Absence of Strong Genetic Linkage Disequilibrium between Single Nucleotide Polymorphisms (SNPs) in the Prion Protein Gene (PRNP) and the Prion-Like Protein Gene (PRND) in the Horse, a Prion-Resistant Species



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Genetic Analysis
2.3. Statistical Analysis
2.4. Analysis of Genetic Linkage between SNPs in the PRNP and PRND Genes
2.5. Measurement of Protein Functional Alterations Induced by Nonsynonymous SNPs
2.6. Physical Map Distance between the PRNP and PRND Genes among Several Species
3. Results
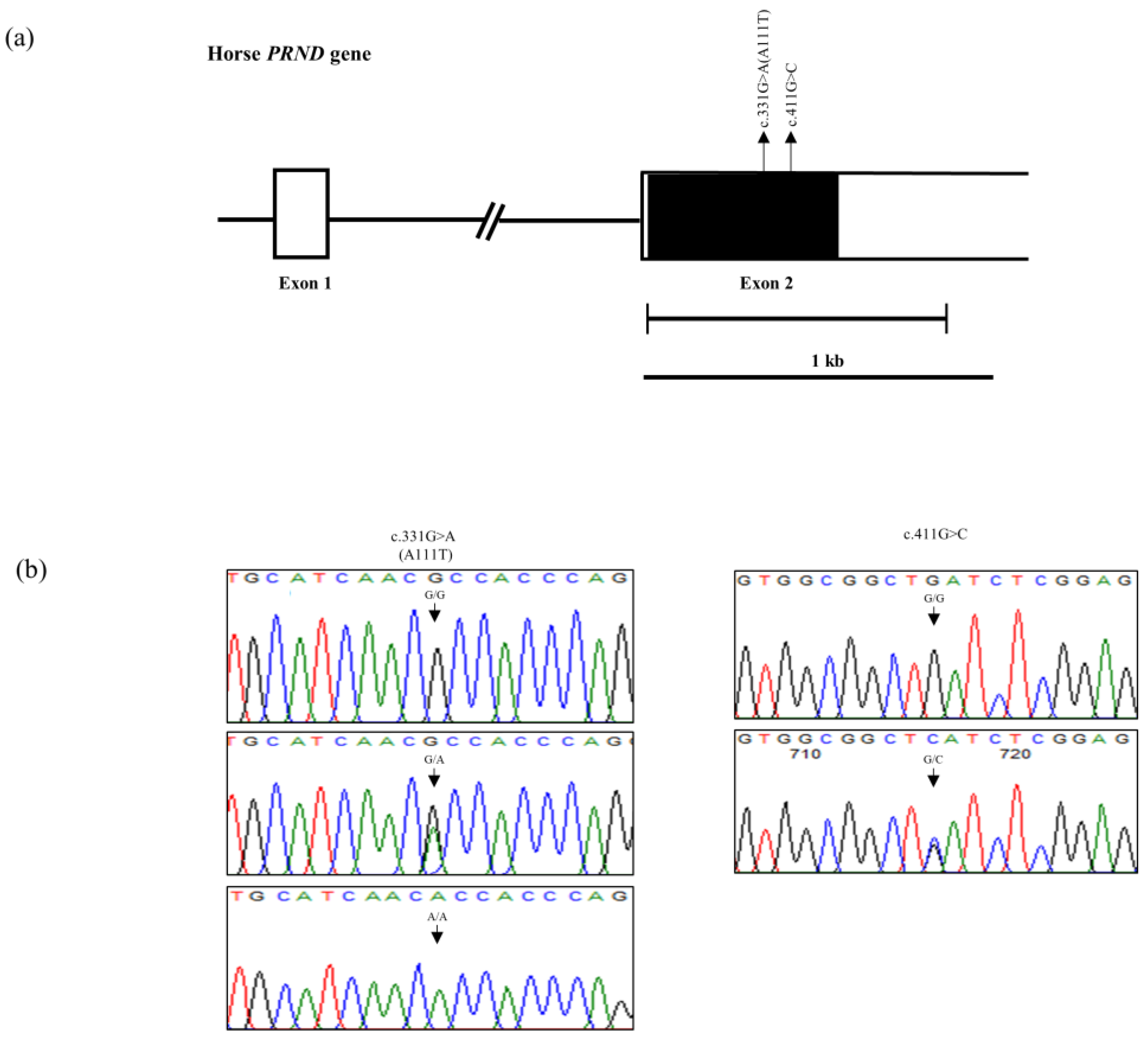
3.1. Investigation of Polymorphisms in the Horse PRND Gene in 140 Jeju and 80 Halla Horses
3.2. Analysis of Genetic Linkage between SNPs in the PRNP and PRND Genes
3.3. Measurement of the Physical Map Distance between the PRNP and PRND Genes among Several Species
3.4. Genetic Linkage between PRNP and PRND Genes among Several Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prusiner, S.B. Prions. Proc. Natl. Acad. Sci. USA 1998, 95, 13363–13383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prusiner, S.B. The Priori Diseases. Brain Pathol. 2006, 8, 499–513. [Google Scholar] [CrossRef]
- DeArmond, S.J.; Bajsarowicz, K. PrPSc accumulation in neuronal plasma membranes links Notch-1 activation to dendritic degeneration in prion diseases. Mol. Neurodegener. 2010, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crozet, C.; Béranger, F.; Lehmann, S. Cellular pathogenesis in prion diseases. Veter. Res. 2008, 39, 44. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-C.; Jeong, B.-H. The first report of prion-related protein gene (PRNT) polymorphisms in goat. Acta Veter. Hung. 2017, 65, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-C.; Jeong, B.-H. First report of prion-related protein gene (PRNT) polymorphisms in cattle. Veter. Rec. 2018, 182, 717. [Google Scholar] [CrossRef]
- Collee, J.G.; Bradley, R. BSE: A decade on—Part I. Lancet 1997, 349, 636–641. [Google Scholar] [CrossRef]
- Lloyd, S.; Mead, S.; Collinge, J. Genetics of Prion Disease. Chem. Diagn. 2011, 305, 1–22. [Google Scholar] [CrossRef]
