Applicability of Obesity-Related SNPs and Their Effect Size Measures Defined on Populations with European Ancestry for Genetic Risk Estimation among Roma

 , ,
, , 
Abstract
:
1. Introduction
2. Material and Methods
2.1. Study Design
2.2. Sampling of Hungarian Roma Subjects
2.2.1. Recruitment of Hungarian Roma Adult Subjects
2.2.2. Recruitment of Hungarian General Adult Subjects
2.3. Genotyping and Anthropometric Data
2.4. Ethical Statement
2.5. Statistical Analyses
3. Results
3.1. Characteristics of the Study Samples
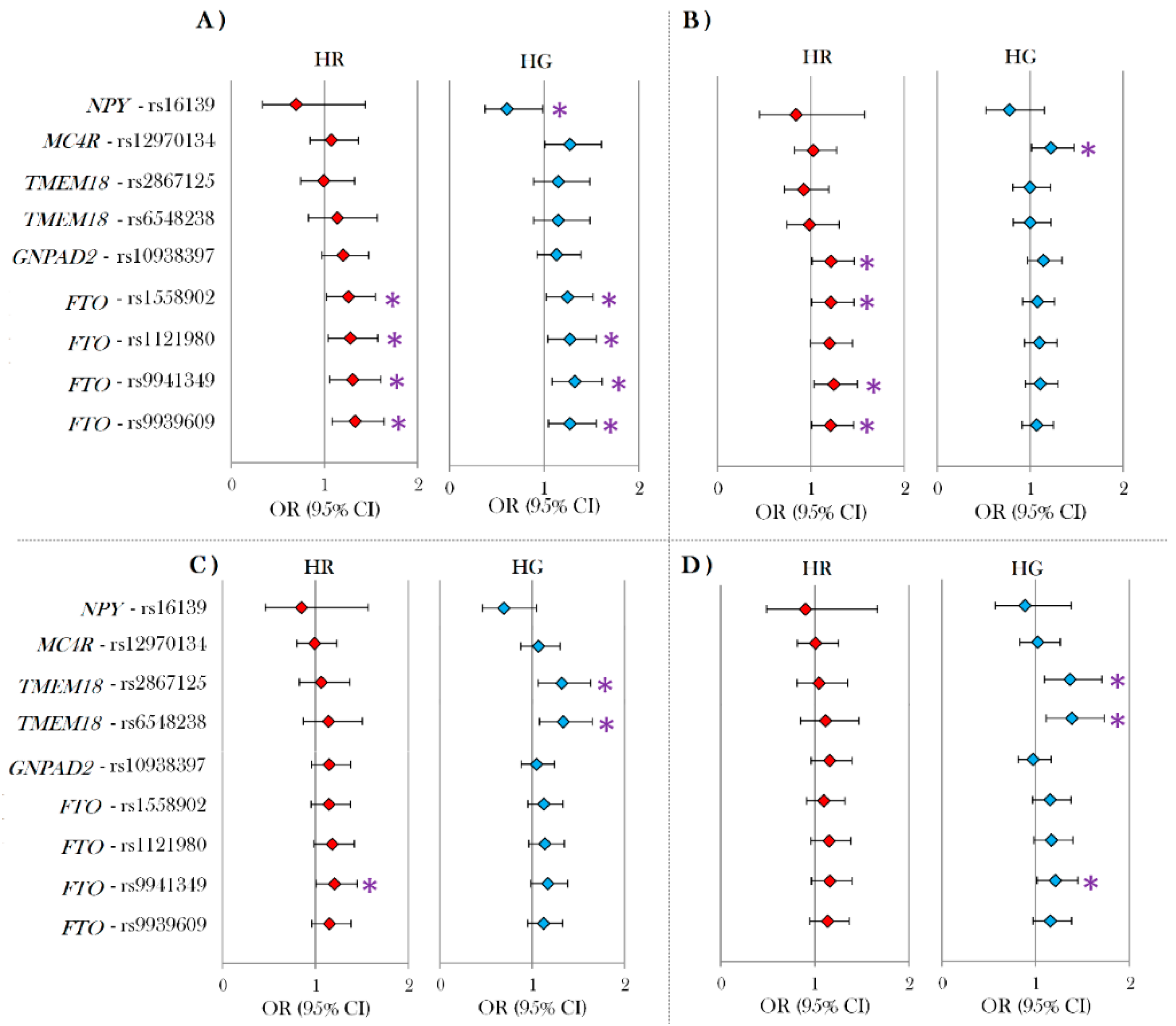
3.2. Individual Effects of SNPs in the Hungarian General and Roma Population
3.3. The Results of Different Genetic Models for the rs1801282 Gene Polymorphism in PPARγ
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- European Commission. Estimates and Official Numbers of Roma in Europe. 2012. Press Corner. Available online: https://ec.europa.eu/commission/presscorner/detail/en/MEMO_14_249 (accessed on 5 May 2020).
