RNA Editors, Cofactors, and mRNA Targets: An Overview of the C-to-U RNA Editing Machinery and Its Implication in Human Disease


1
Division of Immune Diversity, Program in Cancer Immunology, German Cancer Research Centre, 69120 Heidelberg, Germany
2
Division of Biosciences, Uni Heidelberg, 69120 Heidelberg, Germany
*
Authors to whom correspondence should be addressed.
Genes 2019, 10(1), 13; https://doi.org/10.3390/genes10010013
Submission received: 1 November 2018
/
Revised: 10 December 2018
/
Accepted: 20 December 2018
/
Published: 27 December 2018
(This article belongs to the Special Issue The Epitranscriptome in Human Disease)
Abstract
:One of the most prevalent epitranscriptomic modifications is RNA editing. In higher eukaryotes, RNA editing is catalyzed by one of two classes of deaminases: ADAR family enzymes that catalyze A-to-I (read as G) editing, and AID/APOBEC family enzymes that catalyze C-to-U. ADAR-catalyzed deamination has been studied extensively. Here we focus on AID/APOBEC-catalyzed editing, and review the emergent knowledge regarding C-to-U editing consequences in the context of human disease.

{kind=link}
1. Introduction
Epitranscriptomics is a recently coined term that refers to the study of RNA modifications and their effects on the transcriptome. Included under this umbrella is everything from RNA tertiary structure, to processing, stability, localization, and translational efficiency. To date, more than 150 distinct types of RNA modifications have been identified, primarily in ribosomal RNA (rRNA) and transfer RNA (tRNA) [1]. Many of these are suspected to be essential, as loss of tRNA or rRNA modification “writers” or “readers” are often found to be associated with diseases (e.g., cancer—[2])
A small subset of these modifications have also been found in messenger RNA (mRNA) (reviewed in [3]), where they are thought to represent a new mode of regulation of the biological information influencing the fate of each transcript. The most prominent of these modifications is the deamination of adenosine or cytosine (to inosine (decoded as guanosine) or uracil, respectively). Because these modifications effectively lead to single- nucleotide changes in mRNA, the phenomenon has also been termed “RNA editing”. This is distinct from the “editing” described in the mitochondrion of trypanosomes, which refers to the insertion or deletion of uridines to correct the “frame” [4].
mRNA deamination is mediated by two classes of enzymes: the “adenosine deaminases that act on RNA” (ADARs) and the cytidine deaminase family of “Apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like” (also known as the activation-induced deaminase AID/APOBEC family). Here we will focus on AID/APOBEC-mediated RNA editing, and review the impacts of such editing on single transcripts, the cell, and the whole organism.
2. AID/APOBECs: A Common Path through Evolution
APOBEC1 was the first member of the AID/APOBEC family to be discovered and successfully cloned [5]. This enzyme deaminates a specific cytosine in the long apolipoprotein B (Apob) pre-mRNA; the C-to-U editing event recodes a CAA codon (Q2180) to a stop codon, resulting in a truncated form of the APOB protein, called APOB-48 [5,6]. Because of this functionality, the Apob RNA editor was termed “apolipoprotein-B mRNA editing enzyme, catalytic polypeptide-1” (APOBEC1). The rest of the family derives its name from this founding member.
There are several members in the AID/APOBEC family, and all of them (except for APOBEC2 and APOBEC4) have C-to-U deaminase activity on single-stranded DNA or RNA. All family members share the structural and catalytic backbone of the zinc-dependent deaminase superfamily, which also comprises proteins such as ADATs and ADARs, which are involved in the metabolism of purines or pyrimidines. They are also evolutionarily conserved. The first member of the family to appear in the vertebrate lineage is the activation-induced cytidine deaminase (AID), a molecule that is crucial to antibody diversification [7], whose appearance coincides in sharks with the dawn of adaptive immunity [8]. APOBEC1 appeared soon thereafter, from an inverted duplication of the Aid locus [9]. In primates, Aid and Apobec1 are linked, with the Apobec1 locus being only 1 Mb away from the Aid locus [9]. The main difference between AID and APOBEC1 is the primary physiological choice of the nucleic acid substrate; single stranded DNA for AID, and RNA for APOBEC1 [10,11]. A secondary difference is the presence of a longer coding sequence at the 3′ end of the Apobec1 gene, which might have implications for dimerization [12,13], nucleocytoplasmic transport, and possibly substrate binding [14,15].
The Apobec3 (A3) locus [16], appeared after the divergence of the marsupial and placental lineages. A duplication formed the first two A3 genes from which all the other A3s have evolved [9]. In some species, for example in rodents, these two original genes fused into a single gene with a double zinc-coordinating domain.
3. APOBECs as RNA Editors
AID/APOBECs show physiological preference for specific nucleic acid substrates (e.g., RNA or DNA) but most of them can deaminate both, under certain conditions [15]. Here we will focus on the enzymes for which RNA editing activity has been reported, i.e., in APOBEC1, A3A [17], and A3G [18]). We will first review how these molecules were discovered, whether they might require co-factors for activity, and will discuss their known mRNA targets. Finally, we will consider emergent knowledge on how mutations within the editing enzymes or their co-factors (or their mRNA targets) might mediate human disease.
4. APOBEC1
4.1. Key Facts and Controversies
The full length APOB protein (APOB-100) in humans is synthesized in liver, and is the major protein component of plasma low-density lipoproteins (LDLs). In contrast, the edited form (APOB-48) is produced in the small intestine, and is essential for the secretion of chylomicrons [19,20]. Lipoproteins containing APOB-48 are quickly cleared from plasma, and are not converted into LDL. Mammals that edit Apob in liver (horses, rats, mice and dogs) have low levels of plasma LDLs [21]. As soon as APOBEC1 was defined as the key enzyme in Apob editing, the Innerarity group overexpressed APOBEC1 in the liver of mice and rabbits in an attempt to lower plasma LDL and demonstrate therapeutic potential. Instead, they uncovered an unexpected phenotype: these animals developed hepatocellular carcinoma [22]. This was the first association between APOBEC1 editing activity and a specific disease.
Shortly after the discovery of APOBEC1, in vivo experiments showed that RNA editing by APOBEC1 is not redundant; APOBEC1 is the only responsible protein for this deaminase activity [23,24]. Loss of APOBEC1 resulted in viable mice, despite alterations in lipoprotein metabolism [25]. These initial experiments also showed that APOBEC1 was essential for editing, but that additional factors were also important. In vivo biochemical work showed that APOBEC1 alone in the presence of Apob RNA was unable to catalyze editing [5,26]. Indeed, the RNA binding affinity of APOBEC1 is rather low [27]. A few years later, two groups showed that the addition of an RNA-binding protein, termed the APOBEC1 complementation factor or A1CF (ACF), was sufficient in vitro to complement efficient C-to-U editing in the presence of APOBEC1 and Apob mRNA [28,29], thus defining a minimal editing complex.
From there and for the next 15 years, conflicting information was gathered regarding APOBEC1. Apob editing is a nuclear event [30,31], and several papers demonstrated the need for nucleo-cytoplasmic transport. However, the mechanism was debated. Several investigators maintained that APOBEC1 contains an NLS in its N-terminus, and an NES in the C-terminus [5,32,33,34], and that APOBEC1 was necessary and sufficient to shuffle the complex between the cytoplasm and the nucleus [32,33]. However, ACF also contains an NLS [35]. The question became one of dependencies. Some investigators noted that APOBEC1 import/export is importin-α- and exportin-1-dependent respectively, and inhibited by leptomycin B—thus suggesting that the shuttling was APOBEC1-dependent [32,36]. Others suggested that ACF shuttling was dominant, and a transcription-dependent process that is inhibited by actinomycin D treatment [35].
Another dispute regarded the function of the C-terminal part of APOBEC1. This portion of the protein distinguishes APOBEC1 from the rest of the family, and is probably important for homodimerization [12], which in turn was thought to be important for editing activity. C terminally deleted mutants of APOBEC1 cannot dimerize, and are deficient in RNA editing activity [37]. APOBEC1 C terminal domain-interacting proteins were mostly inhibitory to editing [13,38,39,40,41,42,43]. Nevertheless, APOBEC1 with partially deleted C-terminal domains are still able to edit RNA in vitro, indicating that the relation between editing and dimer formation is still not clear [44,45]. It is not a surprise that these data that rely on experiments performed in different cells lines, often overexpressing tagged proteins from different species, resulted in conclusions that were not always in agreement with each other. To date, there is no physiological cell line model for APOBEC1. In addition, good antibodies for APOBEC1 do not exist, and thus the ability to detect physiological levels of the protein in its proper context is missing. Such tools will be necessary to clarify how the APOBEC1 cofactor–RNA complex works.
4.2. Cofactors
Before the discovery of ACF, genetic experiments determined several sequence elements on Apob mRNA that were required for efficient editing by APOBEC1 [46,47]. These sequence elements were found within 50 bp surrounding the target C. Two elements were described: an element 5′ to the edited C, and a 3′ (11 nt) element, termed the mooring sequence. These local sequence elements also benefit from a surrounding region that was AU rich [48,49]. Based on this information, several papers hypothesized the possibility that these elements were important for the creation and stabilization of a secondary RNA structure, amenable to APOBEC1 editing [50,51,52].
The hypothesized minimal editing complex, consisting of ACF, APOBEC1, and Apob mRNA, led to a model in which ACF served as the RNA-binding protein (RBP), binding the mooring sequence [48,53] and recruiting (or positioning) APOBEC1 to the editing site [36,54,55]. Work from several labs suggested that the complex itself was co-ordinately regulated. Both APOBEC1 and ACF have splice variants that modulate expression [56,57,58,59], and both can be posttranscriptionally modified (which impacts their subcellular localization, editosome assembly, and editing activity) [60,61,62].
