C induces epithelial-mesenchymal transition-like change accompanied by SRC activation and focal adhesion kinase phosphorylation in human breast cancer cells. Am. J. Pathol. 2011, 178, 754–763. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Jiang, P.; Li, J.; Peng, M.; Zhao, X.; Zhang, X.; Chen, K.; Zhang, Y.; Liu, H.; Gan, L.; et al. Tumor-derived exosomal lnc-Sox2ot promotes EMT and stemness by acting as a ceRNA in pancreatic ductal adenocarcinoma. Oncogene 2018, 37, 3822–3838. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Shi, J.; Zhang, K.; Xue, J.; Li, J.; Yang, J.; Chen, J.; Wei, J.; Ren, H.; Liu, X. Sox2 inhibits Wnt-β-catenin signaling and metastatic potency of cisplatin-resistant lung adenocarcinoma cells. Mol. Med. Rep. 2017, 15, 1693–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardin, H.; Helein, H.; Meyer, K.; Robertson, S.; Zhang, R.; Zhong, W.; Lloyd, R.V. Thyroid cancer stem-like cell exosomes: Regulation of EMT via transfer of lncRNAs. Lab. Investig. 2018, 98, 1133–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, P.; Zhao, Y.; Li, Z.; Yao, R.; Ma, M.; Gao, Y.; Zhao, L.; Zhang, Y.; Huang, B.; Lu, J. LincRNA-ROR induces epithelial-to-mesenchymal transition and contributes to breast cancer tumorigenesis and metastasis. Cell Death Dis. 2014, 5, e1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.; Jiang, H.; Cui, Z.; Wang, L.; Wang, X.; Tian, T. Linc-ROR induces epithelial-to-mesenchymal transition in ovarian cancer by increasing Wnt/β-catenin signaling. Oncotarget 2017, 8, 69983–69994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Li, C.; Wang, S.; Wang, Z.; Jiang, J.; Wang, W.; Li, X.; Chen, J.; Liu, K.; Li, C.; et al. Exosomes Derived from Hypoxic Oral Squamous Cell Carcinoma Cells Deliver miR-21 to Normoxic Cells to Elicit a Prometastatic Phenotype. Cancer Res. 2016, 76, 1770–1780. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Liu, M.; Wang, Y.; Chen, X.; Xu, J.; Sun, Y.; Zhao, L.; Qu, H.; Fan, Y.; Wu, C. Antagonism of miR-21 reverses epithelial-mesenchymal transition and cancer stem cell phenotype through AKT/ERK1/2 inactivation by targeting PTEN. PLoS ONE 2012, 7, e39520. [Google Scholar] [CrossRef] [Green Version]
- Zerial, M.; McBride, H. Rab proteins as membrane organizers. Nat. Rev. Mol. Cell Biol. 2001, 2, 107–117. [Google Scholar] [CrossRef]
- Qu, J.L.; Qu, X.J.; Zhao, M.F.; Teng, Y.E.; Zhang, Y.; Hou, K.Z.; Jiang, Y.H.; Yang, X.H.; Liu, Y.P. Gastric cancer exosomes promote tumour cell proliferation through PI3K/Akt and MAPK/ERK activation. Dig. Liver Dis. 2009, 41, 875–880. [Google Scholar] [CrossRef]
- He, M.; Qin, H.; Poon, T.C.W.; Sze, S.-C.; Ding, X.; Co, N.N.; Ngai, S.-M.; Chan, T.-F.; Wong, N. Hepatocellular carcinoma-derived exosomes promote motility of immortalized hepatocyte through transfer of oncogenic proteins and RNAs. Carcinogenesis 2015, 36, 1008–1018. [Google Scholar] [CrossRef] [Green Version]
- Berrondo, C.; Flax, J.; Kucherov, V.; Siebert, A.; Osinski, T.; Rosenberg, A.; Fucile, C.; Richheimer, S.; Beckham, C.J. Expression of the Long Non-Coding RNA HOTAIR Correlates with Disease Progression in Bladder Cancer and Is Contained in Bladder Cancer Patient Urinary Exosomes. PLoS ONE 2016, 11, e0147236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conigliaro, A.; Cicchini, C. Exosome-Mediated Signaling in Epithelial to Mesenchymal Transition and Tumor Progression. J. Clin. Med. 2018, 8, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amicone, L.; Citarella, F.; Cicchini, C. Epigenetic regulation in hepatocellular carcinoma requires long noncoding RNAs. BioMed Res. Int. 2015, 2015, 473942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battistelli, C.; Cicchini, C.; Santangelo, L.; Tramontano, A.; Grassi, L.; Gonzalez, F.J.; de Nonno, V.; Grassi, G.; Amicone, L.; Tripodi, M. The Snail repressor recruits EZH2 to