
How Porin Heterogeneity and Trade-Offs Affect the Antibiotic Susceptibility of Gram-Negative Bacteria

Abstract
:1. Introduction

2. Modifications Affecting Porins Influencing Antibiotic Susceptibility
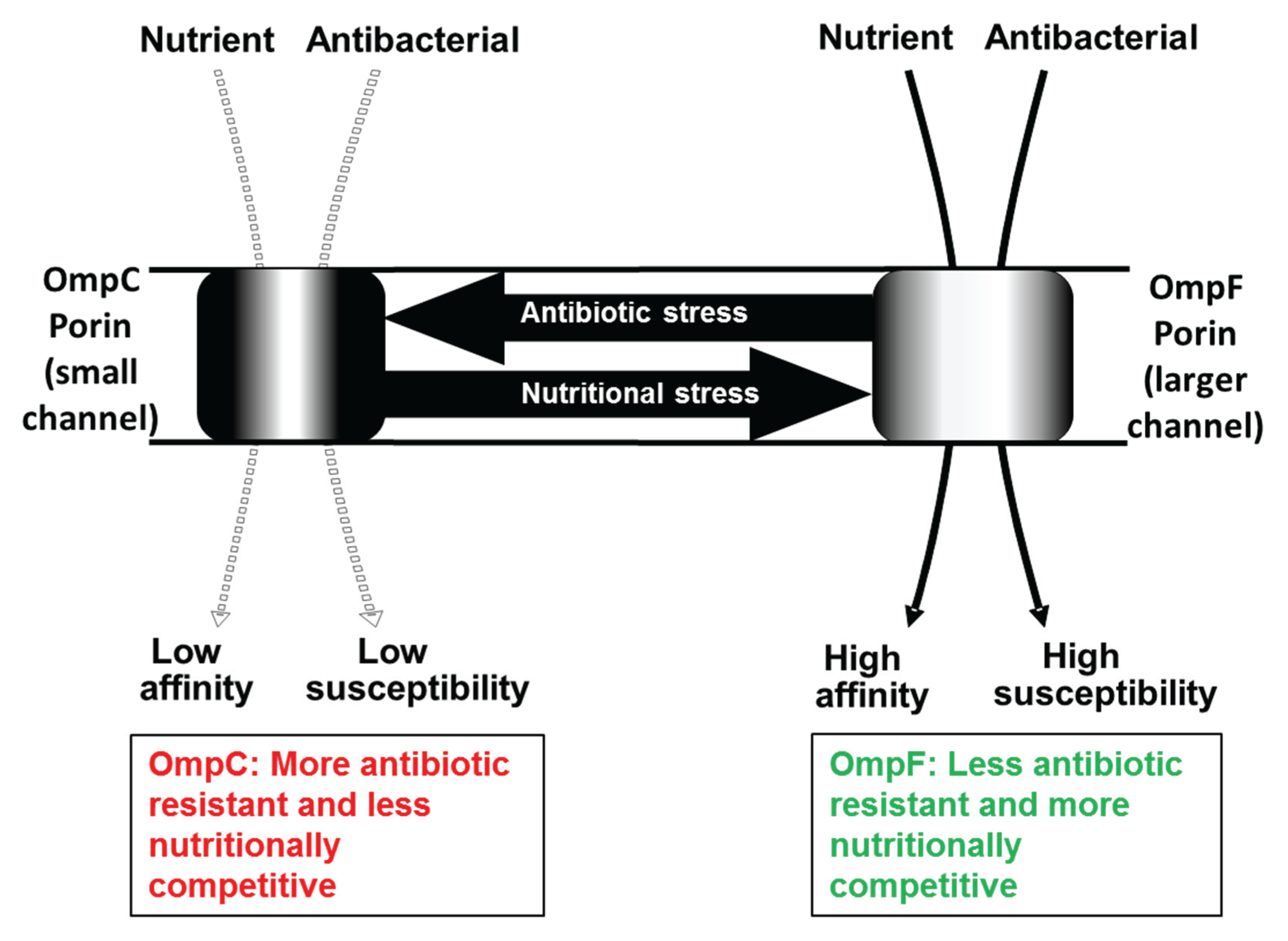
2.1. Regulatory Changes
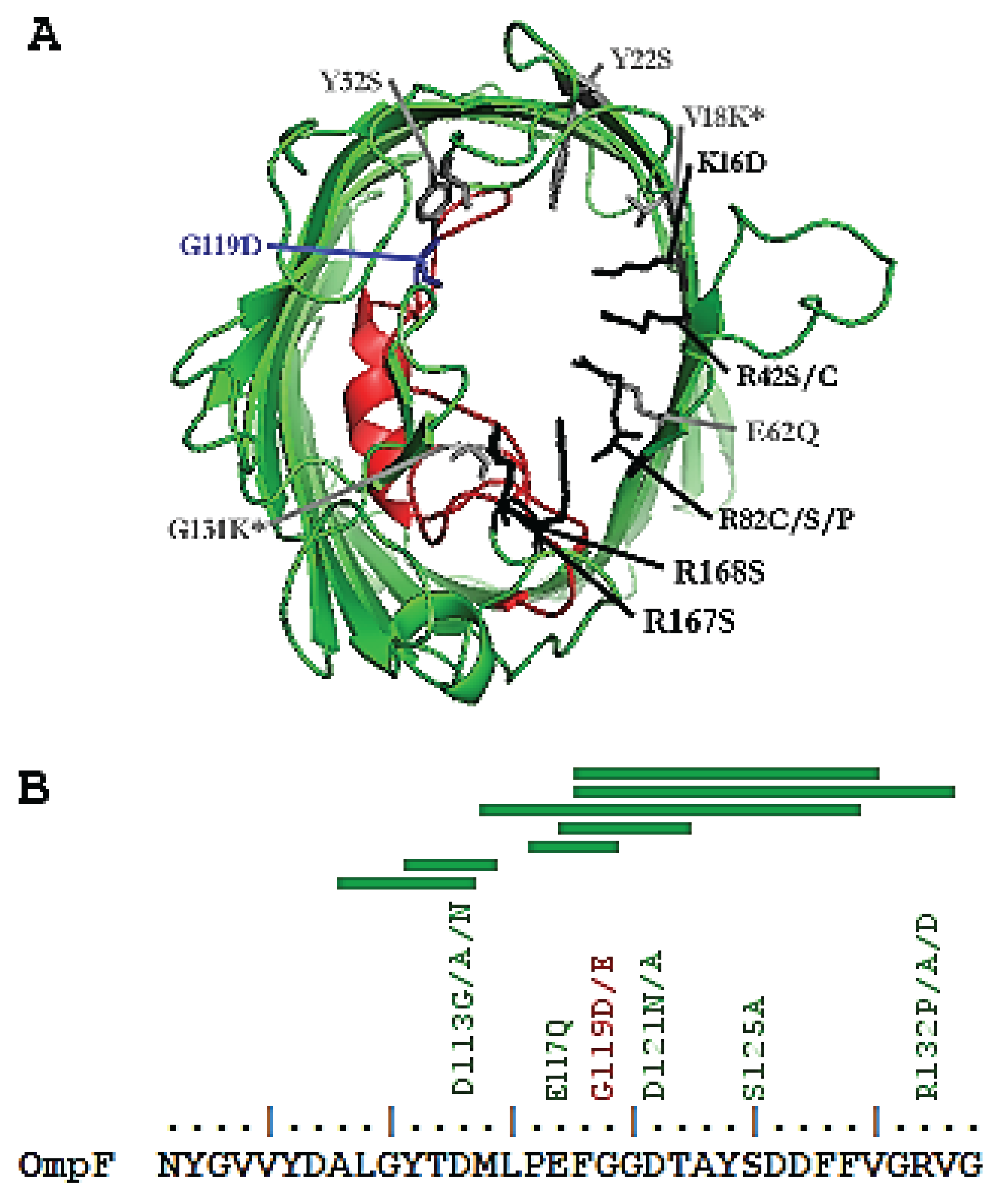
2.2. Structural Mutations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OmpF Sequence Changes | Decreasing (D) or Increasing (I) Permeability | References |
|---|---|---|
| del116–119 | I | [34] |
| del117–122 | I | [34] |
| del108–114 | I | [35,36] |
| del111–123 | I | [37] |
| del114–129 | I | [38] |
| del118–130 | I | [39] |
| del118–133 | I | [38] |
| Y22S | D | [40] |
| D121N/A | I | [40] |
| D113G/A/N | I | [33,34,36] |
| R132A/D/P | I | [33,35,41] |
| G119D/E | D | [42] |
| S125A | I | [40] |
| E117Q | I | [40] |
| R167S | I | [40] |
| R168S | I | [40] |
| R42S/C | I | [33,36,40] |
| K16D | I | [41] |
| R82C/S/P | I | [33,36,41] |
| E62Q | I | [40] |
| Y32S | I | [40] |
| V18K + G131K | D | [35] |

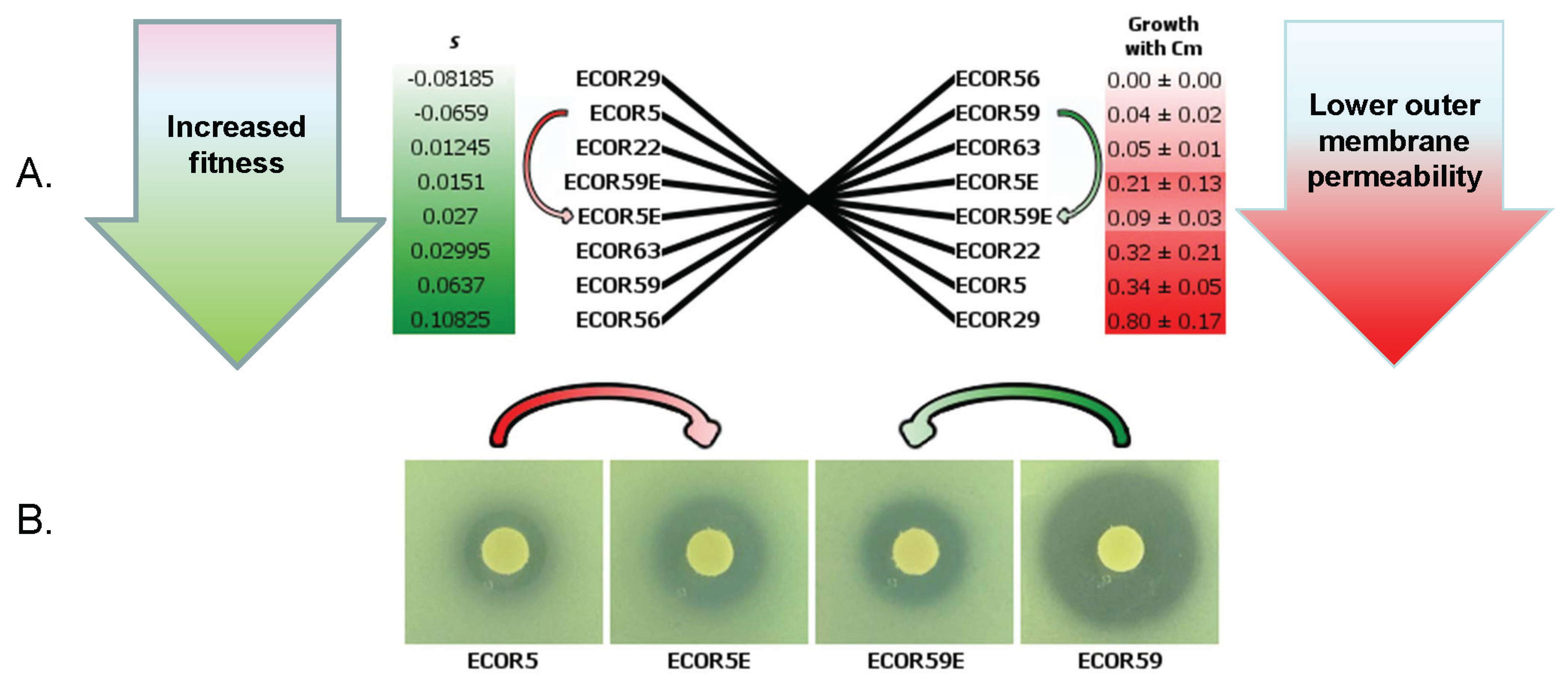
2.3. Mutations Affecting Porins Selected under Nutrient Limitation Stress
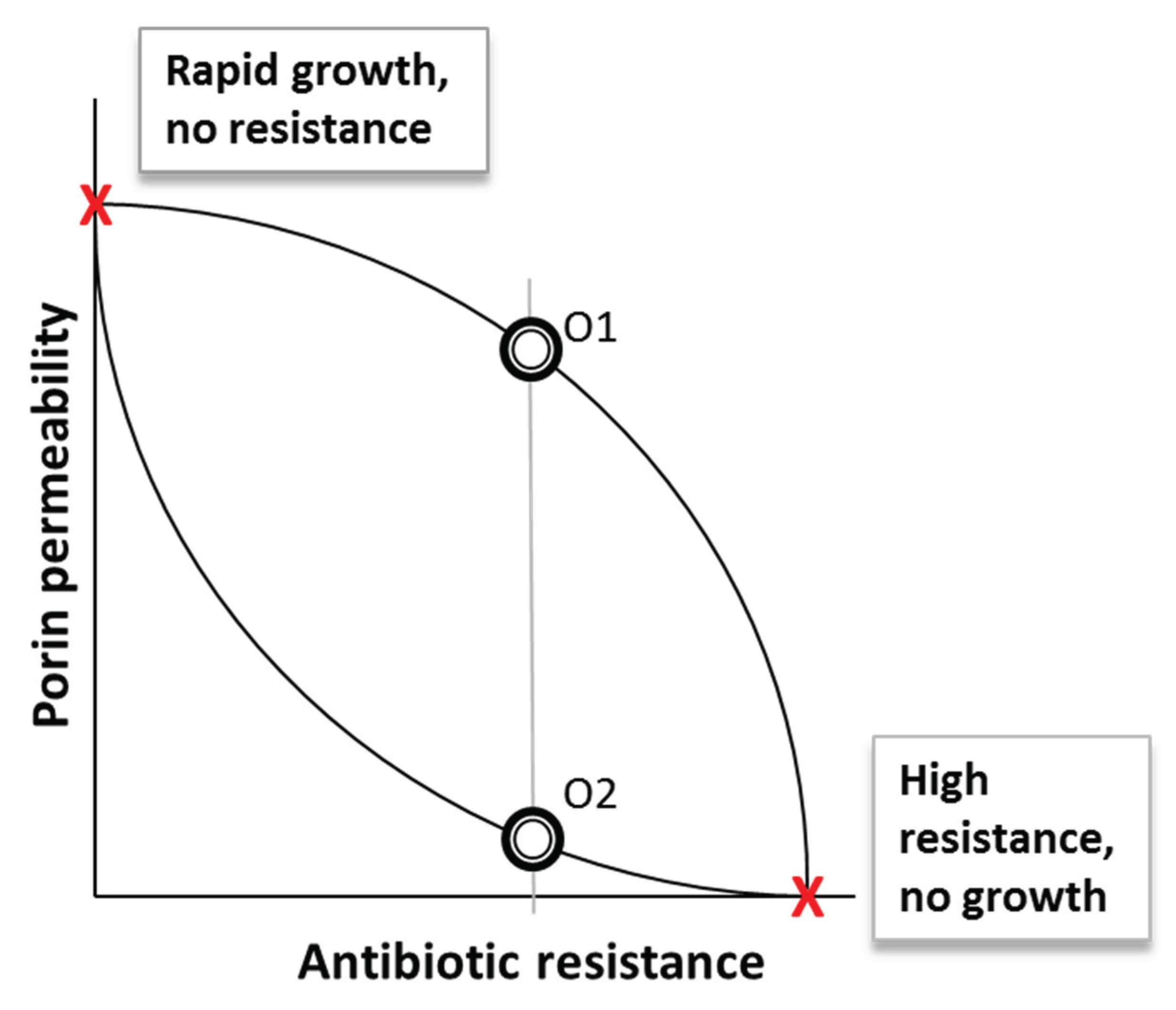
3. Porins as Perfect Examples of a Design Constraint Trade-Off

4. Bacterial Heterogeneity in Porins as a Solution to Alternating Environments

5. Conclusions on the Role of Porins in Antibiotic Resistance
