Differences in Gene Expression Profiles of Seven Target Proteins in Third-Stage Larvae of Anisakis simplex (Sensu Stricto) by Sites of Infection in Blue Whiting (Micromesistius poutassou)

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Anisakis spp. Larvae Sampling
2.2. Total DNA/RNA Isolation and cDNA Synthesis
2.3. DNA Sequencing for Larval Identification
2.4. Gene Expression by Quantitative Real-Time PCR
2.5. Statistical Analyses
3. Results
3.1. Identification of Anisakis spp.
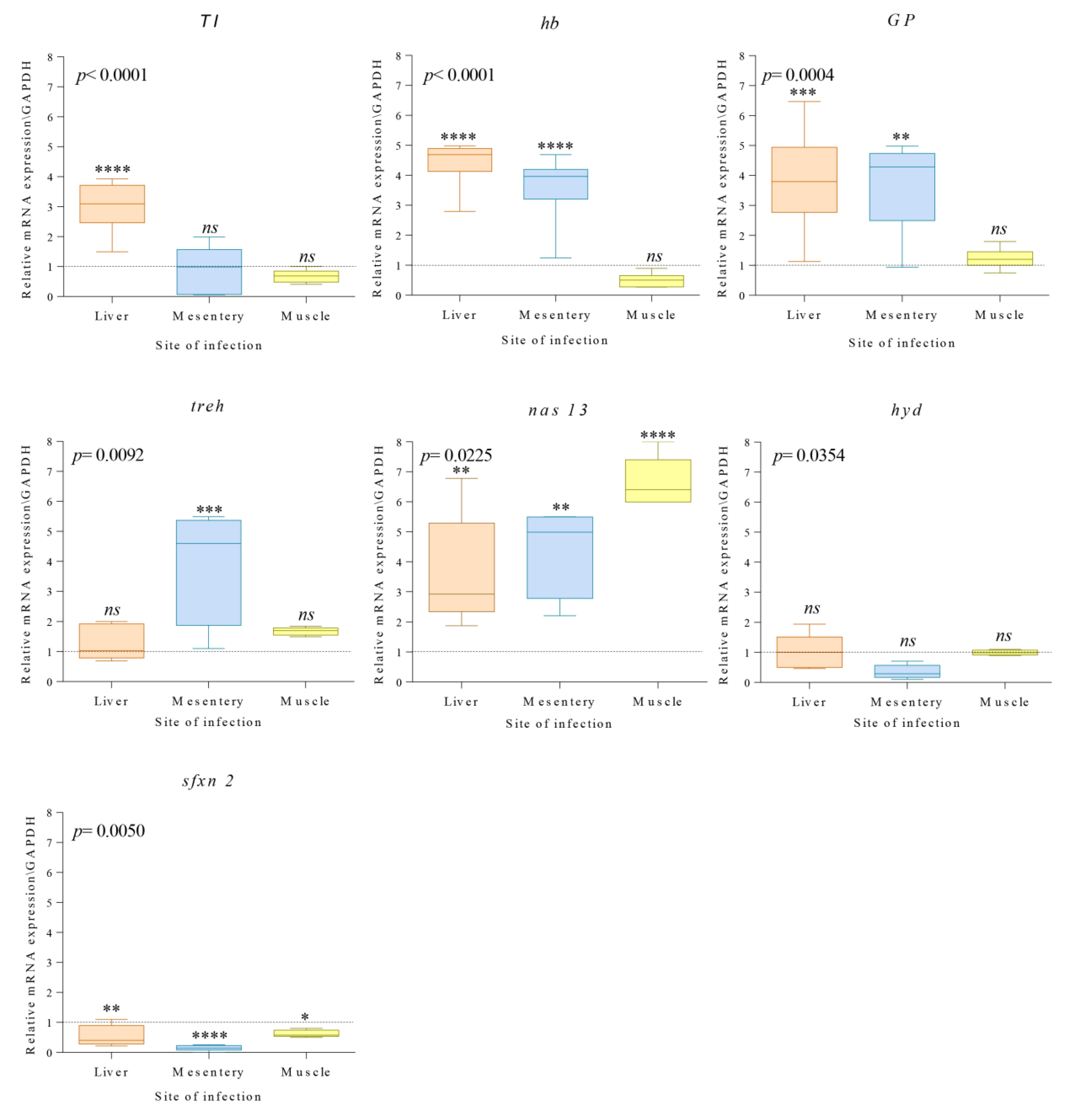
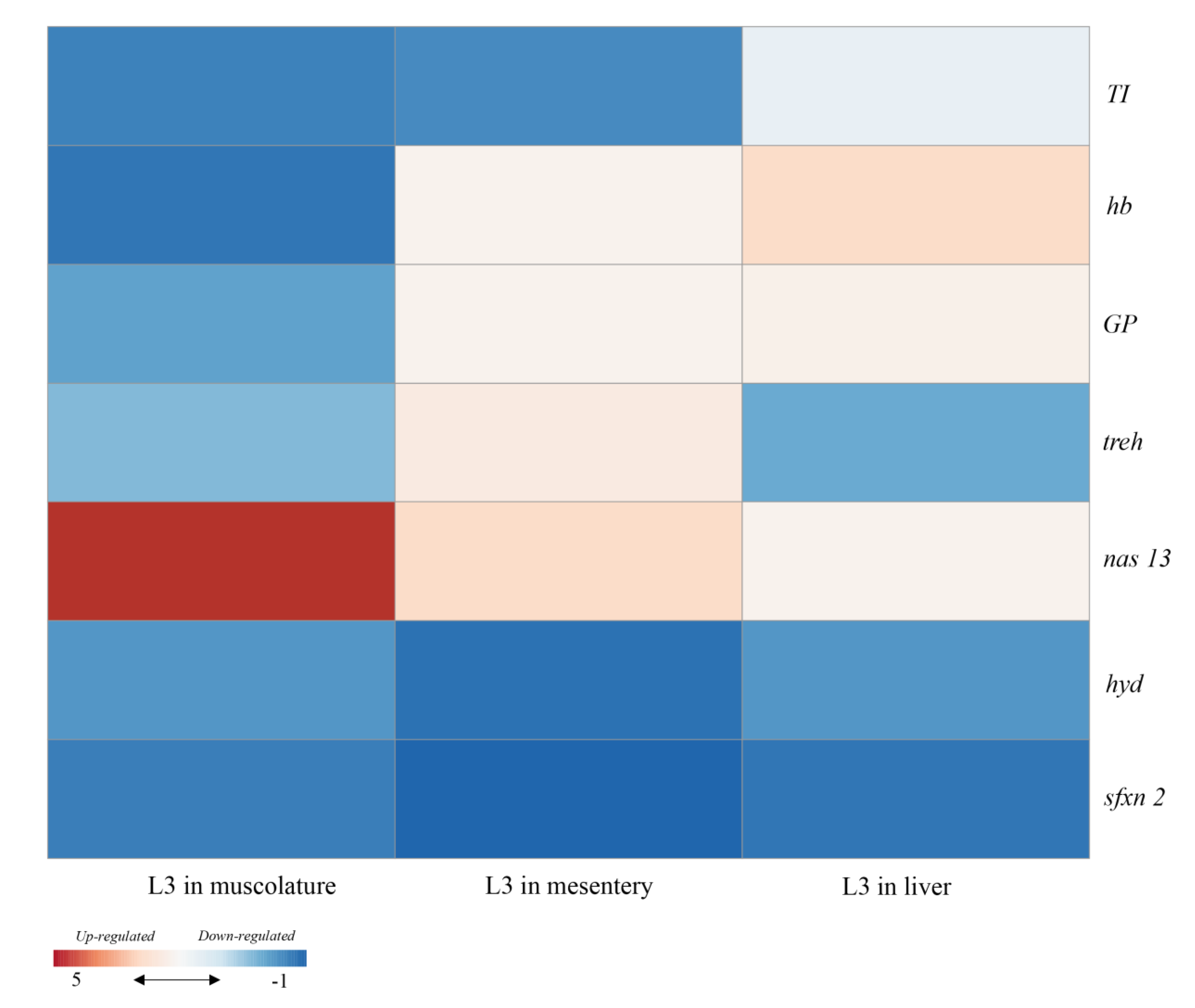
3.2. qPCR Analysis of Gene Coding for Seven Target Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular epidemiology of Anisakis and anisakiasis: An ecological and evolutionary road map. Adv. Parasitol. 2018, 99, 93–263. [Google Scholar] [PubMed]
- Mattiucci, S.; Nascetti, G. Advances and trends in the molecular systematics of anisakids nematodes, with implications for their evolutionary ecology and host-parasite co-evolutionary processes. Adv. Parasitol. 2008, 66, 47–148. [Google Scholar] [PubMed]
- Levsen, A.; Berland, B. Anisakis species. In Fish Parasites: Pathobiology and Protection; Patrick, T.K., Buchmann, K., Eds.; CABI Press: Wallingford, UK, 2012; p. 298. [Google Scholar]
- Smith, J.W. The abundance of Anisakis simplex L3 in the body-cavity and flesh of marine teleosts. Int. J. Parasitol. 1984, 14, 491–495. [Google Scholar] [CrossRef]
- Smith, J.W.; Wootten, R. Experimental studies on the migration of Anisakis sp. larvae (Nematoda: Ascaridida) into the flesh of herring, Clupea harengus L. Int. J. Parasitol. 1975, 5, 133–136. [Google Scholar] [PubMed]
- Karl, H.; Meyer, C.; Banneke, S.; Jark, U.; Feldhusen, F. The abundance of nematode larvae Anisakis sp. in the flesh of fishes and possible post-mortem migration. Archiv. für Lebensmittelhygiene 2002, 53, 118–120. [Google Scholar]
- Quiazon, K.M.; Yoshinaga, T.; Ogawa, K. Experimental challenge of Anisakis simplex sensu stricto and Anisakis pegreffii (Nematoda: Anisakidae) in rainbow trout and olive flounder. Parasitol. Int. 2011, 60, 126–131. [Google Scholar] [CrossRef]
- Cipriani, P.; Acerra, V.; Bellisario, B.; Sbaraglia, G.L.; Cheleschi, R.; Nascetti, G.; Mattiucci, S. Larval migration of the zoonotic parasite Anisakis pegreffii (Nematoda: Anisakidae) in European anchovy, Engraulis encrasicolus: Implications to seafood safety. Food Control 2016, 59, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Cipriani, P.; Sbaraglia, G.; Palomba, M.; Giulietti, L.; Bellisario, B.; Buselic, I.; Mladineo, I.; Cheleschi, R.; Nascetti, G.; Mattiucci, S. Anisakis pegreffii (Nematoda: Anisakidae) in European anchovy Engraulis encrasicolus from the Mediterranean Sea: Considerations in relation to fishing ground as a driver for parasite distribution. Fish. Res. 2017, 202, 59–68. [Google Scholar] [CrossRef]
