The Past, Present, and Future of Human Centromere Genomics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
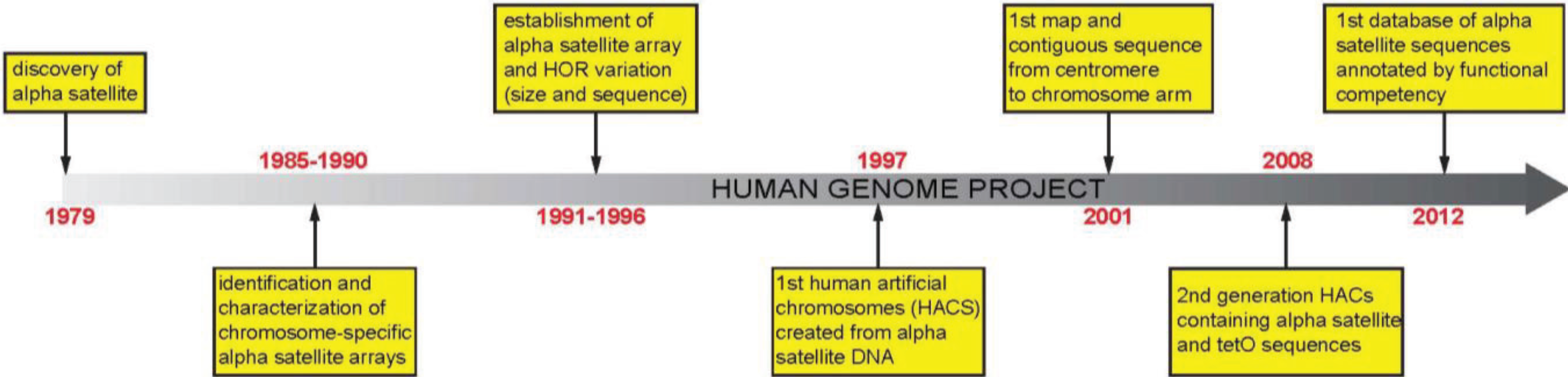
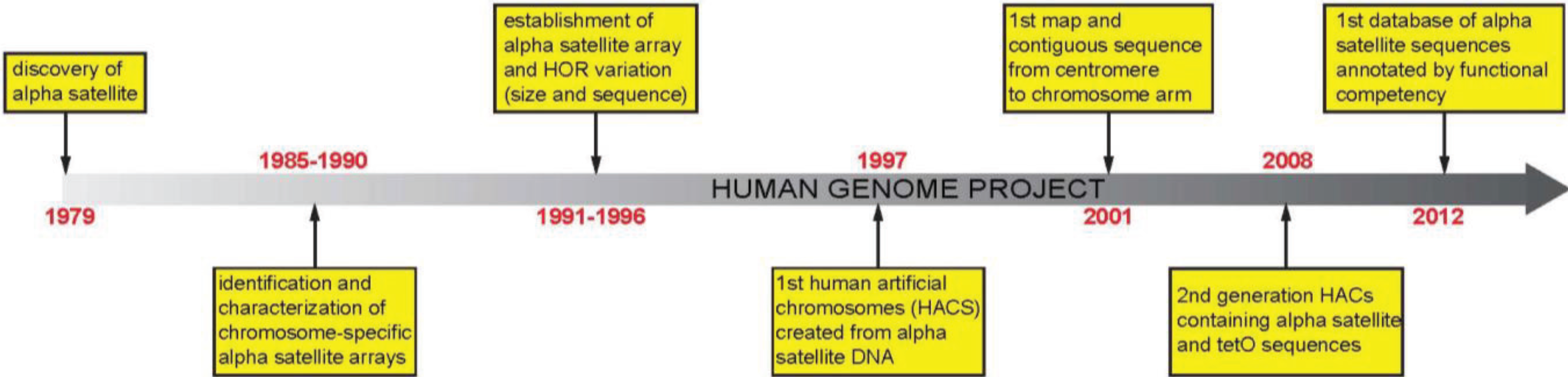
:1. Introduction
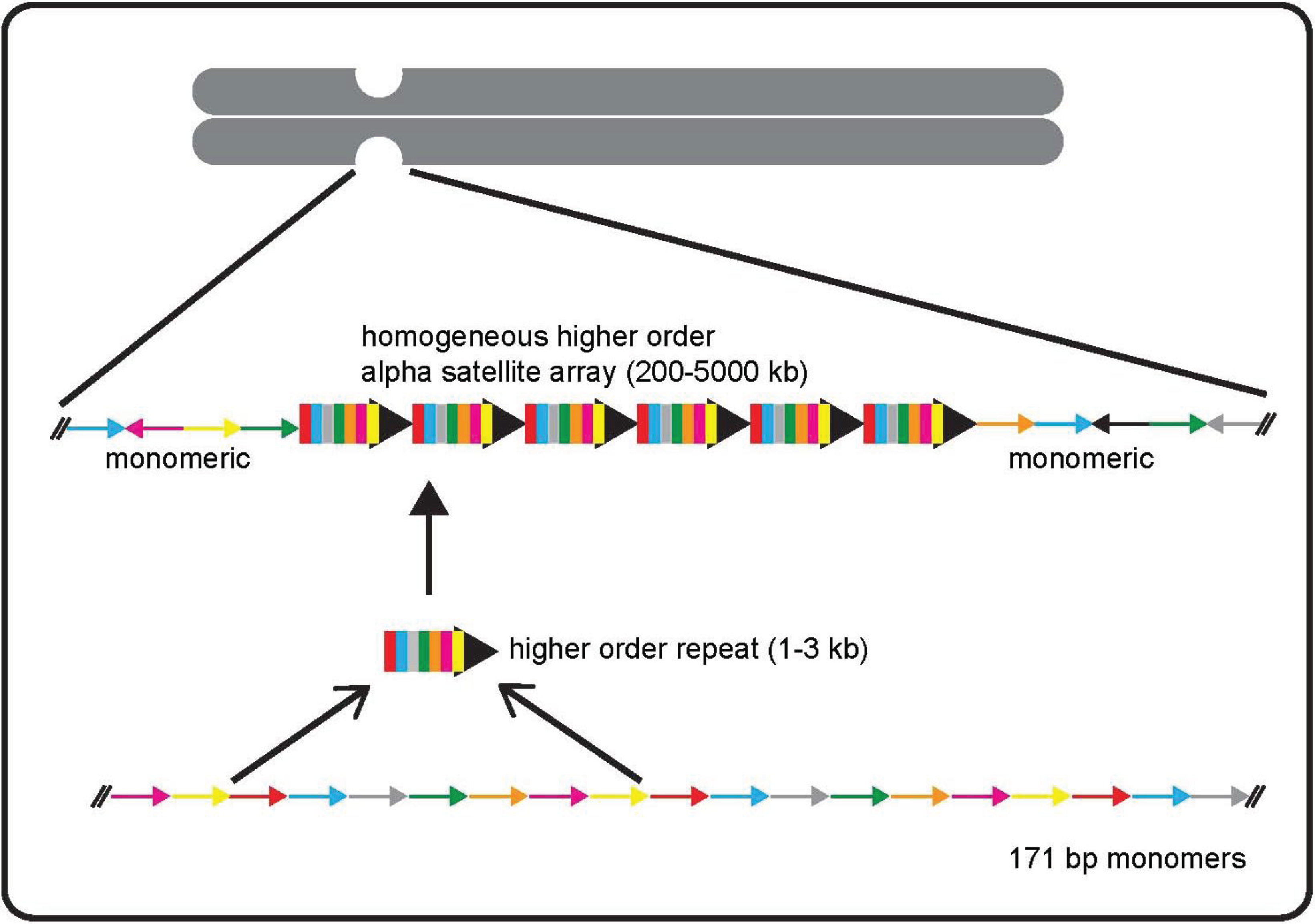
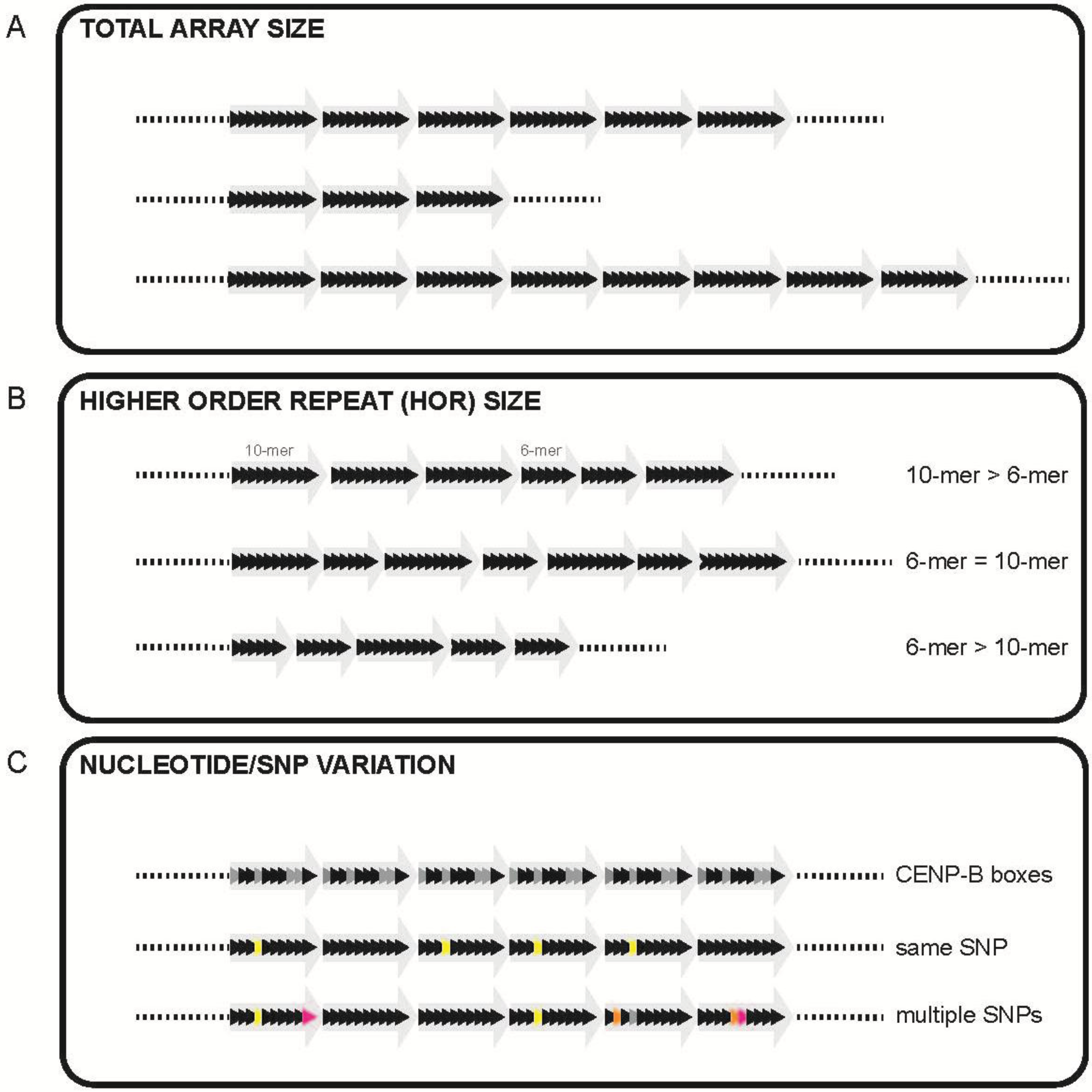
2. Alpha Satellite DNA: Discovery, Organization, and Variation
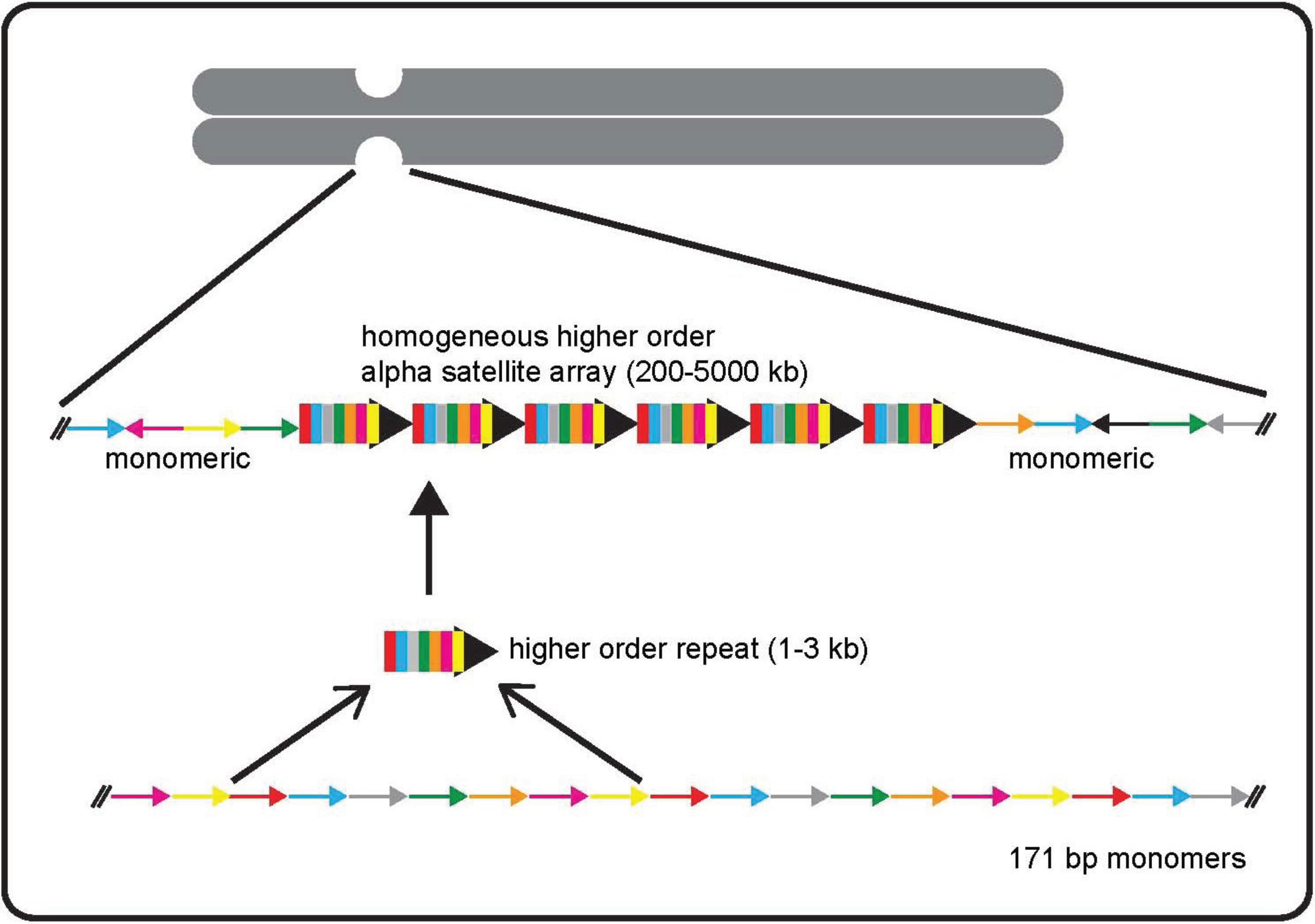

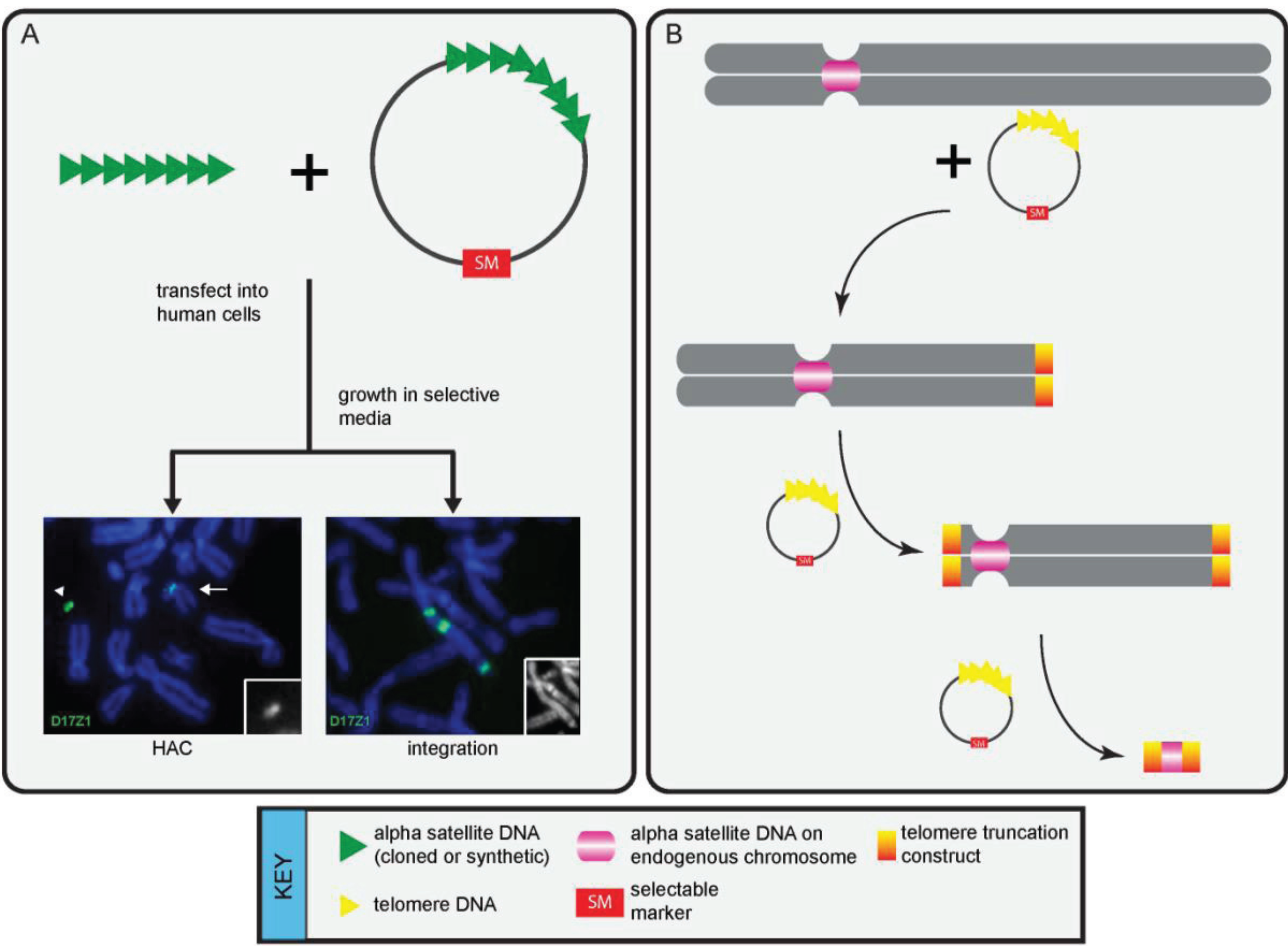
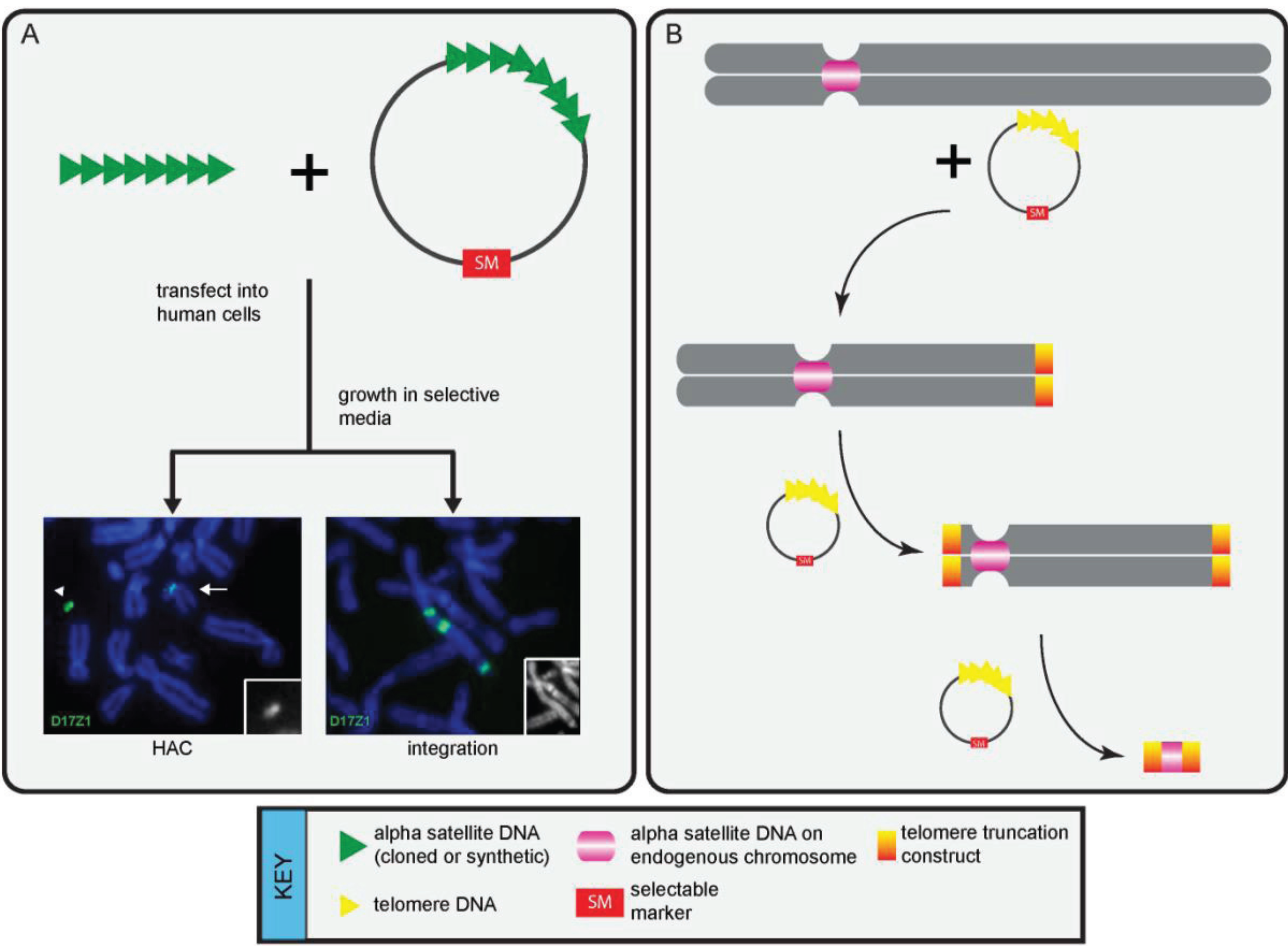
3. Functional Studies that Have Defined Genomic Centromeres
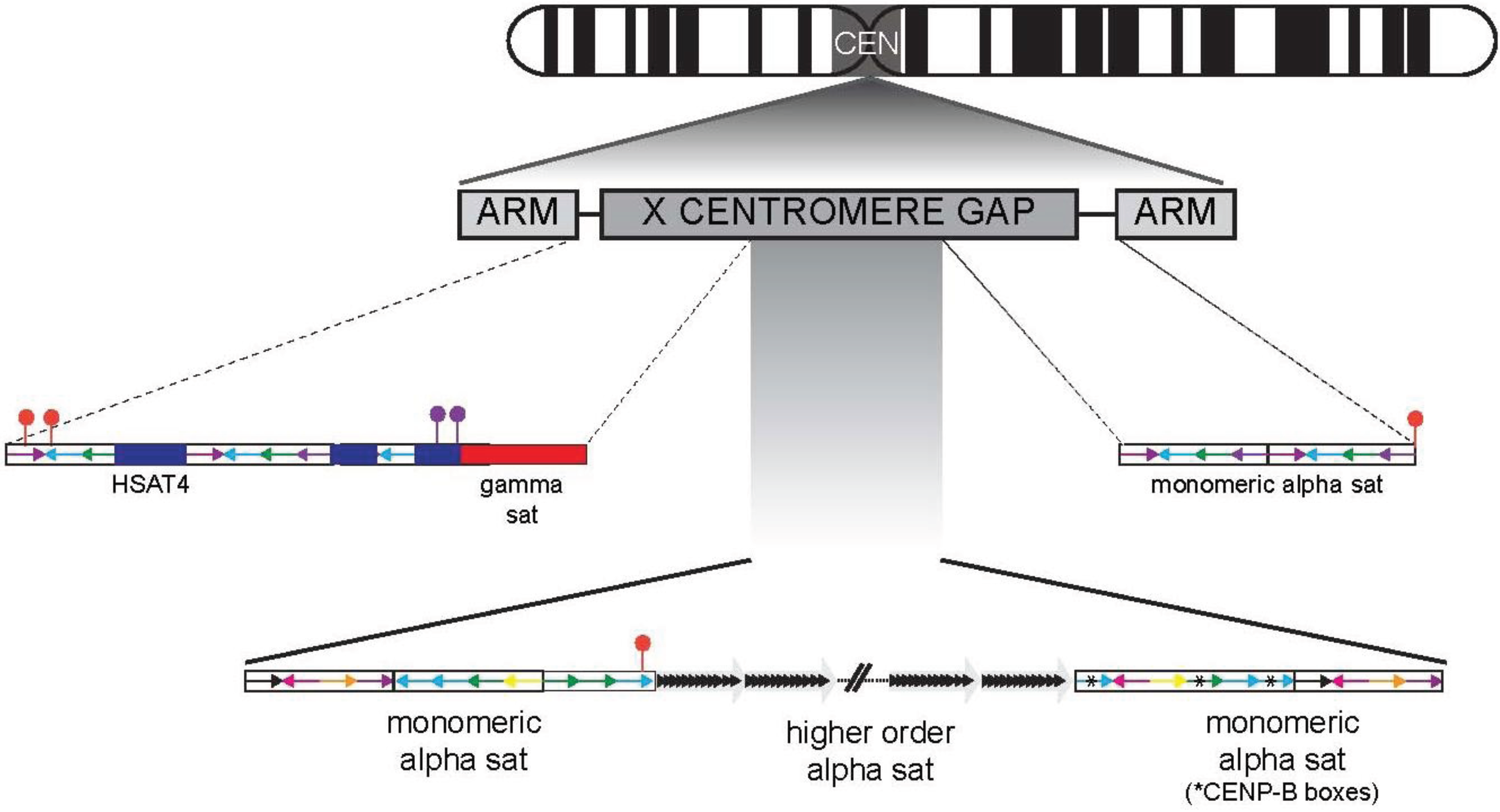
4. Centromere Regions in the Human Genome Project Era
5. Linking Physical and Genetic Maps of Human Centromeres
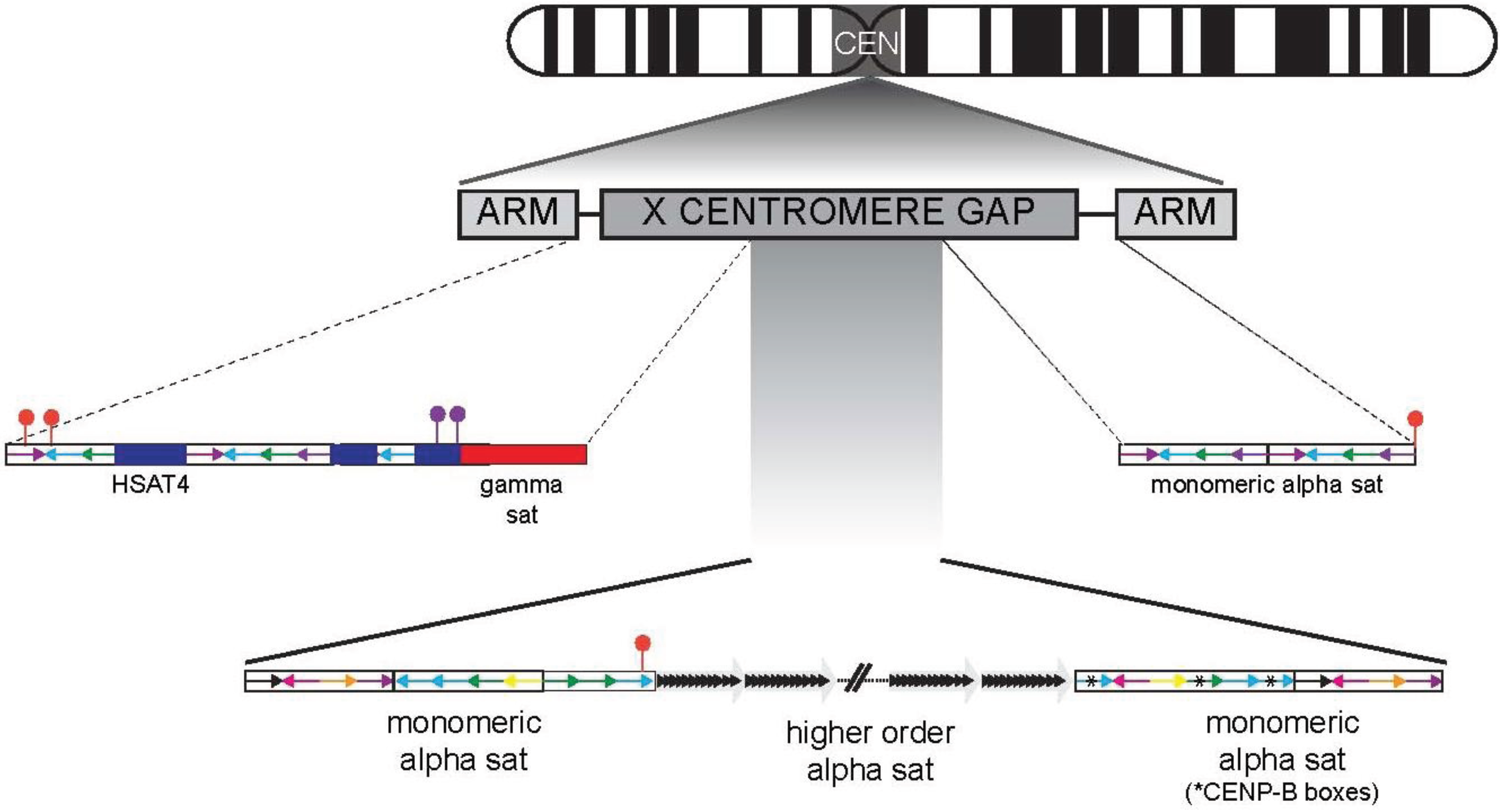
6. Correlating the Genetic and Functional Centromere
7. The Computational Challenge of Alpha Satellite Genome Assemblies
8. Assembling Centromeres in the Present Day
9. Future Perspectives
9.1. Centromere Assemblies
9.2. Centromeric Variation and Functional Capacity
9.3. Maps of Functional Centromeric Domains
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of interest
References
- Panchenko, T.; Black, B.E. The epigenetic basis for centromere identity. Prog. Mol. Subcell. Biol. 2009, 48, 1–32. [Google Scholar] [CrossRef]
- Valente, L.P.; Silva, M.C.; Jansen, L.E. Temporal control of epigenetic centromere specification. Chromosome Res. 2012, 20, 481–492. [Google Scholar] [CrossRef]
- Choo, K.H. Domain organization at the centromere and neocentromere. Dev. Cell 2001, 1, 165–177. [Google Scholar] [CrossRef]
- Warburton, P.E. Chromosomal dynamics of human neocentromere formation. Chromosome Res. 2004, 12, 617–626. [Google Scholar] [CrossRef]
- Manuelidis, L.; Wu, J.C. Homology between human and simian repeated DNA. Nature 1978, 276, 92–94. [Google Scholar] [CrossRef]
- Waye, J.S.; Willard, H.F. Nucleotide sequence heterogeneity of alpha satellite repetitive DNA: A survey of alphoid sequences from different human chromosomes. Nucleic Acids Res. 1987, 15, 7549–7569. [Google Scholar] [CrossRef]
- Willard, H.F. Chromosome-specific organization of human alpha satellite DNA. Am. J. Hum. Genet. 1985, 37, 524–532. [Google Scholar]
- Muro, Y.; Masumoto, H.; Yoda, K.; Nozaki, N.; Ohashi, M.; Okazaki, T. Centromere protein B assembles human centromeric alpha-satellite DNA at the 17-bp sequence, CENP-B box. J. Cell Biol. 1992, 116, 585–596. [Google Scholar] [CrossRef]