specific genomic sites through the enrollment of the lncRNA HOTAIR in epithelial-to-mesenchymal transition. Oncogene 2017, 36, 942–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Chen, B.; Dong, P.; Zheng, J. HOTAIR Epigenetically Modulates PTEN Expression via MicroRNA-29b: A Novel Mechanism in Regulation of Liver Fibrosis. Mol. Ther. 2017, 25, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Ota, Y.; Takahashi, K.; Otake, S.; Tamaki, Y.; Okada, M.; Aso, K.; Makino, Y.; Fujii, S.; Ota, T.; Haneda, M. Extracellular vesicle-encapsulated miR-30e suppresses cholangiocarcinoma cell invasion and migration via inhibiting epithelial-mesenchymal transition. Oncotarget 2018, 9, 16400–16417. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.-M.; Li, Z.; Han, X.-D.; Shi, J.-H.; Tu, D.-Y.; Song, W.; Zhang, J.; Qiu, X.-L.; Ren, Y.; Zhen, L.-L. MiR-30e inhibits tumor growth and chemoresistance via targeting IRS1 in Breast Cancer. Sci. Rep. 2017, 7, 15929. [Google Scholar] [CrossRef]
- Croset, M.; Pantano, F.; Kan, C.W.S.; Bonnelye, E.; Descotes, F.; Alix-Panabières, C.; Lecellier, C.-H.; Bachelier, R.; Allioli, N.; Hong, S.-S.; et al. miRNA-30 Family Members Inhibit Breast Cancer Invasion, Osteomimicry, and Bone Destruction by Directly Targeting Multiple Bone Metastasis-Associated Genes. Cancer Res. 2018, 78, 5259–5273. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Zhao, L.; Zhao, S.; Guo, T.; Li, F.; Li, Z.; Fang, L.; Wu, T.; Gu, C. MicroRNA-30e inhibits proliferation and invasion of non-small cell lung cancer via targeting SOX9. Hum. Cell 2019, 32, 326–333. [Google Scholar] [CrossRef]
- Xu, G.; Cai, J.; Wang, L.; Jiang, L.; Huang, J.; Hu, R.; Ding, F. MicroRNA-30e-5p suppresses non-small cell lung cancer tumorigenesis by regulating USP22-mediated Sirt1/JAK/STAT3 signaling. Exp. Cell Res. 2018, 362, 268–278. [Google Scholar] [CrossRef]
- Liu, K.; Xie, F.; Gao, A.; Zhang, R.; Zhang, L.; Xiao, Z.; Hu, Q.; Huang, W.; Huang, Q.; Lin, B.; et al. SOX2 regulates multiple malignant processes of breast cancer development through the SOX2/miR-181a-5p, miR-30e-5p/TUSC3 axis. Mol. Cancer 2017, 16, 62. [Google Scholar] [CrossRef] [PubMed]
- Laudato, S.; Patil, N.; Abba, M.L.; Leupold, J.H.; Benner, A.; Gaiser, T.; Marx, A.; Allgayer, H. P53-induced miR-30e-5p inhibits colorectal cancer invasion and metastasis by targeting ITGA6 and ITGB1. Int. J. Cancer 2017, 141, 1879–1890. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, X.; Zhu, Z.; Li, W.; Yu, G.; Jia, Z.; Wang, X. Prostate carcinoma cell-derived exosomal MicroRNA-26a modulates the metastasis and tumor growth of prostate carcinoma. Biomed. Pharmacother. 2019, 117, 109109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Duan, W.; Sun, W. LncRNA SNHG6 promotes the migration, invasion, and epithelial-mesenchymal transition of colorectal cancer cells by miR-26a/EZH2 axis. Onco Targets Ther. 2019, 12, 3349–3360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.C.; Li, S.Y.; Jia, Y.F. MicroRNA-26a suppresses the malignant biological behaviors of papillary thyroid carcinoma by targeting ROCK1 and regulating PI3K/AKT signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8940–8949. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Guan, M.; Jia, Y.; Wang, D.; Pang, R.; Lv, F.; Xiao, Z.; Wang, L.; Zhang, H.; Xue, Y. The coordinated roles of miR-26a and miR-30c in regulating TGFβ1-induced epithelial-to-mesenchymal transition in diabetic nephropathy. Sci. Rep. 2016, 6, 37492. [Google Scholar] [CrossRef]
- Chen, X.; Xiao, W.; Chen, W.; Liu, X.; Wu, M.; Bo, Q.; Luo, Y.; Ye, S.; Cao, Y.; Liu, Y. MicroRNA-26a and -26b inhibit lens fibrosis and cataract by negatively regulating Jagged-1/Notch signaling pathway. Cell Death Differ. 2017, 24, 1431–1442. [Google Scholar] [CrossRef] [Green Version]