- Bahlool, Q.Z.M.; Skovgaard, A.; Kania, P.; Haarder, S.; Buchmann, K. Microhabitat preference of Anisakis simplex in three salmonid species: Immunological implication. Vet. Parasitol. 2012, 190, 489–495. [Google Scholar] [CrossRef]
- Abollo, E.; Gestal, C.; Pascal, S. Anisakis infestation in marine fish and cephalopods from Galician waters: And updated perspective. Parasitol. Res. 2001, 87, 492–499. [Google Scholar]
- Dezfuli, B.S.; Giovinazzo, G.; Lui, A.; Giari, L. Inflammatory response to Dentitruncus truttae (Acanthocephala) in the intestine of brown trout. Fish Shellfish Immunol. 2008, 24, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Bahlool, Q.Z.; Skovgaard, A.; Kania, P.W.; Buchmann, K. Effects of excretory/secretory products from Anisakis simplex (Nematoda) on immune gene expression in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2013, 35, 734–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrdana, F.; Kania, P.W.; Nazemi, S.; Buchmann, K. Immunomodulatory effects of excretory/secretory compounds from Contracaecum osculatum larvae in a zebrafish inflammation model. PLoS ONE 2017, 12, e0181277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marnis, H.; Kania, P.W.; Syahputra, K.; Zuo, S.; Dirks, R.P.; Buchmann, K. Transcriptomic analysis of Baltic cod (Gadus morhua) liver infected with Contracaecum osculatum third stage larvae indicated parasitic effects on growth and immune response. Fish Shellfish Immunol. 2019, 93, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Priebe, K.; Huber, C.; Martlbauer, E.; Terplan, G. Detection of antibodies against the larva of Anisakis simplex in the pollock Pollachius virens using ELISA. J. Vet. Med. 1991, 38, 209–214. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, K.H.; Cho, S.; Park, H.Y.; Cho, S.W.; Kim, Y.T.; Joo, K.H.; Lee, J.S. Immunochemical and biological analysis of allergenicity with excretory-secretory products of Anisakis simplex third stage larva. Int. Arch. Allergy Immunol. 2005, 136, 320–328. [Google Scholar] [CrossRef]
- Kuhn, T.; Hailer, F.; Palm, H.W.; Klimpel, S. Global assessment of molecularly identified Anisakis Dujardin, 1845 (Nematoda: Anisakidae) in their teleost intermediate hosts. Folia Parasitol. 2013, 60, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Skrzypczak, M.; Rokicki, J.; Pawliczka, I.; Najda, K.; Dzido, J. Anisakids of seals found on the Southern coast of Baltic Sea. Acta Parasitol. 2014, 59, 165–172. [Google Scholar] [CrossRef]
- Mehrdana, F.; Buchmann, K. Excretory/secretory products of anisakid nematodes: Biological and pathological roles. Acta Vet. Scand. 2017, 59, 42. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, J.O.; Jeon, C.H.; Nam, U.H.; Subramaniyan, S.; Yoo, S.I.; Park, J.H. Comparative transcriptome analyses of the third and fourth stage larvae of Anisakis simplex (Nematoda: Anisakidae). Mol. Biochem. Parasitol. 2018, 226, 24–33. [Google Scholar] [CrossRef]
