
Preliminary Study on the Pathogenic Mechanism of Jujube Flower Disease in Honeybees (Apis mellifera ligustica) Based on Midgut Transcriptomics
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tested Honeybee
2.2. Morphological Observation
2.3. Enzyme Activity Assay
2.4. cDNA Library Construction and Illumina Sequencing
2.5. Validation of DEGs Using qRT-PCR
2.6. Statistical Analyses
3. Results
3.1. Effect of Jujube Flower Disease on Honeybee Midgut Tissue
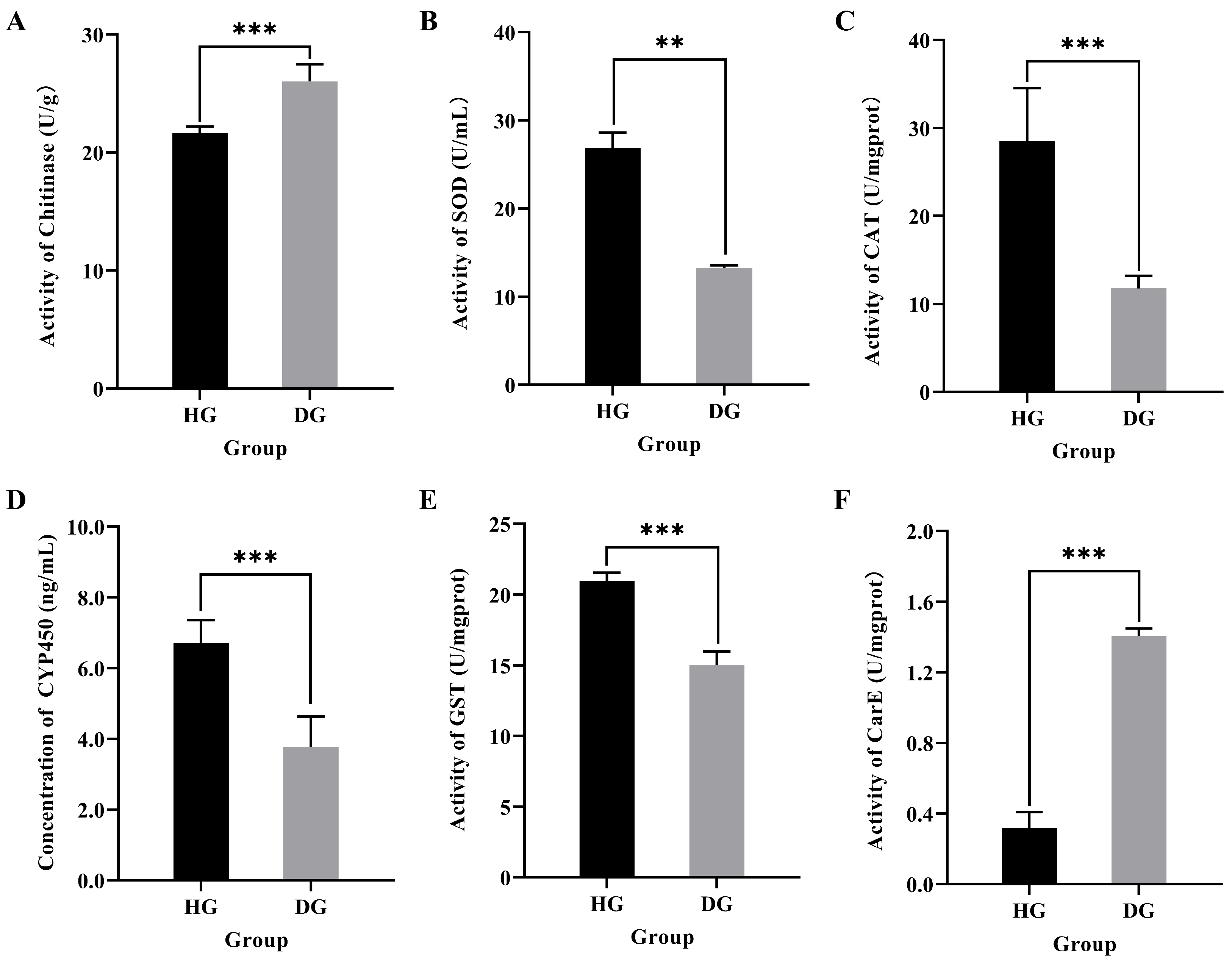
3.2. Effect of Jujube Flower Disease on Antioxidant and Detoxification Capacity in Honeybee Midgut
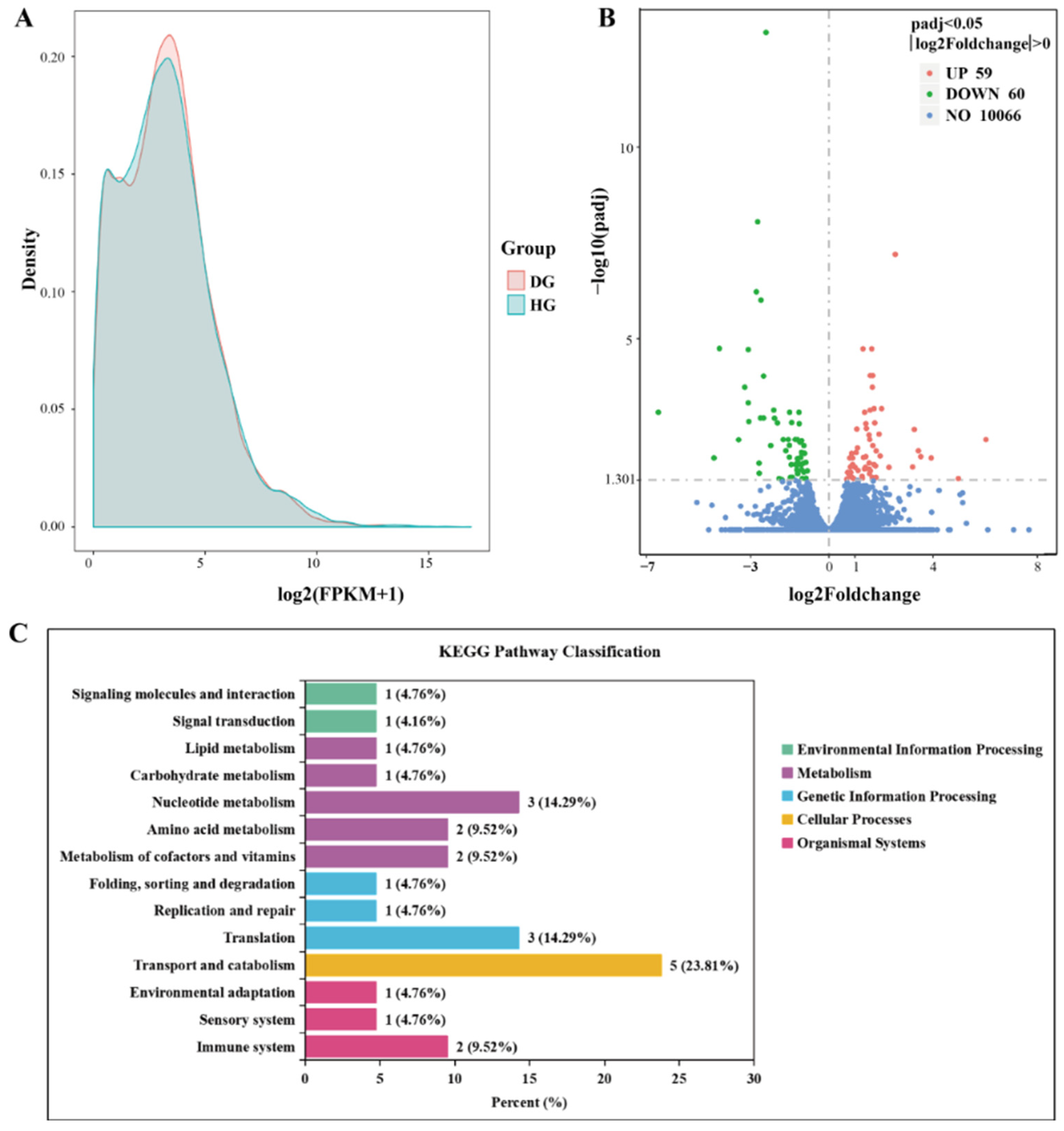
3.3. Transcriptome Analysis
3.4. GO and KEGG Enrichment of DEGs
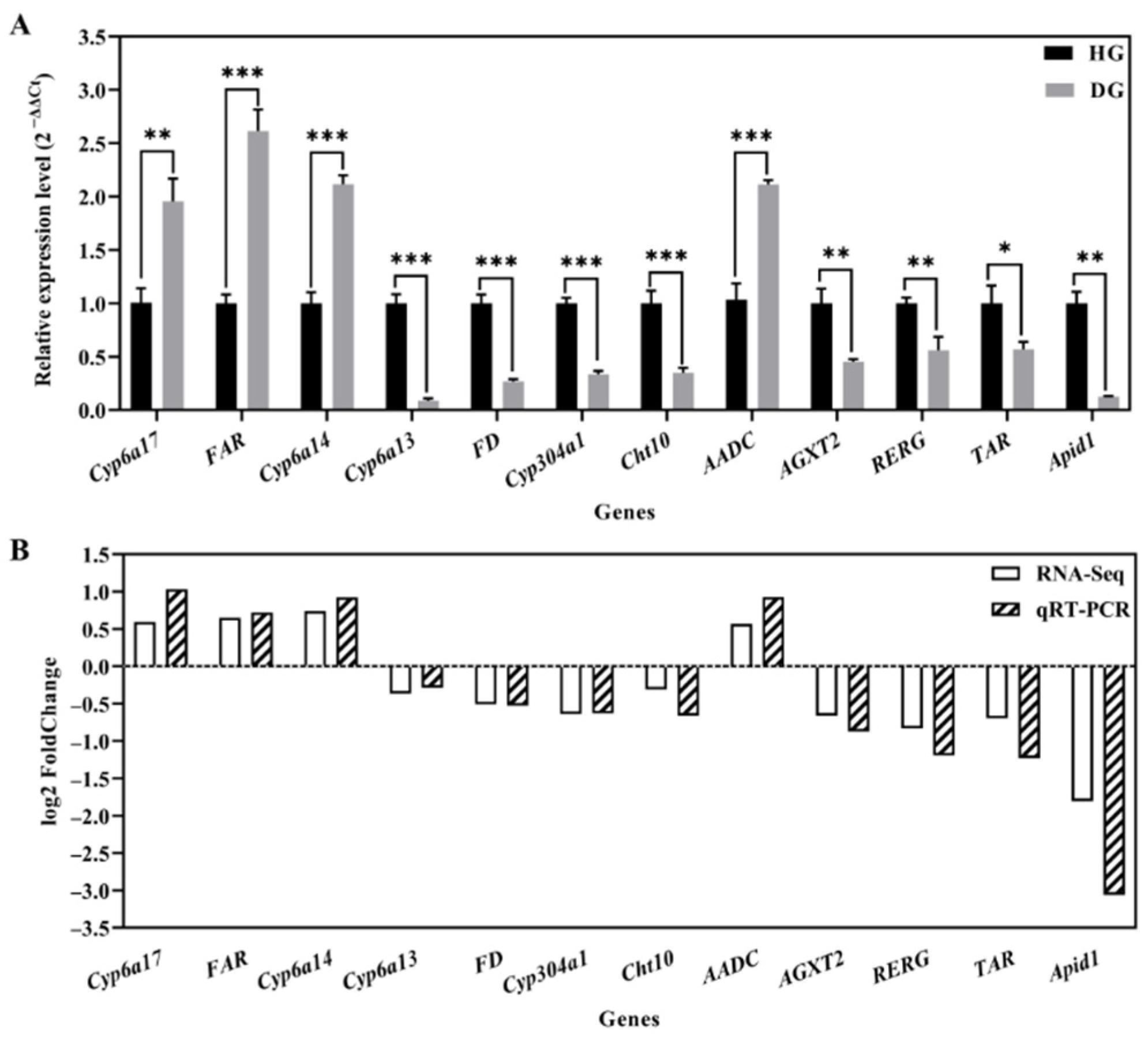
3.5. Transcriptome Verification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, M.; Wang, J.; Wang, L.; Liu, P.; Zhao, J.; Zhao, Z.; Yao, S.; Stănică, F.; Liu, Z.; Wang, L.; et al. The Historical and Current Research Progress on Jujube–a Superfruit for the Future. Hortic. Res. 2020, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Q. The Prevention Management Measures of Honeybee Jujube Flower Disease. Shandong Anim. Husb. Vet. 2016, 37, 68–69. [Google Scholar]
- Lu, X.; Diao, H.X.; Cheng, J. The Diagnosis of Mixed Infection from Jujube Flowers Bees Poisoning and Escherichia coli Disease. Anim. Husb. Vet. Sci. Technol. 2017, 33, 123–124. [Google Scholar]
- Ma, W.H.; Shen, J.S.; Guo, Y.; Zhang, Y.Y.; Shao, Y.Q. Effects of Zaohua Disease on Activities of Digestive Enzymes and Detoxification Enzymes in Apis mellifera ligustica. Acta Entomol. Sci. 2011, 54, 1076–1081. [Google Scholar]
- Yu, H.B. The Etiology and Diagnosis of Bee Poisoning by Toxic Nectars and Pollens. Apic. China 2014, 65, 32–34. [Google Scholar]
