GPR174 and ITM2A Gene Polymorphisms rs3827440 and rs5912838 on the X chromosome in Korean Children with Autoimmune Thyroid Disease

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. DNA Extraction
2.3. Target Gene Primer Design and Multiplex PCR
2.4. Sequencing
2.5. Data Analysis
- (1)
- (2)
- The sequence alignment/map format (BAM) dataset was uploaded in the Galaxy website.
2.6. Statistical Analysis
3. Results
3.1. Comparison of Genotype and Allele Frequencies of GPR174 and ITM2A SNPs on the X chromosome in AITD Patients and Controls
3.2. Comparison of Genotype and Allele Frequencies of GPR174 and ITM2A SNPs on X chromosome in GD Patients with or without TAO and Controls
3.3. Comparison of Genotype and Allele Frequencies of GPR174 and ITM2A SNPs on X chromosome in HD Patients and Controls
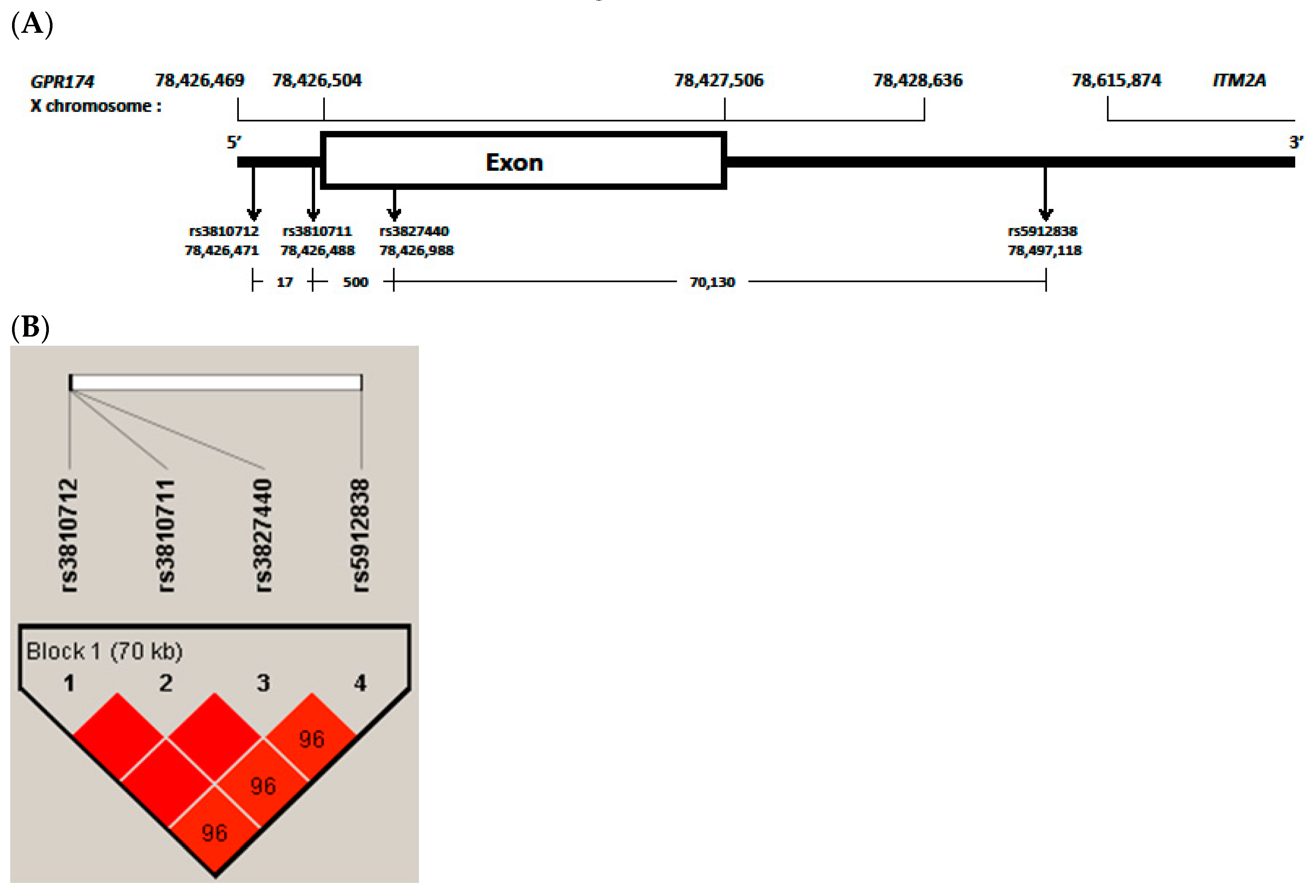
3.4. LD and Haplotype Analysis of the Four GPR174 and ITM2A SNPs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tomer, Y.; Davies, T.F. Searching for the autoimmune thyroid disease susceptibility genes: from gene mapping to gene function. Endocr. Rev. 2003, 24, 694–717. [Google Scholar] [CrossRef] [Green Version]
- Brix, T.H.; Kyvik, K.O.; Christensen, K.; Hegedüs, L. Evidence for a major role of heredity in Graves’ disease: a population-based study of two Danish twin cohorts. J. Clin. Endocrinol. Metab. 2001, 86, 930–934. [Google Scholar] [CrossRef] [PubMed]
- Stassi, G.; De Maria, R. Autoimmune thyroid disease: new models of cell death in autoimmunity. Nat. Rev. Immunol. 2002, 2, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Huh, K.B.; Lee, H.C.; Kim, H.M.; Lee, H.R.; Hong, C.S.; Lee, S.Y.; Choi, H.J.; Park, K.; Kim, C.K. Human leukocyte antigen (HLA) in Korean patients with autoimmune thyroid diseases. Korean J. Int. Med. 1986, 1, 243–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, K.; Tamai, H.; Morita, T.; Kuma, K.; Nishimura, Y.; Sasazuki, T. Hashimoto’s thyroiditis and HLA in Japanese. J. Clin. Endocrinol. Metab. 1989, 69, 1268–1273. [Google Scholar] [CrossRef]
- Webb, R.; Kelly, J.A.; Somers, E.C.; Hughes, T.; Kaufman, K.M.; Sanchez, E.; Nath, S.K.; Bruner, G.; Alarcon-Riquelme, M.E.; Gilkeson, G.S.; et al. Early disease onset is predicted by a higher genetic risk for lupus and is associated with a more severe phenotype in lupus patients. Ann. Rheumatic Diseases 2011, 70, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Cho, W.K.; Jung, M.H.; Choi, E.J.; Choi, H.B.; Kim, T.G.; Suh, B.K. Association of HLA alleles with autoimmune thyroid disease in Korean children. Horm. Res. Paediatr. 2011, 76, 328–334. [Google Scholar] [CrossRef]
- Cho, B.Y.; Rhee, B.D.; Lee, D.S.; Lee, M.S.; Kim, G.Y.; Lee, H.K.; Koh, C.S.; Min, H.K.; Lee, M. HLA and Graves’ disease in Koreans. Tissue Antigens 1987, 30, 119–121. [Google Scholar] [CrossRef]
- Libert, C.; Dejager, L.; Pinheiro, I. The X chromosome in immune functions: when a chromosome makes the difference. Nat. Rev. Immunol. 2010, 10, 594–604. [Google Scholar] [CrossRef]
- Whitacre, C.C. Sex differences in autoimmune disease. Nat. Immunol. 2001, 2, 777–780. [Google Scholar] [CrossRef]
- Dai, R.; Ahmed, S.A. Sexual dimorphism of miRNA expression: a new perspective in understanding the sex bias of autoimmune diseases. Therapeutics Clin. Risk Manag. 2014, 10, 151–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fish, E.N. The X-files in immunity: sex-based differences predispose immune responses. Nat. Rev. Immunol. 