- Fan, B.; Li, L.; Xiao, D.J.; Xu, T. Study of miR-26a inhibiting epithelial-mesenchymal transition and invasion of Tu686 cell line through SMAD1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 217–224. [Google Scholar] [CrossRef]
- Geng, C.; Wei, J.; Wu, C. Yap-Hippo pathway regulates cerebral hypoxia-reoxygenation injury in neuroblastoma N2a cells via inhibiting ROCK1/F-actin/mitochondrial fission pathways. Acta Neurol. Belg. 2018, 1–14. [Google Scholar] [CrossRef]
- Slabáková, E.; Culig, Z.; Remšík, J.; Souček, K. Alternative mechanisms of miR-34a regulation in cancer. Cell Death Dis. 2017, 8, e3100. [Google Scholar] [CrossRef]
- Li, Y.-Y.; Tao, Y.-W.; Gao, S.; Li, P.; Zheng, J.-M.; Zhang, S.-E.; Liang, J.; Zhang, Y. Cancer-associated fibroblasts contribute to oral cancer cells proliferation and metastasis via exosome-mediated paracrine miR-34a-5p. EBioMedicine 2018, 36, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Rokavec, M.; Öner, M.G.; Li, H.; Jackstadt, R.; Jiang, L.; Lodygin, D.; Kaller, M.; Horst, D.; Ziegler, P.K.; Schwitalla, S.; et al. IL-6R/STAT3/miR-34a feedback loop promotes EMT-mediated colorectal cancer invasion and metastasis. J. Clin. Investig. 2014, 124, 1853–1867. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Tian, X.; Li, Y.; Wang, Z.; Li, X.; Zhu, C. miR-27b and miR-34a enhance docetaxel sensitivity of prostate cancer cells through inhibiting epithelial-to-mesenchymal transition by targeting ZEB1. Biomed. Pharmacother. 2018, 97, 736–744. [Google Scholar] [CrossRef]
- Liang, J.; Li, Y.; Daniels, G.; Sfanos, K.; De Marzo, A.; Wei, J.; Li, X.; Chen, W.; Wang, J.; Zhong, X.; et al. LEF1 Targeting EMT in Prostate Cancer Invasion Is Regulated by miR-34a. Mol. Cancer Res. 2015, 13, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, P.; Xiong, Y.; Watari, H.; Hanley, S.J.B.; Konno, Y.; Ihira, K.; Yamada, T.; Kudo, M.; Yue, J.; Sakuragi, N. MiR-137 and miR-34a directly target Snail and inhibit EMT, invasion and sphere-forming ability of ovarian cancer cells. J. Exp. Clin. Cancer Res. 2016, 35, 132. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Wang, G.; Liu, H.; Xiong, C. SATB2 targeted by methylated miR-34c-5p suppresses proliferation and metastasis attenuating the epithelial-mesenchymal transition in colorectal cancer. Cell Prolif. 2018, 51, e12455. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Li, Y.; Pan, Q.; Ye, M.; He, S.; Tian, Q.; Xue, M. MiR-34c/SOX9 axis regulates the chemoresistance of ovarian cancer cell to cisplatin-based chemotherapy. J. Cell Biochem. 2019, 120, 2940–2953. [Google Scholar] [CrossRef]
- Russo, V.; Paciocco, A.; Affinito, A.; Roscigno, G.; Fiore, D.; Palma, F.; Galasso, M.; Volinia, S.; Fiorelli, A.; Esposito, C.L.; et al. Aptamer-miR-34c Conjugate Affects Cell Proliferation of Non-Small-Cell Lung Cancer Cells. Mol. Ther. Nucleic Acids 2018, 13, 334–346. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Li, W.-Z.; Huang, M.-L.; Wei, H.-L.; Wang, T.; Fan, J.; Li, N.-L.; Ling, R. Regulation of cancerous progression and epithelial-mesenchymal transition by miR-34c-3p via modulation of MAP3K2 signaling in triple-negative breast cancer cells. Biochem. Biophys. Res. Commun. 2017, 483, 10–16. [Google Scholar] [CrossRef]
- Wang, J.-X.; Yang, Y.; Li, K. Long noncoding RNA DANCR aggravates retinoblastoma through miR-34c and miR-613 by targeting MMP-9. J. Cell Physiol. 2018, 233, 6986–6995. [Google Scholar] [CrossRef]