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Pagès, J.-M. Structure, function and regulation of outer membrane proteins involved in drug transport in Enterobactericeae: The OmpF/C—TolC case. Open Microbiol. J. 2013, 7, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. Biophys. Acta 2009, 1794, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Q.; Ferenci, T. Regulation of porin-mediated outer membrane permeability by nutrient limitation in Escherichia coli. J. Bacteriol. 1998, 180, 3917–3922. [Google Scholar] [PubMed]
- Death, A.; Notley, L.; Ferenci, T. Derepression of LamB protein facilitates outer membrane permeation of carbohydrates into Escherichia coli under conditions of nutrient stress. J. Bacteriol. 1993, 175, 1475–1483. [Google Scholar] [PubMed]
- Ochs, M.M.; McCusker, M.P.; Bains, M.; Hancock, R.E.W. Negative regulation of the Pseudomonas aeruginosa outer membrane porin OprD selective for imipenem and basic amino acids. Antimicrob. Agents Chemother. 1999, 43, 1085–1090. [Google Scholar] [PubMed]
- Wylie, J.L.; Worobec, E.A. Cloning and nucleotide sequence of the Pseudomonas aeruginosa glucose-selective OprB porin gene and distribution of OprB within the family Pseudomonadaceae. Eur. J. Biochem. 1994, 220, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Pratt, L.A.; Hsing, W.H.; Gibson, K.E.; Silhavy, T.J. From acids to OsmZ -multiple factors influence the synthesis of the OmpF and OmpC porins in Escherichia coli. Mol. Microbiol. 1996, 20, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Delcour, A.H. Solute uptake through general porins. Front. Biosci. 2003, 8, D1055–D1071. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Q.; Ferenci, T. An analysis of multifactorial influences on the transcriptional control of ompF and ompC porin expression under nutrient limitation. Microbiology 2001, 147, 2981–2989. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz, M.A.; Calva, E. The complexities of porin genetic regulation. J. Mol. Microbiol. Biotechnol. 2010, 18, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.P.; McMurry, L.M.; Levy, S.B. MarA locus causes decreased expression of ompF porin in multiple-antibiotic-resistant (mar) mutants of Escherichia coli. J. Bacteriol. 1988, 170, 5416–5422. [Google Scholar] [PubMed]
- Ferenci, T. Maintaining a healthy SPANC balance through regulatory and mutational adaptation. Mol. Microbiol. 2005, 57, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Vasoo, S.; Barreto, J.N.; Tosh, P.K. Emerging issues in gram-negative bacterial resistance: An update for the practicing clinician. Mayo Clinic Proc. 2015, 90, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Davin-Regli, A. Enterobacter aerogenes and Enterobacter cloacae, versatile bacterial pathogens confronting antibiotic treatment. Front. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unemo, M.; Golparian, D.; Nicholas, R.; Ohnishi, M.; Gallay, A.; Sednaoui, P. High-level cefixime- and ceftriaxone-resistant Neisseria gonorrhoeae in France: Novel penA mosaic allele in a successful international clone causes treatment failure. Antimicrob. Agents Chemother. 2012, 56, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.J.; Inouye, M. EnvZ-OmpR interaction and osmoregulation in Escherichia coli. J. Biol. Chem. 2002, 277, 24155–24161. [Google Scholar] [CrossRef] [PubMed]
