
Expression, Polymorphism, and Potential Functional Sites of the BMPR1A Gene in the Sheep Horn
by
 ,
,
Guoqing Zhang
1,2,†,
Mingxing Chu
2,†,
Hao Yang
2,
Hao Li
2,
Jianxin Shi
2,
Pingjie Feng
2,
Shoufeng Wang
1,* and
Zhangyuan Pan
2,* 1
School of Chemistry and Chemical Engineering, University of Jinan, Jinan 250022, China
2
State Key Laboratory of Animal Biotech Breeding, Institute of Animal Science, Chinese Academy of Agricultural Sciences (CAAS), Beijing 100193, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Genes 2024, 15(3), 376; https://doi.org/10.3390/genes15030376
Submission received: 7 January 2024
/
Revised: 15 March 2024
/
Accepted: 16 March 2024
/
Published: 19 March 2024
(This article belongs to the Special Issue Genetic Mechanism Analysis and Application of Important Economic Traits in Sheep and Goats)
Abstract
:Simple Summary
This study investigated the relationship between the BMPR1A gene and horn type in sheep. The study found that the expression of the BMPR1A gene was significantly higher in the normal horn sheep compared to scurred sheep. This trend was observed in multiple sheep breeds. It was also discovered that there is high expression in the skin of three species: cattle, sheep, and pigs. Importantly, it was found that certain nucleotides in the BMPR1A gene are likely to be crucial in horn size and horn type.
Abstract
Sheep horns are composed of bone and sheaths, and the BMPR1A gene is required for cartilage and osteogenic differentiation. Therefore, the BMPR1A gene may have a function related to the sheep horn, but its relationship with the sheep horn remains unclear. In this study, we first utilized RNA sequencing (RNA-seq) data to investigate the expression of the BMPR1A gene in different tissues and breeds of sheep. Second, whole-genome sequencing (WGS) data were used to explore the functional sites of the BMPR1A gene. Lastly, the allele-specific expression of the BMPR1A gene was explored. Our results indicate that BMPR1A gene expression is significantly higher in the normal horn groups than in the scurred groups. Importantly, this trend is consistent across several sheep breeds. Therefore, this finding suggests that the BMPR1A gene may be related to horn type. A total of 43 Single-Nucleotide Polymorphisms (SNPs) (F-statistics > 0.15) and 10 allele-specific expressions (ASEs) exhibited difference between the large and small horn populations. It is probable that these sites significantly impact the size of sheep horns. Compared to other polled species, we discovered ten amino acid sites that could influence horn presence. By combining RNA-seq and WGS functional loci results, we identified a functional site at position 40574836 on chromosome 25 that is both an SNP and exhibits allele-specific expression. In conclusion, we demonstrated that the BMPR1A gene is associated with horn type and identified some important functional sites which can be used as molecular markers in the breeding of sheep horns.
1. Introduction
Sheep are multipurpose animals that produce meat, milk, hides, and wool. Their primary function is meat production [1]. Horns can be used as weapons and play an important role in defense against predators and in sexual selection through their use in intra-male competition [2,3,4,5,6]. However, horns may be harmful during sheep farming. Generally, sheep have several horn phenotypes, namely, large normal horns, scurred horns, and no horns. Rams usually have larger horns, and ewes usually have smaller horns. Bruising reduces meat quality, and severe udder injuries, which may be the result of attack by a horned individual, reduce milk yield [7]. Although dehorning is legal in some countries, it can be painful and distressing for animals [8,9]. At the same time, horns are expensive to grow and require resources beyond those needed for maintenance or growth, especially during the winter months when they are a major source of energy expenditure for sheep [10,11]. The study of horn formation and the presence or absence of horns is essential not only for the study of natural and sexual selection but also for breeding to facilitate modern sheep production.
Sheep horns are hollow, paired structures with a skeletal core that are covered by an integument [12,13]. Horn development is thought to be primarily controlled by the skin [14]. During the initial stages of horn bud development, the epidermis ceases to produce hair and begins to synthesize the horn [15]. Once initiated, the primordium of the bony core of the horn forms a separate ossification center in the dermal connective tissue beneath the region of horn formation, which later fuses with the skull. Although no studies have shown that the BMPR1A gene is associated with horn formation or horn size, several studies have shown that the BMPR1A gene is associated with skeletal development. BMPR1A is a protein-coding gene that has previously been reported to contribute to bone development, and BMPR1A is required for chondrogenic and osteogenic differentiation [16,17,18]. Studies have shown that the BMPR1A gene regulates chondrogenic lineage differentiation and endochondral bone formation through its effects on chondroblast proliferation and chondrocyte differentiation [19,20]. Based on our previous study, we screened the genes that were differentially expressed in the small-horn and large-horn groups. GO (Gene Ontology) enrichment analysis of these genes revealed that the BMPR1A gene is involved in genes associated with epithelial cell proliferation and cartilage development. We therefore hypothesize that the BMPR1A gene is likely to be involved in the formation of the sheep horn. In this study, we first analyzed the expression characteristics of BMPR1A in different tissues of sheep using RNA-seq data to understand the basic function of BMPR1A. Then, we analyzed the horn-related functional sites in BMPR1A using WGS data. This study will provide some valuable molecular makers in sheep horn breeding.
2. Materials and Methods
2.1. Animal and Sample Collection
All the experimental procedures mentioned in the present study were approved by the Science Research Department (in charge of animal welfare issues) of the Institute of Animal Sciences, Chinese Academy of Agricultural Sciences (IAS-CAAS) (Beijing, China). Ethical approval on animal survival was given by the animal ethics committee of IAS-CAAS (No. IASCAAS-AE-03, 12 December 2016). This study used RNA-sequencing (RNA-seq) data previously collected by our laboratory (PRJNA1003277) [21]. A total of fifteen Tibetan sheep samples were collected in Dangxiong, Tibet, China. These Tibetan sheep are all female and aged between 2 and 4.5 years old. Our Tibetan sheep samples were divided into two groups: one group consisted of 7 sheep with scurred horns (0–12 cm), while the other group consisted of 8 sheep with normal horns (>12 cm) (Supplementary Table S1). The soft-horned tissue was collected from these 15 sheep, placed in deep cryopreservation tubes, and stored in liquid nitrogen.
2.2. RNA Sequencing Data Filtering, Comparison, Assembly, and Processing
RNA sequencing (RNA-seq) data consisted of publicly available and laboratory-collected data. We obtained 2915 high-quality sheep RNA-seq data samples from the National Center for Biotechnology Information (NCBI) database (https://www.ncbi.nlm.nih.gov/, accessed on 15 March 2023) and EBI (https://www.ebi.ac.uk/, accessed on 15 March 2023), which are publicly available. Both the public data and the laboratory-collected data underwent the following processing. Raw data with contaminating adapter molecules, reads containing ploy(N), and low-quality reads were removed using Trim Galore (v.0.6.7). The “--chimSegmentMin 10” and “--outFilterMismatchNmax 3” parameters of the STAR (v.2.7.7a) software were used to align clean reads with the sheep reference genome (the ARS-UI_Ramb_v2.0) [22]. Eventually, high-quality RNA-seq clean datasets were obtained for subsequent analysis, with unique mapping reads > 85% and the number of clean reads > 20,000,000. The expression levels of genes were normalized by calculating the number of transcripts per kilobase fragment (FPKM) per million mapped transcripts and the exonic model by the number of transcripts per kilobase (TPM) per million exons using the prepDE.py script of Stringtie (v.2.1.5) [23]. The raw counts of these genes were subsequently extracted using featureCounts (v.2.0.1) [24].