- Cavallero, S.; Lombardo, F.; Su, X.; Salvemini, M.; Cantacessi, C.; D’Amelio, S. Tissue-specific transcriptomes of Anisakis simplex (sensu stricto) and Anisakis pegreffii reveal potential molecular mechanisms involved in pathogenicity. Parasites Vectors 2018, 11, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavallero, S.; Lombardo, F.; Salvemini, M.; Pizzarelli, A.; Cantacessi, C.; D’Amelio, S. Comparative transcriptomics reveals clues for differences in pathogenicity between Hysterothylacium aduncum, Anisakis simplex sensu stricto and Anisakis pegreffii. Genes 2020, 11, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopienska-Biernat, E.; Paukszto, L.; Jastrzebski, J.P.; Myszczynski, K.; Polak, I.; Stryinski, R. Genome-wide analysis of Anisakis simplex sensu latu: The role of carbohydrate metabolism genes in the parasite’s development. Int. J. Parasitol. 2019, 49, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Palomba, M.; Paoletti, M.; Colantoni, A.; Rughetti, A.; Nascetti, G.; Mattiucci, S. Gene expression profiles of antigenic proteins of third stage larvae of the zoonotic nematode Anisakis pegreffii in response to temperature conditions. Parasite 2019, 26, 52. [Google Scholar] [CrossRef] [Green Version]
- Baird, F.J.; Su, X.; Aibinu, I.; Nolan, M.J.; Suglyama, H.; Otranto, D.; Lopata, A.L.; Cantacessi, C. The Anisakis transcriptome provides a resource for fundamental and applied studies on allergy-causing parasites PLOS Neglect. Trop. Dis. 2016, 10, e0004845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prokopchuk, I.; Sentyabov, E. Diets of herring, mackerel and blue whiting in the Norwegian Sea in relation to Calanus finmarchius distribution and temperature conditions. ICES J. Mar. Sci. 2006, 63, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Mattiucci, S.; Cipriani, P.; Webb, S.C.; Paoletti, M.; Marcer, F.; Bellisario, B.; Gibson, D.I.; Nascetti, G. Genetic and morphological approaches distinguishing the three sibling species of the Anisakis simplex species complex, with a species designation as Anisakis berlandi n. sp. for A. simplex sp. C (Nematoda: Anisakidae). J. Parasitol. 2014, 15, 12–15. [Google Scholar]
- Mattiucci, S.; Acerra, V.; Paoletti, M.; Cipriani, P.; Levsen, A.; Webb, S.C.; Canestrelli, D.; Nascetti, G. No more time to stay ‘single’ in the detection of Anisakis pegreffii, A. simplex (s.s.) and hybridization events between them: A multi-marker nuclear genotyping approach. Parasitology 2016, 143, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Cheng, Y.S.; Grabner, D.S.; Chang, S.H.; Shih, H.H. Effect of different temperatures on the expression of the newly characterized heat shock protein 90 (Hsp90) in L3 of Anisakis spp. isolated from Scomber australasicus. Vet. Parasitol. 