- Peng, D.; Wang, H.; Li, L.; Ma, X.; Chen, Y.; Zhou, H.; Luo, Y.; Xiao, Y.; Liu, L. miR-34c-5p promotes eradication of acute myeloid leukemia stem cells by inducing senescence through selective RAB27B targeting to inhibit exosome shedding. Leukemia 2018, 32, 1180–1188. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Du, L.-Y.; Guo, F.; Li, X.; Cheng, B. Exosomes derived from microRNA-101-3p-overexpressing human bone marrow mesenchymal stem cells suppress oral cancer cell proliferation, invasion, and migration. Mol. Cell Biochem. 2019, 458, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Cogdell, D.; Hu, L.; Yang, D.; Sood, A.K.; Xue, F.; Zhang, W. MiR-101 suppresses the epithelial-to-mesenchymal transition by targeting ZEB1 and ZEB2 in ovarian carcinoma. Oncol. Rep. 2014, 31, 2021–2028. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Zhao, Y.; Li, X.; Tao, Z.; Hou, M.; Ma, H. Overexpression of HIF-2α-Dependent NEAT1 Promotes the Progression of Non-Small Cell Lung Cancer through miR-101-3p/SOX9/Wnt/β-Catenin Signal Pathway. Cell Physiol. Biochem. 2019, 52, 368–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Z.; Yin, S.; Su, Z.; Bai, M.; Zhang, H.; Hei, Z.; Cai, S. Downregulation of miR-101 contributes to epithelial-mesenchymal transition in cisplatin resistance of NSCLC cells by targeting ROCK2. Oncotarget 2016, 7, 37524–37535. [Google Scholar] [CrossRef]
- Wei, X.; Tang, C.; Lu, X.; Liu, R.; Zhou, M.; He, D.; Zheng, D.; Sun, C.; Wu, Z. MiR-101 targets DUSP1 to regulate the TGF-β secretion in sorafenib inhibits macrophage-induced growth of hepatocarcinoma. Oncotarget 2015, 6, 18389–18405. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Shao, M.-Y.; Zou, S.-C.; Xiao, Z.-F.; Chen, Z.-C. MiR-101-3p inhibits EMT to attenuate proliferation and metastasis in glioblastoma by targeting TRIM44. J. Neurooncol. 2019, 141, 19–30. [Google Scholar] [CrossRef]
- Che, Y.; Shi, X.; Shi, Y.; Jiang, X.; Ai, Q.; Shi, Y.; Gong, F.; Jiang, W. Exosomes Derived from miR-143-Overexpressing MSCs Inhibit Cell Migration and Invasion in Human Prostate Cancer by Downregulating TFF3. Mol. Ther. Nucleic Acids 2019, 18, 232–244. [Google Scholar] [CrossRef] [Green Version]
- Yusup, A.; Huji, B.; Fang, C.; Wang, F.; Dadihan, T.; Wang, H.-J.; Upur, H. Expression of trefoil factors and TWIST1 in colorectal cancer and their correlation with metastatic potential and prognosis. World J. Gastroenterol. 2017, 23, 110–120. [Google Scholar] [CrossRef]
- Lei, C.; Du, F.; Sun, L.; Li, T.; Li, T.; Min, Y.; Nie, A.; Wang, X.; Geng, L.; Lu, Y.; et al. miR-143 and miR-145 inhibit gastric cancer cell migration and metastasis by suppressing MYO6. Cell Death Dis. 2017, 8, e3101. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Chen, J.; Li, H.; Yang, Y.; Yun, H.; Yang, S.; Mao, X. LncRNA UCA1 promotes the invasion and EMT of bladder cancer cells by regulating the miR-143/HMGB1 pathway. Oncol. Lett 2017, 14, 5556–5562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, L.; Ma, C.; Li, W.; Yang, S.; Liu, Z. miR-143 suppresses epithelial-mesenchymal transition and inhibits tumor growth of breast cancer through down-regulation of ERK5. Mol. Carcinog. 2016, 55, 1990–2000. [Google Scholar] [CrossRef] [PubMed]
- Bufalino, A.; Cervigne, N.K.; de Oliveira, C.E.; Fonseca, F.P.; Rodrigues, P.C.; Macedo, C.C.S.; Sobral, L.M.; Miguel, M.C.; Lopes, M.A.; Paes Leme, A.F.; et al. Low miR-143/miR-145 Cluster Levels Induce Activin A Overexpression in Oral Squamous Cell Carcinomas, Which Contributes to Poor Prognosis. PLoS ONE 2015, 10, e0136599. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Shen, S.; Zheng, X.; Ye, K.; Sun, Y.; Lu, Y.; Ge, H. Long noncoding RNA HAGLR acts as a microRNA-143-5p sponge to regulate epithelial-mesenchymal transition and metastatic potential in esophageal cancer by regulating LAMP3. FASEB J. 2019, 33, 10490–10504. [Google Scholar] [CrossRef]
- Luo, H.; Liang, C. MicroRNA-148b inhibits proliferation and the epithelial-mesenchymal transition and increases radiosensitivity in non-small cell lung carcinomas by regulating ROCK1. Exp. Ther. Med. 2018, 15, 3609–3616. [Google Scholar] [CrossRef]