2.3. Expression of the BMPR1A Gene in Normal Horn and Scurred Groups
To investigate the difference between BMPR1A in normal horn and scurred groups, we produced a boxplot using ggplot2 (v.3.4.0) in R (v.4.2.3). To further understand the distinctions in the expression of BMPR1A exons between the two groups, we utilized the dexseq_prepare_annotation2.py script of Subread_to_DEXSeq (https://github.com/vivekbhr/Subread_to_DEXSeq, accessed on 21 April 2023) to format the genome annotation (GTF) file. Following this, we processed the formatted GTF file and the counts matrix output by featureCounts using load_SubreadOutput.R in Rstudio and constructed the DEXSeqDataSetFromFeatureCounts (dds) object. Finally, we performed an exon difference analysis.
2.4. Analysis of the Expression Profile of the BMPR1A Gene
To explore BMPR1A gene expression across species, we downloaded publicly available RNA-seq data for sheep, pigs, cows, and humans. Additionally, RNA-seq data for 2651 pigs were obtained from the GTEx project, and TPM gene expression values were acquired from http://piggtex.farmgtex.org/ (accessed on 10 September 2023). Furthermore, RNA-seq data for 4359 cows were obtained from the GTEx project, whereas bovine gene expression values in transcripts per million were acquired from https://cgtex.roslin.ed.ac.uk/ (accessed on 1 October 2023) [25]. We downloaded TPM values for genes in the RNA-seq data of 9810 human samples from https://gtexportal.org/home/datasets (accessed on 5 August 2023) [26,27]. Sheep, pig, cow, and human RNA-seq data were pooled to arrange tissue samples, which were subsequently classified into 16 distinct tissues. The average TPM values for the BMPR1A gene were calculated for each species (Supplementary Table S2). To further investigate gender and tissue-specific expression variances in the BMPR1A gene of sheep, we analyzed RNA-seq data. We collected information on sheep breeds from publicly available RNA-seq data and excluded breeds with small sample sizes. The final 8 breeds of sheep obtained were Bashiba, Chinese Merino, Hu, Minxian Black Fur, Spanish Churra, Tan, Texel, and Tibetan. The sheep were divided into three subgroups: polled, scurred, and normal.
2.5. Analysis and Three-Dimensional Structure Prediction of the BMPR1A Protein
To investigate disparities in the BMPR1A protein across various species, we retrieved the FASTa files for this protein from NCBI for 18 different species, including sheep, goat, and cattle. Using MEGA11 software, we produced a protein evolution tree for BMPR1A [28]. The default parameters of the ClustalW method were used for alignment [29,30]. The maximum likelihood method was used to develop a circular evolutionary tree for BMPR1A proteins. To determine whether amino acids are located at key positions in the protein, we used the default parameters of AlphaFold2 to predict the 3D structure of the protein BMPR1A protein [31].
2.6. Whole-Genome Sequencing Analysis
Whole-genome sequencing (WGS) data consisted of publicly available and laboratory-collected data. The publicly available data for 3125 sheep were downloaded from NCBI, including PRJNA304478, PRJNA325682, PRJNA479525, PRJNA624020, PRJNA675420, PRJNA822017, PRJNA30931, PRJNA480684, PRJNA509694, PRJNA779188, and PRJNA783661 [32,33,34,35,36,37,38,39,40,41,42,43]. The sample of 3125 sheep was divided into horned and hornless sheep. F-statistic (Fst) values were calculated using vcftools (v.0.1.16) to discriminate SNP loci that were significantly different in these sheep populations. Small Tail Han sheep were utilized as the laboratory breed, and horn length was quantified for 38 individuals from the root to the outermost end of the horns. The length of the sheep’s right horn was used to determine the length of the sheep’s horn. Trimmomatic (v.0.39) was used to trim the sequencing reads, and FastQC (v. 0.12.1) was used to assess the quality of the raw sequencing data [44]. The qualified reads were then mapped, sorted, and deduped to the sheep reference genome by BWA (v.0.7.17) and Picard (v. 3.1.1) [45,46]. We used the default parameters of the pipeline in the genome analysis toolkit (GATK) (v.4.2.5.0) process to predict the SNP site [47,48]. We used SnpEff (v.4.3) to annotate the mutation information [49]. To determine the relationship between horn length and different genotypes, the dominant model was used. We conducted a deeper analysis of the SNP chain by applying the --maf 0.45 --min-meanDP 5 filters in vcftools (v.0.1.16) and then used LDBlockShow (v.1.40) to identify the chains of SNP loci [50,51].
3. Results
3.1. The Expression of the BMPR1A Gene in Sheep with Different Horn Types
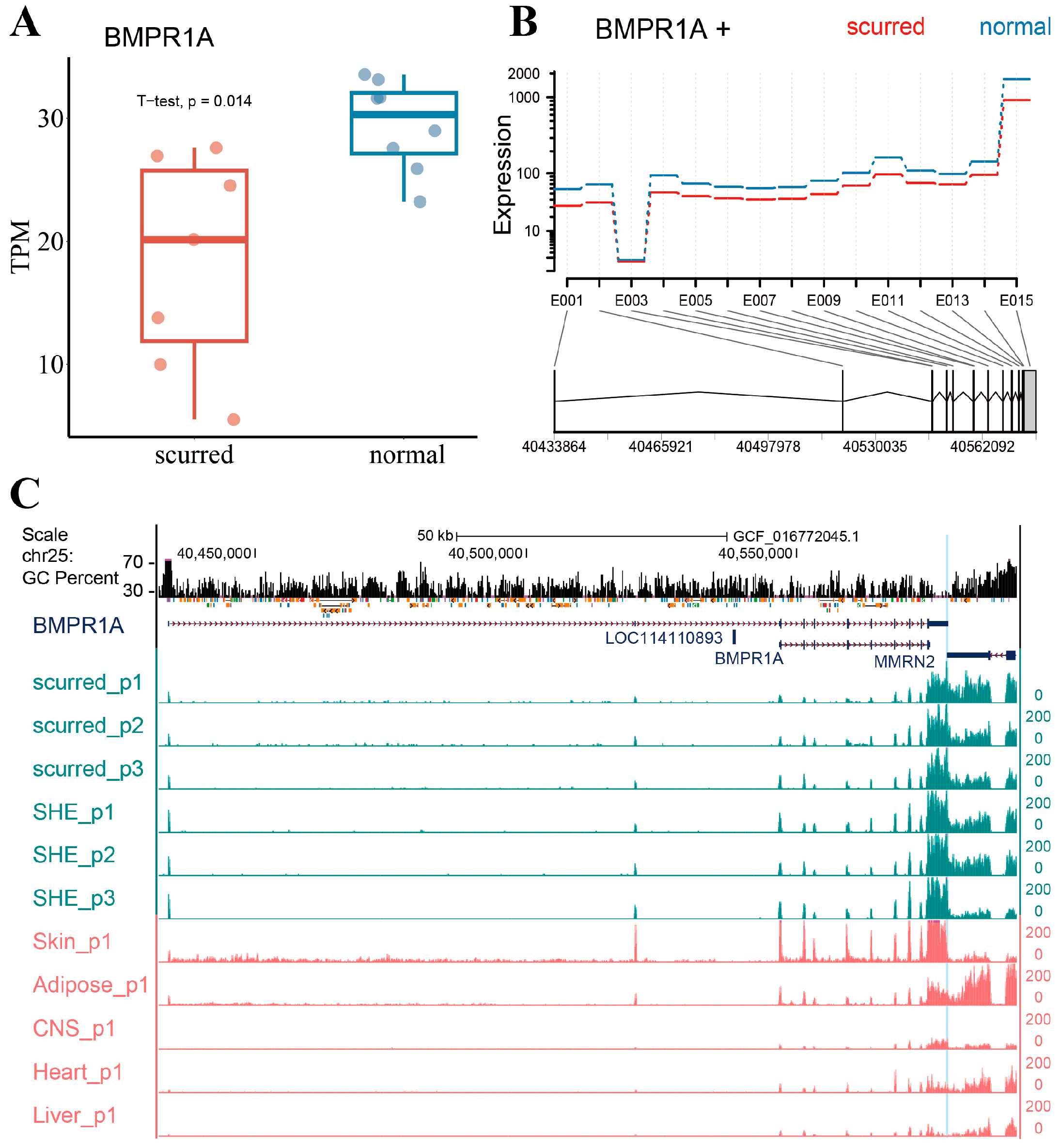
The BMPR1A gene is located on chromosome 25 and has two transcripts: XM_012104920.4 and NM_001280714.1. We use transcript XM_012104920.4 to represent the BMPR1A gene in this paper. As shown in Figure 1A, the expression of the BMPR1A gene was significantly higher in the normal horn group than in the scurred group (p-value = 0.014). The exons of the BMPR1A gene are predominantly located in the 3′ end of the gene. As shown in Figure 1B, all exons of BMPR1A showed higher expression in the normal horn group. We specifically checked the gene expression, GC percent, and repeats of the BMPR1A gene. As shown in Figure 1C, all exons were expressed in both the scurred and normal horn group. However, the expression of exons greatly varied in different tissues. Skin and adipose tissue showed a similar expression pattern to the Exon 13 region of the BMPR1A gene in the sheep horn. Expression of exons except for the Exon 1 region was higher in skin tissues than in other tissues. Additionally, it was observed that Exon 1 and Exon 2 regions were nearly not expressed in the central nervous system (CNS), heart, or liver tissues. Moreover, the expression of various other exon regions was lower in these three tissues compared with skin and horn tissues. Interestingly, we observed that the BMPR1A gene shares a region with the MMRN2 gene. We hypothesize that this region is likely a component of the BMPR1A gene.