- Schueler, M.G.; Sullivan, B.A. Structural and functional dynamics of human centromeric chromatin. Annu. Rev. Genomics Hum. Genet. 2006, 7, 301–313. [Google Scholar] [CrossRef]
- Choo, K.H.; Vissel, B.; Nagy, A.; Earle, E.; Kalitsis, P. A survey of the genomic distribution of alpha satellite DNA on all the human chromosomes, and derivation of a new consensus sequence. Nucleic Acids Res. 1991, 19, 1179–1182. [Google Scholar] [CrossRef]
- Vissel, B.; Choo, K.H. Human alpha satellite DNA—Consensus sequence and conserved regions. Nucleic Acids Res. 1987, 15, 6751–6752. [Google Scholar] [CrossRef]
- Alexandrov, I.A.; Mitkevich, S.P.; Yurov, Y.B. The phylogeny of human chromosome specific alpha satellites. Chromosoma 1988, 96, 443–453. [Google Scholar] [CrossRef]
- Alexandrov, I.; Kazakov, A.; Tumeneva, I.; Shepelev, V.; Yurov, Y. Alpha-satellite DNA of primates: Old and new families. Chromosoma 2001, 110, 253–266. [Google Scholar] [CrossRef]
- Devilee, P.; Kievits, T.; Waye, J.S.; Pearson, P.L.; Willard, H.F. Chromosome-specific alpha satellite DNA: Isolation and mapping of a polymorphic alphoid repeat from human chromosome 10. Genomics 1988, 3, 1–7. [Google Scholar] [CrossRef]
- Mahtani, M.M.; Willard, H.F. Pulsed-field gel analysis of alpha-satellite DNA at the human X chromosome centromere: High-frequency polymorphisms and array size estimate. Genomics 1990, 7, 607–613. [Google Scholar] [CrossRef]
- Greig, G.M.; Parikh, S.; George, J.; Powers, V.E.; Willard, H.F. Molecular cytogenetics of alpha satellite DNA from chromosome 12: Fluorescence in situ hybridization and description of DNA and array length polymorphisms. Cytogenet. Cell Genet. 1991, 56, 144–148. [Google Scholar] [CrossRef]
- Wevrick, R.; Willard, H.F. Long-range organization of tandem arrays of alpha satellite DNA at the centromeres of human chromosomes: High-frequency array-length polymorphism and meiotic stability. Proc. Natl. Acad. Sci. USA 1989, 86, 9394–9398. [Google Scholar] [CrossRef]
- Willard, H.F.; Waye, J.S.; Skolnick, M.H.; Schwartz, C.E.; Powers, V.E.; England, S.B. Detection of restriction fragment length polymorphisms at the centromeres of human chromosomes by using chromosome-specific alpha satellite DNA probes: Implications for development of centromere-based genetic linkage maps. Proc. Natl. Acad. Sci. USA 1986, 83, 5611–5615. [Google Scholar] [CrossRef]
- Abruzzo, M.A.; Griffin, D.K.; Millie, E.A.; Sheean, L.A.; Hassold, T.J. The effect of Y-chromosome alpha-satellite array length on the rate of sex chromosome disomy in human sperm. Hum. Genet. 1996, 97, 819–823. [Google Scholar] [CrossRef]