- Zhou, Z.; Su, Y.; Fa, X. Restoration of BRG1 inhibits proliferation and metastasis of lung cancer by regulating tumor suppressor miR-148b. Onco Targets Ther. 2015, 8, 3603–3612. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Liu, Y.; Qu, Y.; Liu, L.; Li, H. Exosomes Derived From MicroRNA-148b-3p-Overexpressing Human Umbilical Cord Mesenchymal Stem Cells Restrain Breast Cancer Progression. Front. Oncol. 2019, 9, 1076. [Google Scholar] [CrossRef] [Green Version]
- Li, B.-L.; Lu, W.; Qu, J.-J.; Ye, L.; Du, G.-Q.; Wan, X.-P. Loss of exosomal miR-148b from cancer-associated fibroblasts promotes endometrial cancer cell invasion and cancer metastasis. J. Cell Physiol. 2019, 234, 2943–2953. [Google Scholar] [CrossRef]
- Senfter, D.; Holzner, S.; Kalipciyan, M.; Staribacher, A.; Walzl, A.; Huttary, N.; Krieger, S.; Brenner, S.; Jäger, W.; Krupitza, G.; et al. Loss of miR-200 family in 5-fluorouracil resistant colon cancer drives lymphendothelial invasiveness in vitro. Hum. Mol. Genet. 2015, 24, 3689–3698. [Google Scholar] [CrossRef] [Green Version]
- Title, A.C.; Hong, S.-J.; Pires, N.D.; Hasenöhrl, L.; Godbersen, S.; Stokar-Regenscheit, N.; Bartel, D.P.; Stoffel, M. Genetic dissection of the miR-200-Zeb1 axis reveals its importance in tumor differentiation and invasion. Nat. Commun. 2018, 9, 4671. [Google Scholar] [CrossRef]
- Pan, Q.; Meng, L.; Ye, J.; Wei, X.; Shang, Y.; Tian, Y.; He, Y.; Peng, Z.; Chen, L.; Chen, W.; et al. Transcriptional repression of miR-200 family members by Nanog in colon cancer cells induces epithelial-mesenchymal transition (EMT). Cancer Lett. 2017, 392, 26–38. [Google Scholar] [CrossRef]
- Kundu, S.T.; Byers, L.A.; Peng, D.H.; Roybal, J.D.; Diao, L.; Wang, J.; Tong, P.; Creighton, C.J.; Gibbons, D.L. The miR-200 family and the miR-183~96~182 cluster target Foxf2 to inhibit invasion and metastasis in lung cancers. Oncogene 2016, 35, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Le, M.T.N.; Hamar, P.; Guo, C.; Basar, E.; Perdigão-Henriques, R.; Balaj, L.; Lieberman, J. miR-200-containing extracellular vesicles promote breast cancer cell metastasis. J. Clin. Investig. 2014, 124, 5109–5128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De, A.; Powers, B.; De, A.; Zhou, J.; Sharma, S.; Van Veldhuizen, P.; Bansal, A.; Sharma, R.; Sharma, M. Emblica officinalis extract downregulates pro-angiogenic molecules via upregulation of cellular and exosomal miR-375 in human ovarian cancer cells. Oncotarget 2016, 7, 31484–31500. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.-Z.; Lai, M.-F.; Li, Y.-P.; Xu, C.-H.; Zhang, H.-R.; Kuang, J.-G. Human marrow stromal cells secrete microRNA-375-containing exosomes to regulate glioma progression. Cancer Gene Ther. 2019, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.-W.; Lin, J.-S.; He, X.-X. The emerging role of miR-375 in cancer. Int. J. Cancer 2014, 135, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Selth, L.A.; Das, R.; Townley, S.L.; Coutinho, I.; Hanson, A.R.; Centenera, M.M.; Stylianou, N.; Sweeney, K.; Soekmadji, C.; Jovanovic, L.; et al. A ZEB1-miR-375-YAP1 pathway regulates epithelial plasticity in prostate cancer. Oncogene 2017, 36, 24–34. [Google Scholar] [CrossRef]
- Guo, F.; Gao, Y.; Sui, G.; Jiao, D.; Sun, L.; Fu, Q.; Jin, C. miR-375-3p/YWHAZ/β-catenin axis regulates migration, invasion, EMT in gastric cancer cells. Clin. Exp. Pharmacol. Physiol. 2019, 46, 144–152. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Circulating Exosome RNA in Lung Metastases of Primary High-Grade Osteosarcoma. 2017. Available online: https://clinicaltrials.gov/ct2/show/NCT03108677 (accessed on 11 April 2017).
- ClinicalTrials.gov. Identification and Characterization of Predictive Factors of Onset of Bone Metastases in Cancer Patients. Available online: https://clinicaltrials.gov/ct2/show/NCT03895216 (accessed on 29 March 2019).
- ClinicalTrials.gov. Trial of a Vaccination with Tumor Antigen-Loaded Dendritic Cell-Derived Exosomes. Available online: https://clinicaltrials.gov/ct2/show/NCT01159288 (accessed on 9 July 2010).