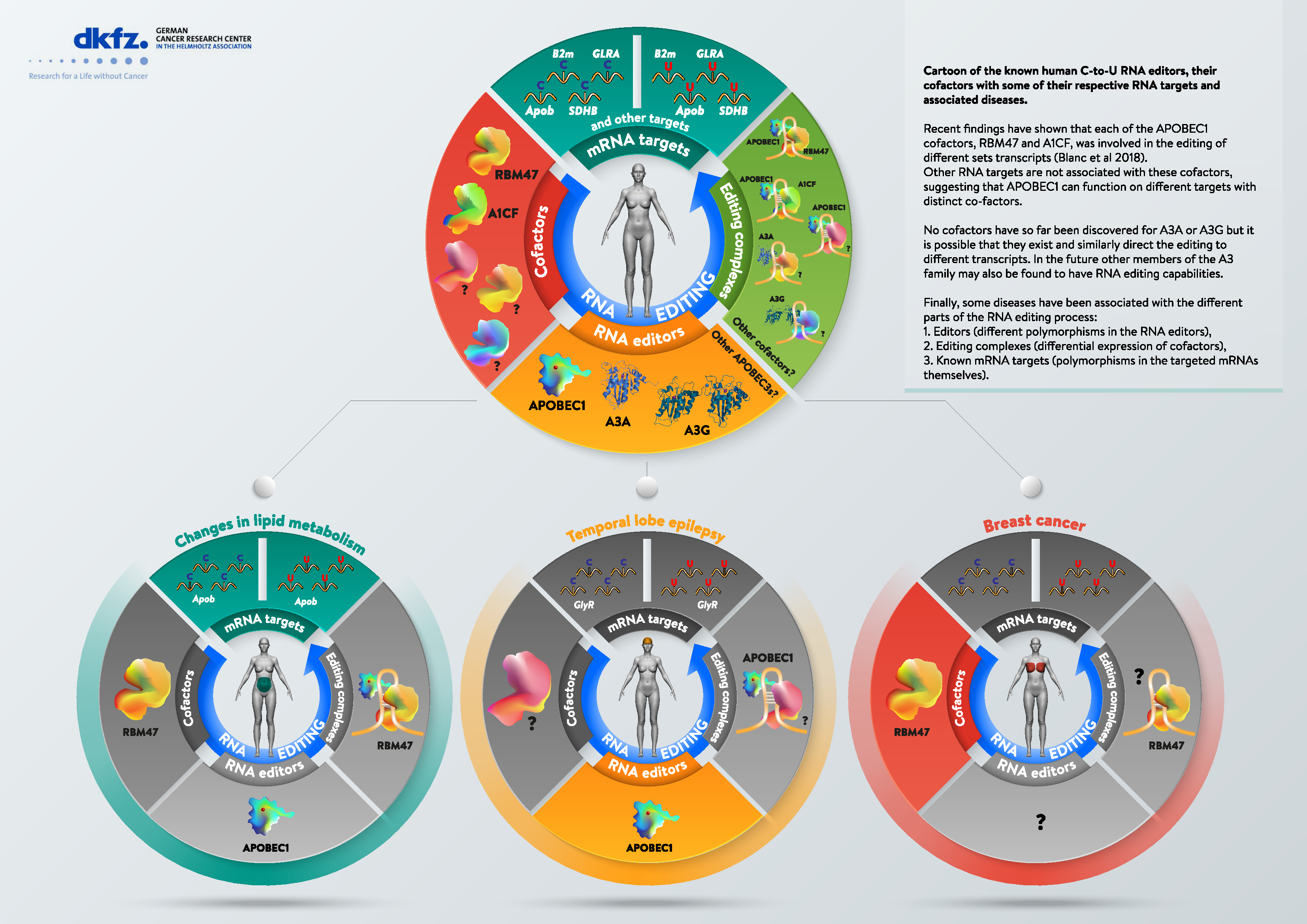
The model of an APOBEC1-ACF editosome was upended with the discovery of another RBP, the RNA-binding motif protein 47 (RBM47), as a cofactor indispensable for Apob editing in vivo, and an effective substitute for ACF in vitro [63]. Although this was not entirely unexpected [42], it was further confirmed by a new Acf−/− mouse model in which a different gene targeting strategy resulted in viable mice [64] (in contrast to the original knockout that suffered from early embryonic lethality [65]). In this model, the loss of ACF did not result in a loss of editing in the liver or small intestine, either in Apob, or in additional known edited transcripts [66,67]. This helped to explained a finding from the original knockout paper that reported that heterozygous Acf+/− mice were viable, but did not show a decrease, but rather an increase in intestinal Apob editing [68]. A very recent paper resolves this conundrum by using genetic means to demonstrate that ACF and RBM47 interact with (and attract APOBEC1 to) different sets of transcripts [69]. Specifically, these authors showed that a loss of ACF or RMB47 results in the loss of non-overlapping sets of transcripts (as assessed by transcriptome sequencing). Moreover, the loss of both ACF and RBM47 does not completely ablate APOBEC1-mediated editing. This suggested that APOBEC1 is able to induce RNA editing alone under physiological settings, or that additional RBP cofactors exist (perhaps expressed in a tissue or stage specific manner) that recruit APOBEC1 to specific transcripts. APOBEC1 is able to edit mRNA on its own in vitro ([70,71]) and in vivo (under overexpression or ectopic expression conditions a phenomenon described as “hyper editing” [22,32]). Whether this editing can happen under normal conditions remains unclear. The alternative possible model suggests that ACF and RBM47 (and perhaps other RBPs) exhibit distinctive tissue- and transcript-specific functions that modulate APOBEC1-dependent RNA editing site selection and activity [69].
This possibility is further supported by the discovery of another putative paralogue of ACF, DND1, which seems to interact with APOBEC1 in vitro though only weakly in 293T transfected cells [72]. Dnd1ter mutant mice develop testicular germ cell tumors (TGCT) [73], whose incidence can be modulated by the dosage of an Apobec1-null allele [74]. In this context, Apobec1 acts as a TGCT enhancer, an effect that is inherited in a conventional manner in the paternal germ-lineage, and is also transgenerationally inherited for at least two subsequent generations (regardless of Apobec1 genotype). These results suggest that a maternally inherited Apobec1 mutation can affect the tumorigenic susceptibility of two generations of offspring in a transgenerational epitranscriptomic manner [74]. The possibility that DND1 is another ACF/RBM47-like cofactor for APOBEC1 had previously been suggested [75]. Future studies will be necessary to confirm the interaction between APOBEC1 and DND1, and to understand which part of APOBEC1 interacts with RBM47 (as has been done for ACF [54]). Finally, it will be important to identify which transcripts are recruited by the distinct RRM motifs of these cofactors (perhaps in a sequence specific manner [76]). In this context, it is interesting to note that DND1 can inhibit miRNA-mediated RNA decay [77], and that APOBEC3G is able to modulate this function of DND1 [78], suggesting binding on similar transcripts/motifs of these two proteins.
4.3. RNA Targets and Their Functional Relevance
The best characterized RNA target of APOBEC1 is Apob, whose editing is essential for proper lipoprotein metabolism. The absence of editing leads to important lipid metabolic phenotypes in Apobec-1−/−. These mice express apoB100-only, and show spontaneous hypercholesterolemia when crossed into the LDLR−/− background [79]. Finally Apobec-1−/− mice crossed into a conditional intestinal Mttp deletor background result in lethal intestinal lipotoxicity [80].
APOBEC1 was initially thought to only edit Apob, and to be expressed only in APOB-expressing tissue (the small intestine in humans and the small intestine and liver in mice [24]). When the dependencies of APOBEC1 editing on the mooring sequence were discovered, other target RNAs were proposed. Neurofibromatosis type 1 (Nf1) mRNA was found to be edited within an alternatively spliced exon in a subset of patients with peripheral nerve sheath tumors [81,82]. Nat1 mRNA was found to be edited in hepatocellular carcinomas, in positions with surrounding sequence homology to the mooring sequence [83].
More recently, it has become clear that APOBEC1 is expressed by most murine immune cell types [67,84,85], where it targets mostly the 3′ UTRs of tens, if not hundreds of transcripts [66,67]. Apobec-1−/− mice show a neurological dysfunction characterized by a proinflammatory phenotype [86]. In this context microglia are more ‘activated’, and C-to-U editing was shown to be relevant in lysosomal-associated protein-2 (LAMP-2) protein production [86]. This suggests that APOBEC1 RNA editing may play an important role in maintaining brain homeostasis [87]. C-to-U editing was also found to create a supramedullary glycine receptor (GlyRs) isoform, GlyRα3P185L [88]. Neurons do not express Acf or Rbm47, again suggesting the possibility of additional co-factors that are involved in editing [86,89]. Lamp1 and related molecules were shown to be edited in murine macrophages (a myeloid population related to microglia) [86]. C-to-U editing is much more widespread than previously thought. Editing is almost always restricted to 3′ UTRs (with a few exceptions), suggesting functions in the regulation of mRNA stability, localization, translation [66,67]. In addition, because C-to-U editing occurs at the same time as splicing and poly-adenylation [30], and also as APOBEC1 prefers to bind to AU-rich regions [90], the intriguing possibility arises that editing can generate polyadenylation signals in targeted transcripts by generating AAUAAA motifs [91].
Recent work has demonstrated that APOBEC1 editing can create sequence variability within a population, generating different subsets of cells with a distinct complement of edited transcripts [84], something that has also been proposed for ADAR-mediated editing [92]. Additionally, it has also been suggested that APOBEC1 editing on different transcripts belonging to the same pathway can be associated with specific cellular functions [85]. This requires further study, potentially by using targeted editing tools on an APOBEC-deficient background. Such tools are already available for A-to-I editing, reviewed by [93], but not for C-to-U. They will be absolutely necessary for a complete understanding of the role of C-to-U editing on single transcripts, pathways, and cells belonging to different tissues.
4.4. Additional Functions
APOBEC1 prefers binding to AU-rich regions common in 3′ UTRs. It has been proposed that the binding of APOBEC1 to the 3′ UTRs of transcripts such as c-Myc, Tnf-α, and Il-2, which normally exhibit rapid turnover, is sufficient to result in increased mRNA stability in vitro [90] and in vivo [94], even in the absence of editing. Similarly, APOBEC1 can bind and stabilize the cyclooxygenase 2 (COX-2) mRNA in mouse enterocytes, resulting in an increase in prostaglandin E2 (PGE2) synthesis [95]. This effect correlates to the inability of Apobec1−/− mice to properly regenerate small intestinal crypts after radiation injury (which is COX-2 and PGE2 dependent) [95]. Increased PGE2 production was also associated with colon tumor development and progression [96,97]. This matches experimental data where Apobec1 deficiency, in mice genetically predisposed to developed intestinal polyposis (Apcmin/+), results in increased apoptosis and reduced proliferation and tumor burden [94]. Editing was not exhaustively examined, in any of those papers (e.g., by transcriptome or even amplicon deep sequencing). However, these findings suggest that APOBEC1 can scan regions of transcripts that are AU-rich, and either do not contain substantial amounts of C (and are therefore not “editable”), or do not contain those in an editable conformation. Mutants that distinguish APOBEC1 binding vs editing in vivo do not exist (some biochemical findings are available [98]). Evidence of separate binding vs catalysis functions must be gathered with the help of specific function mutants, and the generation of mice carrying Apobec1 alleles with mutations that abolish binding vs catalysis.
APOBEC1 is able to deaminate single strand DNA (ssDNA). This mutagenic activity is cofactor-independent and significantly higher than AID [10,99]. APOBEC1 DNA editing activity was also shown on herpes and hepatitis viral genomes [100,101] and in mammalian cells [11]. In this context, APOBEC1 somatic mutations have a mutational signature which is present in esophageal adenocarcinomas [11]. In addition both AID and APOBEC1 were reported to have 5meC deaminase activity, suggesting a new role as DNA modifiers in cell reprogramming [102].
The question of how cells control APOBEC1 substrate selection (RNA over ssDNA) mitigating the oncogenic potential of APOBEC1 has never been formally addressed. However, it is interesting to note that mutations of RBM47, are linked with carcinogenesis, and it is a potential tumor suppressor gene. Therefore is tempting to speculate that cofactors play an essential role in controlling APOBEC1 substrate selection.
5. APOBEC3
5.1. Key FACTS and Controversies
The A3 family members of the AID/APOBEC family are believed to be a result of a process of gene duplications of the AID/APOBEC1 locus, or the APOBEC2 locus and then divergence and evolution of the genes over millions of years [9]. Humans and non-human primates have the highest number of members of the A3 family, seven (A3A, A3B, A3C, A3D, A3F, A3G, and A3H). Other mammals have fewer (e.g., cats and cattle have three members, whereas mice have only one) [103]. The best-studied functions of the A3s are in innate immune responses to retroviruses and endogenous retroelements, although other types of viruses can also be inhibited by these proteins [104,105,106,107,108,109,110,111,112,113,114]. The burden of retroelements is believed to be responsible for the expansion of A3s from one member in mice, to seven in humans, leading to fewer active retroelements in humans, compared to mice [115,116]. Polymorphisms in human A3s can result in variations in the antiviral/retroviral activity of these enzymes affecting a person’s susceptibility to certain viruses and associated co-morbidities, such as HBC-associated hepatocellular carcinoma [117,118].