3.2. The Tissue-Specific Function of the BMPR1A Gene in Sheep
To better understand whether there exists a difference in BMPR1A gene expression among normal horn and scurred groups in different sheep breeds, we generated Figure 2. Figure 2A displays BMPR1A gene expression across 16 tissues in four different species and shows that the BMPR1A gene exhibited high expression levels in three species: sheep, cattle, and pigs. The BMPR1A gene exhibited the second-highest level of expression in sheep and bovine skin, but its expression in porcine skin had the highest TPM level and was much higher than in other tissues. Additional studies were performed to determine if there were sex differences in the expression of the BMPR1A gene in different sheep tissues. As depicted in Figure 2B, significant differences in BMPR1A gene expression were observed among the skin, brain, colon, muscle, blood–immune, and pituitary tissues (p-value < 0.05). In all six tissues displaying significant differences, BMPR1A expression was higher in ewes than rams. We investigated whether the expression of the BMPR1A gene remained consistent across various sheep breeds, as depicted in Figure 2C. The expression of the BMPR1A gene was lower in all four breeds of hornless sheep (Bashibai, Hu, Spanish Churra, and Texel) compared with Chinese Merino, Minxian Black Fur, and Tan sheep and the three breeds of scurred sheep. The above results suggest high expression of the BMPR1A gene in skin tissues across several species, with a sex-specific difference in expression. Additionally, there was variation in expression observed between polled and scurred sheep.
3.3. The Potential Function of BMPR1A Protein
Figure 3A shows that the evolutionary relationship of proteins was consistent with the evolution of species, and the clustering of horned species in Bovidae and Cervidae suggests that there is similarity in the structure of the BMPR1A protein in these antlered animals in Bovidae and Cervidae. Figure 3C shows the amino acid sites specific to horned animals. Amino acids 40, 43, 44, 60, 91, 95, 189, 190, 268, and 529 were specific to horned animals. These amino acids may play a crucial role in the presence or absence of a sheep horn. Figure 3B shows that amino acid 268 is in the α-helix and has a high predictive accuracy, which may have a significant effect on the presence of a sheep horn.
3.4. Allele-Specific Expression in the BMPR1A Gene
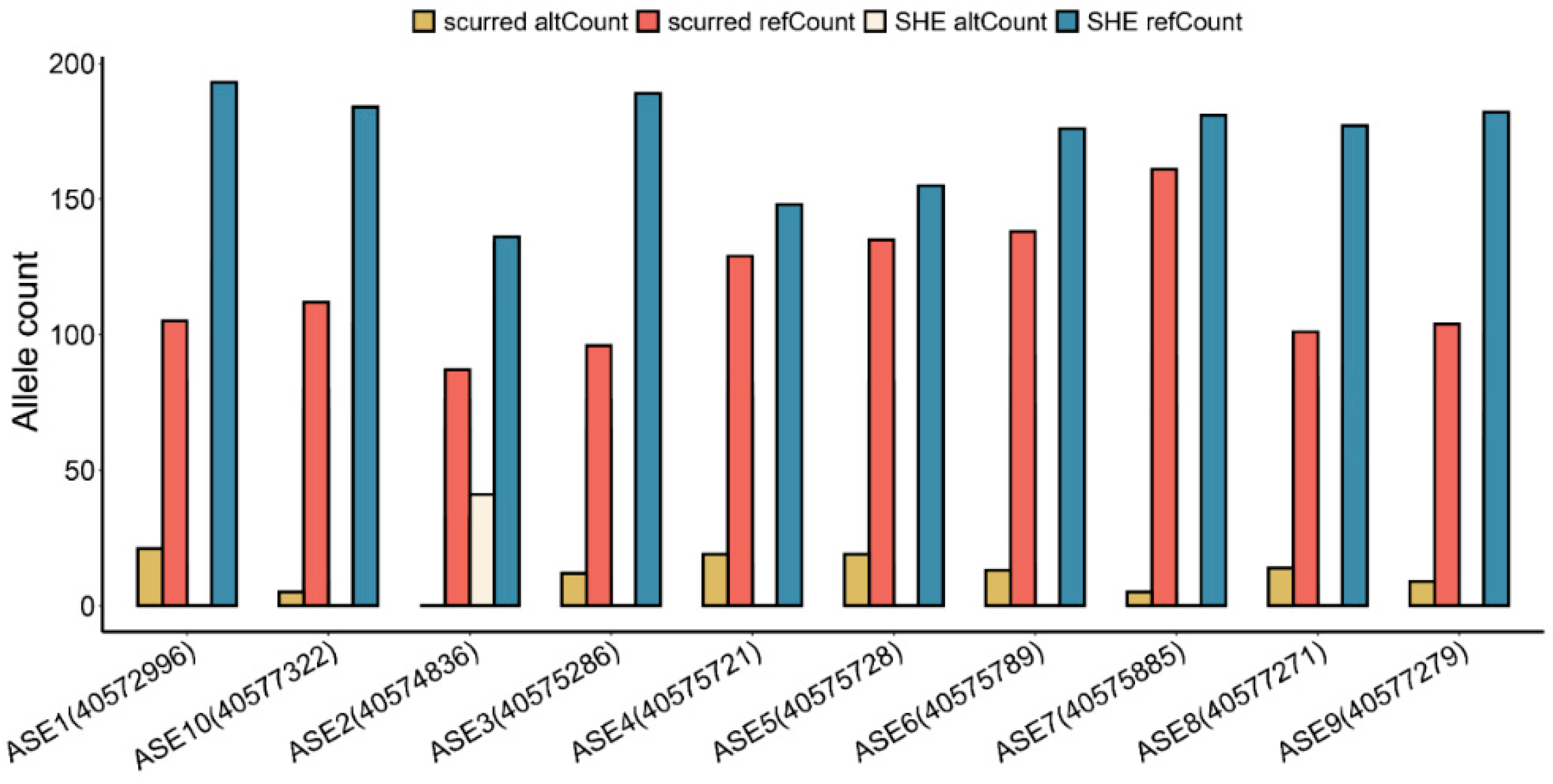
Using RNA-seq data, we found 10 allele-specific expressions (ASEs): ASE1 (chr25:40572996), ASE2 (chr25:40574836), ASE3 (chr25:40575286), ASE4 (chr25:40575721), ASE5 (chr25:40575728), ASE6 (chr25:40575789), ASE7 (chr25:40575885), ASE8 (chr25:40577271), ASE9 (chr25:40577279), and ASE10 (chr25:40577322) (Figure 4). ASE2, ASE3, ASE4, ASE5, ASE6, ASE7, ASE8, ASE9, and ASE10 are located on exon 15, and ASE1 is located on exon 12. The alt (alternative) allele Count of ASE1–ASE10 in each of these ASEs were lower than the reference allele Count. These ASEs, except for ASE2, had an alternative count of 0 in the normal horns group. Therefore, these ASE loci may be closely related to horn type.