2008, 8, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Shen, M.; Xie, F.; Miao, X.J.; Shou, W.H.; Liu, L.; Yang, P.P.; Bai, Y.N.; Zhang, K.Y.; Yang, L.; et al. An X chromosome-wide association analysis identifies variants in GPR174 as a risk factor for Graves’ disease. J. Med. Genetics 2013, 50, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Latif, R.; Tomer, Y.; Davies, T.F. Thyroid epigenetics: X chromosome inactivation in patients with autoimmune thyroid disease. Ann. N. Y. Acad. Sci. 2007, 1110, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Chabchoub, G.; Uz, E.; Maalej, A.; Mustafa, C.A.; Rebai, A.; Mnif, M.; Bahloul, Z.; Farid, N.R.; Ozcelik, T.; Ayadi, H. Analysis of skewed X-chromosome inactivation in females with rheumatoid arthritis and autoimmune thyroid diseases. Arthritis Res. Therapy 2009, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Ishido, N.; Inoue, N.; Watanabe, M.; Hidaka, Y.; Iwatani, Y. The relationship between skewed X chromosome inactivation and the prognosis of Graves’ and Hashimoto’s diseases. Thyroid Off. J. Am. Thyroid Assoc. 2015, 25, 256–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, Y.; Tozaki, T.; Tobe, T.; Ban, Y.; Jacobson, E.M.; Concepcion, E.S.; Tomer, Y. The regulatory T cell gene FOXP3 and genetic susceptibility to thyroid autoimmunity: an association analysis in Caucasian and Japanese cohorts. J. Autoimmun. 2007, 28, 201–207. [Google Scholar] [CrossRef]
- Inoue, N.; Watanabe, M.; Morita, M.; Tomizawa, R.; Akamizu, T.; Tatsumi, K.; Hidaka, Y.; Iwatani, Y. Association of functional polymorphisms related to the transcriptional level of FOXP3 with prognosis of autoimmune thyroid diseases. Clin. Exp. Immunol. 2010, 162, 402–406. [Google Scholar] [CrossRef]
- Xiao, W.; Liu, Z.; Lin, J.; Li, J.; Wu, K.; Ma, Y.; Xiong, C.; Gong, Y.; Liu, Z. Association of Toll-like receptor 7 and 8 gene polymorphisms with Graves’ disease in Chinese Cantonese population. Tissue Antigens 2015, 85, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Song, R.H.; Qin, Q.; Yan, N.; Muhali, F.S.; Meng, S.; He, S.T.; Zhang, J.A. Variants in IRAK1-MECP2 region confer susceptibility to autoimmune thyroid diseases. Mol. Cell. Endocrinol. 2015, 399, 244–249. [Google Scholar] [CrossRef]
- Shin, H.R.; Cho, W.K.; Baek, I.C.; Lee, N.Y.; Lee, Y.J.; Kim, S.K.; Ahn, M.B.; Suh, B.K.; Kim, T.G. Polymorphisms of IRAK1 gene on X chromosome is associated with Hashimoto’s thyroiditis in Korean Children. Endocrinology 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.X.; Xue, L.Q.; Liu, W.; Gu, Z.H.; Pan, C.M.; Yang, S.Y.; Zhan, M.; Wang, H.N.; Liang, J.; Gao, G.Q.; et al. Robust evidence for five new Graves’ disease risk loci from a staged genome-wide association analysis. Hum. Mol. Genetics 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gastaldi, R.; Poggi, E.; Mussa, A.; Weber, G.; Vigone, M.C.; Salerno, M.; Delvecchio, M.; Peroni, E.; Pistorio, A.; Corrias, A. Graves disease in children: thyroid-stimulating hormone receptor antibodies as remission markers. J. Pediatr. 2014, 164, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Watanabe, M.; Morita, M.; Tatusmi, K.; Hidaka, Y.; Akamizu, T.; Iwatani, Y. Association of functional polymorphisms in promoter regions of IL5, IL6 and IL13 genes with development and prognosis of autoimmune thyroid diseases. Clin. Exp. Immunol. 2011, 163, 318–323. [Google Scholar] [CrossRef]
- Graffelman, J.; Weir, B.S. Testing for Hardy-Weinberg equilibrium at biallelic genetic markers on the X chromosome. Heredity 2016, 116, 558–568. [Google Scholar] [CrossRef] [Green Version]
- Fisher, D.A.; Oddie, T.H.; Johnson, D.E.; Nelson, J.C. The diagnosis of Hashimoto’s thyroiditis. J. Clin. Endocrinol. Metab. 1975, 40, 795–801. [Google Scholar] [CrossRef]
- Werner, S.C. Modification of the classification of the eye changes of Graves’ disease: recommendations of the Ad Hoc Committee of the American Thyroid Association. J. Clin. Endocrinol. Metab. 1977, 44, 203–204. [Google Scholar] [CrossRef]
- Frecker, M.; Stenszky, V.; Balazs, C.; Kozma, L.; Kraszits, E.; Farid, N.R. Genetic factors in Graves’ ophthalmopathy. Clin. Endocrinol. 1986, 25, 479–485. [Google Scholar] [CrossRef]