- ClinicalTrials.gov. Study Investigating the Ability of Plant Exosomes to Deliver Curcumin to Normal and Colon Cancer Tissue. 2011. Available online: https://clinicaltrials.gov/ct2/show/NCT01294072 (accessed on 11 February 2011).
- ClinicalTrials.gov. iExosomes in Treating Participants with Metastatic Pancreas Cancer with KrasG12D Mutation. Available online: https://clinicaltrials.gov/ct2/show/NCT03608631 (accessed on 1 August 2018).
- Kamerkar, S.; LeBleu, V.S.; Sugimoto, H.; Yang, S.; Ruivo, C.F.; Melo, S.A.; Lee, J.J.; Kalluri, R. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature 2017, 546, 498–503. [Google Scholar] [CrossRef]
- Theodoraki, M.-N.; Yerneni, S.S.; Brunner, C.; Theodorakis, J.; Hoffmann, T.K.; Whiteside, T.L. Plasma-derived Exosomes Reverse Epithelial-to-Mesenchymal Transition after Photodynamic Therapy of Patients with Head and Neck Cancer. Oncoscience 2018, 5, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Wels, C.; Joshi, S.; Koefinger, P.; Bergler, H.; Schaider, H. Transcriptional activation of ZEB1 by Slug leads to cooperative regulation of the epithelial-mesenchymal transition-like phenotype in melanoma. J. Investig. Dermatol. 2011, 131, 1877–1885. [Google Scholar] [CrossRef] [Green Version]
- Monfared, H.; Jahangard, Y.; Nikkhah, M.; Mirnajafi-Zadeh, J.; Mowla, S.J. Potential Therapeutic Effects of Exosomes Packed With a miR-21-Sponge Construct in a Rat Model of Glioblastoma. Front. Oncol. 2019, 9, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krichevsky, A.M.; Gabriely, G. miR-21: A small multi-faceted RNA. J. Cell Mol. Med. 2009, 13, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.M.; Cardoso, A.L.; Nóbrega, C.; Pereira de Almeida, L.F.; Bruce, J.N.; Canoll, P.; Pedroso de Lima, M.C. MicroRNA-21 silencing enhances the cytotoxic effect of the antiangiogenic drug sunitinib in glioblastoma. Hum. Mol. Genet. 2013, 22, 904–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyuno, D.; Zhao, K.; Bauer, N.; Ryschich, E.; Zöller, M. Therapeutic Targeting Cancer-Initiating Cell Markers by Exosome miRNA: Efficacy and Functional Consequences Exemplified for claudin7 and EpCAM. Transl. Oncol. 2019, 12, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Philip, R.; Heiler, S.; Mu, W.; Büchler, M.W.; Zöller, M.; Thuma, F. Claudin-7 promotes the epithelial-mesenchymal transition in human colorectal cancer. Oncotarget 2015, 6, 2046–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Zhou, C.-Z.; Zhu, R.; Fan, H.; Liu, X.-X.; Duan, X.-Y.; Tang, Q.; Shou, Z.-X.; Zuo, D.-M. miR-200b-containing microvesicles attenuate experimental colitis associated intestinal fibrosis by inhibiting epithelial-mesenchymal transition. J. Gastroenterol. Hepatol. 2017, 32, 1966–1974. [Google Scholar] [CrossRef]
- Almanza, G.; Rodvold, J.J.; Tsui, B.; Jepsen, K.; Carter, H.; Zanetti, M. Extracellular vesicles produced in B cells deliver tumor suppressor miR-335 to breast cancer cells disrupting oncogenic programming in vitro and in vivo. Sci. Rep. 2018, 8, 17581. [Google Scholar] [CrossRef]
- Fujiwara, T.; Eguchi, T.; Sogawa, C.; Ono, K.; Murakami, J.; Ibaragi, S.; Asaumi, J.-I.; Calderwood, S.K.; Okamoto, K.; Kozaki, K.-I. Carcinogenic epithelial-mesenchymal transition initiated by oral cancer exosomes is inhibited by anti-EGFR antibody cetuximab. Oral Oncol. 2018, 86, 251–257. [Google Scholar] [CrossRef]
- Yang, T.; Martin, P.; Fogarty, B.; Brown, A.; Schurman, K.; Phipps, R.; Yin, V.P.; Lockman, P.; Bai, S. Exosome delivered anticancer drugs across the blood-brain barrier for brain cancer therapy in Danio rerio. Pharm. Res. 2015, 32, 2003–2014. [Google Scholar] [CrossRef]
- Vázquez-Ríos, A.J.; Molina-Crespo, Á.; Bouzo, B.L.; López-López, R.; Moreno-Bueno, G.; de la Fuente, M. Exosome-mimetic nanoplatforms for targeted cancer drug delivery. J. Nanobiotechnol. 2019, 17, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic representation of exosomes. Exosomes are generated from multivesicular endosome (MVE), loaded with cellular components in donor cells, and secreted to the extracellular environment. Exosomes transfer proteins, nucleic acids, metabolites, and even organelles to recipient cells in both autocrinal and paracrinal manners. The uptake of exosomes is mediated by receptor activation, endocytosis, and membrane fusion.