3.5. Potential Functional Mutations in the BMPR1A Gene
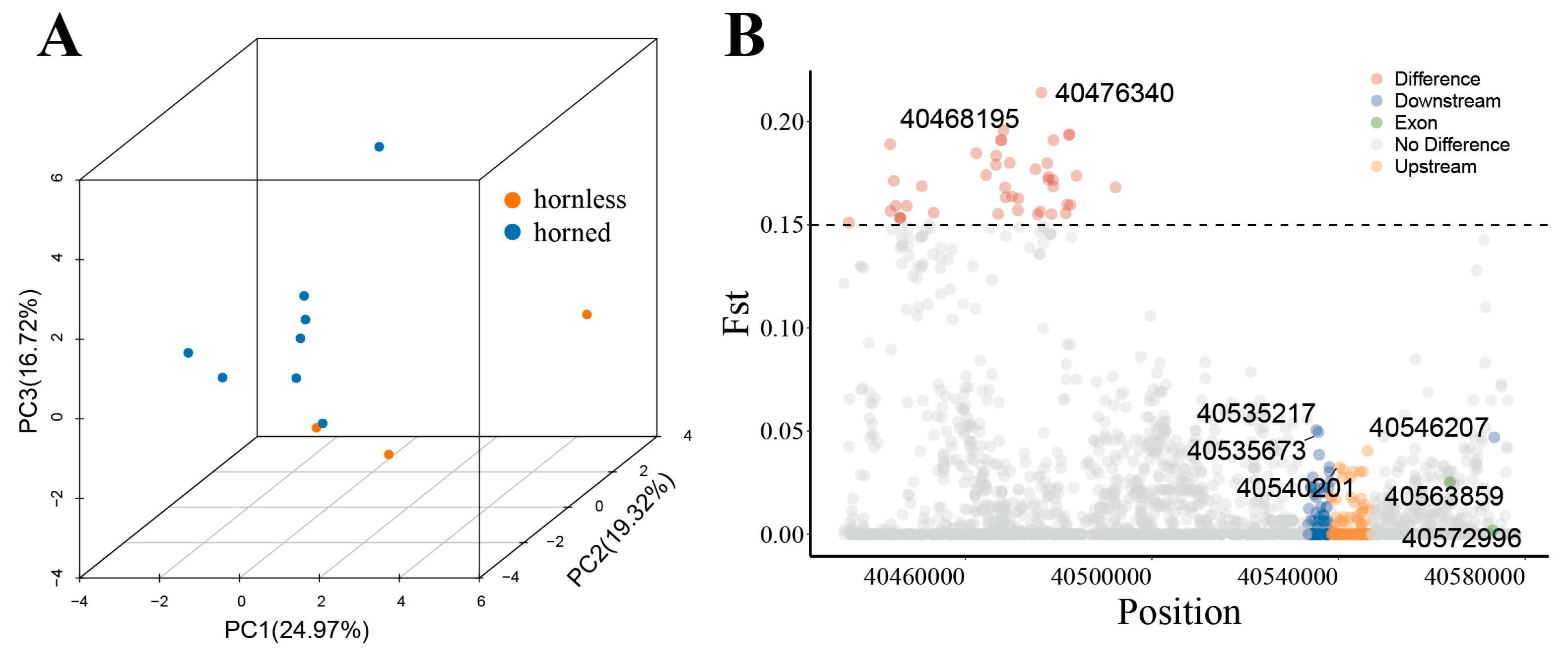
The PCA results showed that these horned breeds of sheep were clearly separated from the polled sheep (Figure 5A). This result indicates that the BMPR1A gene can distinguish between horned and hornless populations to some extent and that loci in the region of the gene play a role in regulating horn. We screened a total of 43 loci with Fst values greater than 0.15 and found 18 loci located in exons of this gene (Supplementary Table S3). As shown in Table 1, there are several potential SNP functional loci of BMPR1A. These functional loci significantly differed in horned and hornless sheep populations and may play a critical role in sheep horn size.
As shown in Figure 6A, there was a significant difference in horn length between sheep with different genotypes at chr25:40468542 according to our results under the dominant model. This SNP probably influences sheep horn size and thus horn length.
We further investigated linkage disequilibrium (LD) among SNPs in the BMPR1A gene. We identified 25 SNPs with significant LD based on our LD analysis. Figure 6B shows three SNP sites (40446552, 40447684, and 40447948) in one LD block, whereas another LD block contained three SNP sites (40512461 and 40512516). We examined the intersection between the ASEs, Fst, and SNP locus and determined that the locus 40574836 showed significant differentiation between the populations of the normal horn and scurred groups (Fst = 0.072885). At the same time, within the normal horn group, ASE2 (40574836) exhibited a significantly higher altcount value than the scurred group, which lacked this expression. These findings indicate a strong likelihood that this genetic locus impacts the size of sheep horns.
4. Discussion
In prior research, RXFP2, FOXL2, ACAN, SFRP2, SFRP4, WNT3, and TNN were identified as important genes that affect horn phenotype [43,52,53,54]. Although the RXFP2 gene has been shown to be associated with large horn size in sheep, a study has demonstrated that the horn phenotype of sheep remains unaffected despite partial disruption of the RXFP2 gene through the use of CRISPR/Cas9 technology [55]. These studies partially explain this phenomenon, indicating that the presence or absence of horns is not solely determined by a single gene or locus.
The function of the BMPR1A gene has been extensively studied. Previous studies of the BMPR1A gene have shown its association with bone quality and bone strength and reproduction [56,57]. It has also been shown that the BMPR1A gene can also promote adipogenesis [58]. Our analysis revealed discrepancies in the BMPR1A gene expression between the normal horn and scurred groups as well as consistency across different sheep breeds. This study did not use qRT-PCR to validate the RNA-seq results. We are aware that this may affect the reliability of our results, and we will add this experiment to future more in-depth studies of the BMPR1A gene on ram’s horn. Using publicly available RNA-seq data, we found that the BMPR1A gene is highly expressed in skin and has similar characteristics in both pigs and cattle. This result has also been demonstrated in studies by others [59]. We also found that the expression of the BMPR1A gene was higher in ewes than in rams, which is consistent with the results of the study by Gwenn-Aël Carré et al. [60]. Our results are consistent with those of previous studies indicating that this gene is expressed at high levels in the ovaries and uterus of ewes and in the testes of rams [29,30]. Using the ASE results, we successfully pinpointed mutation loci in the normal horn and scurred groups that are likely to influence variation in horn size. It is evident from the results that the BMPR1A gene is likely to be located at the amino acid site identified and that sites like ASEs influence BMPR1A expression in the skin, subsequently impacting horn size or shape. In a similar study to ours, it was demonstrated that the reversal of the BMPRIA mutation led to a decrease in bone volume and bone formation in mice [17]. We screened 43 SNPs and identified significant differences between the populations of horned and polled sheep. It is probable that these SNPs influenced the size of the sheep’s horn. Using a dominant model and analyzing WGS data, we discovered an association between the g.40468542T>C variant and sheep horn length. Therefore, we consider that g.40468542T>C could possibly affect horn length. Additionally, we conducted an analysis of SNP position chaining in the BMPR1A gene region and did not identify any significant SNPs chained to other SNPs. Eventually, an important SNP, g.40574836C>T, was identified. It is both an SNP and exhibits allele-specific expression.