- Vaccari, G.; Panagiotidis, C.H.; Acin, C.; Peletto, S.; Barillet, F.; Acutis, P.L.; Bossers, A.; Langeveld, J.; Van Keulen, L.; Sklaviadis, T.; et al. State-of-the-art review of goat TSE in the European Union, with special emphasis on PRNP genetics and epidemiology. Veter. Res. 2009, 40, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-C.; Kim, S.-K.; Jeong, B.-H. Scrapie susceptibility-associated indel polymorphism of shadow of prion protein gene (SPRN) in Korean native black goats. Sci. Rep. 2019, 9, 15261–15267. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-C.; Jeong, B.-H. The first report of polymorphisms and genetic characteristics of the prion protein gene (PRNP) in horses. Prion 2018, 12, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Jeong, B.-H.; Lee, K.-H.; Kim, N.-H.; Jin, J.-K.; Kim, J.-I.; Carp, R.I.; Kim, Y.-S. Association of sporadic Creutzfeldt–Jakob disease with homozygous genotypes at PRNP codons 129 and 219 in the Korean population. Neurogenetics 2005, 6, 229–232. [Google Scholar] [CrossRef]
- Jeong, B.-H.; Kim, Y.-S. Genetic Studies in Human Prion Diseases. J. Korean Med Sci. 2014, 29, 623–632. [Google Scholar] [CrossRef] [Green Version]
- Belt, P.B.G.M.; Muileman, I.H.; Schreuder, B.E.C.; Ruijter, J.B.-D.; Gielkens, A.L.J.; Smits, M. Identification of five allelic variants of the sheep PrP gene and their association with natural scrapie. J. Gen. Virol. 1995, 76, 509–517. [Google Scholar] [CrossRef]
- Hunter, N.; Foster, J.D.; Goldmann, W.; Stear, M.J.; Hope, J.; Bostock, C. Natural scrapie in a closed flock of Cheviot sheep occurs only in specific PrP genotypes. Arch. Virol. 1996, 141, 809–824. [Google Scholar] [CrossRef]
- Ekateriniadou, L.V.; Panagiotidis, C.H.; Terzis, A.; Ploumi, K.; Triantafyllidis, A.; Deligiannidis, P.; Triantaphyllidis, C.; Sklaviadis, T. Genotyping for PrP gene polymorphisms in rare Greek breeds of sheep. Veter. Rec. 2007, 160, 194–195. [Google Scholar] [CrossRef] [PubMed]
- Goldmann, W.; Martin, T.; Foster, J.; Hughes, S.; Smith, G.; Hughes, K.; Dawson, M.; Hunter, N. Novel polymorphisms in the caprine PrP gene: A codon 142 mutation associated with scrapie incubation period. J. Gen. Virol. 1996, 77, 2885–2891. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K.; Kim, Y.-C.; Won, S.-Y.; Jeong, B.-H. Potential scrapie-associated polymorphisms of the prion protein gene (PRNP) in Korean native black goats. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baylis, M.; Goldmann, W. The genetics of scrapie in sheep and goats. Curr. Mol. Med. 2004, 4, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Calvo, P.; Garcia, C.; Espinosa, J.C.; Andréoletti, O.; Torres, J.M. Prion and prion-like diseases in animals. Virus Res. 2015, 207, 82–93. [Google Scholar] [CrossRef]