2014, 205, 540–550. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time quantitative PCR and the 2 (Delta CT) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Molina-Fernandez, D.; Rubio-Calvo, D.; Adroher, F.J.; Benitez, R. Molecular epidemiology of Anisakis spp. in blue whiting Micromesistius poutassou in eastern waters of Spain, western Mediterranean Sea. Int. J. Food Microbiol. 2018, 282, 49–56. [Google Scholar] [PubMed]
- Gomez-Mateos, M.; Valero, A.; Morales-Yuste, M.; Martin-Sanchez, J. Molecular epidemiology and risk factors for Anisakis simplex s. l. infection in blue whiting (Micromesistius poutassou) in a confluence zone of the Atlantic and Mediterranean: Differences between A. simplex s.s. and A. pegreffii. Int. J. Food Microbiol. 2016, 232, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Levsen, A.; Svanevik, C.S.; Cipriani, P.; Mattiucci, S.; Gay, M.; Hastiee, L.C.; Pierce, G.J.; Bušelić, I.; Mladineo, I.; Karl, O.; et al. A survey of zoonotic nematodes of commercial key fish species from major European fishing grounds introducing the FP7 PARASITE exposure assessment study. Fish. Res. 2018, 202, 4–21. [Google Scholar] [CrossRef]
- McKenzie, K. Some parasites and diseases of blue whiting, Micromesistius poutassou (Risso), to the North and West of Scotland and at the Faroe Islands. Scottish Fish. Res. Rep. 1979, 17, 1–14. [Google Scholar]
- Højgaard, D.P. Sníkar í svartkjafti (Parasites of Blue Whiting). Fiskirannsóknir 1998, 8, 114–126, (In Faroese, with English summary). [Google Scholar]
- Berland, B. Musings on onematode parasites. Fisken Oghavet 2006, 11, 1–26. [Google Scholar]
- Karl, H. Nematode larvae in fish on the German market—20 years of consumer related research. Arch. Lebensmittelhyg 2008, 59, 107–116. [Google Scholar]
- Cipriani, P.; Giulietti, L.; Palomba, M.; Mattiucci, S.; Bao, M.; Levesen, A. Occurrence of larval ascaridoid nematodes in the Argentinean short-finned squid Illex argentinus from the Southwest Atlantic Ocean (off Falkland Islands). Int. J. Food. Microbiol. 2019, 297, 27–31. [Google Scholar] [CrossRef]
- Suzuki, J.; Murata, R.; Hosaka, M.; Araki, J. Risk factors for human Anisakis infection and association between the geographic origins of Scomber japonicus and anisakid nematodes. Int. J. Food Microbiol. 2010, 137, 88–93. [Google Scholar] [CrossRef]
- Cipriani, P.; Smaldone, G.; Acerra, V.; D’Angelo, L.; Anastasio, A.; Bellisario, B.; Palma, G.; Nascetti, G.; Mattiucci, S. Genetic identification and distribution of the parasitic larvae of Anisakis pegreffii and A. simplex (s.s.) in European hake Merluccius merluccius from the Tyrrhenian Sea and Spanish Atlantic coast: Implications for food safety. Int. J. Food Microbiol. 2014, 198, 1–8. [Google Scholar] [CrossRef]