Figure 1.
Schematic representation of exosomes. Exosomes are generated from multivesicular endosome (MVE), loaded with cellular components in donor cells, and secreted to the extracellular environment. Exosomes transfer proteins, nucleic acids, metabolites, and even organelles to recipient cells in both autocrinal and paracrinal manners. The uptake of exosomes is mediated by receptor activation, endocytosis, and membrane fusion.

Figure 2.
Comprehensive schematic diagram of signaling pathways related to exosome-induced EMT in cancer. The principal signaling pathways and transcription factors involved in exosome-mediated EMT induction are depicted.
Figure 2.
Comprehensive schematic diagram of signaling pathways related to exosome-induced EMT in cancer. The principal signaling pathways and transcription factors involved in exosome-mediated EMT induction are depicted.

Figure 3.
Schematic representation of exosome-induced EMT in cancer. The involvement of EMT-related signaling pathways during exosome-mediated EMT is depicted. The blue triangle indicates activation and red inverted triangle indicates inactivation of the signal.
Figure 3.
Schematic representation of exosome-induced EMT in cancer. The involvement of EMT-related signaling pathways during exosome-mediated EMT is depicted. The blue triangle indicates activation and red inverted triangle indicates inactivation of the signal.


{kind=link}
{kind=link}
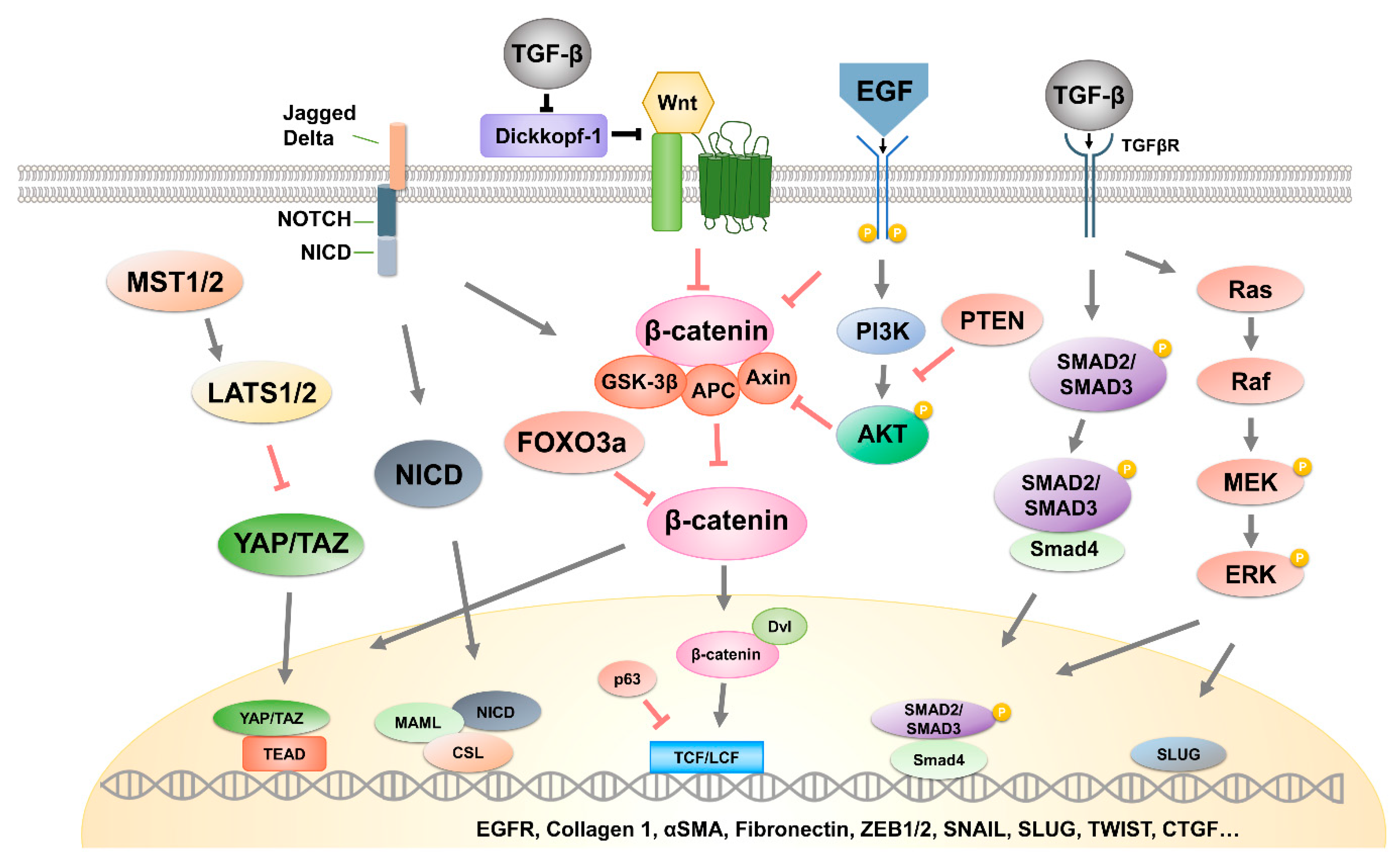{kind=link}
{kind=link}
Table 1.