- Fernández-Borges, N.; Eraña, H.; Castilla, J. Behind the potential evolution towards prion resistant species. Prion 2018, 12, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.; Fernández-Borges, N.; Eraña, H.; Parra, B.; Pintado, B.; Sánchez-Martín, M.A.; Charco, J.M.; Ordóñez, M.; Pérez-Castro, M.A.; Pumarola, M.; et al. Dogs are resistant to prion infection, due to the presence of aspartic or glutamic acid at position 163 of their prion protein. FASEB J. 2020, 34, 3969–3982. [Google Scholar] [CrossRef] [PubMed]
- Perez, D.R.; Damberger, F.F.; Wüthrich, K. Horse Prion Protein NMR Structure and Comparisons with Related Variants of the Mouse Prion Protein. J. Mol. Boil. 2010, 400, 121–128. [Google Scholar] [CrossRef]
- Sanchez-Garcia, J.; Fernandez-Funez, P. D159 and S167 are protective residues in the prion protein from dog and horse, two prion-resistant animals. Neurobiol. Dis. 2018, 119, 1–12. [Google Scholar] [CrossRef]
- Zhang, J. The Structural Stability of Wild-type Horse Prion Protein. J. Biomol. Struct. Dyn. 2011, 29, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Sigurdson, C.J.; Joshi-Barr, S.; Bett, C.; Winson, O.; Manco, G.; Schwarz, P.; Rulicke, T.; Nilsson, K.P.; Margalith, I.; Raeber, A.; et al. Spongiform encephalopathy in transgenic mice expressing a point mutation in the beta2-alpha2 loop of the prion protein. J. Neurosci. 2011, 31, 13840–13847. [Google Scholar] [CrossRef] [Green Version]
- Mesquita, P.; Batista, M.; Marques, M.D.R.; Santos, I.C.; Pimenta, J.; Pereira, M.S.; Carolino, I.; Silva, M.D.F.S.; Sousa, M.C.O.; Gama, L.T.; et al. Prion-like Doppel gene polymorphisms and scrapie susceptibility in portuguese sheep breeds. Anim. Genet. 2010, 41, 311–314. [Google Scholar] [CrossRef]
- Jeong, M.-J.; Kim, Y.-C.; Jeong, B.-H. Prion-like protein gene (PRND) polymorphisms associated with scrapie susceptibility in Korean native black goats. PLoS ONE 2018, 13, e0206209. [Google Scholar] [CrossRef]
- Won, S.-Y.; Kim, Y.-C.; Kim, K.; Kim, A.-D.; Jeong, B.-H. The First Report of Polymorphisms and Genetic Features of the prion-like Protein Gene (PRND) in a Prion Disease-Resistant Animal, Dog. Int. J. Mol. Sci. 2019, 20, 1404. [Google Scholar] [CrossRef] [Green Version]
- Jeong, M.-J.; Jeong, B.-H. No polymorphisms in the coding region of the prion-like protein gene in Thoroughbred racehorses. Acta Veter. Hung. 2019, 67, 174–182. [Google Scholar] [CrossRef]