- Tangden, T.; Adler, M.; Cars, O.; Sandegren, L.; Lowdin, E. Frequent emergence of porin-deficient subpopulations with reduced carbapenem susceptibility in ESBL-producing Escherichia coli during exposure to ertapenem in an in vitro pharmacokinetic model. J. Antimicrob. Chemother. 2013, 68, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Doumith, M.; Ellington, M.J.; Livermore, D.M.; Woodford, N. Molecular mechanisms disrupting porin expression in ertapenem-resistant Klebsiella and Enterobacter spp. Clinical isolates from the UK. J. Antimicrob. Chemother. 2009, 63, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.M.; Li, H.; Wang, C.; Peng, X.X. Proteomic analysis of nalidixic acid resistance in Escherichia coli: Identification and functional characterization of OM proteins. J. Proteome Res. 2008, 7, 2399–2405. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Berg, O.G.; Roth, J.R.; Andersson, D.I. Contribution of gene amplification to evolution of increased antibiotic resistance in Salmonella typhimurium. Genetics 2009, 182, 1183–1195. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, R.A.; Szabo, D. Mechanisms of multidrug resistance in Acinetobacter species and Pseudomonas aeruginosa. Clin. Infect. Dis. 2006, 43, S49–S56. [Google Scholar] [CrossRef] [PubMed]