- Sándor, J.; Kósa, Z.; Boruzs, K.; Boros, J.; Tokaji, I.; McKee, M.; Ádány, R. The decade of Roma Inclusion: Did it make a difference to health and use of health care services? Int. J. Public Health 2017, 62, 803–815. [Google Scholar] [CrossRef] [Green Version]
- Fraser, A. Origins. In The Gypsies, 2nd ed.; Campbell, J., Cunliffe, B., Eds.; Blackwell Publishers Ltd.: Oxford, UK, 1992; pp. 10–28. [Google Scholar]
- Iovita, R.P.; Schurr, T.G. Reconstructing the origins and migrations of diasporic populations: The case of the European gypsies. Am. Anthr. 2004, 106, 267–281. [Google Scholar] [CrossRef]
- Pamjav, H.; Zalán, A.; Béres, J.; Nagy, M.; Chang, Y.M. Genetic structure of the paternal lineage of the Roma people. Am. J. Phys. Anthr. 2011, 145, 21–29. [Google Scholar] [CrossRef]
- Mendizabal, I.; Lao, O.; Marigorta, U.M.; Wollstein, A.; Gusmão, L.; Ferák, V.; Ioana, M.; Jordanova, A.; Kaneva, R.; Kouvatsi, A.; et al. Reconstructing the population history of European Romani from genome-wide data. Curr. Boil. 2012, 22, 2342–2349. [Google Scholar] [CrossRef] [Green Version]
- Melegh, B.; Bánfai, Z.; Hadzsiev, K.; Miseta, A.; Melegh, B. Refining the South Asian origin of the Romani people. BMC Genet. 2017, 18, 82. [Google Scholar] [CrossRef] [Green Version]
- Font-Porterias, N.; Arauna, L.R.; Poveda, A.; Bianco, E.; Rebato, E.; Prata, M.J.; Calafell, F.; Comas, D. European Roma groups show complex West Eurasian admixture footprints and a common South Asian genetic origin. PLoS Genet. 2019, 15, e1008417. [Google Scholar] [CrossRef]
- Regueiro, M.; Rivera, L.; Chennakrishnaiah, S.; Popović, B.; Andjus, S.; Milasin, J.; Herrera, R.J. Ancestral modal Y-STR haplotype shared among Romani and South Indian populations. Gene 2012, 504, 296–302. [Google Scholar] [CrossRef]
- European Commission. Funding, Strategy, Facts and Figures and Contact Details for National Roma Contact points in Hungary, 2015. Roma Integration in Hungary. Available online: https://ec.europa.eu/info/policies/justice-and-fundamental-rights/combatting-discrimination/roma-and-eu/roma-integration-eu-country/roma-integration-hungary_en#facts-and-figures (accessed on 5 May 2020).