- Agius, C.; Roberts, R.J. Melano-macrophage centres and their role in fish pathology. J. Fish Dis. 2003, 26, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Larsen, A.H.; Bresciani, J.; Buchmann, K. Interactions between ecto- and endoparasites in trout Salmo trutta. Vet. Parasitol. 2002, 103, 167–173. [Google Scholar] [CrossRef]
- Rhoads, M.L.; Fetterer, R.H.; Hill, D.E. Trichuris suis: A secretory serine protease inhibitor. Exp. Parasitol. 2000, 94, 1–7. [Google Scholar] [CrossRef]
- Stepek, G.; McCormack, G.; Page, A.P. Collagen processing and cuticle formation is catalysed by the astacin metalloprotease DPY-31 in free-living and parasitic nematodes. Int. J. Parasitol. 2010, 40, 533–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moneo, I.; Caballero, M.L.; Gómez, F.; Ortega, E.; Alonso, M.J. Isolation and characterization of a major allergen from the fish parasite Anisakis simplex. J. Allergy Clin. Immunol. 2000, 106, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Bernard, V.D.; Peanasky, R.J. The serine protease inhibitor family from Ascaris suum: Chemical determination of the five disulphide bridges. Arch. Biochem. Biophys. 1993, 303, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Stanssens, P.; Bergum, P.W.; Gansemans, Y.; Jespers, L.; Laroche, Y.; Huang, S.; Maki, S.; Messens, J.; Lauwereys, M.; Cappello, M.; et al. Anticoagulant repertoire of the hookworm Ancylostoma caninum. Proc. Natl. Acad. Sci. USA 1996, 93, 2149–2154. [Google Scholar] [CrossRef] [Green Version]
- Matthews, B.E. Behaviour and enzyme release by Anisakis sp. larvae (Nematoda: Ascaridida). J. Helminthol. 1982, 56, 177–183. [Google Scholar] [CrossRef]
- Matthews, B.E. The source, release and specificity of proteolytic enzyme activity produced by Anisakis simplex larvae (Nematoda: Ascaridida) in vitro. J. Helminthol. 1984, 58, 175–185. [Google Scholar] [CrossRef]
- Quiazon, K.M.; Santos, M.D.; Yoshinaga, T. Anisakis species (Nematoda: Anisakidae) of Dwarf Sperm Whale Kogia sima (Owen, 1866) stranded off the Pacific coast of southern Philippine archipelago. Vet. Parasitol. 2013, 197, 221–230. [Google Scholar] [CrossRef]
- Hawley, J.; Peanasky, R.J. Ascaris suum: Are trypsin inhibitors involved in species specificity of Ascarid nematodes? Exp. Parasitol. 1992, 75, 112–128. [Google Scholar] [CrossRef]
- Khangembam, B.K.; Sharma, K.; Chakrabarti, R. Purification and characterization of trypsin from the digestive system of carp Catla catla (Hamilton). Int. Aquat. Res. 2012, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Klomklao, S.; Benjakul, S. Two trypsin isoforms from albacore tuna (Thunnus alalonga) liver: Purification and physicochemical and biochemical characterization. Int. J. Biol. Macro. 2018, 107, 1864–1870. [Google Scholar] [CrossRef] [PubMed]