The antiviral activity of the A3s was originally defined for A3G; it inhibited HIV in the absence of its viral neutralization partner Vif [119]. After its discovery, A3G created some controversy with conflicting data being published. There was some debate about the relation of the antiviral activity with reverse transcription; some groups claimed that the activity was independent of reverse transcription [120], while others claimed that A3G interacts with and inhibits reverse transcription [121]. The current general consensus is that deamination of the viral genome occurs at the same time as reverse transcription. Other controversies arise, surrounding the packaging of A3G into virons. The importance of A3G packaging and anti-viral activity was not debated but the mechanism and elements involved were debated. Different groups argued that the mechanism for packaging is RNA-independent, and mediated only by an interaction with viral Gag [122,123] versus those arguing that packaging is RNA-dependent [124,125]. The mechanism of the packaging of A3G remains unclear to date.
The prevailing model is that all A3s are more or less associated with the deamination of deoxycytidine to deoxyuridine in ssDNA formed during the viral/retroelement life cycle. This process leads to G to A hypermutations in the viral genome, ultimately reducing viral fitness [126]. It is not entirely clear how the A3s specifically identify their targets, and there are a few insights on how cellular DNA is generally avoided. Most of the A3s (A3G, A3D, A3F, and A3H haplotype II) do not come in contact with cellular DNA, and are maintained in the cytoplasm [127]. A3G binds RNA, and this is believed to be part of the mechanism that maintains its location in the cytoplasm (but not the only mechanism, as it is retained in the cytoplasm even without RNA binding) and inhibits its hypermutation of ssDNA [127,128].
However, not all A3 family members are restricted to the cytoplasm. Specifically, the A3H haplotype II which is cytoplasmic also has RNA binding capabilities, whereas haplotypes I, II, and VI which are all nuclear do not bind RNA [129]. As well, A3C can be found both in the nucleus and in the cytoplasm. A3A is mostly expressed in monocytes/macrophages, and under normal circumstances, it is cytoplasmic and therefore not genotoxic. However, when it is expressed/overexpressed in other cell types, it can also be found in the nucleus [130]. This suggests that some factor in the monocytes/macrophages that is generally absent in other cells is actively maintaining A3A in the cytoplasm. The human Tribbles 3 protein, a pseudokinase, could be such a candidate, as it interacts with both A3A and A3C and was found to decrease A3A editing of nuclear DNA in a concentration-dependent manner, thus protecting genetic integrity [131]. Finally, A3B has a strong NLS, resulting in only nuclear expression [132]. A3B genotoxicity seems to be mitigated under normal conditions by very low expression, although what exactly regulates its expression is yet unknown.
5.2. Additional Functions and Disease Associations
5.2.1. DNA Mutation (Kataegis)
Despite careful control of A3 expression and localization, sometimes misregulation occurs, and this is associated with cancer. In particular, A3B and A3A are found to be overexpressed in some cancers and/or are associated with kataegis mutation (literally describing “rainfalls” of C-to-T mutation, particularly at promoter regions [133]. This has since been termed mutational signature 2 and 13 in these cancers [134,135,136,137,138,139,140,141,142]. A polymorphism where A3B is deleted and its 3′UTR replaces a deleted A3A 3′UTR results in higher levels of A3A and an increase in cancer risk in Asian populations [143]. It is unknown whether these genes are cancer drivers or a downstream consequence of other cancer driver genes. They do contribute to genetic instability and heterogeneity with important consequences to prognosis (which can be both positive and negative, depending on the cancer type) and cancer therapy [144,145,146]. A3G has also been associated with a negative prognosis in colon carcinoma patients with hepatic metastasis [147,148] and in T cells, it was associated with increased tumor infiltration and a positive prognosis [149]. It is also a target for cancer therapy, as it enhances tumor resistance to radiotherapy [150,151]. The mechanism of action of A3G in cancer remains unclear, and the mechanism appears to be unrelated to DNA deamination; however, in the case of resistance to radiotherapy, it is believed to promote DNA repair in a deaminase-dependent manner [147,148,149,150,151].
5.2.2. RNA Editing
Traditionally, the A3s were considered to work only on ssDNA, and to bind RNA in a non-specific and non-selective manner [152]. However, in 2015, an RNA editing function for A3A was described in human monocytes/macrophages [17]. Editing levels were affected by hypoxia and the polarization of the macrophages [17] and transient ectopic overexpression of A3A in HEK293Ts also resulted in RNA editing of a large number of genes [153]. Later the same group went on to describe the RNA editing capabilities for A3G, again in the HEK293T cell system [18]. They also suggest that there is an increase in RNA editing as a response to mitochondrial stress in NK cells and lymphocytes [154].
Whereas most of the work on A3s and RNA editing involved overexpression, it remains clear that many of the A3s do bind RNA (reviewed in [155]). C-to-U editing in 3′ UTRs is readily apparent in transcriptome data from human cells or tissues (e.g., macrophages and others), where APOBEC1 is not expressed. A3A has been found to be associated with the editing of similar sets of transcripts as APOBEC1 in mouse macrophages [85], an activity that was upregulated upon interferon stimulation [156]. Thus, while the relevance of editing remains unclear in the context of the A3 family, it remains largely unexamined. Since APOBEC1 is absent in most human tissues, including macrophages and other immune cells, it is possible that A3A (and perhaps additional family members) have adapted and evolved to carry out similar roles in humans. In that regard, it would be interesting to assess the known SNPs and polymorphisms in A3s (especially SNPs that have not been associated with an antiviral/anti-retroviral function, or a decrease in such functions) in the context of their RNA editing capabilities. Such potential differences in RNA editing could contribute to the complex differences in immune responses in different individuals, and susceptibility to the development of autoimmune diseases.
Interestingly, A3s are also considered to have an antiviral activity that is deaminase-independent. In double deaminase domain A3s, a mutation in the non-catalytically active domain results in a reduction in antiviral activity [157,158,159,160]. The mechanism of this deaminase independent activity is unknown with some suggesting that RNA binding is important for this process [158], and as previously mentioned, RNA binding inhibits ssDNA deamination [127,128]. In the case of A3A and A3G, where RNA editing has been described, it may be worthy to consider that this ‘deaminase independent’ activity may in fact be DNA deaminase-independent and be a result of the RNA editing of cellular transcripts that by some mechanism increases the intrinsic cellular immunity or RNA editing of the viruses/retroelements. Much more research into these links and the functions of A3 mediated RNA editing is necessary.
Overall, very little is known about C-to-U RNA editing, especially in humans, and much more interest and effort are required to begin to understand its in vivo consequences. It is certainly worthwhile to re-evaluate the A3s’ DNA editing abilities, and to consider other A3s for possible RNA editing functions. Most of the inquiry into the functions of the A3s has involved overexpression studies where proteins are then found in levels that are higher than possible in nature, and activities and targets that may not occur in vivo may be observed. Often, the A3s are studied as a tagged fusion protein; while in some cases, no differences in DNA deamination can be seen, in others, such as human and rhesus monkey A3A a C-terminal tag, greatly reduced anti-viral activity against SHIV [161]. Therefore, this group of proteins should be studied further in their native form under native expression conditions to determine whether humans have evolved a host of different RNA editors. In that context, it is interesting to note that DND1 can also interact with APOBEC3 in mice, with inhibitory effects on miRNA functions [72,78]. In humans A3G appears to be able to interact with HNRNPC1, HNRNPAB, SYNCRIP, and HNRNPR [162,163]; most of these proteins are involved in C-to-U RNA editing and/or related to ACF [164]. Considering the potential role of A3G and A3A in RNA editing [18], it is tempting to speculate that different RBPs, in different tissues, could work as cofactors in bringing the A3s to their specific targets (and consequently, it remains possible that APOBEC1 and A3s contain a conserved region for the interaction with these RBPs). Overall, it remains to be determined if A3A and A3G have RBP interaction partners like ACF and RBM47 (the APOBEC1 interactors) that are a part of the human RNA editing machinery, and what, if anything, determines the specificity of the targets between the proteins and the different cell types.
6. Human Diseases Related to C-to-U Editing
Whereas it is clear that C-to-U editing occurs with reasonable frequency, because APOBEC1 expression is thought to be tissue-restricted in humans, most of the disease burden must be associated with A3 RNA editors, and these have not been extensively studied, as noted above. At the same time, whereas all AID/APOBEC family members were originally characterized exclusively, either as DNA mutators or RNA editors, this distinction is not supported either by structural data (all active sites appear near-identical—reviewed in [14,15]) or by cell biological findings (where APOBEC1 can function as a robust DNA mutator [11]). Because most deaminases can edit both DNA and RNA, it is important to keep this in mind as we assess the relevance of RNA editing to specific diseases.
Even though APOBEC1 is not supposed to be expressed outside the human intestinal tract, all genetic dysfunctions recently ascribed to APOBEC1 are within the human brain. GlyR editing is important in disease progression of temporal lobe epilepsy (TLE) [165]. Specifically, GlyR editing was shown to be increased in the hippocampus of patients with pharmacoresistant temporal lobe epilepsy (TLE) [165]. RNA-edited GlyRs are expressed at the presynaptic terminals of hippocampal neurons [166], and even small differences in editing can induce dysfunction (probably because the low levels of editing identified in bulk represent editing of GlyR within specific subsets of neurons) [166,167]. At the same time, it was shown that APOBEC1 is able to edit the GlyR mRNA in vitro. Whereas this is only suggestive (as noted above), recent bioinformatic analyses have identified two genetic variants of APOBEC1, 80M and 80I, which are associated with GlyR editing levels [168]. Further characterization of patients with intractable TLE (iTLE) for APOBEC1 dimorphism, showed that patients with 80I variant suffer from simple or complex seizures, whereas patients with 80M exhibit neurodegeneration and secondary generalized seizure activity [168]. The I80M polymorphism in human APOBEC1 was reported previously in the small intestine, with no effect on Apob RNA editing [169]. The identification of the APOBEC1 80I/M-coding allele as a new genetic risk factor for iTLE patients is the first example of a direct human disease association between APOBEC1 and a tissue other than the small intestine. Similar studies must be carried out with the A3 family, as well as disease entities that APOBEC1 and A3 are involved in.
In the nervous system, C-to-U editing of the transcript encoding tryptophan hydroxylase 2 (TPH2a) can lead to alternative splicing. In the inserted exon, 3b, there is a C-to-U editing event that leads to a recoding mutation (Q129X substitution) and a truncated protein [170]. Editing at this position decreased substantially (by 50% and 30%) in suicide-prone and schizophrenia patients, respectively [170]. To date, the responsible C-to-U editing enzyme remains unknown.
Mutations in the APOBEC1 cofactors are also associated with disease. Mutations in RBM47 have been associated with breast cancer progression, and specifically with an increase in the fitness of certain cancer clones and increased metastatic potential [171]. In addition, expression of RBM47 correlates with a good prognosis in patients with lung, breast, and gastric cancer suggesting a tumor-suppressive role [172]. RBM47 robustly binds around 2500 transcripts, but mRNA editing has not been assessed in these studies [171]. It would be important to assess whether the loss of the RNA editing function in some transcripts due to loss of the editing co-factor is directly responsible for disease progression. In support of this hypothesis, it is important to note that some targets identified as RBM47-bound RNA [171] were also identified as APOBEC1 RNA editing targets or as APOBEC1 RNA interactors, such as β-2 microglobulin (B2m) and interleukin-8 (IL8) [173], respectively.