- Shin, D.H.; Baek, I.C.; Kim, H.J.; Choi, E.J.; Ahn, M.; Jung, M.H.; Suh, B.K.; Cho, W.K.; Kim, T.G. HLA alleles, especially amino-acid signatures of HLA-DPB1, might contribute to the molecular pathogenesis of early-onset autoimmune thyroid disease. PLoS ONE 2019, 14, e0216941. [Google Scholar] [CrossRef]
- Schirmer, M.; Ijaz, U.Z.; D’Amore, R.; Hall, N.; Sloan, W.T.; Quince, C. Insight into biases and sequencing errors for amplicon sequencing with the Illumina MiSeq platform. Nucleic Acids Res. 2015, 43, e37. [Google Scholar] [CrossRef]
- Jäger, A.C.; Alvarez, M.L.; Davis, C.P.; Guzmán, E.; Han, Y.; Way, L.; Walichiewicz, P.; Silva, D.; Pham, N.; Caves, G.; et al. Developmental validation of the MiSeq FGx Forensic Genomics System for Targeted Next Generation Sequencing in Forensic DNA Casework and Database Laboratories. Forensic. Sci. Int. Genet. 2017, 28, 52–70. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravi, R.K.; Walton, K.; Khosroheidari, M. MiSeq: A Next Generation Sequencing Platform for Genomic Analysis. Methods Mol. Biol. (Clifton, N.J.) 2018, 1706, 223–232. [Google Scholar] [CrossRef]
- Sikkema-Raddatz, B.; Johansson, L.F.; de Boer, E.N.; Almomani, R.; Boven, L.G.; van den Berg, M.P.; van Spaendonck-Zwarts, K.Y.; van Tintelen, J.P.; Sijmons, R.H.; Jongbloed, J.D.; et al. Targeted next-generation sequencing can replace Sanger sequencing in clinical diagnostics. Hum. Mutat. 2013, 34, 1035–1042. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.T.; Thorvaldsdottir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Blankenberg, D.; Von Kuster, G.; Bouvier, E.; Baker, D.; Afgan, E.; Stoler, N.; Taylor, J.; Nekrutenko, A. Dissemination of scientific software with Galaxy ToolShed. Genome Biol. 2014, 15, 403. [Google Scholar] [CrossRef] [Green Version]
- Rebolledo-Jaramillo, B.; Su, M.S.; Stoler, N.; McElhoe, J.A.; Dickins, B.; Blankenberg, D.; Korneliussen, T.S.; Chiaromonte, F.; Nielsen, R.; Holland, M.M.; et al. Maternal age effect and severe germ-line bottleneck in the inheritance of human mitochondrial DNA. Proc. Natl. Acad. Sci. USA 2014, 111, 15474–15479. [Google Scholar] [CrossRef] [Green Version]
- Jun, J.K.; Kim, S.M. Association study of fibroblast growth factor 2 and fibroblast growth factor receptors gene polymorphism in korean ossification of the posterior longitudinal ligament patients. J. Korean Neurosurg. Soc. 2012, 52, 7–13. [Google Scholar] [CrossRef]
- Waldenlind, K.; Saevarsdottir, S.; Bengtsson, C.; Askling, J. Risk of Thyroxine-Treated Autoimmune Thyroid Disease Associated With Disease Onset in Patients With Rheumatoid Arthritis. JAMA Netw. Open 2018, 1, e183567. [Google Scholar] [CrossRef]
- Kirchner, J.; Bevan, M.J. ITM2A is induced during thymocyte selection and T cell activation and causes downregulation of CD8 when overexpressed in CD4(+)CD8(+) double positive thymocytes. J. Exp. Med. 1999, 190, 217–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeters, M.C.; van Westen, G.J.; Li, Q.; AP, I.J. Importance of the extracellular loops in G protein-coupled receptors for ligand recognition and receptor activation. Trends Pharmacol. Sci. 2011, 32, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.ncbi.nlm.nih.gov/snp/rs3827440 (accessed on 26 June 2020).
- Szymanski, K.; Miskiewicz, P.; Pirko, K.; Jurecka-Lubieniecka, B.; Kula, D.; Hasse-Lazar, K.; Krajewski, P.; Bednarczuk, T.; Ploski, R. rs3827440, a nonsynonymous single nucleotide polymorphism within GPR174 gene in X chromosome, is associated with Graves’ disease in Polish Caucasian population. Tissue Antigens 2014, 83, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.P.; Yuan, F.F.; Zhang, L.L.; Ma, Y.R.; Zhang, M.M.; Liu, W.; Sun, F.; Wu, J.; Lu, M.; Xue, L.Q.; et al. ITM2A Expands Evidence for Genetic and Environmental Interaction in Graves Disease Pathogenesis. J. Clin. Endocrinol. Metab. 2017, 102, 652–660. [Google Scholar] [CrossRef]