- Oakey, R.; Tyler-Smith, C. Y chromosome DNA haplotyping suggests that most European and Asian men are descended from one of two males. Genomics 1990, 7, 325–330. [Google Scholar] [CrossRef]
- Willard, H.F. Evolution of alpha satellite. Curr. Opin. Genet. Dev. 1991, 1, 509–514. [Google Scholar] [CrossRef]
- Waye, J.S.; Willard, H.F. Molecular analysis of a deletion polymorphism in alpha satellite of human chromosome 17: Evidence for homologous unequal crossing-over and subsequent fixation. Nucleic Acids Res. 1986, 14, 6915–6927. [Google Scholar] [CrossRef]
- Waye, J.S.; Willard, H.F. Structure, organization, and sequence of alpha satellite DNA from human chromosome 17: Evidence for evolution by unequal crossing-over and an ancestral pentamer repeat shared with the human X chromosome. Mol. Cell. Biol. 1986, 6, 3156–3165. [Google Scholar]
- Willard, H.F.; Greig, G.M.; Powers, V.E.; Waye, J.S. Molecular organization and haplotype analysis of centromeric DNA from human chromosome 17: Implications for linkage in neurofibromatosis. Genomics 1987, 1, 368–373. [Google Scholar] [CrossRef]
- Warburton, P.E.; Willard, H.F. Interhomologue sequence variation of alpha satellite DNA from human chromosome 17: Evidence for concerted evolution along haplotypic lineages. J. Mol. Evol. 1995, 41, 1006–1015. [Google Scholar]
- Maloney, K.A.; Sullivan, L.L.; Matheny, J.E.; Strome, E.D.; Merrett, S.L.; Ferris, A.; Sullivan, B.A. Functional epialleles at an endogenous human centromere. Proc. Natl. Acad. Sci. USA 2012, 109, 13704–13709. [Google Scholar] [CrossRef]
- Brown, K.E.; Barnett, M.A.; Burgtorf, C.; Shaw, P.; Buckle, V.J.; Brown, W.R. Dissecting the centromere of the human Y chromosome with cloned telomeric DNA. Hum. Mol. Genet. 1994, 3, 1227–1237. [Google Scholar] [CrossRef]
- Farr, C.J.; Bayne, R.A.; Kipling, D.; Mills, W.; Critcher, R.; Cooke, H.J. Generation of a human X-derived minichromosome using telomere-associated chromosome fragmentation. EMBO J. 1995, 14, 5444–5454. [Google Scholar]
- Harrington, J.J.; van Bokkelen, G.; Mays, R.W.; Gustashaw, K.; Willard, H.F. Formation of de novo centromeres and construction of first-generation human artificial microchromosomes. Nat. Genet. 1997, 15, 345–355. [Google Scholar] [CrossRef]
- Ikeno, M.; Grimes, B.R.; Okazaki, T.; Nakano, M.; Saitoh, K.; Hoshino, H.; McGill, N.I.; Cooke, H.; Masumoto, H. Construction of YAC-based mammalian artificial chromosomes. Nat. Biotechnol. 1998, 16, 431–439. [Google Scholar] [CrossRef]