- Belak, A.; Madarasova-Geckova, A.; Van Dijk, J.P.; Reijneveld, S.A. Why don’t segregated Roma do more for their health? An explanatory framework from an ethnographic study in Slovakia. Int. J. Public Health 2018, 63, 1123–1131. [Google Scholar] [CrossRef] [Green Version]
- Jarcuska, P.; Team, H.-M.; Bobakova, D.F.; Uhrin, J.; Bobák, L.; Babinska, I.; Kolarčik, P.; Veselska, Z.; Madarasova-Geckova, A.; Dankulincova, Z. Are barriers in accessing health services in the Roma population associated with worse health status among Roma? Int. J. Public Health 2013, 58, 427–434. [Google Scholar] [CrossRef] [Green Version]
- La Parra-Casado, D.; Gil-González, D.; Esteve, M.D.L.T. The social class gradient in health in Spain and the health status of the Spanish Roma. Ethn. Health 2015, 21, 1–12. [Google Scholar] [CrossRef]
- Le Cook, B.; Wayne, G.F.; Valentine, A.; Lessios, A.; Yeh, E. Revisiting the evidence on health and health care disparities among the Roma: a systematic review 2003–2012. Int. J. Public Health 2013, 58, 885–911. [Google Scholar] [CrossRef] [PubMed]
- Pázstor, I.Z.; Pénzes, J.; Tátrai, P. Pálóczi, Á. The number and spatial distribution of the Roma population in Hungary–in the light of different approaches. Folia Geograph 2016, 58, 5–21. [Google Scholar]
- Kósa, Z.; Moravcsik-Kornyicki, Á.; Diószegi, J.; Roberts, B.; Szabó, B.; Sándor, J.; Ádány, R. Prevalence of metabolic syndrome among Roma: A comparative health examination survey in Hungary. Eur. J. Public Health 2014, 25, 299–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD. OECD Health Statistics 2017. 2017. OECD Obesity Update. Available online: https://www.oecd.org/els/health-systems/Obesity-Update-2017.pdf (accessed on 5 May 2020).
- González-Muniesa, P.; Mártinez-González, M.A.; Hu, F.B.; Després, J.P.; Matsuzawa, Y.; Loos, R.J.F.; Moreno, L.A.; Bray, G.A.; Martinez, J.A. Obesity. Nat. Rev. Dis. Primers 2017, 3, 17034. [Google Scholar] [CrossRef]
- Ghosh, S.; Bouchard, C. Convergence between biological, behavioural and genetic determinants of obesity. Nat. Rev. Genet. 2017, 18, 731–748. [Google Scholar] [CrossRef]
- Barroso, I.; McCarthy, M.I. The genetic basis of metabolic disease. Cell 2019, 177, 146–161. [Google Scholar] [CrossRef] [Green Version]
- Stryjecki, C.; Alyass, A.; Meyre, D. Ethnic and population differences in the genetic predisposition to human obesity. Obes. Rev. 2017, 19, 62–80. [Google Scholar] [CrossRef]
- Evangelatos, N.; Satyamoorthy, K.; Brand, A. Personalized health in a public health perspective. Int. J. Public Health 2017, 63, 433–434. [Google Scholar] [CrossRef] [Green Version]
- Topol, E.J. Individualized medicine from prewomb to tomb. Cell 2014, 157, 241–253. [Google Scholar] [CrossRef] [Green Version]
- Yuzhalin, A.E.; Kutikhin, A.G. Integrative systems of genomic risk markers for cancer and other diseases: future of predictive medicine. Cancer Manag. Res. 2012, 4, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, M.O. Genetics of obesity: what genetic association studies have taught us about the biology of obesity and its complications. Lancet Diabetes Endocrinol. 2018, 6, 223–236. [Google Scholar] [CrossRef]