- Yang, Y.H.; Kim, K.I.; Cothran, E.G.; Flannery, A.R. Genetic diversity of Cheju horses (Equus caballus) determined by using mitochondrial DNA D-loop polymorphism. Biochem. Genet. 2002, 40, 175–186. [Google Scholar] [CrossRef]
- Available online: http://allhorse.kra.co.kr/ked/index.jsp (accessed on 15 March 2019).
- Kim, Y.-C.; Jeong, B.-H. In Silico Evaluation of Acetylation Mimics in the 27 Lysine Residues of Human Tau Protein. Curr. Alzheimer Res. 2019, 16, 379–387. [Google Scholar] [CrossRef]
- Kim, Y.-C.; Jeong, M.-J.; Jeong, B.-H. Strong association of regulatory single nucleotide polymorphisms (SNPs) of the IFITM3 gene with influenza H1N1 2009 pandemic virus infection. Cell. Mol. Immunol. 2019, in press. [Google Scholar] [CrossRef]
- Adzhubei, I.; Jordan, D.M.; Sunyaev, S.R. Predicting Functional Effect of Human Missense Mutations Using PolyPhen-2. Curr. Protoc. Hum. Genet. 2013, 76, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-C.; Jeong, M.-J.; Jeong, B.-H. The first report of genetic variations in the chicken prion protein gene. Prion 2018, 12, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mead, S.; Mahal, S.P.; Beck, J.; Campbell, T.; Farrall, M.; Fisher, E.; Collinge, J. Sporadic—But Not Variant—Creutzfeldt-Jakob Disease Is Associated with Polymorphisms Upstream of PRNP Exon 1. Am. J. Hum. Genet. 2001, 69, 1225–1235. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Total, n | Genotype Frequency, n (%) | p-Value | Allele Frequency, n (%) | p-Value | HWE | Ref | |||
|---|---|---|---|---|---|---|---|---|---|---|
| GG | GA | AA | G | A | ||||||
| Jeju | 140 | 83 (59.3) | 52 (37.1) | 5 (3.6) | - | 218 (77.9) | 62 (22.1) | - | 0.360 | Current study |
| Halla | 80 | 78 (97.5) | 2 (2.5) | 0 (0) | <0.0001 | 158 (98.8) | 2 (1.2) | <0.0001 | 0.909 | Current study |
| Thoroughbred | 242 | 242 (100) | 0 (0) | 0 (0) | <0.0001 | 484 (100) | 0 (0) | <0.0001 | NA * | [30] |
| Breed | Total, n | Genotype Frequency, n (%) | p-Value | Allele Frequency, n (%) | p-Value | HWE | Ref | |||
|---|---|---|---|---|---|---|---|---|---|---|
| GG | GC | CC | G | C | ||||||
| Jeju | 140 | 130 (92.9) | 10 (7.1) | 0 (0) | - | 270 (96.4) | 10 (3.6) | - | 0.661 | Current study |
| Halla | 80 | 79 (98.8) | 1 (1.2) | 0 (0) | 0.06 | 159 (99.4) | 1 (0.6) | 0.0631 | 0.955 | Current study |
| Thoroughbred | 242 | 242 (100) | 0 (0) | 0 (0) | <0.0001 | 484 (100) | 0 (0) | 0.0022 | NA | [30] |
| Haplotype | Frequency | p-Value | |
|---|---|---|---|
| Jeju | Halla | ||
| GG | 208 (0.743) | 157 (0.981) | |
| AG | 62 (0.221) | 2 (0.013) | <0.0001 |
| GC | 10 (0.036) | 1 (0.006) | |