- Prohinar, P.; Forst, S.A.; Reed, D.; Mandic-Mulec, I.; Weiss, J. OmpR-dependent and OmpR-independent responses of Escherichia coli to sublethal attack by the neutrophil bactericidal/permeability increasing protein. Mol. Microbiol. 2002, 43, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, M.L.; Sanna, M.G.; Fontaine, A.; Sansonetti, P.J. OmpC is involved in invasion of epithelial cells by Shigella flexneri. Infect. Immun. 1993, 61, 3625–3635. [Google Scholar] [PubMed]
- Dorman, C.J.; Chatfield, S.; Higgins, C.F.; Hayward, C.; Dougan, G. Characterization of porin and OmpR mutants of a virulent strain of Salmonella typhimurium: OmpR mutants are attenuated in vivo. Infect. Immun. 1989, 57, 2136–2140. [Google Scholar] [PubMed]
- Cohen, S.P.; McMurry, L.M.; Hooper, D.C.; Wolfson, J.S.; Levy, S.B. Cross-resistance to fluoroquinolones in multiple-antibiotic-resistant (mar) Escherichia coli selected by tetracycline or chloramphenicol: Decreased drug accumulation associated with membrane changes in addition to OmpF reduction. Antimicrob. Agents Chemother. 1989, 33, 1318–1325. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.K.; Fung, C.P.; Lin, J.C.; Chen, J.H.; Chang, F.Y.; Chen, T.L.; Siu, L.K. Klebsiella pneumoniae outer membrane porins OmpK35 and OmpK36 play roles in both antimicrobial resistance and virulence. Antimicrob. Agents Chemother. 2011, 55, 1485–1493. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Ginn, A.N.; Wiklendt, A.M.; Ellem, J.; Wong, J.S.J.; Ingram, P.; Guy, S.; Garner, S.; Iredell, J.R. Emergence of blaKPC carbapenemase genes in Australia. Int. J. Antimicrob. Agents 2015, 45, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Tavio, M.M.; Vila, J.; Perilli, M.; Casansa, L.T.; Macia, L.; Amicosante, G.; de Anta, M.T.J. Enhanced active efflux, repression of porin synthesis and development of Mar phenotype by diazepam in two enterobacteria strains. J. Med. Microbiol. 