Multiple studies have demonstrated that BMPR1A functions via the transforming growth factor β (TGF-β)/BMP pathway and Wnt signaling in the skin [59,61,62]. According to previous studies, it is proposed that the BMPR1A protein activates SMAD proteins by binding to ligands in the TGF-β signaling pathway [63,64,65]. The Smad2/3/4 signaling pathway in osteoblasts regulates osteogenic differentiation of MC3T3-E1 cells through modulation of ClC-3 chloride channels [66]. The TGF-β family is believed to have arisen during multicellular (metazoan) evolution and is highly conserved [67,68,69,70]. In numerous species, TGF-βs facilitate a range of signaling functions during embryonic and adult stages that regulate tissue-specific differentiation, proliferation, as well as movement of cell-specific or tissue-specific motility [71,72,73]. Members of this family comprise activins, BMPs, growth differentiation factors (GDFs), Müllerian-inhibiting substance (MIS), the nodal, and TGF-βs [73]. TGF-β and BMP signaling pathways played important roles in secondary palate formation in a study of palate development [74]. Chloride-conducting ion channels are present in almost all organisms, including members in every mammalian tissue. These channels play a crucial role in regulating cellular excitability, trans-epithelial transport, cell volume regulation, and intracellular organelle acidification [56]. The function of ClC-3 in bone differentiation may be through the Runx2 gene pathway, which mediates bone formation and remodeling [75]. Xiaolin Lu et al. discovered that changes in ClC-3 chloride channels affect Smad2/3 proteins, demonstrating that the Smad2/3/4 signaling pathway inhibits the regulation of osteogenic differentiation in MC3T3-E1 cells [66]. ClC-3 is expressed in intracellular organelles of osteoclasts and promotes osteoclast bone resorption in vitro via organelle acidification [76].
Molecular selection based on the loci identified for horn shape and size offers several advantages for captive sheep breeding. Firstly, it simplifies management as horns can lead to issues during breeding, such as fights between sheep or snagging on fences. Sheep without horns are easier to manage, and the occurrence of these problems can be reduced. Secondly, in intensive farming environments, sheep may injure each other or even the breeder due to their horns. Therefore, sheep without horns reduce the risk of such injuries. Additionally, sheep without horns can use the nutrients that would have been used for horn growth for growth and reproduction, thus improving their survival and reproductive success. This is particularly advantageous in cold regions. For example, Tibetan sheep, which have large horns, may not be as successful in these regions. Finally, animal welfare can be improved by avoiding the painful process of dehorning sheep through selective breeding for hornless sheep.
5. Conclusions
The study concludes that there is a correlation between the BMPR1A gene and the size and type of horn in ewes. Further analysis indicated that ewes had elevated BMPR1A expression levels in skin tissues compared with rams. We identified several potential loci that impact sheep horns, including SNPs and ASEs, along with 10 amino acid sites on the BMPR1A protein that are specifically expressed in horned species. It was found that g.40574836C > T is located on exons, which may have a significant effect on the size of the ewe horns. These loci may significantly impact the size and type of ewe horns and can be used as molecular markers for horn breeding.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes15030376/s1, Table S1: Sample Information; Table S2: Mean TPM of BMPR1A gene in different tissues of Cattle, Human, Pig, and Sheep; Table S3: SNP details identified within the BMPR1A gene region.
Author Contributions
The division of labor among the authors of this article is as follows, methodology, G.Z., H.Y. and H.L.; formal analysis G.Z.; data curation, G.Z. and J.S.; writing—original draft preparation, G.Z., H.Y. and M.C.; Investigation, P.F. and S.W.; writing—review and editing, Z.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key R&D Program Young Scientist (2023YFF1001800), National Key R&D Program of China (2022YFF1000103), National Natural Science Foundation of China (31802031 and 31960659), Agricultural Science and Technology Innovation Program of China (CAAS-ZDRW202106 and ASTIP-IAS13), and China Agriculture Research System of MOF and MARA (CARS-38).
Institutional Review Board Statement
All the experimental procedures mentioned in the present study were approved by the Science Research Department (in charge of animal welfare issues) of the Institute of Animal Sciences, Chinese Academy of Agricultural Sciences (IAS-CAAS) (Beijing, China). Ethical approval on animal survival was given by the animal ethics committee of IAS-CAAS (No. IASCAAS-AE-03, 12 December 2016).
Informed Consent Statement
Not applicable.
Data Availability Statement
Information about the RNA-seq data in this article can be found at https://www.ncbi.nlm.nih.gov/sra/PRJNA1003277 (accessed on 6 January 2024).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zygoyiannis, D. Sheep Production in the World and in Greece. Small Rumin. Res. 2006, 62, 143–147. [Google Scholar] [CrossRef]
- Poissant, J.; Davis, C.S.; Malenfant, R.M.; Hogg, J.T.; Coltman, D.W. QTL Mapping for Sexually Dimorphic Fitness-Related Traits in Wild Bighorn Sheep. Heredity 2012, 108, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.G.; Bateman, R.S.; Morris, P.J. Cerebral Infarction and Meningoencephalitis Following Hot-Iron Disbudding of Goat Kids. N. Z. Vet. J. 2005, 53, 368–370. [Google Scholar] [CrossRef]
- Hempstead, M.N.; Waas, J.R.; Stewart, M.; Cave, V.M.; Turner, A.R.; Sutherland, M.A. The Effectiveness of Clove Oil and Two Different Cautery Disbudding Methods on Preventing Horn Growth in Dairy Goat Kids. PLoS ONE 2018, 13, e0198229. [Google Scholar] [CrossRef] [PubMed]
- Kardos, M.; Luikart, G.; Bunch, R.; Dewey, S.; Edwards, W.; McWilliam, S.; Stephenson, J.; Allendorf, F.W.; Hogg, J.T.; Kijas, J. Whole-Genome Resequencing Uncovers Molecular Signatures of Natural and Sexual Selection in Wild Bighorn Sheep. Mol. Ecol. 2015, 24, 5616–5632. [Google Scholar] [CrossRef]
- Johnston, S.E.; Beraldi, D.; McRae, A.F.; Pemberton, J.M.; Slate, J. Horn Type and Horn Length Genes Map to the Same Chromosomal Region in Soay Sheep. Heredity 2010, 104, 196–205. [Google Scholar] [CrossRef]
- Simon, R.; Drögemüller, C.; Lühken, G. The Complex and Diverse Genetic Architecture of the Absence of Horns (Polledness) in Domestic Ruminants, Including Goats and Sheep. Genes 2022, 13, 832. [Google Scholar] [CrossRef] [PubMed]
- Knierim, U.; Irrgang, N.; Roth, B.A. To Be or Not to Be Horned—Consequences in Cattle. Livest. Sci. 2015, 179, 29–37. [Google Scholar] [CrossRef]