- Barnes, M.J.; Cyster, J.G. Lysophosphatidylserine suppression of T-cell activation via GPR174 requires Gαs proteins. Immunol. Cell Biol. 2018, 96, 439–445. [Google Scholar] [CrossRef]
- Napier, C.; Mitchell, A.L.; Gan, E.; Wilson, I.; Pearce, S.H. Role of the X-linked gene GPR174 in autoimmune Addison’s disease. J. Clin. Endocrinol. Metab. 2015, 100, E187–190. [Google Scholar] [CrossRef] [Green Version]
- Puig, X.; Ginebra, J.; Graffelman, J. A Bayesian test for Hardy-Weinberg equilibrium of biallelic X-chromosomal markers. Heredity 2017, 119, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Wellek, S.; Ziegler, A. Testing for goodness rather than lack of fit of an X-chromosomal SNP to the Hardy-Weinberg model. PLoS ONE 2019, 14, e0212344. [Google Scholar] [CrossRef]


{kind=link}
| AITD (n = 115) | Controls (n = 204) | ||
|---|---|---|---|
| GD (n = 74) | HD (n = 41) | ||
| Females (%) | 52 (70.3%) | 38 (92.7%) | 100 (49.0%) |
| Age at enrollment (years) | 14.7 ± 3.6 | 13.4 ± 3.2 | 22.7 ± 3.8 |
| Goiter (%) | 62 (83.8%) | 25 (61.0%) | |
| T3 at diagnosis, 0.78–1.82 ng/mL | 3.79 ± 2.05 | 1.22 ± 0.55 | n.d. |
| Free T4 at diagnosis, 0.85–1.86 ng/dL | 3.17 ± 1.32 | 1.07 ± 0.74 | n.d. |
| TSH at diagnosis, 0.17–4.05 mIU/L | 0.06 ± 0.20 | 30.57 ± 48.36 | n.d. |
| TSHR Ab positive at diagnosis | 72 (97.3%) | n.d. | |
| Tg Ab positive at diagnosis | 32 (78%) | n.d. | |
| TPO Ab positive at diagnosis | 36 (87.8%) | n.d. | |
| Clinically evident TAO (NOSPECS class II or higher), n (%) | 40 (54.1%) | ||
| Remission | 23 (31.1%) | ||
| Gene | RS Number (SNP) | SNP Position (hg19) | Direction | Sequence (5′-3′) | Span a | Specific | |||
|---|---|---|---|---|---|---|---|---|---|
| Tm | Insert Length | Amplicon Size (bp) | |||||||
| PCR1 primer | GPR174 | rs3810712 | 78,426,471 | Forward | ACACTCTTTCCCTACACGACGCTCTTCCGATCT TTG GAA GGA ACA GCA GTT GAT TG | 34 | 63 | 65 | 95 |
| Reverse | GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCT ACG TGT AAT TAG CAG GCA TGA TTC TCT CTA | 65 | |||||||
| GPR174 | rs3810711 | 78,426,488 | Forward | ACACTCTTTCCCTACACGACGCTCTTCCGATCT TTG GAA GGA ACA GCA GTT GAT TGT GAA TTT A | 34 | 65 | 95 | ||
| Reverse | GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCT ACG TGT AAT TAG CAG GCA TGA TTC TCT CTA | 65 | |||||||
| GPR174 | rs3827440 | 78,426,988 | Forward | ACACTCTTTCCCTACACGACGCTCTTCCGATCT CCT GTG TAC TCT TTC CAC TCC TCA GAA | 39 | 61 | 97 | ||
| Reverse | GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCT GCC AGG TTG ACA TTC CTG GTA GGA AGA TCC A | 66 | |||||||
| ITM2A-GPR174 | rs5912838 | 78,497,118 | Forward | ACACTCTTTCCCTACACGACGCTCTTCCGATCT TTC CAC TTC ATG TTA GAT AAA TTT GGA TGT CA | 25 | 66 | 86 | ||
| Reverse | GTGACTGGAGTTCAGACGTGTGCTCTTCCGATCT ACT ATG ATC ACA TTT CTC TGG ATA CTT GA | 64 | |||||||
| PCR2 Illumina adapters (sequencing) | MiSeq_F | AATGATACGGCGACCACCGAGATCTACAC | - | 59 | - | - | |||
| MiSeq_R | CAAGCAGAAGACGGCATACGAGAT | ||||||||
| Controls | AITD (GD and HD) * | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Analysis Type | n = 204 (%) | n = 115 (%) | χ2 | p-Value | Pc | OR | 95CI (Low) | 95CI (High) | ||||
| F100, | M104 | F90, | M25 | |||||||||
| GPR174 rs3810712 C>T | F genotype | CC | 28 | (28.0) | 39 | (43.3) | 4.879 | 0.027 | 0.082 | 2.0 | 1.1 | 3.6 |
| CT | 48 | (48.0) | 41 | (45.6) | 0.114 | 0.736 | 2.208 | NA | NA | NA | ||
| TT | 24 | (24.0) | 10 | (11.1) | 5.356 | 0.021 | 0.062 | 0.4 | 0.2 | 0.9 | ||
| Comb Genotype | CC/C | 66 | (32.4) | 55 | (47.8) | 7.479 | 0.006 | 0.019 | 1.9 | 1.2 | 3.1 | |
| CT | 48 | (23.5) | 41 | (35.7) | 5.373 | 0.020 | 0.061 | 1.8 | 1.1 | 3.0 | ||