Polymorphisms in A3 RNA editors could affect human disease progression by influencing immune responses or through changes in RNA editing levels or targets. In autoimmune diseases, systemic lupus erythematosus (SLE) patients often have high levels of circulating type I interferon, and elevated expression of interferon-stimulated genes, which include various A3s [174,175]. These SLE patients can also exhibit increased levels of A-to-I (by adenosine deaminases) and C-to-U RNA editing [176]. This increased level of editing can result in amino acid recoding and the development of potential MHC class I epitopes, which are possible contributors to disease development [176].
The RNA editing of A3A and A3G appears to be more scrupulous than their DNA editing with a more specific sequence signature [177]. Where DNA editing by A3A and A3G appears to be generally non-specific with only the dinucleotide [T/C]C as a preference, RNA editing was more likely to be observed in stem-loop structures, with the target C contained in the loop; the more stable the stem, the higher the levels of editing [177]. The rs172378 C1q synonymous SNP associated with nephritis in SLE patients [178,179], has increased C-to-U editing within three nucleotides from the polymorphism, likely by altering the RNA secondary structure stabilizing a stem-loop [177]. This type of SNP affects how RNA may be targeted by the A3s. While it is unknown whether this change directly contributes to SLE, as with other RNA editing events, it could contribute to the transcript fate and subsequently to the outcome of the protein, and thus contribute to the disease. Likewise, it is possible that other such SNPs, including synonymous ones, could result in changes in RNA editing affecting protein diversity.
A3A and A3G have both also been implicated to play various roles in cancers. It stands to reason that the question of what the additional involvement of RNA editing by these editors in these cancers should be addressed. RNA editing may also account for some of the unclear mechanisms of the involvement of A3G in cancer.
7. Conclusions
Research in the field of RNA editing has really taken off, with the advent of NGS technologies [17,18,69], which have demonstrated that C-to-U editing in human is not limited to APOBEC1, but can also be catalyzed by APOBEC3 family enzymes. In that context, it is important to note the discovery of RBM47 as an APOBEC1 co-factor even for Apob editing [63]. Additionally, recent work has indicated that co-factors, which are crucial to targeting the editing enzyme to the right transcript, might have distinct transcript preferences. Future work will need to clarify the identity and role of co-factors for APOBEC3 editases, and further clarify the role of co-factors other than ACF and RBM47 for APOBEC1 editing.
Author Contributions
T.L. and R.P. contributed to writing of the original draft; F.N.P. was involved in the review and editing of the draft, together with the other two authors.
Funding
Research in the Papavasiliou lab is funded by ERC grant #649019 (RNAEDIT). We also acknowledge funding from COST action CA16120 (EPITRAN).
Acknowledgments
The authors would like to acknowledge Erec Stebbins for critical reading of the manuscript, and Piero Sisti for help with the production of the graphical abstract.
Conflicts of Interest
The authors declare no conflict of interest, and the funders had no role in the writing of the manuscript.
References
- Boccaletto, P.; MacHnicka, M.A.; Purta, E.; Pitkowski, P.; Baginski, B.; Wirecki, T.K.; De Crécy-Lagard, V.; Ross, R.; Limbach, P.A.; Kotter, A.; et al. MODOMICS: A database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2018, 46. [Google Scholar] [CrossRef] [PubMed]
- Frye, M.; Harada, B.T.; Behm, M.; He, C. RNA modifications modulate gene expression during development. Science 2018. [Google Scholar] [CrossRef] [PubMed]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA modifications in gene expression regulation. Cell 2017, 169, 1187–1200. [Google Scholar] [CrossRef] [PubMed]
- Benne, R.; Van Den Burg, J.; Brakenhoff, J.P.J.; Sloof, P.; Van Boom, J.H.; Tromp, M.C. Major transcript of the frameshifted coxll gene from trypanosome mitochondria contains four nucleotides that are not encoded in the DNA. Cell 1986, 46, 819–826. [Google Scholar] [CrossRef]
- Teng, B.; Burant, C.; Davidson, N. Molecular cloning of an apolipoprotein B messenger RNA editing protein. Science 1993, 260, 1816–1820. [Google Scholar] [CrossRef] [PubMed]
- Navaratnam, N.; Morrison, J.R.; Bhattacharya, S.; Patel, D.; Funahashi, T.; Giannoni, F.; Teng, B.B.; Davidson, N.O.; Scott, J. The p27 catalytic subunit of the apolipoprotein B mRNA editing enzyme is a cytidine deaminase. J. Biol. Chem. 1993, 268, 20709–20712. [Google Scholar] [PubMed]
- Muramatsu, M.; Kinoshita, K.; Fagarasan, S.; Yamada, S.; Shinkai, Y.; Honjo, T. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell 2000. [Google Scholar] [CrossRef]
- Conticello, S.G. The AID/APOBEC family of nucleic acid mutators. Genome Biol. 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Conticello, S.G.; Thomas, C.J.F.; Petersen-Mahrt, S.K.; Neuberger, M.S. Evolution of the AID/APOBEC family of polynucleotide (deoxy)cytidine deaminases. Mol. Biol. Evol. 2005, 22, 367–377. [Google Scholar] [CrossRef]
- Harris, R.S.; Petersen-Mahrt, S.K.; Neuberger, M.S. RNA editing enzyme APOBEC1 and some of its homologs can act as DNA mutators. Mol. Cell 2002. [Google Scholar] [CrossRef]
- Saraconi, G.; Severi, F.; Sala, C.; Mattiuz, G.; Conticello, S.G. The RNA editing enzyme APOBEC1 induces somatic mutations and a compatible mutational signature is present in esophageal adenocarcinomas. Genome Biol. 2014. [Google Scholar] [CrossRef]
- Lau, P.P.; Zhu, H.J.; Baldini, A.; Charnsangavej, C.; Chan, L. Dimeric structure of a human apolipoprotein B mRNA editing protein and cloning and chromosomal localization of its gene. Proc. Natl. Acad. Sci. USA 1994, 91, 8522–8526. [Google Scholar] [CrossRef] [PubMed]
- Chieca, M.; Montini, M.; Severi, F.; Pecori, R.; Silvestro, G. Dimerisation of APOBEC1 is dispensable for its RNA editing activity. Biorxiv 2018. [Google Scholar] [CrossRef]
- Salter, J.D.; Smith, H.C. Modeling the embrace of a mutator: APOBEC selection of nucleic acid ligands. Trends Biochem. Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Salter, J.D.; Bennett, R.P.; Smith, H.C. The APOBEC Protein Family: United by Structure, Divergent in Function. Trends Biochem. Sci. 2016, 41, 578–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarmuz, A.; Chester, A.; Bayliss, J.; Gisbourne, J.; Dunham, I.; Scott, J.; Navaratnam, N. An anthropoid-specific locus of orphan C to U RNA-editing enzymes on chromosome 22. Genomics 2002, 79, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Patnaik, S.K.; Thomas Taggart, R.; Kannisto, E.D.; Enriquez, S.M.; Gollnick, P.; Baysal, B.E. APOBEC3A cytidine deaminase induces RNA editing in monocytes and macrophages. Nat. Commun. 2015, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Patnaik, S.K.; Taggart, R.T.; Baysal, B.E. The double-domain cytidine deaminase APOBEC3G is a cellular site-specific RNA editing enzyme. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Young, S.G. Recent progress in understanding apolipoprotein B. Circulation 1990. [Google Scholar] [CrossRef]
- Yao, Z.; McLeod, R.S. Synthesis and secretion of hepatic apolipoprotein B-containing lipoproteins. Biochim. Biophys. Acta Lipids Lipid Metab. 1994, 1212, 152–166. [Google Scholar] [CrossRef]
- Greeve, J.; Altkemper, I.; Dieterich, J.H.; Greten, H.; Windler, E. Apolipoprotein B mRNA editing in 12 different mammalian species: Hepatic expression is reflected in low concentrations of apoB-containing plasma lipoproteins. J. Lipid Res. 1993, 34, 1367–1383. [Google Scholar] [PubMed]
- Yamanaka, S.; Balestra, M.E.; Ferrell, L.D.; Fan, J.; Arnold, K.S.; Taylor, S.; Taylor, J.M.; Innerarity, T.L. Apolipoprotein B mRNA-editing protein induces hepatocellular carcinoma and dysplasia in transgenic animals. Proc. Natl. Acad. Sci. USA 1995, 92, 8483–8487. [Google Scholar] [CrossRef] [PubMed]
- Nakamuta, M.; Chang, B.H.; Zsigmond, E.; Kobayashi, K.; Lei, H.; Ishida, B.Y.; Oka, K.; Li, E.; Chan, L. Complete phenotypic characterization of apobec-1 knockout mice with a wild-type genetic background and a human apolipoprotein B transgenic background, and restoration of apolipoprotein B mRNA editing by somatic gene transfer of Apobec-1. J. Biol. 1996, 271, 25981–25988. [Google Scholar]
- Hirano, K.I.; Young, S.G.; Farese, R.V.; Ng, J.; Sande, E.; Warburton, C.; Powell-Braxton, L.M.; Davidson, N.O. Targeted disruption of the mouse apobec-1 gene abolishes apolipoprotein B mRNA editing and eliminates apolipoprotein B48. J. Biol. Chem. 1996. [Google Scholar] [CrossRef]
- Morrison, J.R.; Pászty, C.; Stevens, M.E.; Hughes, S.D.; Forte, T.; Scott, J.; Rubin, E.M. Apolipoprotein B RNA editing enzyme-deficient mice are viable despite alterations in lipoprotein metabolism. Proc. Natl. Acad. Sci. USA 1996, 93, 7154–7159. [Google Scholar] [CrossRef] [PubMed]
- Giannoni, F.; Bonen, D.K.; Funahashi, T.; Hadjiagapiou, C.; Burant, C.F.; Davidson, N.O. Complementation of apolipoprotein B mRNA editing by human liver accompanied by secretion of apolipoprotein B48. J. Biol. Chem. 1994, 269, 5932–5936. [Google Scholar] [PubMed]
- Anant, S.; MacGinnitie, A.J.; Davidson, N.O. apobec-1, the catalytic subunit of the mammalian apolipoprotein B mRNA editing enzyme, is a novel RNA-binding protein. J. Biol. Chem. 1995, 270, 14762–14767. [Google Scholar] [CrossRef]
- Mehta, A.; Kinter, M.T.; Sherman, N.E.; Driscoll, D.M. Molecular cloning of apobec-1 complementation factor, a novel RNA-binding protein involved in the editing of apolipoprotein B mRNA. Mol. Cell. Biol. 2000. [Google Scholar] [CrossRef]