- Poveda, A.; Ibáñez, M.E.; Rebato, E. Common variants in BDNF, FAIM2, FTO, MC4R, NEGR1, and SH2B1 show association with obesity-related variables in Spanish Roma population. Am. J. Hum. Biol. 2014, 26, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Maceková, S.; Bernasovský, I.; Gabriková, D.; Bôžiková, A.; Bernasovská, J.; Boronová, I.; Behulova, R.; Švícková, P.; Petrejcíková, E.; Soták, M.; et al. Association of the FTO rs9939609 polymorphism with obesity in Roma/Gypsy population. Am. J. Phys. Anthr. 2011, 147, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Nagy, K.; Fiatal, S.; Sándor, J.; Ádány, R. Distinct Penetrance of Obesity-Associated Susceptibility Alleles in the Hungarian General and Roma Populations. Obes. Facts 2017, 10, 444–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ádány, R.; Kósa, K.; Sándor, J.; Papp, M.; Fürjes, G. General practitioners’ cluster: a model to reorient primary health care to public health services. Eur. J. Public Health 2013, 23, 529–530. [Google Scholar] [CrossRef] [Green Version]
- Széles, G.; Vokó, Z.; Jenei, T.; Kardos, L.; Pocsai, Z.; Bajtay, A.; Papp, E.; Pásti, G.; Kósa, Z.; Molnár, I.; et al. A preliminary evaluation of a health monitoring programme in Hungary. Eur. J. Public Health 2005, 15, 26–32. [Google Scholar] [CrossRef]
- Szigethy, E.; Széles, G.; Horvath, A.; Hídvégi, T.; Jermendy, G.; Paragh, G.; Blaskó, G.; Ádány, R.; Vokó, Z. Epidemiology of the metabolic syndrome in Hungary. Public Health 2012, 126, 143–149. [Google Scholar] [CrossRef]
- Expert Panel on Detection Evaluation and Treatment of High Blood Cholesterol in Adults. Executive summary of the third report of the national cholesterol education program (NCEP) expert panel on detection, evaluation, and treatment of high blood cholesterol in adults (adult treatment panel III). JAMA 2001, 285, 2486–2497. [Google Scholar] [CrossRef]
- Zimmet, P.; Alberti, K.G.M.; Ríos, M.S. A new international diabetes federation (IDF) worldwide definition of the metabolic syndrome: the rationale and the results. Rev Esp Cardiol (English Edition) 2005, 58, 1371–1375. [Google Scholar] [CrossRef]
- Templeton, G.F. A two-step approach for transforming continuous variables to normal: Implications and recommendations for IS research. Commun. Assoc. Inf. Syst. 2011, 28, 4. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauderman, W.J. Sample size requirements for matched case-control studies of gene-environment interaction. Stat Med. 2002, 21, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Meyre, D. Challenges in reproducibility of genetic association studies: lessons learned from the obesity field. Int. J. Obes. 2012, 37, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Baldani, D.P.; Škrgatić, L.; Cerne, J.Z.; Ferk, P.; Simunic, V.; Gersak, K. Association of PPARG Pro12Ala polymorphism with insulin sensitivity and body mass index in patients with polycystic ovary syndrome. Biomed. Rep. 2013, 2, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, D.; Mann, W.A.; De Heer, J.; Michel, U.; Wendt, D.; Kortner, B.; Wolf, A.; Beisiegel, U. Variation in the gene for human peroxisome proliferator activated receptor γ (PPARγ) does not play a major role in the development of morbid obesity. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 647–651. [Google Scholar] [CrossRef] [Green Version]
- Goni, L.; García-Granero, M.; Milagro, F.I.; Cuervo, M.; Martínez, J.A. Phenotype and genotype predictors of BMI variability among European adults. Nutr. Diabetes 2018, 8, 27. [Google Scholar] [CrossRef]
- Fajas, L.; Auboeuf, D.; Raspé, E.; Schoonjans, K.; Lefebvre, A.-M.; Saladin, R.; Najib, J.; Laville, M.; Fruchart, J.C.; Deeb, S.; et al. The organization, promoter analysis, and expression of the human PPARγ gene. J. Boil. Chem. 1997, 272, 18779–18789. [Google Scholar] [CrossRef] [Green Version]