| TT/T | 87 | (42.6) | 19 | (16.5) | 22.623 | 0.000 | 0.000 | 0.3 | 0.2 | 0.5 | ||
| F Allele | C | 104 | (52.0) | 119 | (66.1) | 7.780 | 0.005 | 0.011 | 1.8 | 1.2 | 2.7 | |
| T | 96 | (48.0) | 61 | (33.9) | 7.780 | 0.005 | 0.011 | 0.6 | 0.4 | 0.8 | ||
| M Allele | C | 41 | (39.4) | 16 | (64.0) | 4.936 | 0.026 | 0.053 | 2.7 | 1.1 | 6.8 | |
| T | 66 | (63.5) | 9 | (36.0) | 6.245 | 0.012 | 0.025 | 0.3 | 0.1 | 0.8 | ||
| Comb Allele | C | 145 | (47.7) | 135 | (65.9) | 16.308 | 0.000 | 0.000 | 2.1 | 1.5 | 3.0 | |
| T | 162 | (53.3) | 70 | (34.1) | 18.089 | 0.000 | 0.000 | 0.5 | 0.3 | 0.7 | ||
| GPR174 rs3810711 T>C | F genotype | TT | 28 | (28.0) | 39 | (43.3) | 4.879 | 0.027 | 0.082 | 2.0 | 1.1 | 3.6 |
| TC | 48 | (48.0) | 41 | (45.6) | 0.114 | 0.736 | 2.208 | NA | NA | NA | ||
| CC | 24 | (24.0) | 10 | (11.1) | 5.356 | 0.021 | 0.062 | 0.4 | 0.2 | 0.9 | ||
| Comb Genotype | TT/T | 66 | (32.4) | 55 | (47.8) | 7.479 | 0.006 | 0.019 | 1.9 | 1.2 | 3.1 | |
| TC | 48 | (23.5) | 41 | (35.7) | 5.373 | 0.020 | 0.061 | 1.8 | 1.1 | 3.0 | ||
| CC/C | 87 | (42.6) | 19 | (16.5) | 22.623 | 0.000 | 0.000 | 0.3 | 0.2 | 0.5 | ||
| F Allele | T | 104 | (52.0) | 119 | (66.1) | 7.780 | 0.005 | 0.011 | 1.8 | 1.2 | 2.7 | |
| C | 96 | (48.0) | 61 | (33.9) | 7.780 | 0.005 | 0.011 | 0.6 | 0.4 | 0.8 | ||
| M Allele | T | 41 | (39.4) | 16 | (64.0) | 4.936 | 0.026 | 0.053 | 2.7 | 1.1 | 6.8 | |
| C | 66 | (63.5) | 9 | (36.0) | 6.245 | 0.012 | 0.025 | 0.3 | 0.1 | 0.8 | ||
| Comb Allele | T | 145 | (47.7) | 135 | (65.9) | 16.308 | 0.000 | 0.000 | 2.1 | 1.5 | 3.0 | |
| C | 162 | (53.3) | 70 | (34.1) | 18.089 | 0.000 | 0.000 | 0.5 | 0.3 | 0.7 | ||
| GPR174 rs3827440 T>C | F genotype | TT | 28 | (28.0) | 39 | (43.3) | 4.879 | 0.027 | 0.082 | 2.0 | 1.1 | 3.6 |
| TC | 48 | (48.0) | 41 | (45.6) | 0.114 | 0.736 | 2.208 | NA | NA | NA | ||
| CC | 24 | (24.0) | 10 | (11.1) | 5.356 | 0.021 | 0.062 | 0.4 | 0.2 | 0.9 | ||
| Comb Genotype | TT/T | 66 | (32.4) | 55 | (47.8) | 7.479 | 0.006 | 0.019 | 1.9 | 1.2 | 3.1 | |
| TC | 48 | (23.5) | 41 | (35.7) | 5.373 | 0.020 | 0.061 | 1.8 | 1.1 | 3.0 | ||
| CC/C | 87 | (42.6) | 19 | (16.5) | 22.623 | 0.000 | 0.000 | 0.3 | 0.2 | 0.5 | ||
| F Allele | T | 104 | (52.0) | 119 | (66.1) | 7.780 | 0.005 | 0.011 | 1.8 | 1.2 | 2.7 | |
| C | 96 | (48.0) | 61 | (33.9) | 7.780 | 0.005 | 0.011 | 0.6 | 0.4 | 0.8 | ||
| M Allele | T | 41 | (39.4) | 16 | (64.0) | 4.936 | 0.026 | 0.053 | 2.7 | 1.1 | 6.8 | |
| C | 66 | (63.5) | 9 | (36.0) | 6.245 | 0.012 | 0.025 | 0.3 | 0.1 | 0.8 | ||
| Comb Allele | T | 145 | (47.7) | 135 | (65.9) | 16.308 | 0.000 | 0.000 | 2.1 | 1.5 | 3.0 | |
| C | 162 | (53.3) | 70 | (34.1) | 18.089 | 0.000 | 0.000 | 0.5 | 0.3 | 0.7 | ||
| ITM2A-GPR174 rs5912838 A>C | F genotype | AA | 29 | (29.0) | 39 | (43.3) | 4.235 | 0.040 | 0.119 | 1.9 | 1.0 | 3.4 |
| AC | 44 | (44.0) | 41 | (45.6) | 0.046 | 0.830 | 2.489 | NA | NA | NA | ||
| CC | 27 | (27.0) | 10 | (11.1) | 7.626 | 0.006 | 0.017 | 0.3 | 0.2 | 0.7 | ||
| Comb Genotype | AA/A | 68 | (33.3) | 55 | (47.8) | 6.520 | 0.011 | 0.032 | 1.8 | 1.1 | 2.9 | |
| AC | 44 | (21.6) | 41 | (35.7) | 7.463 | 0.006 | 0.019 | 2.0 | 1.2 | 3.3 | ||
| CC/C | 89 | (43.6) | 19 | (16.5) | 24.129 | 0.000 | 0.000 | 0.3 | 0.1 | 0.4 | ||
| F Allele | A | 102 | (51.0) | 119 | (66.1) | 8.890 | 0.003 | 0.006 | 1.9 | 1.2 | 2.8 | |
| C | 98 | (49.0) | 61 | (33.9) | 8.890 | 0.003 | 0.006 | 0.5 | 0.4 | 0.8 | ||
| M Allele | A | 42 | (40.4) | 16 | (64.0) | 4.542 | 0.033 | 0.066 | 2.6 | 1.1 | 6.5 | |
| C | 65 | (62.5) | 9 | (36.0) | 5.787 | 0.016 | 0.032 | 0.34 | 0.1 | 0.8 | ||
| Comb Allele | A | 144 | (47.4) | 135 | (65.9) | 16.891 | 0.000 | 0.000 | 2.1 | 1.5 | 3.1 | |
| C | 163 | (53.6) | 70 | (34.1) | 18.703 | 0.000 | 0.000 | 0.4 | 0.3 | 0.6 | ||