- McNeil, H.P. Mast cell-restricted tryptases: Structure and function in inflammation and pathogen defense. J. Biol. Chem. 2007, 282, 20785–20789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Fernandez, J.; Daschner, A.; Nieuwenhuizen, N.E.; Lopata, A.L.; Frutos, C.D.; Valls, A.; Cuellar, C. Haemoglobin, a new major allergen of Anisakis simplex. Int. J. Parasitol. 2015, 45, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Abe, N.; Teramoto, I. Anisakis haemoglobin is a main antigen indicung strong and prolonged immunoreactions in rats. Parasitol. Res. 2017, 116, 2035–2039. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, J.; Rivas, L.; Luque-Ortega, J.R.; Nunez-Ramirez, R.; Campioli, P.; Garate, T.; Perteguer, M.J.; Daschner, A.; Cuellar, C. Recombinant vs native Anisakis haemoglobin (Ani s 13): Its appraisal as a new gold standard for the diagnosis of allergy. Exp. Parasitol. 2017, 181, 119–129. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.E.; Meter, J.M.; Horsnell, W.G.; Hoving, J.C.; Fick, L.; Sharp, M.F.; Darby, M.G.; Parihar, S.P.; Brombacher, F.; Lopata, A.L. A cross-reactive monoclonal antibody to nematode haemoglobin enhances protective immune responses to Nippostrongylus brasiliensis. PLoS Negl. Trop. Dis. 2013, 7, e2395. [Google Scholar] [CrossRef] [Green Version]
- Secombes, C.J.; Wang, T. The Innate and Adaptive Immune System of Fish. In Infectious Disease in Aquaculture: Prevention and Control; Austin, B., Ed.; Woodhead Publishing Limited: Aberdeen, UK, 2012; pp. 1–67. [Google Scholar]
- Prasanphanich, N.S.; Mickum, M.L.; Molinaro, J.H.; Cummings, R.D. Glycoconjugates in host-helminth interactions. Front. Immunol. 2013, 4, 240. [Google Scholar] [CrossRef] [Green Version]
- White, R.R.; Artavanis-Tsakonas, K. How helminths use excretory secretory fractions to modulate dendritic cells. Virulence 2012, 3, 668–677. [Google Scholar] [CrossRef] [Green Version]
- Steinfelder, S. The major component in schistosome eggs responsible for conditioning dendritic cells for Th2 polarization is a T2 ribonuclease (omega-1). J. Exp. Med. 2009, 206, 1681–1690. [Google Scholar] [CrossRef] [PubMed]
- Hrabar, J.; Trumbic, Z.; Bocina, I.; Buselic, I.; Vrbatovic, A.; Mladineo, I. Interplay between proinfiammatory cytokines, miRNA, and tissue lesions in Anisakis-infected Sprague-Dawley rats. PLoS Neglect. Trop. Dis. 2019, 13, e0007397. [Google Scholar] [CrossRef] [PubMed]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003 13, 17R–27R. [CrossRef]
- Tang, B.; Wang, S.; Wang, S.G.; Wang, H.J.; Zhang, J.Y.; Cui, S.Y. Invertebrate trehalose-6-phosphate synthase gene: Genetic architecture, biochemistry, physiological function, and potential applications. Front. Physiol. 2018, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łopienska-Biernat, E.; Zaobidna, E.A.; Dmitryjuk, M. Expression of genes encoding the enzymes for glycogen and trehalose metabolism in L3 and L4 larvae of Anisakis simplex. J. Parasitol. Res. 2015, 2015, 438145. [Google Scholar] [CrossRef] [Green Version]