- Lellek, H.; Kirsten, R.; Diehl, I.; Apostel, F.; Buck, F.; Greeve, J. Purification and molecular cloning of a novel essential component of the apolipoprotein B mRNA editing enzyme-complex. J. Biol. Chem. 2000. [Google Scholar] [CrossRef]
- Lau, P.P.; Xiong, W.; Zhu, H.J.; Chen, S.H.; Chan, L. Apolipoprotein B mRNA editing is an intranuclear event that occurs posttranscriptionally coincident with splicing and polyadenylation. J. Biol. Chem. 1991, 266, 20550–20554. [Google Scholar]
- Sowden, M.; Hamm, J.K.; Spinelli, S.; Smith1, H.C.; Smith, H.C. Determinants involved in regulating the proportion of edited apolipoprotein B RNAs. Rna 1996, 2, 274–288. [Google Scholar] [PubMed]
- Chester, A.; Somasekaram, A.; Tzimina, M.; Jarmuz, A.; Gisbourne, J.; O’Keefe, R.; Scott, J.; Navaratnam, N. The apolipoprotein B mRNA editing complex performs a multifunctional cycle and suppresses nonsense-mediated decay. EMBO J. 2003. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sowden, M.P.; Yang, Y.; Smith, H.C. Intracellular trafficking determinants in APOBEC-1, the catalytic subunit for cytidine to uridine editing of apolipoprotein B mRNA. Exp. Cell Res. 2001. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Smith, H.C. Multiple protein domains determine the cell type-specific nuclear distribution of the catalytic subunit required for apolipoprotein B mRNA editing. Proc. Natl. Acad. Sci. USA 1997. [Google Scholar] [CrossRef]
- Blanc, V.; Kennedy, S.; Davidson, N.O. A novel nuclear localization signal in the auxiliary domain of Apobec-1 complementation factor regulates nucleocytoplasmic import and shuttling. J. Biol. Chem. 2003, 278, 41198–41204. [Google Scholar] [CrossRef] [PubMed]
- Sowden, M.P.; Ballatori, N.; Jensen, K.L.D.M.; Reed, L.H.; Smith, H.C. The editosome for cytidine to uridine mRNA editing has a native complexity of 27S: Identification of intracellular domains containing active and inactive editing factors. J. Cell Sci. 2002, 115, 1027–1039. [Google Scholar] [PubMed]
- Oka, K.; Kobayashi, K.; Sullivan, M.; Martinez, J.; Teng, B.B.; Ishimura-Oka, K.; Chan, L. Tissue-specific inhibition of apolipoprotein B mRNA editing in the liver by adenovirus-mediated transfer of a dominant negative mutant APOBEC-1 leads to increased low density lipoprotein in mice. J. Biol. Chem. 1997. [Google Scholar] [CrossRef]
- Anant, S.; Henderson, J.O.; Mukhopadhyay, D.; Navaratnam, N.; Kennedy, S.; Min, J.; Davidson, N.O. Novel role for RNA-binding protein CUGBP2 in mammalian RNA editing: CUGBP2 modulates C to U editing of apolipoprotein B mRNA by interacting with apobec-1 and ACF, the apobec-1 complementation factor. J. Biol. Chem. 2001. [Google Scholar] [CrossRef]
- Lau, P.P.; Villanueva, H.; Kobayashi, K.; Nakamuta, M.; Chang, B.H.J.; Chan, L. A DnaJ Protein, Apobec-1-binding Protein-2, Modulates Apolipoprotein B mRNA Editing. J. Biol. Chem. 2001. [Google Scholar] [CrossRef]
- Lau, P.P.; Chan, L. Involvement of a Chaperone Regulator, Bcl2-associated Athanogene-4, in Apolipoprotein B mRNA Editing. J. Biol. Chem. 2003. [Google Scholar] [CrossRef]
- Greeve, J.; Lellek, H.; Rautenberg, P.; Greten, H. Inhibition of the apolipoprotein B mRNA editing enzyme-complex by hnRNP C1 protein and 40S hnRNP complexes. Biol. Chem. 1998. [Google Scholar] [CrossRef]
- Blanc, V.; Navaratnam, N.; Henderson, J.O.; Anant, S.; Kennedy, S.; Jarmuz, A.; Scott, J.; Davidson, N.O. Identification of GRY-RBP as an apolipoprotein B RNA-binding protein that interacts with both Apobec-1 and Apobec-1 complementation factor to modulate C to U editing. J. Biol. Chem. 2001. [Google Scholar] [CrossRef]
- Chen, Z.; Eggerman, T.L.; Patterson, A.P. ApoB mRNA editing is mediated by a coordinated modulation of multiple apoB mRNA editing enzyme components. Am. J. Physiol. Liver Physiol. 2007. [Google Scholar] [CrossRef]
- Navaratnam, N.; Fujino, T.; Bayliss, J.; Jarmuz, A.; How, A.; Richardson, N.; Somasekaram, A.; Bhattacharya, S.; Carter, C.; Scott, J. Escherichia coli cytidine deaminase provides a molecular model for ApoB RNA editing and a mechanism for RNA substrate recognition. J. Mol. Biol. 1998. [Google Scholar] [CrossRef] [PubMed]
- Teng, B.B.; Ochsner, S.; Zhang, Q.; Soman, K.V; Lau, P.P.; Chan, L. Mutational analysis of apolipoprotein B mRNA editing enzyme (APOBEC1): structure–function relationships of RNA editing and dimerization. J. Lipid Res. 1999. [Google Scholar] [CrossRef]
- Blanc, V.; Davidson, N.O. C-to-U RNA editing: Mechanisms leading to genetic diversity. J. Biol. Chem. 2003, 278, 1395–1398. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, D.M.; Lakhe-Reddy, S.; Oleksa, L.M.; Martinez, D. Induction of RNA editing at heterologous sites by sequences in apolipoprotein B mRNA. Mol. Cell. Biol. 1993, 13, 7288–7294. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.R.; Knott, T.J.; Legros, J.E.; Navaratnam, N.; Greeve, J.C.; Scott, J. Sequence requirements for the editing of apolipoprotein B mRNA. J. Biol. Chem. 1991, 266, 16301–16304. [Google Scholar] [PubMed]
- Backus, J.W.; Smith, H.C. 3 Distinct RNA sequence elements are required for efficient apolipoprotein-B (apoB) RNA editing in vitro. Nucleic Acids Res 1992, 20, 6007–6014. [Google Scholar] [CrossRef] [PubMed]
- Maris, C.; Masse, J.; Chester, A.N.N.; Navaratnam, N.; Allain, F.H. NMR structure of the apoB mRNA stem-loop and its interaction with the C to U editing APOBEC1 complementary factor NMR structure of the apoB mRNA stem-loop and its interaction with the C to U editing APOBEC1 complementary factor. RNA 2005, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Hersberger, M.; Patarroyo-white, S.; Arnold, K.S.; Innerarity, T.L. Phylogenetic analysis of the apolipoprotein B mRNA-editing region. J. Biol. Chem. 1999, 274, 34590–34597. [Google Scholar] [CrossRef] [PubMed]
- Richardson, N.; Navaratnam, N.; Scott, J. Secondary structure for the apolipoprotein B mRNA editing site. J.Biol.Chem. 1998, 273, 31707–31717. [Google Scholar] [CrossRef] [PubMed]
- Backus, J.W.; Schock, D.; Smith, H.C. Only cytidines 5′ of the apolipoprotein B mRNA mooring sequence are edited. BBA Gene Struct. Expr. 1994. [Google Scholar] [CrossRef]
- Mehta, A.; Driscoll, D.M. Identification of domains in apobec-1 complementation factor required for RNA binding and apolipoprotein-B mRNA editing. RNA 2002, 8, 69–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, V.; Henderson, J.O.; Kennedy, S.; Davidson, N.O. Mutagenesis of apobec-1 complementation factor reveals distinct domains that modulate RNA binding, protein-protein interaction with apobec-1, and complementation of C to U RNA-editing activity. J. Biol. Chem. 2001. [Google Scholar] [CrossRef]
- Nakamuta, M.; Oka, K.; Krushkal, J.; Kobayashi, K.; Yamamoto, M.; Li, W.H.; Chan, L. Alternative mRNA splicing and differential promoter utilization determine tissue-specific expression of the apolipoprotein B mRNA-editing protein (apobec1) gene in mice: Structure and evolution of apobec1 and related nucleoside/nucleotide deaminases. J. Biol. Chem. 1995, 270, 13042–13056. [Google Scholar] [CrossRef]
- Dance, G.S.; Sowden, M.P.; Cartegni, L.; Cooper, E.; Krainer, A.R.; Smith, H.C. Two proteins essential for apolipoprotein B mRNA editing are expressed from a single gene through alternative splicing. J. Biol. Chem. 2002. [Google Scholar] [CrossRef]
- Lee, R.M.; Hirano, K.; Anant, S.; Baunoch, D.; Davidson, N.O. An alternatively spliced form of apobec-1 messenger RNA is overexpressed in human colon cancer. Gastroenterology 1998, 115, 1096–1103. [Google Scholar] [CrossRef]
- Sowden, M.P.; Lehmann, D.M.; Lin, X.; Smith, C.O.; Smith, H.C. Identification of novel alternative splice variants of APOBEC-1 complementation factor with different capacities to support apolipoprotein B mRNA editing. J. Biol. Chem. 2004. [Google Scholar] [CrossRef]
- Lehmann, D.M.; Galloway, C.A.; Sowden, M.P.; Smith, H.C. Metabolic regulation of ApoB mRNA editing is associated with phosphorylation of APOBEC-1 complementation factor. Nucleic Acids Res. 2006. [Google Scholar] [CrossRef]
- Lehmann, D.M.; Galloway, C.A.; MacElrevey, C.; Sowden, M.P.; Wedekind, J.E.; Smith, H.C. Functional characterization of APOBEC-1 complementation factor phosphorylation sites. Biochim. Biophys. Acta Mol. Cell Res. 2007. [Google Scholar] [CrossRef] [PubMed]
- Galloway, C.A.; Ashton, J.; Sparks, J.D.; Mooney, R.A.; Smith, H.C. Metabolic regulation of APOBEC-1 Complementation Factor trafficking in mouse models of obesity and its positive correlation with the expression of ApoB protein in hepatocytes. Biochim. Biophys. Acta Mol. Basis Dis. 2010. [Google Scholar] [CrossRef] [PubMed]
- Fossat, N.; Tourle, K.; Radziewic, T.; Barratt, K.; Liebhold, D.; Studdert, J.B.; Power, M.; Jones, V.; Loebel, D.A.F.; Tam, P.P.L. C to U RNA editing mediated by APOBEC 1 requires RNA-binding protein RBM47. EMBO Rep. 2014, 15, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Snyder, E.M.; McCarty, C.; Mehalow, A.; Svenson, K.L.; Murray, S.A.; Korstanje, R.; Braun, R.E. APOBEC1 complementation factor (A1CF) is dispensable for C-to-U RNA editing in vivo. RNA 2017, 23. [Google Scholar] [CrossRef] [PubMed]
- Blanc, V.; Henderson, J.O.; Newberry, E.P.; Kennedy, S.; Luo, J.; Davidson, N.O. Targeted deletion of the murine apobec-1 complementation factor (acf) gene results in embryonic lethality. Mol. Cell. Biol. 2005. [Google Scholar] [CrossRef]
- Rosenberg, B.R.; Hamilton, C.E.; Mwangi, M.M.; Dewell, S.; Papavasiliou, F.N. Transcriptome-wide sequencing reveals numerous APOBEC1 mRNA-editing targets in transcript 3′ UTRs. Nat. Struct. Mol. Biol. 2011. [Google Scholar] [CrossRef]