| Controls | GD | GD_TAO | GD_w/o TAO | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotype Analysis | Total n = 204 | Total n = 74 | χ2 | p-Value | Pc | OR | 95CI (Low) | 95CI (High) | Total n = 40 | χ2 | p-Value | Pc | OR | 95CI (Low) | 95CI (High) | Total n = 34 | χ2 | p-Value | Pc | OR | 95CI (Low) | 95CI (High) | ||||||
| F100 M104 (%) | F52 M22 (%) | F27 M13 (%) | F25 M9 (%) | |||||||||||||||||||||||||
| GPR174 rs3810712 C>T | F | CC | 28 | (28.0) | 26 | (50.0) | 7.229 | 0.007 | 0.022 | 2.6 | 1.3 | 5.2 | 12 | (44.4) | 2.665 | 0.103 | 0.308 | NA | NA | NA | 14 | (56.0) | 7.028 | 0.008 | 0.024 | 3.3 | 1.3 | 8.1 |
| CT | 48 | (48.0) | 24 | (46.2) | 0.047 | 0.829 | 2.486 | NA | NA | NA | 14 | (51.9) | 0.126 | 0.722 | 2.167 | NA | NA | NA | 10 | (40.0) | 0.515 | 0.473 | 1.419 | NA | NA | NA | ||
| TT | 24 | (24.0) | 2 | (3.8) | 9.800 | 0.002 | 0.005 | 0.1 | 0.0 | 0.6 | 1 | (3.7) | 5.539 | 0.019 | 0.056 | 0.1 | 0.0 | 0.9 | 1 | (4.0) | 5.000 | 0.025 | 0.076 | 0.1 | 0.0 | 1.0 | ||
| Comb | CC/C | 66 | (32.4) | 39 | (52.7) | 9.567 | 0.002 | 0.006 | 2.3 | 1.4 | 4.0 | 19 | (47.5) | 3.380 | 0.066 | 0.198 | NA | NA | NA | 20 | (58.8) | 8.849 | 0.003 | 0.009 | 3.0 | 1.4 | 6.3 | |
| TC | 48 | (23.5) | 24 | (32.4) | 0.047 | 0.829 | 2.486 | NA | NA | NA | 14 | (35.0) | 0.126 | 0.722 | 2.167 | NA | NA | NA | 10 | (29.4) | 0.515 | 0.473 | 1.419 | NA | NA | NA | ||
| TT/T | 87 | (42.6) | 11 | (14.9) | 18.841 | 0.000 | 0.000 | 0.2 | 0.1 | 0.5 | 7 | (17.5) | 8.930 | 0.003 | 0.008 | 0.3 | 0.1 | 0.7 | 4 | (11.8) | 11.769 | 0.001 | 0.002 | 0.2 | 0.1 | 0.5 | ||
| GPR174 rs3810711 T>C | F | TT | 28 | (28.0) | 26 | (50.0) | 7.229 | 0.007 | 0.022 | 2.6 | 1.3 | 5.2 | 12 | (44.4) | 2.665 | 0.103 | 0.308 | NA | NA | NA | 14 | (56.0) | 7.028 | 0.008 | 0.024 | 3.3 | 1.3 | 8.1 |
| TC | 48 | (48.0) | 24 | (46.2) | 0.047 | 0.829 | 2.486 | NA | NA | NA | 14 | (51.9) | 0.126 | 0.722 | 2.167 | NA | NA | NA | 10 | (40.0) | 0.515 | 0.473 | 1.419 | NA | NA | NA | ||
| CC | 24 | (24.0) | 2 | (3.8) | 9.800 | 0.002 | 0.005 | 0.1 | 0.0 | 0.6 | 1 | (3.7) | 5.539 | 0.019 | 0.056 | 0.1 | 0.0 | 0.9 | 1 | (4.0) | 5.000 | 0.025 | 0.076 | 0.1 | 0.0 | 1.0 | ||
| Comb | TT/T | 66 | (32.4) | 39 | (52.7) | 9.567 | 0.002 | 0.006 | 2.3 | 1.4 | 4.0 | 19 | (47.5) | 3.380 | 0.066 | 0.198 | NA | NA | NA | 20 | (58.8) | 8.849 | 0.003 | 0.009 | 3.0 | 1.4 | 6.3 | |
| TC | 48 | (23.5) | 24 | (32.4) | 0.047 | 0.829 | 2.486 | NA | NA | NA | 14 | (35.0) | 0.126 | 0.722 | 2.167 | NA | NA | NA | 10 | (29.4) | 0.515 | 0.473 | 1.419 | NA | NA | NA | ||
| CC/C | 87 | (42.6) | 11 | (14.9) | 18.841 | 0.000 | 0.000 | 0.2 | 0.1 | 0.5 | 7 | (17.5) | 8.930 | 0.003 | 0.008 | 0.3 | 0.1 | 0.7 | 4 | (11.8) | 11.769 | 0.001 | 0.002 | 0.2 | 0.1 | 0.5 | ||
| GPR174 rs3827440 T>C | F | TT | 28 | (28.0) | 26 | (50.0) | 7.229 | 0.007 | 0.022 | 2.6 | 1.3 | 5.2 | 12 | (44.4) | 2.665 | 0.103 | 0.308 | NA | NA | NA | 14 | (56.0) | 7.028 | 0.008 | 0.024 | 3.3 | 1.3 | 8.1 |
| TC | 48 | (48.0) | 24 | (46.2) | 0.047 | 0.829 | 2.486 | NA | NA | NA | 14 | (51.9) | 0.126 | 0.722 | 2.167 | NA | NA | NA | 10 | (40.0) | 0.515 | 0.473 | 1.419 | NA | NA | NA | ||
| CC | 24 | (24.0) | 2 | (3.8) | 9.800 | 0.002 | 0.005 | 0.1 | 0.0 | 0.6 | 1 | (3.7) | 5.539 | 0.019 | 0.056 | 0.1 | 0.0 | 0.9 | 1 | (4.0) | 5.000 | 0.025 | 0.076 | 0.1 | 0.0 | 1.0 | ||
| Comb | TT/T | 66 | (32.4) | 39 | (52.7) | 9.567 | 0.002 | 0.006 | 2.3 | 1.4 | 4.0 | 19 | (47.5) | 3.380 | 0.066 | 0.198 | NA | NA | NA | 20 | (58.8) | 8.849 | 0.003 | 0.009 | 3.0 | 1.4 | 6.3 | |
| TC | 48 | (23.5) | 24 | (32.4) | 0.047 | 0.829 | 2.486 | NA | NA | NA | 14 | (35.0) | 0.126 | 0.722 | 2.167 | NA | NA | NA | 10 | (29.4) | 0.515 | 0.473 | 1.419 | NA | NA | NA | ||
| CC/C | 87 | (42.6) | 11 | (14.9) | 18.841 | 0.000 | 0.000 | 0.2 | 0.1 | 0.5 | 7 | (17.5) | 8.930 | 0.003 | 0.008 | 0.3 | 0.1 | 0.7 | 4 | (11.8) | 11.769 | 0.001 | 0.002 | 0.2 | 0.1 | 0.5 | ||