- Still Brooks, K.M.; Hempstead, M.N.; Anderson, J.L.; Parsons, R.L.; Sutherland, M.A.; Plummer, P.J.; Millman, S.T. Characterization of Efficacy and Animal Safety across Four Caprine Disbudding Methodologies. Animals 2021, 11, 430. [Google Scholar] [CrossRef]
- Monteith, K.L.; Long, R.A.; Stephenson, T.R.; Bleich, V.C.; Bowyer, R.T.; Lasharr, T.N. Horn Size and Nutrition in Mountain Sheep: Can Ewe Handle the Truth? J. Wildl. Manag. 2018, 82, 67–84. [Google Scholar] [CrossRef]
- Picard, K.; Festa-Bianchet, M.; Thomas, D. The Cost of Horniness: Heat Loss May Counter Sexual Selection for Large Horns in Temperate Bovids. Écoscience 1996, 3, 280–284. [Google Scholar] [CrossRef]
- Davis, E.B.; Brakora, K.A.; Lee, A.H. Evolution of Ruminant Headgear: A Review. Proc. R. Soc. B Biol. Sci. 2011, 278, 2857–2865. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Wang, N.; Li, Z.; Heller, R.; Liu, R.; Zhao, Y.; Han, J.; Pan, X.; Zheng, Z.; et al. Genetic Basis of Ruminant Headgear and Rapid Antler Regeneration. Science 2019, 364, eaav6335. [Google Scholar] [CrossRef]
- Jheon, A.H.; Mitgutsch, C. Bones and Cartilage: Developmental and Evolutionary Skeletal Biology. Acta Zool. 2008, 89, 277–278. [Google Scholar] [CrossRef]
- Dove, W.F. The Physiology of Horn Growth: A Study of the Morphogenesis, the Interaction of Tissues, and the Evolutionary Processes of a Mendelian Recessive Character by Means of Transplantation of Tissues. J. Exp. Zool. 1935, 69, 347–405. [Google Scholar] [CrossRef]
- Pan, H.; Zhang, H.; Abraham, P.; Komatsu, Y.; Lyons, K.; Kaartinen, V.; Mishina, Y. BmpR1A Is a Major Type 1 BMP Receptor for BMP-Smad Signaling during Skull Development. Dev. Biol. 2017, 429, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Mishina, Y.; Starbuck, M.W.; Gentile, M.A.; Fukuda, T.; Kasparcova, V.; Seedor, J.G.; Hanks, M.C.; Amling, M.; Pinero, G.J.; Harada, S.; et al. Bone Morphogenetic Protein Type IA Receptor Signaling Regulates Postnatal Osteoblast Function and Bone Remodeling. J. Biol. Chem. 2004, 279, 27560–27566. [Google Scholar] [CrossRef] [PubMed]
- Mang, T.; Kleinschmidt-Doerr, K.; Ploeger, F.; Schoenemann, A.; Lindemann, S.; Gigout, A. BMPR1A Is Necessary for Chondrogenesis and Osteogenesis, Whereas BMPR1B Prevents Hypertrophic Differentiation. J. Cell Sci. 2020, 133, jcs246934. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Hinton, R.J.; Mishina, Y.; Liu, Y.; Zhou, X.; Feng, J.Q. Critical Role of Bmpr1a in Mandibular Condyle Growth. Connect. Tissue Res. 2014, 55, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Hinton, R.J.; Feng, J.Q. Chapter Nine—Bmpr1a Signaling in Cartilage Development and Endochondral Bone Formation. In Vitamins & Hormones; Litwack, G., Ed.; Bone Morphogenic Protein; Academic Press: Cambridge, MA, USA, 2015; Volume 99, pp. 273–291. [Google Scholar]
- Li, H.; Du, X.; Li, X.; Feng, P.; Chu, M.; Jin, Y.; Pan, Z. Genetic Diversity, Tissue-Specific Expression, and Functional Analysis of the ATP7A Gene in Sheep. Front. Genet. 2023, 14, 1239979. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinforma. Oxf. Engl. 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinforma. Oxf. Engl. 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Gao, Y.; Canela-Xandri, O.; Wang, S.; Yu, Y.; Cai, W.; Li, B.; Xiang, R.; Chamberlain, A.J.; Pairo-Castineira, E.; et al. A Multi-Tissue Atlas of Regulatory Variants in Cattle. Nat. Genet. 2022, 54, 1438–1447. [Google Scholar] [CrossRef] [PubMed]
- The Gtex Consortium. The GTEx Consortium Atlas of Genetic Regulatory Effects across Human Tissues. Science 2020, 369, 1318–1330. [Google Scholar] [CrossRef]
- The Gtex Consortium; Ardlie, K.G.; Deluca, D.S.; Segrè, A.V.; Sullivan, T.J.; Young, T.R.; Gelfand, E.T.; Trowbridge, C.A.; Maller, J.B.; Tukiainen, T.; et al. The Genotype-Tissue Expression (GTEx) Pilot Analysis: Multitissue Gene Regulation in Humans. Science 2015, 348, 648–660. [Google Scholar]
- MEGA11: Molecular Evolutionary Genetics Analysis Version 11 | Molecular Biology and Evolution | Oxford Academic. Available online: https://academic.oup.com/mbe/article/38/7/3022/6248099?login=true (accessed on 8 October 2023).
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Wang, J.; Hua, G.; Cai, G.; Ma, Y.; Yang, X.; Zhang, L.; Li, R.; Liu, J.; Ma, Q.; Wu, K.; et al. Genome-Wide DNA Methylation and Transcriptome Analyses Reveal the Key Gene for Wool Type Variation in Sheep. J. Anim. Sci. Biotechnol. 2023, 14, 88. [Google Scholar] [CrossRef]
- Marina, H.; Gutiérrez-Gil, B.; Esteban-Blanco, C.; Suárez-Vega, A.; Pelayo, R.; Arranz, J.J. Analysis of Whole Genome Resequencing Datasets from a Worldwide Sample of Sheep Breeds to Identify Potential Causal Mutations Influencing Milk Composition Traits. Anim. Open Access J. 2020, 10, 1542. [Google Scholar] [CrossRef]
- Li, C.; Li, M.; Li, X.; Ni, W.; Xu, Y.; Yao, R.; Wei, B.; Zhang, M.; Li, H.; Zhao, Y.; et al. Whole-Genome Resequencing Reveals Loci Associated With Thoracic Vertebrae Number in Sheep. Front. Genet. 2019, 10, 674. [Google Scholar] [CrossRef]
- Allais-Bonnet, A.; Hintermann, A.; Deloche, M.-C.; Cornette, R.; Bardou, P.; Naval-Sanchez, M.; Pinton, A.; Haruda, A.; Grohs, C.; Zakany, J.; et al. Analysis of Polycerate Mutants Reveals the Evolutionary Co-Option of HOXD1 for Horn Patterning in Bovidae. Mol. Biol. Evol. 2021, 38, 2260–2272. [Google Scholar] [CrossRef]
- Qiao, G.; Xu, P.; Guo, T.; Wu, Y.; Lu, X.; Zhang, Q.; He, X.; Zhu, S.; Zhao, H.; Lei, Z.; et al. Genetic Basis of Dorper Sheep (Ovis Aries) Revealed by Long-Read De Novo Genome Assembly. Front. Genet. 2022, 13, 846449. [Google Scholar] [CrossRef]
- Luo, R.; Dai, X.; Zhang, L.; Li, G.; Zheng, Z. Genome-Wide DNA Methylation Patterns of Muscle and Tail-Fat in DairyMeade Sheep and Mongolian Sheep. Anim. Open Access J. 2022, 12, 1399. [Google Scholar] [CrossRef] [PubMed]
- Schultz, D.T.; Haddock, S.H.D.; Bredeson, J.V.; Green, R.E.; Simakov, O.; Rokhsar, D.S. Ancient Gene Linkages Support Ctenophores as Sister to Other Animals. Nature 2023, 618, 110. [Google Scholar] [CrossRef] [PubMed]
- Posbergh, C.J.; Staiger, E.A.; Huson, H.J. A Stop-Gain Mutation within MLPH Is Responsible for the Lilac Dilution Observed in Jacob Sheep. Genes 2020, 11, 618. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, X.; Li, F.; Zhang, D.; Zhang, Y.; Li, X.; Song, Q.; Zhou, B.; Zhao, L.; Wang, J.; et al. Whole Genome Sequencing Analysis to Identify Candidate Genes Associated With the Rib Eye Muscle Area in Hu Sheep. Front. Genet. 2022, 13, 824742. [Google Scholar] [CrossRef]
- Guo, Y.; Liang, J.; Lv, C.; Wang, Y.; Wu, G.; Ding, X.; Quan, G. Sequencing Reveals Population Structure and Selection Signatures for Reproductive Traits in Yunnan Semi-Fine Wool Sheep (Ovis Aries). Front. Genet. 2022, 13, 812753. [Google Scholar] [CrossRef]