- Robinson, E.; Grieve, D.J. Significance of peroxisome proliferator-activated receptors in the cardiovascular system in health and disease. Pharmacol. Ther. 2009, 122, 246–263. [Google Scholar] [CrossRef]
- Heikkinen, S.; Argmann, C.; Feige, J.N.; Koutnikova, H.; Champy, M.F.; Dali-Youcef, N.; Schadt, E.E.; Laakso, M.; Auwerx, J. The Pro12Ala PPARγ2 variant determines metabolism at the gene-environment interface. Cell Metabol. 2009, 9, 88–98. [Google Scholar] [CrossRef]
- Galbete, C.; Toledo, E.; A Martínez-González, M.; Martinez, J.; Guillen-Grima, F.; Marti, A. Pro12Ala variant of the PPARG2 gene increases body mass index: An updated meta-analysis encompassing 49,092 subjects. Obesity 2013, 21, 1486–1495. [Google Scholar] [CrossRef]
- Mansoori, A.; Amini, M.; Kolahdooz, F.; Seyedrezazadeh, E. Obesity and Pro12Ala polymorphism of peroxisome proliferator-activated receptor-gamma gene in healthy adults: A systematic review and meta-analysis. Ann. Nutr. Metab. 2015, 67, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Deeb, S.S.; Fajas, L.; Nemoto, M.; Pihlajamäki, J.; Mykkänen, L.; Kuusisto, J.; Laakso, M.; Fujimoto, W.; Auwerx, J. A Pro12Ala substitution in PPARγ2 associated with decreased receptor activity, lower body mass index and improved insulin sensitivity. Nat. Genet. 1998, 20, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, T.-J.; Lin, E. The Pro12Ala polymorphism in the peroxisome proliferator-activated receptor gamma (PPARG) gene in relation to obesity and metabolic phenotypes in a Taiwanese population. Endocrine 2014, 48, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Randall, J.; Winkler, T.W.; Kutalik, Z.; Berndt, S.I.; Jackson, A.U.; Monda, K.L.; Kilpeläinen, T.O.; Esko, T.; Magi, R.; Li, S.; et al. Sex-stratified genome-wide association studies including 270,000 individuals show sexual dimorphism in genetic loci for anthropometric traits. PLoS Genet. 2013, 9, e1003500. [Google Scholar] [CrossRef]
- Lamri, A.; Abi-Khalil, C.; Jaziri, R.; Velho, G.; Lantieri, O.; Vol, S.; Froguel, P.; Balkau, B.; Marre, M.; Fumeron, F. Dietary fat intake and polymorphisms at the PPARG locus modulate BMI and type 2 diabetes risk in the D.E.S.I.R. prospective study. Int. J. Obes. (Lond) 2012, 36, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Verier, C.; Meirhaeghe, A.; Bokor, S.; Breidenassel, C.; Manios, Y.; Molnár, D.; Artero, E.G.; Nova, E.; De Henauw, S.; Moreno, L.A.; et al. Breast-feeding modulates the influence of the peroxisome proliferator–activated receptor-γ (PPARG2) Pro12Ala polymorphism on adiposity in adolescents. Diabetes Care 2009, 33, 190–196. [Google Scholar] [CrossRef] [Green Version]
- Dedoussis, G.V.; Manios, Y.; Kourlaba, G.; Kanoni, S.; Lagou, V.; Butler, J.; Papoutsakis, C.; Scott, R.A.; Yannakoulia, M.; Pitsiladis, Y.P.; et al. An age-dependent diet-modified effect of the PPARγ Pro12Ala polymorphism in children. Metabolism 2011, 60, 467–473. [Google Scholar] [CrossRef]
- Mathieson, I.; Alpaslan-Roodenberg, S.; Posth, C.; Szécsényi-Nagy, A.; Rohland, N.; Mallick, S.; Olalde, I.; Broomandkhoshbacht, N.; Candilio, F.; Cheronet, O.; et al. The genomic history of southeastern Europe. Nature 2018, 555, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Laayouni, H.; Oosting, M.; Luisi, P.; Ioana, M.; Alonso, S.; Ricano-Ponce, I.; Trynka, G.; Zhernakova, A.; Plantinga, T.; Cheng, S.-C.; et al. Convergent evolution in European and Rroma populations reveals pressure exerted by plague on Toll-like receptors. Proc. Natl. Acad. Sci. USA 2014, 111, 2668–2673. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Characteristics | Hungarian General (N = 1496) | Hungarian Roma (N = 1141) | p-Value |