| ITM2A-GPR174 rs5912838 A>C | F | AA | 29 | (29.0) | 26 | (50.0) | 6.534 | 0.011 | 0.032 | 2.4 | 1.2 | 4.9 | 12 | (44.4 | 2.320 | 0.128 | 0.383 | NA | NA | NA | 14 | (56.0) | 6.461 | 0.011 | 0.033 | 3.1 | 1.3 | 7.7 |
| AC | 44 | (44.0) | 24 | (46.2) | 0.064 | 0.800 | 2.400 | NA | NA | NA | 14 | (51.9 | 0.528 | 0.467 | 1.402 | NA | NA | NA | 10 | (40.0) | 0.130 | 0.718 | 2.154 | NA | NA | NA | ||
| CC | 27 | (27.0) | 2 | (3.8) | 11.879 | 0.001 | 0.002 | 0.1 | 0.0 | 0.5 | 1 | (3.7) | 6.713 | 0.010 | 0.029 | 0.1 | 0.0 | 0.8 | 1 | (4.0) | 6.087 | 0.014 | 0.041 | 0.1 | 0.0 | 0.9 | ||
| Comb | AA/A | 68 | (33.3) | 39 | (52.7) | 8.605 | 0.003 | 0.010 | 2.2 | 1.3 | 3.8 | 19 | (47.5) | 2.925 | 0.087 | 0.262 | NA | NA | NA | 20 | (58.8) | 8.126 | 0.004 | 0.013 | 2.9 | 1.4 | 6.0 | |
| AC | 44 | (21.6) | 24 | (32.4) | 0.064 | 0.800 | 2.400 | NA | NA | NA | 14 | (35.0) | 0.528 | 0.467 | 1.402 | NA | NA | NA | 10 | (29.4) | 0.130 | 0.718 | 2.154 | NA | NA | NA | ||
| CC/C | 89 | (43.6) | 11 | (14.9) | 19.505 | 0.000 | 0.000 | 0.2 | 0.1 | 0.5 | 7 | (17.5) | 9.566 | 0.002 | 0.006 | 0.3 | 0.1 | 0.6 | 4 | (11.8) | 12.428 | 0.000 | 0.001 | 0.2 | 0.1 | 0.5 | ||
| Controls | GD | GD_TAO | GD_w/o TAO | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total n = 204 | Total n = 74 | χ2 | p-Value | Pc | OR | 95CI (Low) | 95CI (High) | Total n = 40 | χ2 | p-Value | Pc | OR | 95CI (Low) | 95CI (High) | Total n = 34 | χ2 | p-Value | Pc | OR | 95CI (Low) | 95CI (High) | |||||||
| Allele | F100 M104 (%) | F52 M22 (%) | F27 M13 (%) | F25 M9 (%) | ||||||||||||||||||||||||
| GPR174 rs3810712 C>T | F | C | 104 | (52.0) | 76 | (73.1) | 12.585 | 0.000 | 0.001 | 2.5 | 1.5 | 4.2 | 38 | (70.4) | 5.821 | 0.016 | 0.032 | 2.2 | 1.1 | 4.2 | 38 | (76.0) | 9.390 | 0.002 | 0.004 | 2.9 | 1.4 | 5.9 |
| T | 96 | (48.0) | 28 | (26.9) | 12.585 | 0.000 | 0.001 | 0.4 | 0.2 | 0.7 | 16 | (29.6) | 5.821 | 0.016 | 0.032 | 0.5 | 0.2 | 0.9 | 12 | (24.0) | 9.390 | 0.002 | 0.004 | 0.3 | 0.2 | 0.7 | ||
| M | C | 41 | (39.4) | 13 | (59.1) | 2.868 | 0.090 | 0.181 | NA | NA | NA | 7 | (53.8) | 0.994 | 0.319 | 0.638 | NA | NA | NA | 6 | (66.7) | 2.531 | 0.112 | 0.223 | NA | NA | NA | |
| T | 66 | (63.5) | 9 | (40.9) | 3.833 | 0.050 | 0.100 | NA | NA | NA | 6 | (46.2) | 1.463 | 0.227 | 0.453 | NA | NA | NA | 3 | (33.3) | 3.162 | 0.075 | 0.151 | NA | NA | NA | ||
| Comb | C | 145 | (47.7) | 89 | (70.6) | 18.894 | 0.000 | 0.000 | 2.6 | 1.7 | 4.1 | 45 | (67.2) | 8.327 | 0.004 | 0.008 | 2.2 | 1.3 | 3.9 | 44 | (74.6) | 14.304 | 0.000 | 0.000 | 3.2 | 1.7 | 6.0 | |
| T | 162 | (53.3) | 37 | (29.4) | 20.508 | 0.000 | 0.000 | 0.4 | 0.2 | 0.6 | 22 | (32.8) | 9.188 | 0.002 | 0.005 | 0.4 | 0.2 | 0.7 | 15 | (25.4) | 15.356 | 0.000 | 0.000 | 0.3 | 0.2 | 0.6 | ||
| GPR174 rs3810711 T>C | F | T | 104 | (52.0) | 76 | (73.1) | 12.585 | 0.000 | 0.001 | 2.5 | 1.5 | 4.2 | 38 | (70.4) | 5.821 | 0.016 | 0.032 | 2.2 | 1.1 | 4.2 | 38 | (76.0) | 9.390 | 0.002 | 0.004 | 2.9 | 1.4 | 5.9 |
| C | 96 | (48.0) | 28 | (26.9) | 12.585 | 0.000 | 0.001 | 0.4 | 0.2 | 0.7 | 16 | (29.6) | 5.821 | 0.016 | 0.032 | 0.5 | 0.2 | 0.9 | 12 | (24.0) | 9.390 | 0.002 | 0.004 | 0.3 | 0.2 | 0.7 | ||
| M | T | 41 | (39.4) | 13 | (59.1) | 2.868 | 0.090 | 0.181 | NA | NA | NA | 7 | (53.8) | 0.994 | 0.319 | 0.638 | NA | NA | NA | 6 | (66.7) | 2.531 | 0.112 | 0.223 | NA | NA | NA | |
| C | 66 | (63.5) | 9 | (40.9) | 3.833 | 0.050 | 0.100 | NA | NA | NA | 6 | (46.2) | 1.463 | 0.227 | 0.453 | NA | NA | NA | 3 | (33.3) | 3.162 | 0.075 | 0.151 | NA | NA | NA | ||
| Comb | T | 145 | (47.7) | 89 | (70.6) | 18.894 | 0.000 | 0.000 | 2.6 | 1.7 | 4.1 | 45 | (67.2) | 8.327 | 0.004 | 0.008 | 2.2 | 1.3 | 3.9 | 44 | (74.6) | 14.304 | 0.000 | 0.000 | 3.2 | 1.7 | 6.0 | |