- Zhang, X.G. Prevention and Treatment of Jujube Flower Disease. Apic. China 1959, 10, 43. [Google Scholar]
- Xi, G. Discussion on Jujube Flower Disease. Apic. China 1960, 11, 274–275. [Google Scholar]
- Li, X.L.; Fan, Z.Y. Discussion on the Cause of Bee Jujube Flower Disease. Apic. China 1991, 42, 5–8. [Google Scholar]
- Jin, F. Prevention and Treatment of Bee Poisoning during Jujube Flowering. Apic. China 1994, 45, 22. [Google Scholar]
- Ma, W.H.; Zheng, X.; Li, L.; Shen, J.S.; Li, W.; Gao, Y. Changes in the Gut Microbiota of Honey Bees Associated with Jujube Flower Disease. Ecotoxicol. Environ. Saf. 2020, 198, 110616. [Google Scholar] [CrossRef]
- Du, Y.L.; Xu, K.; Zheng, L.F.; Liu, Y.L.; Jiang, Y.S. Study on the Pathogenesis of Jujube Flower Disease of Apis mellifera liguatica Workers Based on Midgut Metabolomics. Acta Entomol. Sci. 2023, 66, 1171–1182. [Google Scholar]
- Sun, Q.X.; Li, X.C.; Tan, X.H.; You, C.H.; Dong, Y.W.; Gao, H.Y.; Huang, Y.H.; Zhou, M. Observations on Morphology and Histology of Digestive Tract in Black Soldier Fly Hermetia illucens. J. Environ. Entomol. 2023, 45, 1463–1475. [Google Scholar]
- Zheng, H.; Steele, M.I.; Leonard, S.P.; Motta, E.V.S.; Moran, N.A. Honey Bees as Models for Gut Microbiota Research. Lab. Anim. 2018, 47, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Xiong, M.Q.; Qin, G.; Wang, L.Z.; Huang, S.K.; Li, J.H.; Duan, X.L. Toxic Effects of Fungicides on Physiology and Behavior of Honeybee. Asian J. Ecotoxicol. 2022, 17, 163–175. [Google Scholar]
- Dai, P.; Yan, Z.; Ma, S.; Yang, Y.; Wang, Q.; Hou, C.; Wu, Y.; Liu, Y.; Diao, Q. The Herbicide Glyphosate Negatively Affects Midgut Bacterial Communities and Survival of Honey Bee during Larvae Reared in Vitro. J. Agric. Food Chem. 2018, 66, 7786–7793. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Z.; Ma, L.; Li, G.; Han, K.; Liu, Z.; Wang, H.; Xu, B. The Native Dietary Habits of the Two Sympatric Bee Species and Their Effects on Shaping Midgut Microorganisms. Front. Microbiol. 2021, 12, 738226. [Google Scholar] [CrossRef]