- Grimes, B.R.; Babcock, J.; Rudd, M.K.; Chadwick, B.; Willard, H.F. Assembly and characterizationof heterochromatin and euchromatin on human artificial chromosomes. Genome Biol. 2004, 5, R89. [Google Scholar] [CrossRef] [Green Version]
- Grimes, B.R.; Rhoades, A.A.; Willard, H.F. Alpha-satellite DNA and vector composition influence rates of human artificial chromosome formation. Mol. Ther. 2002, 5, 798–805. [Google Scholar] [CrossRef]
- Ohzeki, J.; Nakano, M.; Okada, T.; Masumoto, H. CENP-B box is required for de novo centromere chromatin assembly on human alphoid DNA. J. Cell Biol. 2002, 159, 765–775. [Google Scholar] [CrossRef]
- Haaf, T.; Warburton, P.E.; Willard, H.F. Integration of human alpha-satellite DNA into simian chromosomes: Centromere protein binding and disruption of normal chromosome segregation. Cell 1992, 70, 681–696. [Google Scholar] [CrossRef]
- Nakashima, H.; Nakano, M.; Ohnishi, R.; Hiraoka, Y.; Kaneda, Y.; Sugino, A.; Masumoto, H. Assembly of additional heterochromatin distinct from centromere-kinetochore chromatin is required for de novo formation of human artificial chromosome. J. Cell Sci. 2005, 118, 5885–5898. [Google Scholar] [CrossRef]
- Nakano, M.; Cardinale, S.; Noskov, V.N.; Gassmann, R.; Vagnarelli, P.; Kandels-Lewis, S.; Larionov, V.; Earnshaw, W.C.; Masumoto, H. Inactivation of a human kinetochore by specific targeting of chromatin modifiers. Dev. Cell 2008, 14, 507–522. [Google Scholar] [CrossRef]
- Bergmann, J.H.; Rodriguez, M.G.; Martins, N.M.; Kimura, H.; Kelly, D.A.; Masumoto, H.; Larionov, V.; Jansen, L.E.; Earnshaw, W.C. Epigenetic engineering shows H3K4me2 is required for HJURP targeting and CENP-A assembly on a synthetic human kinetochore. EMBO J. 2011, 30, 328–340. [Google Scholar] [CrossRef]
- Cardinale, S.; Bergmann, J.H.; Kelly, D.; Nakano, M.; Valdivia, M.M.; Kimura, H.; Masumoto, H.; Larionov, V.; Earnshaw, W.C. Hierarchical inactivation of a synthetic human kinetochore by a chromatin modifier. Mol. Biol. Cell 2009, 20, 4194–4204. [Google Scholar] [CrossRef]
- Kononenko, A.V.; Lee, N.C.; Earnshaw, W.C.; Kouprina, N.; Larionov, V. Re-engineering an alphoid(tetO)-HAC-based vector to enable high-throughput analyses of gene function. Nucleic Acids Res. 2013, 41, e107. [Google Scholar] [CrossRef]
- Murphy, T.D.; Karpen, G.H. Centromeres take flight: Alpha satellite and the quest for the human centromere. Cell 1998, 93, 317–320. [Google Scholar] [CrossRef]