2004, 53, 1119–1122. [Google Scholar] [CrossRef] [PubMed]
- Mallea, M.; Chevalier, J.; Bornet, C.; Eyraud, A.; Davinregli, A.; Bollet, C.; Pages, J.M. Porin alteration and active efflux—Two in vivo drug resistance strategies used by Enterobacter aerogenes. Microbiology 1998, 144, 3003–3009. [Google Scholar] [CrossRef] [PubMed]
- Basle, A.; Rummel, G.; Storici, P.; Rosenbusch, J.P.; Schirmer, T. Crystal structure of osmoporin OmpC from Escherichia coli at 2.0 Angstrom. J. Mol. Biol. 2006, 362, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Cowan, S.W.; Schirmer, T.; Rummel, G.; Steiert, M.; Ghosh, R.; Pauptit, R.A.; Jansonius, J.N.; Rosenbusch, J.P. Crystal structures explain functional properties of two E. coli porins. Nature 1992, 358, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Saint, N.; Lou, K.L.; Widmer, C.; Luckey, M.; Schirmer, T.; Rosenbusch, J.P. Structural and functional characterization of OmpF porin mutants selected for larger pore size. 2. Functional characterization. J. Biol. Chem. 1996, 271, 20676–20680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Ferenci, T. OmpF changes and the complexity of Echerichia coli adaptation to prolonged lactose limitation. FEMS Microbiol. Lett. 1999, 176, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Phale, P.S.; Philippsen, A.; Widmer, C.; Phale, V.P.; Rosenbusch, J.P.; Schirmer, T. Role of charged residues at the OmpF porin channel constriction probed by mutagenesis and simulation. Biochemistry 2001, 40, 6319–6325. [Google Scholar] [CrossRef] [PubMed]