- Łopienska-Biernat, E.; Stryinski, R.; Dmitryjuk, M.; Wasilewska, B. Infective larvae of Anisakis simplex (Nematoda) accumulate trehalose and glycogen in response to starvation and temperature stress. Biol. Open 2019, 8, bio040014. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.W.; Wootten, R. Anisakis and anisakiasis. Adv. Parasitol. 1978, 16, 93–163. [Google Scholar]
- Grabda, J. Studies on the life cycle and morphogenesis of Anisakis simplex (Rudolphi, 1809) (Nematoda: Anisakidae) cultured in vitro. Acta Ichthyol. Piscat. 1976, 6, 119–141. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, L.; Valero, A.; Benítez, R.; Adroher, F.J. In vitro cultivation of Anisakis simplex: Pepsin increases survival and moulting from fourth larval to adult stage. Parasitology 2001, 1123, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Łopienska-Biernat, E.; Zółtowska, K.; Rokicki, J. The content of carbohydrates in larval stages of Anisakis simplex (Nematoda, Anisakidae). Helmintologia 2006, 43, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Levsen, A.; Lunestad, B.T.; Berland, B. Parasites in farmed fish and fishery products. In Improving Farmed Fish Quality and Safety; Oyvind, L., Ed.; WP: New York, NY, USA, 2008; pp. 428–445. [Google Scholar]
- Sajid, M.; McKerrow, J.H. Cysteine proteases of parasitic organism. Mol. Biochem. Parasit. 2002, 120, 1–21. [Google Scholar] [CrossRef]
- Malagon, D.; Benitez, R.; Kasny, M.; Adroher, F.J. Peptidases in parasitic nematodes: A review. In Parasites: Ecology, Diseases and Management; Erzinger, G.S., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2013; pp. 61–102. [Google Scholar]
- Hotez, P.J.; Pritchard, D.I. Hookworm infection. Sci. Am. 1995, 272, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Todorova, V.K.; Stoyanov, D.I. Partial characterization of serine proteinases secreted by adult Trichinella spiralis. Parasitol. Res. 2000, 86, 684–687. [Google Scholar] [CrossRef] [PubMed]
- Malagón, D.; Benítez, R.; Adroher, F.J.; Díaz-López, M. Proteolytic activity in Hysterothylacium aduncum (Nematoda: Anisakidae), a fish gastrointestinal parasite of worldwide distribution. Vet. Parasitol. 2011, 183, 95–102. [Google Scholar] [CrossRef]
- Bruschi, F.; Pinto, B. Metalloproteinases in Parasitic Infections. In Pathophysiological Aspects of Proteases; Chakraborti, S., Dhalla, N., Eds.; Springer: Singapore, 2017; pp. 61–102. [Google Scholar]
- Adhikari, B.N.; Adams, B.J. Molecular Analyses of Desiccation Survival in Antarctic Nematodes. In Molecular and Physiological Basis of Nematode Survival; Perry, R.N., Wharton, D.A., Eds.; CAB International: Oxford, UK, 2011; pp. 205–232. [Google Scholar]