| C | 162 | (53.3) | 37 | (29.4) | 20.508 | 0.000 | 0.000 | 0.4 | 0.2 | 0.6 | 22 | (32.8) | 9.188 | 0.002 | 0.005 | 0.4 | 0.2 | 0.7 | 15 | (25.4) | 15.356 | 0.000 | 0.000 | 0.3 | 0.2 | 0.6 | ||
| GPR174 rs3827440 T>C | F | T | 104 | (52.0) | 76 | (73.1) | 12.585 | 0.000 | 0.001 | 2.5 | 1.5 | 4.2 | 38 | (70.4) | 5.821 | 0.016 | 0.032 | 2.2 | 1.1 | 4.2 | 38 | (76.0) | 9.390 | 0.002 | 0.004 | 2.9 | 1.4 | 5.9 |
| C | 96 | (48.0) | 28 | (26.9) | 12.585 | 0.000 | 0.001 | 0.4 | 0.2 | 0.7 | 16 | (29.6) | 5.821 | 0.016 | 0.032 | 0.5 | 0.2 | 0.9 | 12 | (24.0) | 9.390 | 0.002 | 0.004 | 0.3 | 0.2 | 0.7 | ||
| M | T | 41 | (39.4) | 13 | (59.1) | 2.868 | 0.090 | 0.181 | NA | NA | NA | 7 | (53.8) | 0.994 | 0.319 | 0.638 | NA | NA | NA | 6 | (66.7) | 2.531 | 0.112 | 0.223 | NA | NA | NA | |
| C | 66 | (63.5) | 9 | (40.9) | 3.833 | 0.050 | 0.100 | NA | NA | NA | 6 | (46.2) | 1.463 | 0.227 | 0.453 | NA | NA | NA | 3 | (33.3) | 3.162 | 0.075 | 0.151 | NA | NA | NA | ||
| Comb | T | 145 | (47.7) | 89 | (70.6) | 18.894 | 0.000 | 0.000 | 2.6 | 1.7 | 4.1 | 45 | (67.2) | 8.327 | 0.004 | 0.008 | 2.2 | 1.3 | 3.9 | 44 | (74.6) | 14.304 | 0.000 | 0.000 | 3.2 | 1.7 | 6.0 | |
| C | 162 | (53.3) | 37 | (29.4) | 20.508 | 0.000 | 0.000 | 0.4 | 0.2 | 0.6 | 22 | (32.8) | 9.188 | 0.002 | 0.005 | 0.4 | 0.2 | 0.7 | 15 | (25.4) | 15.356 | 0.000 | 0.000 | 0.3 | 0.2 | 0.6 | ||
| ITM2A-GPR174 rs5912838 A>C | F | A | 102 | (51.0) | 76 | (73.1) | 13.741 | 0.000 | 0.000 | 2.6 | 1.6 | 4.4 | 38 | (70.4) | 6.449 | 0.011 | 0.022 | 2.3 | 1.2 | 4.4 | 38 | (76.0) | 10.146 | 0.001 | 0.003 | 3.0 | 1.5 | 6.2 |
| C | 98 | (49.0) | 28 | (26.9) | 13.741 | 0.000 | 0.000 | 0.4 | 0.2 | 0.6 | 16 | (29.6) | 6.449 | 0.011 | 0.022 | 0.4 | 0.2 | 0.8 | 12 | (24.0) | 10.146 | 0.001 | 0.003 | 0.3 | 0.2 | 0.7 | ||
| M | A | 42 | (40.4) | 13 | (59.1) | 2.583 | 0.108 | 0.216 | NA | NA | NA | 7 | (53.8) | 0.860 | 0.354 | 0.707 | NA | NA | NA | 6 | (66.7) | 2.342 | 0.126 | 0.252 | NA | NA | NA | |
| C | 65 | (62.5) | 9 | (40.9) | 3.492 | 0.062 | 0.123 | NA | NA | NA | 6 | (46.2) | 1.294 | 0.255 | 0.511 | NA | NA | NA | 3 | (33.3) | 2.940 | 0.086 | 0.173 | NA | NA | NA | ||
| Comb | A | 144 | (47.4) | 89 | (70.6) | 19.425 | 0.000 | 0.000 | 2.7 | 1.7 | 4.2 | 45 | (67.2) | 8.609 | 0.003 | 0.007 | 2.3 | 1.3 | 4.0 | 44 | (74.6) | 14.650 | 0.000 | 0.000 | 3.3 | 1.7 | 6.1 | |
| C | 163 | (53.6) | 37 | (29.4) | 21.062 | 0.000 | 0.000 | 0.4 | 0.2 | 0.6 | 22 | (32.8) | 9.485 | 0.002 | 0.004 | 0.4 | 0.2 | 0.7 | 15 | (25.4) | 15.717 | 0.000 | 0.000 | 0.3 | 0.2 | 0.6 | ||
| Haplotype | Freq. | Case, Control Ratio Counts | Case, Control Frequencies | Chi Square | p-Value | ||
|---|---|---|---|---|---|---|---|
| AITD | Control | AITD | Control | ||||
| + #x2003;- | +#x2003;#x2003;- | ||||||
| CTTA | 0.537 | 135.0:70.0 | 137.0:164.0 | 0.658 | 0.455 | 20.29 | 0.000 |
| TCCC | 0.445 | 70.0:135.0 | 155.0:146.0 | 0.341 | 0.515 | 14.87 | 0.000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, W.K.; Shin, H.-R.; Lee, N.Y.; Kim, S.K.; Ahn, M.B.; Baek, I.-C.; Kim, T.-G.; Suh, B.-K. GPR174 and ITM2A Gene Polymorphisms rs3827440 and rs5912838 on the X chromosome in Korean Children with Autoimmune Thyroid Disease. Genes 2020, 11, 858. https://doi.org/10.3390/genes11080858
Cho WK, Shin H-R, Lee NY, Kim SK, Ahn MB, Baek I-C, Kim T-G, Suh B-K. GPR174 and ITM2A Gene Polymorphisms rs3827440 and rs5912838 on the X chromosome in Korean Children with Autoimmune Thyroid Disease. Genes. 2020; 11(8):858. https://doi.org/10.3390/genes11080858
Chicago/Turabian StyleCho, Won Kyoung, Hye-Ri Shin, Na Yeong Lee, Seul Ki Kim, Moon Bae Ahn, In-Cheol Baek, Tai-Gyu Kim, and Byung-Kyu Suh. 2020. "GPR174 and ITM2A Gene Polymorphisms rs3827440 and rs5912838 on the X chromosome in Korean Children with Autoimmune Thyroid Disease" Genes 11, no. 8: 858. https://doi.org/10.3390/genes11080858