- Konno, K.; Mitsuhashi, W. The Peritrophic Membrane as a Target of Proteins That Play Important Roles in Plant Defense and Microbial Attack. J. Insect Physiol. 2019, 117, 103912. [Google Scholar] [CrossRef]
- Terra, W.R. The Origin and Functions of the Insect Peritrophic Membrane and Peritrophic Gel. Arch. Insect Biochem. Physiol. 2001, 47, 47–61. [Google Scholar] [CrossRef]
- Rabadiya, D.; Behr, M. The Biology of Insect Chitinases and Their Roles at Chitinous Cuticles. Insect Biochem. Mol. Biol. 2024, 165, 104071. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Zhou, Y.; Chen, L.; Yang, R.K.; Feng, L.C. Structural Changes and Its Relationship with Activity of Related Enzymes in Silkworm Peritrophic Membrane during Regeneration. Sci. Seric. 2019, 45, 921–926. [Google Scholar]
- Rzezniczak, T.Z.; Douglas, L.A.; Watterson, J.H.; Merritt, T.J.S. Paraquat Administration in Drosophila for Use in Metabolic Studies of Oxidative Stress. Anal. Biochem. 2011, 419, 345–347. [Google Scholar] [CrossRef] [PubMed]
- Taric, E.; Glavinic, U.; Vejnovic, B.; Stanojkovic, A.; Aleksic, N.; Dimitrijevic, V.; Stanimirovic, Z. Oxidative Stress, Endoparasite Prevalence and Social Immunity in Bee Colonies Kept Traditionally vs. Those Kept for Commercial Purposes. Insects 2020, 11, 266. [Google Scholar] [CrossRef] [PubMed]
- Rand, E.E.D.; Smit, S.; Beukes, M.; Apostolides, Z.; Pirk, C.W.W.; Nicolson, S.W. Detoxification Mechanisms of Honey Bees (Apis mellifera) Resulting in Tolerance of Dietary Nicotine. Sci. Rep. 2015, 5, 11779. [Google Scholar] [CrossRef] [PubMed]