- Blanc, V.; Park, E.; Schaefer, S.; Miller, M.; Lin, Y.; Kennedy, S.; Billing, A.M.; Hamidane, H.B.; Graumann, J.; Mortazavi, A.; et al. Genome-wide identification and functional analysis of Apobec-1-mediated C-to-U RNA editing in mouse small intestine and liver. Genome Biol. 2014. [Google Scholar] [CrossRef]
- Blanc, V.; Sessa, K.J.; Kennedy, S.; Luo, J.; Davidson, N.O. Apobec-1 complementation factor modulates liver regeneration by post-transcriptional regulation of interleukin-6 mRNA stability. J. Biol. Chem. 2010. [Google Scholar] [CrossRef]
- Blanc, V.; Xie, Y.; Kennedy, S.; Riordan, J.D.; Rubin, D.C.; Blair, B.; Mills, J.C.; Nadeau, J.H.; Davidson, N.O. APOBEC1 complementation factor (A1CF) and RBM47 interact in tissue-specific regulation of C to U RNA editing in mouse intestine and liver. RNA 2018. [Google Scholar] [CrossRef]
- Greeve, J.; Navaratnam, N.; Scott, J. Characterization of the apolipoprotein B mRNA editing enzyme: No similarity to the proposed mechanism of RNA editing in kinetoplastid protozoa. Nucleic Acids Res. 1991. [Google Scholar] [CrossRef]
- Chester, A.; Weinreb, V.; Carter, C.W.; Navaratnam, N. Optimization of apolipoprotein B mRNA editing by APOBEC1 apoenzyme and the role of its auxiliary factor, ACF. RNA 2004. [Google Scholar] [CrossRef]
- Bhattacharya, C.; Aggarwal, S.; Kumar, M.; Ali, A.; Matin, A. Mouse apolipoprotein B editing complex 3 (APOBEC3) is expressed in germ cells and interacts with dead-end (DND1). PLoS ONE 2008, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Youngren, K.K.; Coveney, D.; Peng, X.; Bhattacharya, C.; Schmidt, L.S.; Nickerson, M.L.; Lamb, B.T.; Deng, J.M.; Behringer, R.R.; Capel, B.; et al. The Ter mutation in the dead end gene causes germ cell loss and testicular germ cell tumours. Nature 2005. [Google Scholar] [CrossRef] [PubMed]
- Nelson, V.R.; Heaney, J.D.; Tesar, P.J.; Davidson, N.O.; Nadeau, J.H. Transgenerational epigenetic effects of the Apobec1 cytidine deaminase deficiency on testicular germ cell tumor susceptibility and embryonic viability. Proc. Natl. Acad. Sci. USA 2012, 109, E2766–E2773. [Google Scholar] [CrossRef] [PubMed]
- Fossat, N.; Tam, P.P.L. Re-editing the paradigm of Cytidine (C) to Uridine (U) RNA editing. RNA Biol. 2014, 11, 1233–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez, D.; Freese, P.; Alexis, M.S.; Su, A.; Hochman, M.; Palden, T.; Bazile, C.; Lambert, N.J.; Van Nostrand, E.L.; Pratt, G.A.; et al. Sequence, structure, and context preferences of human RNA binding proteins. Mol. Cell 2018. [Google Scholar] [CrossRef]
- Kedde, M.; Strasser, M.J.; Boldajipour, B.; Vrielink, J.A.F.O.; Slanchev, K.; le Sage, C.; Nagel, R.; Voorhoeve, P.M.; van Duijse, J.; Ørom, U.A.; et al. RNA-binding protein Dnd1 inhibits microRNA access to target mRNA. Cell 2007, 131, 1273–1286. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Karki, N.; Bhattacharya, C.; Zhu, R.; MacDuff, D.A.; Stenglein, M.D.; Schumacher, A.J.; Demorest, Z.L.; Harris, R.S.; Matin, A.; et al. APOBEC3 inhibits DEAD-END function to regulate microRNA activity. BMC Mol. Biol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Powell-Braxton, L.; Véniant, M.; Latvala, R.D.; Hirano, K.I.; Won, W.B.; Ross, J.; Dybdal, N.; Zlot, C.H.; Young, S.G.; Davidson, N.O. A mouse model of human familial hypercholesterolemia: Markedly elevated low density lipoprotein cholesterol levels and severe atherosclerosis on a low-fat chow diet. Nat. Med. 1998. [Google Scholar] [CrossRef]
- Xie, Y.; Luo, J.; Kennedy, S.; Davidson, N.O. Conditional intestinal lipotoxicity in Apobec-1-/-Mttp-IKO mice: A survival advantage for mammalian intestinal apolipoprotein B mRNA editing. J. Biol. Chem. 2007, 282, 33043–33051. [Google Scholar] [CrossRef]
- Skuse, G.R.; Cappione, A.J.; Sowden, M.; Metheny, L.J.; Smith, H.C. The neurofibromatosis type I messenger RNA undergoes base-modification RNA editing. Nucleic Acids Res. 1996, 24, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, D.; Anant, S.; Lee, R.M.; Kennedy, S.; Viskochil, D.; Davidson, N.O. C→U editing of neurofibromatosis 1 mRNA occurs in tumors that express both the type II transcript and apobec-1, the catalytic subunit of the apolipoprotein B mRNA–editing enzyme. Am. J. Hum. Genet. 2002, 70, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, S.; Poksay, K.S.; Arnold, K.S.; Innerarity, T.L. A novel translational repressor mRNA is edited extensively in livers containing tumors caused by the transgene expression of the apoB mRNA-editing enzyme. Genes Dev. 1997, 11, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Harjanto, D.; Papamarkou, T.; Oates, C.J.; Rayon-Estrada, V.; Papavasiliou, F.N.; Papavasiliou, A. RNA editing generates cellular subsets with diverse sequence within populations. Nat. Commun. 2016. [Google Scholar] [CrossRef] [PubMed]
- Rayon-Estrada, V.; Harjanto, D.; Hamilton, C.E.; Berchiche, Y.A.; Gantman, E.C.; Sakmar, T.P.; Bulloch, K.; Gagnidze, K.; Harroch, S.; McEwen, B.S.; et al. Epitranscriptomic profiling across cell types reveals associations between APOBEC1-mediated RNA editing, gene expression outcomes, and cellular function. Proc. Natl. Acad. Sci. USA 2017. [Google Scholar] [CrossRef] [PubMed]
- Cole, D.C.; Chung, Y.; Gagnidze, K.; Hajdarovic, K.H.; Rayon-Estrada, V.; Harjanto, D.; Bigio, B.; Gal-Toth, J.; Milner, T.A.; McEwen, B.S.; et al. Loss of APOBEC1 RNA-editing function in microglia exacerbates age-related CNS pathophysiology. Proc. Natl. Acad. Sci. USA 2017. [Google Scholar] [CrossRef] [PubMed]
- Gagnidze, K.; Rayon-Estrada, V.; Harroch, S.; Bulloch, K.; Papavasiliou, F.N. A new chapter in genetic medicine: RNA editing and its role in disease pathogenesis. Trends Mol. Med. 2018. [Google Scholar] [CrossRef]
- Meier, J.C.; Henneberger, C.; Melnick, I.; Racca, C.; Harvey, R.J.; Heinemann, U.; Schmieden, V.; Grantyn, R. RNA editing produces glycine receptor α3P185L, resulting in high agonist potency. Nat. Neurosci. 2005, 8, 736–744. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014. [Google Scholar] [CrossRef]
- Anant, S.; Davidson, N.O. An AU-rich sequence element (UUUN[A/U]U) downstream of the edited C in apolipoprotein B mRNA is a high-affinity binding site for Apobec-1: Binding of Apobec-1 to this motif in the 3’ untranslated region of c-myc increases mRNA stability. Mol. Cell. Biol. 2000, 20, 1982–1992. [Google Scholar] [CrossRef]
- Beaudoing, E.; Freier, S.; Wyatt, J.R.; Claverie, J.M.; Gautheret, D. Patterns of variant polyadenylation signal usage in human genes. Genome Res. 2000, 10, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Picardi, E.; Horner, D.S.; Pesole, G. Single-cell transcriptomics reveals specific RNA editing signatures in the human brain. RNA 2017. [Google Scholar] [CrossRef] [PubMed]
- Vogel, P.; Stafforst, T. Critical review on engineering deaminases for site-directed RNA editing. Curr. Opin. Biotechnol. 2018, 55, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Blanc, V.; Henderson, J.O.; Newberry, R.D.; Xie, Y.; Cho, S.J.; Newberry, E.P.; Kennedy, S.; Rubin, D.C.; Wang, H.L.; Luo, J.; et al. Deletion of the AU-rich RNA binding protein apobec-1 reduces intestinal tumor burden in Apcminmice. Cancer Res. 2007. [Google Scholar] [CrossRef] [PubMed]
- Anant, S.; Murmu, N.; Houchen, C.W.; Mukhopadhyay, D.; Riehl, T.E.; Young, S.G.; Morrison, A.R.; Stenson, W.F.; Davidson, N.O. Apobec-1 protects intestine from radiation injury through posttranscriptional regulation of cyclooxygenase-2 expression. Gastroenterology 2004. [Google Scholar] [CrossRef]
- Tsujii, M.; DuBois, R.N. Alterations in cellular adhesion and apoptosis in epithelial cells overexpressing prostaglandin endoperoxide synthase 2. Cell 1995. [Google Scholar] [CrossRef]
- Eberhart, C.E.; Coffey, R.J.; Radhika, A.; Giardiello, F.M.; Ferrenbach, S.; DuBois, R.N. Up-regulation of cyclooxygenase 2 gene expression in human colorectal adenomas and adenocarcinomas. Gastroenterology 1994. [Google Scholar] [CrossRef]
- MacGinnitie, A.J.; Anant, S.; Davidson, N.O. Mutagenesis of apobec-1, the catalytic subunit of the mammalian apolipoprotein B mRNA editing enzyme, reveals distinct domains that mediate cytosine nucleoside deaminase, RNA binding, and RNA editing activity. J. Biol. Chem. 1995, 270, 14768–14775. [Google Scholar] [CrossRef]
- Petersen-Mahrt, S.K.; Neuberger, M.S. In vitro deamination of cytosine to uracil in single-stranded DNA by apolipoprotein B editing complex catalytic subunit 1 (APOBEC1). J. Biol. Chem. 2003. [Google Scholar] [CrossRef]
- Gee, P.; Ando, Y.; Kitayama, H.; Yamamoto, S.P.; Kanemura, Y.; Ebina, H.; Kawaguchi, Y.; Koyanagi, Y. APOBEC1-mediated editing and attenuation of herpes simplex virus 1 DNA indicate that neurons have an antiviral role during herpes simplex encephalitis. J. Virol. 2011. [Google Scholar] [CrossRef]
- Gonzalez, M.C.; Suspène, R.; Henry, M.; Guétard, D.; Wain-Hobson, S.; Vartanian, J.P. Human APOBEC1 cytidine deaminase edits HBV DNA. Retrovirology 2009. [Google Scholar] [CrossRef] [PubMed]
- Morgan, H.D.; Dean, W.; Coker, H.A.; Reik, W.; Petersen-Mahrt, S.K. Activation-induced cytidine deaminase deaminates 5-methylcytosine in DNA and is expressed in pluripotent tissues: Implications for epigenetic reprogramming. J. Biol. Chem. 2004. [Google Scholar] [CrossRef] [PubMed]
- LaRue, R.S.; Andresdottir, V.; Blanchard, Y.; Conticello, S.G.; Derse, D.; Emerman, M.; Greene, W.C.; Jonsson, S.R.; Landau, N.R.; Lochelt, M.; et al. Guidelines for naming nonprimate APOBEC3 Genes and proteins. J. Virol. 2009, 83, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Orecchini, E.; Frassinelli, L.; Galardi, S.; Ciafrè, S.A.; Michienzi, A. Post-transcriptional regulation of LINE-1 retrotransposition by AID/APOBEC and ADAR deaminases. Chromosome Res. 2018, 26, 45–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, V.; Bloch, N.; Landau, N.R. Intrinsic host restrictions to HIV-1 and mechanisms of viral escape. Nat. Immunol. 2015, 16, 546–553. [Google Scholar] [CrossRef]