- Shang, F.; Gong, X.; Taylor, A. Activity of ubiquitin-dependent pathway in response to oxidative stress: Ubiquitin-activating enzye (E1) is transiently upregulated. J. Biol. Chem. 1997, 276, 23086–23093. [Google Scholar] [CrossRef] [Green Version]
- Miotto, G.; Tessaro, S.; Rotta, G.A.; Bonatto, D. In silico analyses of Fsf1 sequences, a new group of fungal proteins orthologous to the metazoan sideroblastic anemia-related sideroflexin family. Fungal. Genet. Biol. 2007, 44, 740–753. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, w566–w570. [Google Scholar] [CrossRef]
- Read, A.; Sharping, A. The evolution of tissue migration by parasitic nematode larvae. Parasitology 1995, 111, 359–371. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene Locus | Sequence (5′ to 3′) | Ta/°C | Product Size (bp) | Reference |
|---|---|---|---|---|
| TI | CATGTGCCGATAAATGCGGG | 57 °C | 130 | [25] |
| CCCTGTGAGCATGCATCCTT | ||||
| hb | AAACATTCGACGCCTACACC | 60 °C | 108 | [25] |
| CATCGTGGTCTTCTCTGCGA | ||||
| GP | TATCGGAATGCGTGACTGCA | 57 °C | 130 | [25] |
| AGGCAGTTTCCATGGTGTATG | ||||
| treh | TCAGCAAGCATTTGAGTGAAGAGT | 60 °C | 117 | [21] |
| TAACATGATTGAAAACTTCGCAACA | ||||
| nas 13 | AGCAATAGCAGCACGATGA | 60 °C | 131 | [21] |
| CTGCGGTAGCCAATGCTTTT | ||||
| hyd | CCATCCAGTGAAGAAGGATTCC | 60 °C | 116 | [21] |
| GAATAGAGCGGTAAGTAGAGCCTTGA | ||||
| sfxn 2 | TTAGAATGGCGTTGAAGCAGTAGTAG | 60 °C | 130 | [21] |
| AGTATCGGTTCTGACCAGTTTTTTG | ||||
| gpd | CCCCTTCATCAACATCGACT | 60 °C | 152 | [30] |
| TCAGCTCCCCATTTGATTTC |
| Gene Locus | Liver vs. Mesentery | Liver vs. Muscle | Mesentery vs. Muscle |
|---|---|---|---|
| TI | **** | **** | ns |
| hb | ns | **** | **** |
| GP | ns | *** | ** |
| treh | * | ns | * |
| nas 13 | ns | * | ns |
| hyd | * | ns | ns |
| sfxn 2 | * | ns | **** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palomba, M.; Cipriani, P.; Giulietti, L.; Levsen, A.; Nascetti, G.; Mattiucci, S. Differences in Gene Expression Profiles of Seven Target Proteins in Third-Stage Larvae of Anisakis simplex (Sensu Stricto) by Sites of Infection in Blue Whiting (Micromesistius poutassou). Genes 2020, 11, 559. https://doi.org/10.3390/genes11050559
Palomba M, Cipriani P, Giulietti L, Levsen A, Nascetti G, Mattiucci S. Differences in Gene Expression Profiles of Seven Target Proteins in Third-Stage Larvae of Anisakis simplex (Sensu Stricto) by Sites of Infection in Blue Whiting (Micromesistius poutassou). Genes. 2020; 11(5):559. https://doi.org/10.3390/genes11050559
Chicago/Turabian StylePalomba, Marialetizia, Paolo Cipriani, Lucilla Giulietti, Arne Levsen, Giuseppe Nascetti, and Simonetta Mattiucci. 2020. "Differences in Gene Expression Profiles of Seven Target Proteins in Third-Stage Larvae of Anisakis simplex (Sensu Stricto) by Sites of Infection in Blue Whiting (Micromesistius poutassou)" Genes 11, no. 5: 559. https://doi.org/10.3390/genes11050559