|---|---|---|---|
| Females (%) / Males (%) | 52.87/47.13 | 59.10/40.90 | 0.001 |
| Age (mean value (years) ± SD) | 44.17 ± 11.81 | 40.22 ± 13.00 | <0.001 |
| BMI (mean value (kg/m2) 95% CI) | 28.03 (27.75–28.32) | 26.15 (25.69–26.61) | <0.001 |
| WC (mean value (cm) 95% CI) | 94.89 (94.14–95.65) | 90.09 (89.08–91.10) | <0.001 |
| Abdominal/central obesity (%) ATPIII 1 | 47.79 | 40.23 | <0.001 |
| Abdominal/central obesity (%) IDFEURO 2 | 69.85 | 56.35 | <0.001 |
| Abdominal/central obesity (%) IDFASIA 3 | 75.47 | 59.16 | <0.001 |
| SNP | Gene † | Risk Allele | Phenotype | HG (N = 1496) | HR (N = 1141) | p-Value ‡ | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| β | SE | p-Value | β | SE | p-Value | |||||
| rs1121980 | FTO | A | BMI | 0.553 | 0.205 | 0.007 | 0.604 | 0.356 | 0.090 | 0.888 |
| WC | 1.516 | 0.472 | 0.001 | 1.682 | 0.752 | 0.025 | 0.679 | |||
| rs1558902 | FTO | A | BMI | 0.482 | 0.205 | 0.019 | 0.628 | 0.360 | 0.081 | 0.697 |
| WC | 1.318 | 0.472 | 0.005 | 1.513 | 0.758 | 0.046 | 0.730 | |||
| rs9939609 | FTO | A | BMI | 0.515 | 0.205 | 0.012 | 0.756 | 0.357 | 0.035 | 0.525 |
| WC | 1.333 | 0.472 | 0.005 | 1.596 | 0.754 | 0.035 | 0.618 | |||
| rs9941349 | FTO | T | BMI | 0.618 | 0.206 | 0.003 | 0.770 | 0.357 | 0.031 | 0.666 |
| WC | 1.567 | 0.474 | 0.001 | 1.546 | 0.754 | 0.040 | 0.801 | |||
| rs1801282 | PPARγ | C | BMI | −0.005 | 0.314 | 0.988 | −1.188 | 0.807 | 0.141 | 0.177 |
| WC | 0.340 | 0.724 | 0.638 | −3.507 | 1.699 | 0.039 | 0.029 * | |||
| Hungarian Roma | |||||
|---|---|---|---|---|---|
| Genetic Model | Genotype | Difference (95%CI) | p-Value | AIC ** | BIC ** |
| Codominant | C/C | ref. | 0.140 | 8928.1 | 8957.9 |
| C/G | 3.44 (–0.05 –6.93) | ||||
| G/G | 4.72 (–4.20 –23.65) | ||||
| Dominant | C/C | ref. | 0.048 | 8926.1 | 8950.9 |
| C/G and G/G | 3.48 (0.04 –6.92) | ||||
| Recessive | C/C and C/G | ref. | 0.650 | 8929.9 | 8954.7 |
| G/G | 4.38 (–14.57 –23.32) | ||||
| Over-dominant | C/C and G/G | ref. | 0.054 | 8926.3 | 8951.2 |
| C/G | 3.43 (–0.06 –6.91) | ||||
| Hungarian General | |||||
| Genetic Model | Genotype | Difference (95%CI) | p-Value | AIC ** | BIC ** |
| Codominant | C/C | ref. | 0.870 | 11812.4 | 11844.1 |
| C/G | –0.25 (–1.86 –1.35) | ||||
| G/G | –1.21 (–6.60 –4.19) | ||||
| Dominant | C/C | ref. | 0.690 | 11810.5 | 11837 |
| C/G and G/G | –0.32 (–1.88 –1.25) | ||||
| Recessive | C/C and C/G | ref. | 0.680 | 11810.4 | 11836.9 |
| G/G | –1.15 (–6.53 –4.23) | ||||
| Over-dominant | C/C and G/G | ref. | 0.780 | 11810.5 | 11837 |
| C/G | –0.23 (–1.83 –1.37) | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Llanaj, E.; Pikó, P.; Nagy, K.; Rácz, G.; János, S.; Kósa, Z.; Fiatal, S.; Ádány, R. Applicability of Obesity-Related SNPs and Their Effect Size Measures Defined on Populations with European Ancestry for Genetic Risk Estimation among Roma. Genes 2020, 11, 516. https://doi.org/10.3390/genes11050516
Llanaj E, Pikó P, Nagy K, Rácz G, János S, Kósa Z, Fiatal S, Ádány R. Applicability of Obesity-Related SNPs and Their Effect Size Measures Defined on Populations with European Ancestry for Genetic Risk Estimation among Roma. Genes. 2020; 11(5):516. https://doi.org/10.3390/genes11050516
Chicago/Turabian StyleLlanaj, Erand, Péter Pikó, Károly Nagy, Gábor Rácz, Sándor János, Zsigmond Kósa, Szilvia Fiatal, and Róza Ádány. 2020. "Applicability of Obesity-Related SNPs and Their Effect Size Measures Defined on Populations with European Ancestry for Genetic Risk Estimation among Roma" Genes 11, no. 5: 516. https://doi.org/10.3390/genes11050516