- Cullen, B.R. Role and mechanism of action of the APOBEC3 family of antiretroviral resistance factors. J. Virol. 2006, 80, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Delebecque, F.; Suspene, R.; Calattini, S.; Casartelli, N.; Saıb, A.; Froment, A.; Wain-Hobson, S.; Gessain, A.; Vartanian, J.-P.; Schwartz, O. Restriction of foamy viruses by APOBEC cytidine deaminases. J. Virol. 2006, 80, 605–614. [Google Scholar] [CrossRef]
- Harris, R.S.; Dudley, J.P. APOBECs and virus restriction. Virology 2015, 479–480, 131–145. [Google Scholar] [CrossRef]
- Desimmie, B.A.; Burdick, R.C.; Izumi, T.; Doi, H.; Shao, W.; Alvord, W.G.; Sato, K.; Koyanagi, Y.; Jones, S.; Wilson, E.; et al. APOBEC3 proteins can copackage and comutate HIV-1 genomes. Nucleic Acids Res. 2016, 44, 7848–7865. [Google Scholar] [CrossRef] [Green Version]
- Zennou, V.; Perez-caballero, D.; Gottlinger, H.; Bieniasz, P.D. APOBEC3G incorporation into human immunodeficiency virus type 1 particles. J. Virol. 2004, 78, 12058–12061. [Google Scholar] [CrossRef]
- Mangeat, B.; Turelli, P.; Caron, G.; Friedli, M.; Perrin, L.; Trono, D. Broad antiretroviral defence by human APOBEC3G through lethal editing of nascent reverse transcripts. Nature 2003, 424, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X.; Ma, J.; Zhang, L.; Ma, L.; Mi, Z.; Zhou, J.; Guo, F.; Kleiman, L.; Cen, S. Human APOBEC3F incorporation into human immunodeficiency virus type 1 particles. Virus Res. 2014, 191, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Sheehy, A.M.; Gaddis, N.C.; Malim, M.H. The antiretroviral enzyme APOBEC3G is degraded by the proteasome in response to HIV-1 Vif. Nat. Med. 2003, 9, 1404–1407. [Google Scholar] [CrossRef]
- Stavrou, S.; Crawford, D.; Blouch, K.; Browne, E.P.; Kohli, R.M.; Ross, S.R. Different modes of retrovirus restriction by human APOBEC3A and APOBEC3G in vivo. PLoS Pathog. 2014, 10, e1004145. [Google Scholar] [CrossRef] [PubMed]
- Münk, C.; Willemsen, A.; Bravo, I.G. An ancient history of gene duplications, fusions and losses in the evolution of APOBEC3 mutators in mammals. BMC Evol. Biol. 2012, 12, 71. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, M.; Bleiber, G.; Martinez, R.; Kaessmann, H.; Telenti, A. Patterns of evolution of host proteins involved in retroviral pathogenesis. Retrovirology 2006, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- He, X.-T.; Xu, H.-Q.; Wang, X.-M.; He, X.-S.; Niu, J.-Q.; Gao, P.-J. Association between polymorphisms of the APOBEC3G gene and chronic hepatitis B viral infection and hepatitis B virus-related hepatocellular carcinoma. World J. Gastroenterol. 2017, 23, 232. [Google Scholar] [CrossRef]
- Duggal, N.K.; Fub, W.; Akeyb, J.M.; Emerman, M. Identification and antiviral activity of common polymorphisms in the APOBEC3 locus in human populations. Virology 2013, 72, 329–337. [Google Scholar] [CrossRef]
- Sheehy, A.M.; Gaddis, N.C.; Choi, J.D.; Malim, M.H. Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature 2002, 418, 646–650. [Google Scholar] [CrossRef]
- Suspène, R.; Sommer, P.; Henry, M.; Ferris, S.; Guétard, D.; Pochet, S.; Chester, A.; Navaratnam, N.; Wain-Hobson, S.; Vartanian, J.P. APOBEC3G is a single-stranded DNA cytidine deaminase and functions independently of HIV reverse transcriptase. Nucleic Acids Res. 2004, 32, 2421–2429. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ao, Z.; Chen, L.; Kobinger, G.; Peng, J.; Yao, X. The cellular antiviral protein APOBEC3G interacts with HIV-1 reverse transcriptase and inhibits its function during viral replication. J. Virol. 2012, 86, 3777–3786. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Goila-Gaur, R.; Opi, S.; Miyagi, E.; Takeuchi, H.; Kao, S.; Strebel, K. Analysis of the contribution of cellular and viral RNA to the packaging of APOBEC3G into HIV-1 virions. Retrovirology 2007, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bach, D.; Peddi, S.; Mangeat, B.; Lakkaraju, A.; Strub, K.; Trono, D. Characterization of APOBEC3G binding to 7SL RNA. Retrovirology 2008, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Svarovskaia, E.S.; Xu, H.; Mbisa, J.L.; Barr, R.; Gorelick, R.J.; Ono, A.; Freed, E.O.; Hu, W.S.; Pathak, V.K. Human apolipoprotein B mRNA-editing enzyme-catalytic polypeptide-like 3G (APOBEC3G) is incorporated into HIV-1 virions through interactions with viral and nonviral RNAs. J. Biol. Chem. 2004, 279, 35822–35828. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Tian, C.; Zhang, W.; Luo, K.; Sarkis, P.T.N.; Yu, L.; Liu, B.; Yu, Y.; Yu, X.-F. 7SL RNA mediates virion packaging of the antiviral cytidine deaminase APOBEC3G. J. Virol. 2007, 81, 13112–13124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, B.; Pomerantz, R.J.; Zhang, C.; Arunachalam, S.C.; Gao, L. The cytidine deaminase CEM15 induces hypermutation in newly synthesized HIV-1 DNA. Nature 2003, 424, 94–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, R.P.; Presnyak, V.; Wedekind, J.E.; Smith, H.C. Nuclear exclusion of the HIV-1 host defense factor APOBEC3G requires a novel cytoplasmic retention signal and is not dependent on RNA binding. J. Biol. Chem. 2008, 283, 7320–7327. [Google Scholar] [CrossRef]
- Gallois-Montbrun, S.; Kramer, B.; Swanson, C.M.; Byers, H.; Lynham, S.; Ward, M.; Malim, M.H. Antiviral protein APOBEC3G localizes to ribonucleoprotein complexes found in P bodies and stress granules. J. Virol. 2007, 81, 2165–2178. [Google Scholar] [CrossRef]
- Zhen, A.; Du, J.; Zhou, X.; Xiong, Y.; Yu, X.F. Reduced APOBEC3H variant anti-viral activities are associated with altered RNA binding activities. PLoS ONE 2012, 7, 1–10. [Google Scholar] [CrossRef]
- Land, A.M.; Law, E.K.; Carpenter, M.A.; Lackey, L.; Brown, W.L.; Harris, R.S. Endogenous APOBEC3A DNA cytosine deaminase is cytoplasmic and nongenotoxic. J. Biol. Chem. 2013, 288, 17253–17260. [Google Scholar] [CrossRef]
- Aynaud, M.-M.M.; Suspène, R.; Vidalain, P.-O.O.; Mussil, B.; Guétard, D.; Tangy, F.; Wain-Hobson, S.; Vartanian, J.-P.P. Human tribbles 3 protects nuclear DNA from cytidine deamination by APOBEC3A. J. Biol. Chem. 2012, 287, 39182–39192. [Google Scholar] [CrossRef] [PubMed]
- Lackey, L.; Demorest, Z.L.; Land, A.M.; Hultquist, J.F.; Brown, W.L.; Harris, R.S. APOBEC3B and AID have similar nuclear import mechanisms. J. Mol. Biol. 2012, 419, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Nik-Zainal, S.; Alexandrov, L.B.; Wedge, D.C.; Van Loo, P.; Greenman, C.D.; Raine, K.; Jones, D.; Hinton, J.; Marshall, J.; Stebbings, L.A.; et al. Mutational processes molding the genomes of 21 breast cancers. Cell 2012, 149, 979–993. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.J.R.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.-L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, M.B.; Temiz, N.A.; Harris, R.S. Evidence for APOBEC3B mutagenesis in multiple human cancers. Nat. Genet. 2013, 45, 977–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caval, V.; Suspène, R.; Shapira, M.; Vartanian, J.; Wain-Hobson, S. A prevalent cancer susceptibility APOBEC3A hybrid allele bearing APOBEC3B 3′UTR enhances chromosomal DNA damage. Nat. Commun. 2014, 5, 5129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowarski, R.; Kotler, M. APOBEC3 cytidine deaminases in double-strand DNA break repair and cancer promotion. Cancer Res. 2013, 73, 3494–3498. [Google Scholar] [CrossRef]
- Seplyarskiy, V.B.; Soldatov, R.A.; Popadin, K.Y.; Antonarakis, S.E.; Bazykin, G.A.; Nikolaev, S.I. APOBEC-induced mutations in human cancers are strongly enriched on the lagging DNA strand during replication. Genome Res. 2016, 26, 174–182. [Google Scholar] [CrossRef] [Green Version]
- Swanton, C.; McGranahan, N.; Starrett, G.J.; Harris, R.S. APOBEC enzymes: mutagenic fuel for cancer evolution and heterogeneity. Cancer Discov. 2015, 5, 704–712. [Google Scholar] [CrossRef]
- Roberts, S.A.; Lawrence, M.S.; Klimczak, L.J.; Grimm, S.A.; Fargo, D.; Stojanov, P.; Kiezun, A.; Kryukov, G.V.; Carter, S.L.; Saksena, G.; et al. An APOBEC cytidine deaminase mutagenesis pattern is widespread in human cancers. Nat. Genet. 2013, 45, 970–976. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.; Roberts, S.A.; Klimczak, L.J.; Sterling, J.F.; Saini, N.; Malc, E.P.; Kim, J.; Kwiatkowski, D.J.; Fargo, D.C.; Mieczkowski, P.A.; et al. An APOBEC3A hypermutation signature is distinguishable from the signature of background mutagenesis by APOBEC3B in human cancers. Nat. Genet. 2015, 47, 1067–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, M.B.; Lackey, L.; Carpenter, M.A.; Rathore, A.; Land, A.M.; Leonard, B.; Refsland, E.W.; Kotandeniya, D.; Tretyakova, N.; Nikas, J.B.; et al. APOBEC3B is an enzymatic source of mutation in breast cancer. Nature 2013, 494, 366–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gansmo, L.B.; Romundstad, P.; Hveem, K.; Vatten, L.; Nik-Zainal, S.; Lønning, P.E.; Knappskog, S. APOBEC3A/B deletion polymorphism and cancer risk. Carcinogenesis 2018, 39, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Law, E.K.; Sieuwerts, A.M.; Lapara, K.; Leonard, B.; Starrett, G.J.; Molan, A.M.; Temiz, N.A.; Vogel, R.I.; Meijer-Van Gelder, M.E.; Sweep, F.C.G.J.; et al. The DNA cytosine deaminase APOBEC3B promotes tamoxifen resistance in ER-positive breast cancer. Sci. Adv. 2016, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sieuwerts, A.M.; Willis, S.; Burns, M.B.; Look, M.P.; Gelder, M.E.M.; Schlicker, A.; Heideman, M.R.; Jacobs, H.; Wessels, L.; Leyland-Jones, B.; et al. Elevated APOBEC3B correlates with poor outcomes for estrogen-receptor-positive breast cancers. Horm. Cancer 2014, 5, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Glaser, A.P.; Fantini, D.; Wang, Y.; Yu, Y.; Rimar, K.J.; Podojil, J.R.; Miller, S.D.; Meeks, J.J. APOBEC-mediated mutagenesis in urothelial carcinoma is associated with improved survival, mutations in DNA damage response genes, and immune response. Oncotarget 2018, 9, 4537–4548. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Chang, C.; Xie, X.; Xia, W.; Yang, J.; Wang, S.; Wang, Y.; Xia, J.; Chen, L.; Cai, C.; et al. APOBEC3G promotes liver metastasis in an orthotopic mouse model of colorectal cancer and predicts human hepatic metastasis. Cancer 2011, 121. [Google Scholar] [CrossRef]