- Henikoff, S. Near the edge of a chromosome’s “black hole”. Trends Genet. 2002, 18, 165–167. [Google Scholar] [CrossRef]
- Collins, F.S.; Patrinos, A.; Jordan, E.; Chakravarti, A.; Gesteland, R.; Walters, L. New goals for the U.S. Human Genome Project: 1998–2003. Science 1998, 282, 682–689. [Google Scholar] [CrossRef]
- Eichler, E.E. Repetitive conundrums of centromere structure and function. Hum. Mol. Genet. 1999, 8, 151–155. [Google Scholar] [CrossRef]
- Horvath, J.E.; Bailey, J.A.; Locke, D.P.; Eichler, E.E. Lessons from the human genome: Transitions between euchromatin and heterochromatin. Hum. Mol. Genet. 2001, 10, 2215–2223. [Google Scholar] [CrossRef]
- Horvath, J.E.; Viggiano, L.; Loftus, B.J.; Adams, M.D.; Archidiacono, N.; Rocchi, M.; Eichler, E.E. Molecular structure and evolution of an alpha satellite/non-alpha satellite junction at 16p11. Hum. Mol. Genet. 2000, 9, 113–123. [Google Scholar] [CrossRef]
- She, X.; Horvath, J.E.; Jiang, Z.; Liu, G.; Furey, T.S.; Christ, L.; Clark, R.; Graves, T.; Gulden, C.L.; Alkan, C.; et al. The structure and evolution of centromeric transition regions within the human genome. Nature 2004, 430, 857–864. [Google Scholar] [CrossRef]
- Hoskins, R.A.; Carlson, J.W.; Kennedy, C.; Acevedo, D.; Evans-Holm, M.; Frise, E.; Wan, K.H.; Park, S.; Mendez-Lago, M.; Rossi, F.; et al. Sequence finishing and mapping of Drosophila melanogaster heterochromatin. Science 2007, 316, 1625–1628. [Google Scholar] [CrossRef]
- Smith, C.D.; Shu, S.; Mungall, C.J.; Karpen, G.H. The Release 5.1 annotation of Drosophila melanogaster heterochromatin. Science 2007, 316, 1586–1591. [Google Scholar] [CrossRef]
- Kalitsis, P.; Griffiths, B.; Choo, K.H. Mouse telocentric sequences reveal a high rate of homogenization and possible role in Robertsonian translocation. Proc. Natl. Acad. Sci. USA 2006, 103, 8786–8791. [Google Scholar] [CrossRef]
- Mouse Genome Sequencing Consortium; Waterston, R.H.; Lindblad-Toh, K.; Birney, E.; Rogers, J.; Abril, J.F.; Agarwal, P.; Agarwala, R.; Ainscough, R.; Alexandersson, M.; et al. Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar] [CrossRef]
- Hayden, K.E.; Strome, E.D.; Merrett, S.E.; Lee, H.R.; Rudd, M.K.; Willard, H.F. Sequences associated with centromere competency in the human genome. Mol. Cell. Biol. 2012, 33, 763–772. [Google Scholar]