- Su, B.; Gao, K.K.; Wang, R.M.; Yang, X.H. Research Advances of Cytochrome P450 in Honeybees. J. Qilu Univ. Technol. 2022, 36, 1–6. [Google Scholar]
- Dermauw, W.; Van Leeuwen, T.; Feyereisen, R. Diversity and Evolution of the P450 Family in Arthropods. Insect Biochem. Mol. Biol. 2020, 127, 103490. [Google Scholar] [CrossRef] [PubMed]
- Ueland, P.M.; McCann, A.; Midttun, Ø.; Ulvik, A. Inflammation, Vitamin B6 and Related Pathways. Mol. Asp. Med. 2017, 53, 10–27. [Google Scholar] [CrossRef]
- Han, S.W.; Shin, J.S. Aromatic L-Amino Acid Decarboxylases: Mechanistic Features and Microbial Applications. Appl. Microbiol. Biotechnol. 2022, 106, 4445–4458. [Google Scholar] [CrossRef] [PubMed]
- Tanianskii, D.; Jarzebska, N.; Birkenfeld, A.; O’Sullivan, J.; Rodionov, R. Beta-Aminoisobutyric Acid as a Novel Regulator of Carbohydrate and Lipid Metabolism. Nutrients 2019, 11, 524. [Google Scholar] [CrossRef] [PubMed]
- Rodionov, R.N.; Jarzebska, N.; Weiss, N.; Lentz, S.R. AGXT2: A Promiscuous Aminotransferase. Trends Pharmacol. Sci. 2014, 35, 575–582. [Google Scholar] [CrossRef]
- Rodionov, R.N.; Jarzebska, N.; Burdin, D.; Todorov, V.; Martens-Lobenhoffer, J.; Hofmann, A.; Kolouschek, A.; Cordasic, N.; Jacobi, J.; Rubets, E.; et al. Overexpression of Alanine-Glyoxylate Aminotransferase 2 Protects from Asymmetric Dimethylarginine-Induced Endothelial Dysfunction and Aortic Remodeling. Sci. Rep. 2022, 12, 9381. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | ID | GO Description | p-Value | Upregulated Genes | Downregulated Genes |
|---|---|---|---|---|---|
| Molecular Function | GO:0005506 | iron ion binding | 0.001551 | cytochrome P450 6a17 (LOC412209) cytochrome P450 6a14 (LOC112935903) | cytochrome P450 6a13 (LOC112939925) cytochrome P450 304a1 (LOC724175) |