- Li, X.; Yang, J.; Shen, M.; Xie, X.-L.; Liu, G.-J.; Xu, Y.-X.; Lv, F.-H.; Yang, H.; Yang, Y.-L.; Liu, C.-B.; et al. Whole-Genome Resequencing of Wild and Domestic Sheep Identifies Genes Associated with Morphological and Agronomic Traits. Nat. Commun. 2020, 11, 2815. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Li, S.; Liu, Q.; Wang, Z.; Zhou, Z.; Di, R.; Miao, B.; Hu, W.; Wang, X.; Hu, X.; et al. Whole-Genome Sequences of 89 Chinese Sheep Suggest Role of RXFP2 in the Development of Unique Horn Phenotype as Response to Semi-Feralization. GigaScience 2018, 7, giy019. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinforma. Oxf. Engl. 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and Accurate Long-Read Alignment with Burrows-Wheeler Transform. Bioinforma. Oxf. Engl. 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce Framework for Analyzing next-Generation DNA Sequencing Data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ Data to High Confidence Variant Calls: The Genome Analysis Toolkit Best Practices Pipeline. Curr. Protoc. Bioinforma. 2013, 43, 11.10.1–11.10.33. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A Program for Annotating and Predicting the Effects of Single Nucleotide Polymorphisms, SnpEff. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The Variant Call Format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Dong, S.-S.; He, W.-M.; Ji, J.-J.; Zhang, C.; Guo, Y.; Yang, T.-L. LDBlockShow: A Fast and Convenient Tool for Visualizing Linkage Disequilibrium and Haplotype Blocks Based on Variant Call Format Files. Brief. Bioinform. 2021, 22, bbaa227. [Google Scholar] [CrossRef]
- Lühken, G.; Krebs, S.; Rothammer, S.; Küpper, J.; Mioč, B.; Russ, I.; Medugorac, I. The 1.78-Kb Insertion in the 3′-Untranslated Region of RXFP2 Does Not Segregate with Horn Status in Sheep Breeds with Variable Horn Status. Genet. Sel. Evol. 2016, 48, 78. [Google Scholar] [CrossRef]
- Johnston, S.E.; McEWAN, J.C.; Pickering, N.K.; Kijas, J.W.; Beraldi, D.; Pilkington, J.G.; Pemberton, J.M.; Slate, J. Genome-Wide Association Mapping Identifies the Genetic Basis of Discrete and Quantitative Variation in Sexual Weaponry in a Wild Sheep Population. Mol. Ecol. 2011, 20, 2555–2566. [Google Scholar] [CrossRef]
- Luan, Y.; Wu, S.; Wang, M.; Pu, Y.; Zhao, Q.; Ma, Y.; Jiang, L.; He, X. Identification of Critical Genes for Ovine Horn Development Based on Transcriptome during the Embryonic Period. Biology 2023, 12, 591. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Xi, S.; Cai, B.; Wu, T.; Wang, Q.; Kalds, P.; Huang, S.; Wang, Y.; Han, S.; Pan, M.; et al. Sheep with Partial RXFP2 Knockout Exhibit Normal Horn Phenotype but Unilateral Cryptorchidism1. J. Integr. Agric. 2023; in press. [Google Scholar]
- Edson, M.A.; Nalam, R.L.; Clementi, C.; Franco, H.L.; DeMayo, F.J.; Lyons, K.M.; Pangas, S.A.; Matzuk, M.M. Granulosa Cell-Expressed BMPR1A and BMPR1B Have Unique Functions in Regulating Fertility but Act Redundantly to Suppress Ovarian Tumor Development. Mol. Endocrinol. 2010, 24, 1251–1266. [Google Scholar] [CrossRef]
- Kasiriyan, M.M.; Gholizadeh, M.; Rahimi-Mianji, G.; Moradi, M.H. Selection Signatures Associated with the Number of Lambs per Lambing in Baluchi Ewes. Anim. Prod. Res. 2023, 11, 47–60. [Google Scholar]
- Zielins, E.R.; Paik, K.; Ransom, R.C.; Brett, E.A.; Blackshear, C.P.; Luan, A.; Walmsley, G.G.; Atashroo, D.A.; Senarath-Yapa, K.; Momeni, A.; et al. Enrichment of Adipose-Derived Stromal Cells for BMPR1A Facilitates Enhanced Adipogenesis. Tissue Eng. Part A 2016, 22, 214–221. [Google Scholar] [CrossRef]
- Zhu, X.-J.; Liu, Y.; Dai, Z.-M.; Zhang, X.; Yang, X.; Li, Y.; Qiu, M.; Fu, J.; Hsu, W.; Chen, Y.; et al. BMP-FGF Signaling Axis Mediates Wnt-Induced Epidermal Stratification in Developing Mammalian Skin. PLOS Genet. 2014, 10, e1004687. [Google Scholar] [CrossRef]
- Carré, G.-A.; Couty, I.; Hennequet-Antier, C.; Govoroun, M.S. Gene Expression Profiling Reveals New Potential Players of Gonad Differentiation in the Chicken Embryo. PLoS ONE 2011, 6, e23959. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.C.; Bernard, D.J. Bone Morphogenetic Protein 2 Signals via BMPR1A to Regulate Murine Follicle-Stimulating Hormone Beta Subunit Transcription1. Biol. Reprod. 2009, 81, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Chen, N.; Wang, H.; Li, Q.; Shi, H.; Su, M.; Zhang, Y.; Ma, Y.; Li, T. The Regulatory Role of BMP4 in Testicular Sertoli Cells of Tibetan Sheep. J. Anim. Sci. 2023, 101, skac393. [Google Scholar] [CrossRef]
- Andl, T.; Ahn, K.; Kairo, A.; Chu, E.Y.; Wine-Lee, L.; Reddy, S.T.; Croft, N.J.; Cebra-Thomas, J.A.; Metzger, D.; Chambon, P.; et al. Epithelial Bmpr1a Regulates Differentiation and Proliferation in Postnatal Hair Follicles and Is Essential for Tooth Development. Development 2004, 131, 2257–2268. [Google Scholar] [CrossRef]
- Ming Kwan, K.; Li, A.G.; Wang, X.-J.; Wurst, W.; Behringer, R.R. Essential Roles of BMPR-IA Signaling in Differentiation and Growth of Hair Follicles and in Skin Tumorigenesis. Genesis 2004, 39, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Aretz, S.; Stienen, D.; Uhlhaas, S.; Stolte, M.; Entius, M.M.; Loff, S.; Back, W.; Kaufmann, A.; Keller, K.; Blaas, S.H.; et al. High Proportion of Large Genomic Deletions and a Genotype–Phenotype Update in 80 Unrelated Families with Juvenile Polyposis Syndrome. J. Med. Genet. 2007, 44, 702–709. [Google Scholar] [CrossRef]
- Calva-Cerqueira, D.; Chinnathambi, S.; Pechman, B.; Bair, J.; Larsen-Haidle, J.; Howe, J. The Rate of Germline Mutations and Large Deletions of SMAD4 and BMPR1A in Juvenile Polyposis. Clin. Genet. 2009, 75, 79–85. [Google Scholar] [CrossRef]
- Howe, J.R.; Sayed, M.G.; Ahmed, A.F.; Ringold, J.; Larsen-Haidle, J.; Merg, A.; Mitros, F.A.; Vaccaro, C.A.; Petersen, G.M.; Giardiello, F.M.; et al. The Prevalence of MADH4 and BMPR1A Mutations in Juvenile Polyposis and Absence of BMPR2, BMPR1B, and ACVR1 Mutations. J. Med. Genet. 2004, 41, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Li, W.; Wang, H.; Cao, M.; Jin, Z. The Role of the Smad2/3/4 Signaling Pathway in Osteogenic Differentiation Regulation by ClC-3 Chloride Channels in MC3T3-E1 Cells. J. Orthop. Surg. 2022, 17, 338. [Google Scholar] [CrossRef] [PubMed]
- Huminiecki, L.; Goldovsky, L.; Freilich, S.; Moustakas, A.; Ouzounis, C.; Heldin, C.-H. Emergence, Development and Diversification of the TGF-β Signalling Pathway within the Animal Kingdom. BMC Evol. Biol. 2009, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, A.; Heldin, C.-H. The Regulation of TGFβ Signal Transduction. Development 2009, 136, 3699–3714. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J. TGFβ Signalling in Context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Wakefield, L.M.; Hill, C.S. Beyond TGFβ: Roles of Other TGFβ Superfamily Members in Cancer. Nat. Rev. Cancer 2013, 13, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-β and the TGF-β Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harb. Perspect. Biol. 2016, 8, a021873. [Google Scholar] [CrossRef] [PubMed]