- Melters, D.P.; Bradnam, K.R.; Young, H.A.; Telis, N.; May, M.R.; Ruby, J.G.; Sebra, R.; Peluso, P.; Eid, J.; Rank, D.; et al. Comparative analysis of tandem repeats from hundreds of species reveals unique insights into centromere evolution. Genome Biol. 2013, 14, R10. [Google Scholar] [CrossRef]
- Mahtani, M.M.; Willard, H.F. Physical and genetic mapping of the human X chromosome centromere: Repression of recombination. Genome Res. 1998, 8, 100–110. [Google Scholar]
- Puechberty, J.; Laurent, A.M.; Gimenez, S.; Billault, A.; Brun-Laurent, M.E.; Calenda, A.; Marçais, B.; Prades, C.; Ioannou, P.; Yurov, Y.; et al. Genetic and physical analyses of the centromeric and pericentromeric regions of human chromosome 5: Recombination across 5cen. Genomics 1999, 56, 274–287. [Google Scholar] [CrossRef]
- Vermeesch, J.R.; Duhamel, H.; Raeymaekers, P.; van Zand, K.; Verhasselt, P.; Fryns, J.P.; Marynen, P. A physical map of the chromosome 12 centromere. Cytogenet. Genome Res. 2003, 103, 63–73. [Google Scholar] [CrossRef]
- Horvath, J.E.; Schwartz, S.; Eichler, E.E. The mosaic structure of human pericentromeric DNA: A strategy for characterizing complex regions of the human genome. Genome Res. 2000, 10, 839–852. [Google Scholar] [CrossRef]
- Bailey, J.A.; Yavor, A.M.; Massa, H.F.; Trask, B.J.; Eichler, E.E. Segmental duplications: Organization and impact within the current human genome project assembly. Genome Res. 2001, 11, 1005–1017. [Google Scholar] [CrossRef]
- Cheung, V.G.; Nowak, N.; Jang, W.; Kirsch, I.R.; Zhao, S.; Chen, X.N.; Furey, T.S.; Kim, U.J.; Kuo, W.L.; Olivier, M.; et al. Integration of cytogenetic landmarks into the draft sequence of the human genome. Nature 2001, 409, 953–958. [Google Scholar] [CrossRef]
- Genovese, G.; Handsaker, R.E.; Li, H.; Kenny, E.E.; McCarroll, S.A. Mapping the human reference genome’s missing sequence by three-way admixture in Latino genomes. Am. J. Hum. Genet. 2013, 93, 411–421. [Google Scholar] [CrossRef]
- Schueler, M.G.; Higgins, A.W.; Rudd, M.K.; Gustashaw, K.; Willard, H.F. Genomic and genetic definition of a functional human centromere. Science 2001, 294, 109–115. [Google Scholar] [CrossRef]
- Rudd, M.K.; Willard, H.F. Analysis of the centromeric regions of the human genome assembly. Trends Genet. 2004, 20, 529–533. [Google Scholar] [CrossRef]
- Nusbaum, C.; Mikkelsen, T.S.; Zody, M.C.; Asakawa, S.; Taudien, S.; Garber, M.; Kodira, C.D.; Schueler, M.G.; Shimizu, A.; Whittaker, C.A.; et al. DNA sequence and analysis of human chromosome 8. Nature 2006, 439, 331–335. [Google Scholar] [CrossRef]