| Variation | Method | Score | Prediction |
|---|---|---|---|
| c.331G > A (A111T) | PolyPhen-2 | 0.006 | Benign |
| PROVEAN | −0.969 | Neutral | |
| PANTHER | 176 | Probably benign |
| Jeju | Halla | ||||
|---|---|---|---|---|---|
| c.331G > A (Codon 111) | c.411G > C (Codon 137) | c.331G > A (Codon 111) | c.411G > C (Codon 137) | ||
| c.331G > A (codon 111) | - | 1.0 | - | 1.0 | D’ |
| c.411G > C (codon 137) | 0.011 | - | 0 | - | r2 |
| PRNP | PRND | |||
|---|---|---|---|---|
| Jeju | Halla | |||
| c.331G > A (Codon 111) | c.411G > C (Codon 137) | c.331G > A (Codon 111) | c.411G > C (Codon 137) | |
| c.-3A > G | 0.005 | 0.002 | 0.025 | 0.001 |
| c.301T > A (codon 101) | 0.0 | 0.0 | 0.0 | 0.0 |
| c.525C > A (codon 175) | 0.089 | 0.003 | 0.01 | 0.005 |
| c.570G > A (codon 190) | 0.002 | 0.002 | 0.003 | 0.002 |
| Common Name | Scientific Name | Prion Disease Susceptibility | Method | Genetic Linkage | References |
|---|---|---|---|---|---|
| Human | Homo sapiens | Susceptible animal | Pairwise disequilibrium | Strong linkage | [37] |
| Sheep | Ovis aries | Susceptible animal | Haplotype analysis | Strong linkage | [27] |
| Goat | Capra hircus | Susceptible animal | Pairwise disequilibrium | Strong linkage | [28] |
| Dog | Canis lupus familiaris | Resistant animal | Pairwise disequilibrium | Weak linkage | [29] |
| Horse | Equs canbalus | Resistant animal | Pairwise disequilibrium | Weak linkage | Current study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Won, S.-Y.; Kim, Y.-C.; Do, K.; Jeong, B.-H. Absence of Strong Genetic Linkage Disequilibrium between Single Nucleotide Polymorphisms (SNPs) in the Prion Protein Gene (PRNP) and the Prion-Like Protein Gene (PRND) in the Horse, a Prion-Resistant Species. Genes 2020, 11, 518. https://doi.org/10.3390/genes11050518
Won S-Y, Kim Y-C, Do K, Jeong B-H. Absence of Strong Genetic Linkage Disequilibrium between Single Nucleotide Polymorphisms (SNPs) in the Prion Protein Gene (PRNP) and the Prion-Like Protein Gene (PRND) in the Horse, a Prion-Resistant Species. Genes. 2020; 11(5):518. https://doi.org/10.3390/genes11050518
Chicago/Turabian StyleWon, Sae-Young, Yong-Chan Kim, Kyoungtag Do, and Byung-Hoon Jeong. 2020. "Absence of Strong Genetic Linkage Disequilibrium between Single Nucleotide Polymorphisms (SNPs) in the Prion Protein Gene (PRNP) and the Prion-Like Protein Gene (PRND) in the Horse, a Prion-Resistant Species" Genes 11, no. 5: 518. https://doi.org/10.3390/genes11050518