- Lan, H.; Jin, K.; Gan, M.; Wen, S.; Bi, T.; Zhou, S.; Zhu, N.; Teng, L.; Yu, W. APOBEC3G expression is correlated with poor prognosis in colon carcinoma patients with hepatic metastasis. Int. J. Clin. Exp. Med. 2014, 7, 665–672. [Google Scholar]
- Leonard, B.; Starrett, G.J.; Maurer, M.J.; Oberg, A.L.; Van Bockstal, M.; Van Dorpe, J.; De Wever, O.; Helleman, J.; Sieuwerts, A.M.; Berns, E.M.J.J.; et al. APOBEC3G expression correlates with T-cell infiltration and improved clinical outcomes in high-grade serous ovarian carcinoma. Clin. Cancer Res. 2016, 22, 4746–4755. [Google Scholar] [CrossRef]
- Nowarski, R.; Wilner, O.I.; Cheshin, O.; Shahar, O.D.; Kenig, E.; Baraz, L.; Britan-Rosich, E.; Nagler, A.; Harris, R.S.; Goldberg, M.; et al. APOBEC3G enhances lymphoma cell radioresistance by promoting cytidine deaminase-dependent DNA repair. Blood 2012, 120, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, S.; Zheng, S.; Wang, S.; Wali, A.; Ezhilarasan, R.; Sulman, E.P.; Koul, D.; Alfred Yung, W.K. APOBEC3G acts as a therapeutic target in mesenchymal gliomas by sensitizing cells to radiation-induced cell death. Oncotarget 2017, 8, 54285–54296. [Google Scholar] [CrossRef] [PubMed]
- Prohaska, K.M.; Bennett, R.P.; Salter, J.D.; Smith, H.C. The multifaceted roles of RNA binding in APOBEC cytidine deaminase functions. Wiley Interdiscip. Rev. RNA 2014, 5, 493–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Patnaik, S.K.; Kemer, Z.; Baysal, B.E. Transient overexpression of exogenous APOBEC3A causes C-to-U RNA editing of thousands of genes. RNA Biol. 2017, 14, 603–610. [Google Scholar] [CrossRef]
- Sharma, S.; Wang, J.; Portwood, S.; Cortes-Gomez, E.; Maguire, O.; Basse, P.H.; Wang, E.S.; Baysal, B.E. Mitochondrial hypoxic stress induces widespread RNA editing by APOBEC3G in lymphocytes. BioRxiv 2018. [Google Scholar] [CrossRef]
- Smith, H.C. RNA binding to APOBEC deaminases; Not simply a substrate for C to U editing. RNA Biol. 2017, 14, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Koning, F.A.; Newman, E.N.C.; Kim, E.-Y.; Kunstman, K.J.; Wolinsky, S.M.; Malim, M.H. Defining APOBEC3 expression patterns in human tissues and hematopoietic cell subsets. J. Virol. 2009, 83, 9474–9485. [Google Scholar] [CrossRef] [PubMed]
- Polevoda, B.; McDougall, W.M.; Tun, B.N.; Cheung, M.; Salter, J.D.; Friedman, A.E.; Smith, H.C. RNA binding to APOBEC3G induces the disassembly of functional deaminase complexes by displacing single-stranded DNA substrates. Nucleic Acids Res. 2015, 43, 9434–9445. [Google Scholar] [CrossRef] [Green Version]
- Bélanger, K.; Savoie, M.; Rosales Gerpe, M.C.; Couture, J.F.; Langlois, M.A. Binding of RNA by APOBEC3G controls deamination-independent restriction of retroviruses. Nucleic Acids Res. 2013, 41, 7438–7452. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Goubran, M.H.; Follack, T.B.; Chelico, L. Deamination-independent restriction of LINE-1 retrotransposition by APOBEC3H. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Kobayashi, T.; Koizumi, Y.; Takeuchi, J.S.; Misawa, N.; Kimura, Y.; Morita, S.; Aihara, K.; Koyanagi, Y.; Iwami, S.; Sato, K. Quantification of deaminase activity-dependent and -independent restriction of HIV-1 replication mediated by APOBEC3F and APOBEC3G through experimental-mathematical investigation. J. Virol. 2014, 88, 5881–5887. [Google Scholar] [CrossRef]
- Schmitt, K.; Guo, K.; Katuwal, M.; Wilson, D.; Prochnow, C.; Bransteitter, R.; Chen, X.S.; Santiago, M.L.; Stephens, E.B. Lentivirus restriction by diverse primate APOBEC3A proteins. Virology 2013, 442, 82–96. [Google Scholar] [CrossRef] [Green Version]
- Kozak, S.L.; Marin, M.; Rose, K.M.; Bystrom, C.; Kabat, D. The anti-HIV-1 editing enzyme APOBEC3G binds HIV-1 RNA and messenger RNAs that shuttle between polysomes and stress granules. J. Biol. Chem. 2006. [Google Scholar] [CrossRef] [PubMed]
- Gallois-Montbrun, S.; Holmes, R.K.; Swanson, C.M.; Fernandez-Ocana, M.; Byers, H.L.; Ward, M.A.; Malim, M.H. Comparison of cellular ribonucleoprotein complexes associated with the APOBEC3F and APOBEC3G antiviral proteins. J. Virol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Blanc, V.; Davidson, N.O. APOBEC-1 mediated RNA editing. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 594–602. [Google Scholar] [CrossRef]
- Eichler, S.A.; Kirischuk, S.; Jüttner, R.; Schafermeier, P.K.; Legendre, P.; Lehmann, T.N.; Gloveli, T.; Grantyn, R.; Meier, J.C. Glycinergic tonic inhibition of hippocampal neurons with depolarizing GABAergic transmission elicits histopathological signs of temporal lobe epilepsy. J. Cell. Mol. Med. 2008. [Google Scholar] [CrossRef] [PubMed]
- Winkelmann, A.; Maggio, N.; Eller, J.; Caliskan, G.; Semtner, M.; Häussler, U.; Jüttner, R.; Dugladze, T.; Smolinsky, B.; Kowalczyk, S.; et al. Changes in neural network homeostasis trigger neuropsychiatric symptoms. J. Clin. Investig. 2014. [Google Scholar] [CrossRef] [PubMed]
- Çalişkan, G.; Müller, I.; Semtner, M.; Winkelmann, A.; Raza, A.S.; Hollnagel, J.O.; Rösler, A.; Heinemann, U.; Stork, O.; Meier, J.C. Identification of parvalbumin interneurons as cellular substrate of fear memory persistence. Cereb. Cortex 2016. [Google Scholar] [CrossRef] [PubMed]
- Kankowski, S.; Förstera, B.; Winkelmann, A.; Knauff, P.; Wanker, E.E.; You, X.A.; Semtner, M.; Hetsch, F.; Meier, J.C. A Novel RNA editing sensor tool and a specific agonist determine neuronal protein expression of RNA-edited glycine receptors and identify a genomic APOBEC1 dimorphism as a new genetic risk factor of epilepsy. Front. Mol. Neurosci. 2018, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Min, J.; Funahashi, T.; Baunoch, D.A.; Davidson, N.O. Characterization of the human apobec-1 gene: Expression in gastrointestinal tissues determined by alternative splicing with production of a novel truncated peptide. J. Lipid Res. 1997, 38, 847–859. [Google Scholar]
- Grohmann, M.; Hammer, P.; Walther, M.; Paulmann, N.; Büttner, A.; Eisenmenger, W.; Baghai, T.C.; Schüle, C.; Rupprecht, R.; Bader, M.; et al. Alternative splicing and extensive RNA editing of human TPH2 transcripts. PLoS ONE 2010. [Google Scholar] [CrossRef]
- Vanharanta, S.; Marney, C.B.; Shu, W.; Valiente, M.; Zou, Y.; Mele, A.; Darnell, R.B.; Massagué, J. Loss of the multifunctional RNA-binding protein RBM47 as a source of selectable metastatic traits in breast cancer. Elife 2014. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Isogaya, K.; Sakai, S.; Morikawa, M.; Morishita, Y.; Ehata, S.; Miyazono, K.; Koinuma, D. RNA-binding motif protein 47 inhibits Nrf2 activity to suppress tumor growth in lung adenocarcinoma. Oncogene 2017, 35, 5000–5009. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Nishitsuji, H.; Marusawa, H.; Ujino, S.; Takaku, H.; Shimotohno, K. The RNA-editing enzyme APOBEC1 requires heterogeneous nuclear ribonucleoprotein Q Isoform 6 for efficient interaction with interleukin-8 mRNA. J. Biol. Chem. 2014, 289, 26226–26238. [Google Scholar] [CrossRef] [PubMed]
- Chiche, L.; Jourde-Chiche, N.; Whalen, E.; Presnell, S.; Gersuk, V.; Dang, K.; Anguiano, E.; Quinn, C.; Burtey, S.; Berland, Y.; et al. Modular transcriptional repertoire analyses of adults with systemic lupus erythematosus reveal distinct type I and type II interferon signatures. Arthritis Rheumatol. 2014, 66, 1583–1595. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, N.; Rönnblom, L. Systemic Lupus Erythematosus—A disease with a dysregulated type I interferon system. Scand. J. Immunol. 2015, 82, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Roth, S.H.; Danan-Gotthold, M.; Ben-Izhak, M.; Rechavi, G.; Cohen, C.J.; Louzoun, Y.; Levanon, E.Y. Increased RNA editing may provide a source for autoantigens in Systemic Lupus Erythematosus. Cell Rep. 2018, 23, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Baysal, B.E. Stem-loop structure preference for site-specific RNA editing by APOBEC3A and APOBEC3G. PeerJ 2017, 5, e4136. [Google Scholar] [CrossRef]
- Radanova, M.; Vasilev, V.; Dimitrov, T.; Deliyska, B.; Ikonomov, V.; Ivanova, D. Association of rs172378 C1q gene cluster polymorphism with lupus nephritis in Bulgarian patients. Lupus 2015, 24, 280–289. [Google Scholar] [CrossRef]
- Namjou, B.; Gray-McGuire, C.; Sestak, A.L.; Gilkeson, G.S.; Jacob, C.O.; Merrill, J.T.; James, J.A.; Wakeland, E.K.; Li, Q.Z.; Langefeld, C.D.; et al. Evaluation of C1q genomic region in minority racial groups of lupus. Genes Immun. 2009, 10, 517–524. [Google Scholar] [CrossRef] [Green Version]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lerner, T.; Papavasiliou, F.N.; Pecori, R. RNA Editors, Cofactors, and mRNA Targets: An Overview of the C-to-U RNA Editing Machinery and Its Implication in Human Disease. Genes 2019, 10, 13. https://doi.org/10.3390/genes10010013
AMA Style
Lerner T, Papavasiliou FN, Pecori R. RNA Editors, Cofactors, and mRNA Targets: An Overview of the C-to-U RNA Editing Machinery and Its Implication in Human Disease. Genes. 2019; 10(1):13. https://doi.org/10.3390/genes10010013
Chicago/Turabian StyleLerner, Taga, F. Nina Papavasiliou, and Riccardo Pecori. 2019. "RNA Editors, Cofactors, and mRNA Targets: An Overview of the C-to-U RNA Editing Machinery and Its Implication in Human Disease" Genes 10, no. 1: 13. https://doi.org/10.3390/genes10010013
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.