| GO:0016705 | oxidoreductase activity, acting on paired donors, with incorporation or reduction of molecular oxygen | 0.001905 | cytochrome P450 6a17 (LOC412209) cytochrome P450 6a14 (LOC112935903) | cytochrome P450 6a13 (LOC112939925) cytochrome P450 304a1 (LOC724175) | |
| GO:0020037 | heme binding | 0.002311 | cytochrome P450 6a17 (LOC412209) cytochrome P450 6a14 (LOC112935903) | cytochrome P450 6a13 (LOC112939925) cytochrome P450 304a1 (LOC724175) | |
| GO:0046906 | tetrapyrrole binding | 0.002459 | cytochrome P450 6a17 (LOC412209) cytochrome P450 6a14 (LOC112935903) | cytochrome P450 6a13 (LOC112939925) cytochrome P450 304a1 (LOC724175) | |
| GO:0048037 | cofactor binding | 0.009924 | cytochrome P450 6a17 (LOC412209) aromatic-L-amino-acid decarboxylase (LOC410638) cytochrome P450 6a14 (LOC112935903) | cytochrome P450 6a13 (LOC112939925) cytochrome P450 304a1 (LOC724175) Alanine-glyoxylate aminotransferase 2 (LOC408817) | |
| GO:0030170 | pyridoxal phosphate binding | 0.039579 | aromatic-L-amino-acid decarboxylase (LOC410638) | Alanine-glyoxylate aminotransferase 2 (LOC408817) | |
| GO:0070279 | vitamin B6 binding | 0.039579 | aromatic-L-amino-acid decarboxylase (LOC410638) | Alanine-glyoxylate aminotransferase 2 (LOC408817) | |
| GO:0019842 | vitamin binding | 0.046380 | aromatic-L-amino-acid decarboxylase (LOC410638) | Alanine-glyoxylate aminotransferase 2 (LOC408817) | |
| Biological Process | GO:0007155 | cell adhesion | 0.017147 | Vinculin (LOC552082) | Nidogen-2 (LOC408797) |
| GO:0022610 | biological adhesion | 0.017147 | Vinculin (LOC552082) | Nidogen-2 (LOC408797) |
| KEGG ID | Pathway Name | p-Value | Upregulated Genes | Downregulated Genes |
|---|---|---|---|---|
| ame00130 | Ubiquinone and other terpenoid-quinone biosynthesis | 0.099986 | — | 4-coumarate--CoA ligase 1 (LOC726625) |
| ame00350 | Tyrosine metabolism | 0.107270 | Aromatic-L-amino-acid decarboxylase (LOC410638) | — |