- Ross, M.T.; Grafham, D.V.; Coffey, A.J.; Scherer, S.; McLay, K.; Muzny, D.; Platzer, M.; Howell, G.R.; Burrows, C.; Bird, C.P.; et al. The DNA sequence of the human X chromosome. Nature 2005, 434, 325–337. [Google Scholar] [CrossRef]
- Roizes, G. Human centromeric alphoid domains are periodically homogenized so that they vary substantially between homologues. Mechanism and implications for centromere functioning. Nucleic Acids Res. 2006, 34, 1912–1924. [Google Scholar] [CrossRef]
- Paar, V.; Pavin, N.; Rosandic, M.; Gluncic, M.; Basar, I.; Pezer, R.; Zinic, S.D. ColorHOR—Novel graphical algorithm for fast scan of alpha satellite higher-order repeats and HOR annotation for GenBank sequence of human genome. Bioinformatics 2005, 21, 846–852. [Google Scholar] [CrossRef]
- Rosandic, M.; Paar, V.; Gluncic, M.; Basar, I.; Pavin, N. Key-string algorithm—Novel approach to computational analysis of repetitive sequences in human centromeric DNA. Croat. Med. J. 2003, 44, 386–406. [Google Scholar]
- Rudd, M.K.; Schueler, M.G.; Willard, H.F. Sequence organization and functional annotation of human centromeres. Cold Spring Harb. Symp. Quant. Biol. 2003, 68, 141–149. [Google Scholar] [CrossRef]
- Alkan, C.; Ventura, M.; Archidiacono, N.; Rocchi, M.; Sahinalp, S.C.; Eichler, E.E. Organization and evolution of primate centromeric DNA from whole-genome shotgun sequence data. PLoS Comput. Biol. 2007, 3, 1807–1818. [Google Scholar]
- Alexandrov, I.A.; Mashkova, T.D.; Akopian, T.A.; Medvedev, L.I.; Kisselev, L.L.; Mitkevich, S.P.; Yurov, Y.B. Chromosome-specific alpha satellites: Two distinct families on human chromosome 18. Genomics 1991, 11, 15–23. [Google Scholar] [CrossRef]
- Choo, K.H.; Earle, E.; Vissel, B.; Filby, R.G. Identification of two distinct subfamilies of alpha satellite DNA that are highly specific for human chromosome 15. Genomics 1990, 7, 143–151. [Google Scholar] [CrossRef]
- Wevrick, R.; Willard, H.F. Physical map of the centromeric region of human chromosome 7: Relationship between two distinct alpha satellite arrays. Nucleic Acids Res. 1991, 19, 2295–2301. [Google Scholar] [CrossRef]
- Miga, K.H.; Newton, Y.; Jain, M.; Altemose, N.; Willard, H.F.; Kent, W.J. Centromere reference models for human chromosomes X and Y satellite arrays. arXiv 2013. arXiv:1307.0035v3[q-bio.GN]. [Google Scholar]
- Schulze, S.; Sinclair, D.A.; Silva, E.; Fitzpatrick, K.A.; Singh, M.; Lloyd, V.K.; Morin, K.A.; Kim, J.; Holm, D.G.; Kennison, J.A.; et al. Essential genes in proximal 3L heterochromatin of Drosophila melanogaster. Mol. Gen. Genet. 2001, 264, 782–789. [Google Scholar] [CrossRef]