The contents of exosomes and their mechanisms leading to EMT in cancer.
| Name | Cancer Type | Target | Signaling Pathway | Reference | |
|---|---|---|---|---|---|
| miRNA | miR-665 | Hepatocellular carcinoma | PTPRB | Hippo pathway | [23,24] |
| miR-34c | Nasopharyngeal carcinoma | β-catenin, SOX9, SATB2, MAP3K2, DANCR, RAB27B | Wnt/β-catenin pathway | [92,93,94,95,96,97] | |
| miR-301a | GBM | TCEAL7, p63, ZEB1, ZEB2, E-cadherin | [31,32] | ||
| miR-92a | Colorectal cancer | FBXW7, MOAP1 | [36] | ||
| miR-155 | Breast cancer | FOXO3a, SGK3, p85a | [38,39,40,41,42,43] | ||
| miR-30c | Esophageal cancer | circPRMT5, BCL9 | [44,45,46] | ||
| miR-1260b | Lung adenocarcinoma | SOCS6, YY1 | [33] | ||
| miR-375 | Gastric cancer | 14-3-3ζ, YAP, SNAI1, SLC31A1 | [120,121,122,123,124] | ||
| miR-34a | OSCC | STAT3, LEF1, ZEB1, SNAI1 | NOTCH signaling pathway | [87,88,89,90,91] | |
| miR-1260b | NSCLC | KIT, MEK, ERK | ERK pathway | [34,35] | |
| miR-21 | OSCC | HIF-1a, HIF-2a, PTEN | [64] | ||
| miR-92a | NSCLC | PTEN, PI3K, AKT | PI3K/AKT pathway | [37] | |
| Protein | AREG | Chronic myelogenous leukemia | EGFR, SNAI1, YAP/TAZ | Hippo pathway | [25,26] |
| linc-ROR | Breast cancer | miRNP, miR-205, ZEB2 | Wnt/β-catenin pathway | [69] | |
| Cir-ABCC1 | Colorectal cancer | Unclear | [47] | ||
| UBR2 | Gastric cancer | p53 | [60] | ||
| TNC | PDAC | E-cadherin, CD44, MMP-7, MET | [61] | ||
| TNC | Breast cancer | p-Y418, p-FAK, SRC | [62] | ||
| Wnt1 | Colorectal cancer | TCF/LEF | [48] | ||
| Wnt3a | Colorectal cancer | TCF/LEF | [49] | ||
| Wnt4 | Colorectal cancer | HIF1α | [54] | ||
| Wnt5a | Breast cancer | p38 | [50,57,58] | ||
| Wnt10b | Breast cancer | p85a | [51] | ||
| Wnt11a | Breast cancer | PCP | [52,53] | ||
| HOTAIR | Bladder cancer | PTEN, miR-29b, DNMT3b | MAPK/ERK pathway | [81,82,83,84,85] | |
| Cbl | Liver cancerGastric cancer | Rab27a, AKT | [65,66,67] | ||
| lncRNA-Sox2ot | PDAC | Sox2ot, miR-200 | Sox2ot pathway | [68] | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, H.; Lee, S.; Shin, E.; Seong, K.M.; Jin, Y.W.; Youn, H.; Youn, B. The Emerging Roles of Exosomes as EMT Regulators in Cancer. Cells 2020, 9, 861. https://doi.org/10.3390/cells9040861
AMA Style
Kim H, Lee S, Shin E, Seong KM, Jin YW, Youn H, Youn B. The Emerging Roles of Exosomes as EMT Regulators in Cancer. Cells. 2020; 9(4):861. https://doi.org/10.3390/cells9040861
Chicago/Turabian StyleKim, Hyunwoo, Sungmin Lee, Eunguk Shin, Ki Moon Seong, Young Woo Jin, HyeSook Youn, and BuHyun Youn. 2020. "The Emerging Roles of Exosomes as EMT Regulators in Cancer" Cells 9, no. 4: 861. https://doi.org/10.3390/cells9040861
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.