| ame00760 | Nicotinate and nicotinamide metabolism | 0.128784 | — | Purine nucleoside phosphorylase (LOC408299) |
| ame00380 | Tryptophan metabolism | 0.163549 | Aromatic-L-amino-acid decarboxylase (LOC410638) | — |
| ame00240 | Pyrimidine metabolism | 0.210006 | — | Purine nucleoside phosphorylase (LOC408299) |
| ame00270 | Cysteine and methionine metabolism | 0.283985 | — | Methylthioribose-1-phosphate isomerase (LOC409023) |
| ame00010 | Glycolysis/Gluconeogenesis | 0.318452 | Multiple inositol polyphosphate phosphatase 1 (LOC409751) | — |
| ame00564 | Glycerophospholipid metabolism | 0.392835 | — | Alanine--glyoxylate aminotransferase 2 (LOC408817) |
| ame00562 | Inositol phosphate metabolism | 0.318452 | Multiple inositol polyphosphate phosphatase 1 (LOC409751) | — |
| ame00230 | Purine metabolism | 0.110121 | — | Purine nucleoside phosphorylase (LOC408299) Bifunctional 3′-phosphoadenosine 5′-phosphosulfate synthase 2 (LOC408299) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.; Xu, K.; Zhao, H.; Wu, Y.; Jiang, H.; He, J.; Jiang, Y. Preliminary Study on the Pathogenic Mechanism of Jujube Flower Disease in Honeybees (Apis mellifera ligustica) Based on Midgut Transcriptomics. Genes 2024, 15, 533. https://doi.org/10.3390/genes15050533
Du Y, Xu K, Zhao H, Wu Y, Jiang H, He J, Jiang Y. Preliminary Study on the Pathogenic Mechanism of Jujube Flower Disease in Honeybees (Apis mellifera ligustica) Based on Midgut Transcriptomics. Genes. 2024; 15(5):533. https://doi.org/10.3390/genes15050533
Chicago/Turabian StyleDu, Yali, Kai Xu, Huiting Zhao, Ying Wu, Haibin Jiang, Jinming He, and Yusuo Jiang. 2024. "Preliminary Study on the Pathogenic Mechanism of Jujube Flower Disease in Honeybees (Apis mellifera ligustica) Based on Midgut Transcriptomics" Genes 15, no. 5: 533. https://doi.org/10.3390/genes15050533