- David, C.J.; Massagué, J. Contextual Determinants of TGFβ Action in Development, Immunity and Cancer. Nat. Rev. Mol. Cell Biol. 2018, 19, 419–435. [Google Scholar] [CrossRef]
- Derynck, R.; Budi, E.H. Specificity, Versatility, and Control of TGF-β Family Signaling. Sci. Signal. 2019, 12, eaav5183. [Google Scholar] [CrossRef]
- Yuan, G.; Zhan, Y.; Gou, X.; Chen, Y.; Yang, G. TGF-β Signaling Inhibits Canonical BMP Signaling Pathway during Palate Development. Cell Tissue Res. 2018, 371, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Mao, Y.; Zhang, B.; Wang, T.; Li, F.; Fu, S.; Xue, Y.; Yang, T.; Wen, X.; Ding, Y.; et al. Chloride Channel ClC-3 Promotion of Osteogenic Differentiation through Runx2. J. Cell. Biochem. 2010, 111, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, F.; Kajiya, H.; Toh, K.; Uchida, S.; Yoshikawa, M.; Sasaki, S.; Kido, M.A.; Tanaka, T.; Okabe, K. Intracellular ClC-3 Chloride Channels Promote Bone Resorption in Vitro through Organelle Acidification in Mouse Osteoclasts. Am. J. Physiol.-Cell Physiol. 2008, 294, C693–C701. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(A) Differential expression of BMPR1A in scurred and normal horn group groups. (B) Expression of BMPR1A exons in scurred and normal horn groups. Expression refers to the fitted expression estimates from the GLM regression, and E001–E015 represents the number of exons. The red line represents the scurred group, whereas the blue line represents the normal horn group. (C) Differential expression of the BMPR1A gene in horn tissues and other tissues. The region labeled in blue represents the overlap between the BMPR1A and MMRN2 genes.
Figure 1.
(A) Differential expression of BMPR1A in scurred and normal horn group groups. (B) Expression of BMPR1A exons in scurred and normal horn groups. Expression refers to the fitted expression estimates from the GLM regression, and E001–E015 represents the number of exons. The red line represents the scurred group, whereas the blue line represents the normal horn group. (C) Differential expression of the BMPR1A gene in horn tissues and other tissues. The region labeled in blue represents the overlap between the BMPR1A and MMRN2 genes.

Figure 2.
(A) Expression of BMPR1A in various tissues of four species. Darker colors and larger dots indicate higher TPM values for the BMPR1A gene in this tissue. (B) Sex differences in the expression of BMPR1A were observed in various tissues of sheep. Asterisks indicate the t-test p-value: * indicates p-value < 0.05, ** indicates p-value < 0.005, **** indicates p-value < 0.00005. (C) Expression of BMPR1A in different sheep breeds.
Figure 2.
(A) Expression of BMPR1A in various tissues of four species. Darker colors and larger dots indicate higher TPM values for the BMPR1A gene in this tissue. (B) Sex differences in the expression of BMPR1A were observed in various tissues of sheep. Asterisks indicate the t-test p-value: * indicates p-value < 0.05, ** indicates p-value < 0.005, **** indicates p-value < 0.00005. (C) Expression of BMPR1A in different sheep breeds.

Figure 3.
(A) Circular evolutionary tree of the BMPR1A protein. (B) Three-dimensional (3D) structure of sheep BMPR1A protein. The 3D structure was predicted via AlphaFold2 with a high model confidence level indicated by the color spectrum ranging from yellow to blue. (C) The amino acid sites of BMPR1A proteins unique to horned animals. Amino acids marked in red are those that differ between horned and hornless animals.
Figure 3.
(A) Circular evolutionary tree of the BMPR1A protein. (B) Three-dimensional (3D) structure of sheep BMPR1A protein. The 3D structure was predicted via AlphaFold2 with a high model confidence level indicated by the color spectrum ranging from yellow to blue. (C) The amino acid sites of BMPR1A proteins unique to horned animals. Amino acids marked in red are those that differ between horned and hornless animals.

Figure 4.
Allele count of 10 ASEs that differed between the scurred and normal horn groups.

Figure 5.
(A) Three-dimensional (3D) principal component analysis (PCA) of the BMPR1A gene. Each point represents a breed of sheep, categorized as either horned or hornless based on their breed information. (B) F-statistics (Fst) of functional loci of the BMPR1A gene in horned and hornless populations.
Figure 5.
(A) Three-dimensional (3D) principal component analysis (PCA) of the BMPR1A gene. Each point represents a breed of sheep, categorized as either horned or hornless based on their breed information. (B) F-statistics (Fst) of functional loci of the BMPR1A gene in horned and hornless populations.

Figure 6.
(A) Box plots of individual horn lengths for different genotypes, with p-values calculated based on the dominant inheritance model and t-test. (B) LD heatmap of the BMPR1A gene. Darker colors represent higher LD values, and black triangles represent LD blocks, which are collections of SNPs with higher LD values. (C) Intersection of ASEs, Fst, and SNPs.
Figure 6.
(A) Box plots of individual horn lengths for different genotypes, with p-values calculated based on the dominant inheritance model and t-test. (B) LD heatmap of the BMPR1A gene. Darker colors represent higher LD values, and black triangles represent LD blocks, which are collections of SNPs with higher LD values. (C) Intersection of ASEs, Fst, and SNPs.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Potential SNP functional loci of BMPR1A.
| INFO | Mutation Type | Position | Fst | Ref | Alt |
|---|---|---|---|---|---|
| SNP1 | intron | 40476340 | 0.214027 | G | A |
| SNP2 | intron | 40468195 | 0.19581 | A | G |
| SNP3 | downstream | 40535217 | 0.0505623 | A | G |
| SNP4 | downstream | 40535673 | 0.0494824 | C | G |
| SNP5 | upstream | 40540201 | 0.0325271 | G | T |
| SNP6 | upstream | 40546207 | 0.040404 | T | G |
| SNP7 | exon | 40563859 | 0.0252525 | A | G |
| SNP8 | exon | 40572996 | 0.0019107 | G | A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, G.; Chu, M.; Yang, H.; Li, H.; Shi, J.; Feng, P.; Wang, S.; Pan, Z. Expression, Polymorphism, and Potential Functional Sites of the BMPR1A Gene in the Sheep Horn. Genes 2024, 15, 376. https://doi.org/10.3390/genes15030376
AMA Style
Zhang G, Chu M, Yang H, Li H, Shi J, Feng P, Wang S, Pan Z. Expression, Polymorphism, and Potential Functional Sites of the BMPR1A Gene in the Sheep Horn. Genes. 2024; 15(3):376. https://doi.org/10.3390/genes15030376
Chicago/Turabian StyleZhang, Guoqing, Mingxing Chu, Hao Yang, Hao Li, Jianxin Shi, Pingjie Feng, Shoufeng Wang, and Zhangyuan Pan. 2024. "Expression, Polymorphism, and Potential Functional Sites of the BMPR1A Gene in the Sheep Horn" Genes 15, no. 3: 376. https://doi.org/10.3390/genes15030376
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.