- Ryan, D.P.; da Silva, M.R.; Soong, T.W.; Fontaine, B.; Donaldson, M.R.; Kung, A.W.; Jongjaroenprasert, W.; Liang, M.C.; Khoo, D.H.; Cheah, J.S.; et al. Mutations in potassium channel Kir2.6 cause susceptibility to thyrotoxic hypokalemic periodic paralysis. Cell 2010, 140, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Masumoto, H.; Nakano, M.; Ohzeki, J. The role of CENP-B and alpha-satellite DNA: De novo assembly and epigenetic maintenance of human centromeres. Chromosome Res. 2004, 12, 543–556. [Google Scholar] [CrossRef]
- Blower, M.D.; Sullivan, B.A.; Karpen, G.H. Conserved organization of centromeric chromatin in flies and humans. Dev. Cell 2002, 2, 319–330. [Google Scholar] [CrossRef]
- Sullivan, B.A.; Karpen, G.H. Centromeric chromatin exhibits a histone modification pattern that is distinct from both euchromatin and heterochromatin. Nat. Struct. Mol. Biol. 2004, 11, 1076–1083. [Google Scholar] [CrossRef]
- Hori, T.; Fukagawa, T. Establishment of the vertebrate kinetochores. Chromosome Res. 2012, 20, 547–561. [Google Scholar] [CrossRef]
- Spence, J.M.; Critcher, R.; Ebersole, T.A.; Valdivia, M.M.; Earnshaw, W.C.; Fukagawa, T.; Farr, C.J. Co-localization of centromere activity, proteins and topoisomerase II within a subdomain of the major human X alpha-satellite array. EMBO J. 2002, 21, 5269–5280. [Google Scholar] [CrossRef]
- Sullivan, L.L.; Boivin, C.D.; Mravinac, B.; Song, I.Y.; Sullivan, B.A. Genomic size of CENP-A domain is proportional to total alpha satellite array size at human centromeres and expands in cancer cells. Chromosome Res. 2011, 19, 457–470. [Google Scholar] [CrossRef]
- Hasson, D.; Panchenko, T.; Salimian, K.J.; Salman, M.U.; Sekulic, N.; Alonso, A.; Warburton, P.E.; Black, B.E. The octamer is the major form of CENP-A nucleosomes at human centromeres. Nat. Struct. Mol. Biol. 2013, 20, 687–695. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Aldrup-MacDonald, M.E.; Sullivan, B.A. The Past, Present, and Future of Human Centromere Genomics. Genes 2014, 5, 33-50. https://doi.org/10.3390/genes5010033
Aldrup-MacDonald ME, Sullivan BA. The Past, Present, and Future of Human Centromere Genomics. Genes. 2014; 5(1):33-50. https://doi.org/10.3390/genes5010033
Chicago/Turabian StyleAldrup-MacDonald, Megan E., and Beth A. Sullivan. 2014. "The Past, Present, and Future of Human Centromere Genomics" Genes 5, no. 1: 33-50. https://doi.org/10.3390/genes5010033