- Benson, S.A.; Occi, J.L.; Sampson, B.A. Mutations that alter the pore function of the OmpF porin of Escherichia coli K12. J. Mol. Biol. 1988, 203, 961–970. [Google Scholar] [CrossRef]
- Sampson, B.A.; Misra, R.; Benson, S.A. Identification and characterization of a new gene of Escherichia coli K-12 involved in outer membrane permeability. Genetics 1989, 122, 491–501. [Google Scholar] [PubMed]
- Klebba, P.E.; Benson, S.A.; Bala, S.; Abdullah, T.; Reid, J.; Singh, S.P.; Nikaido, H. Determinants of OmpF porin antigenicity and structure. J. Biol. Chem. 1990, 265, 6800–6810. [Google Scholar] [PubMed]
- Benson, S.A.; DeCloux, A.M.; Munro, J. Mutant bias in nonlethal selections results from selective recovery of mutants. Genetics 1991, 129, 647–658. [Google Scholar] [PubMed]
- Ziervogel, B.K.; Roux, B. The binding of antibiotics in OmpF porin. Structure 2013, 21, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Bredin, J.; Saint, N.; Malléa, M.; Emmanuelle, D.; Molle, G.; Pages, J.-M.; Simonet, V. Alteration of pore properties of Escherichia coli OmpF induced by mutation of key residues in anti-loop 3 region. Biochem. J. 2002, 363, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Jeanteur, D.; Schirmer, T.; Fourel, D.; Simonet, V.; Rummel, G.; Widmer, C.; Rosenbusch, J.P.; Pattus, F.; Pages, J.M. Structural and functional alterations of a colicin-resistant mutant of OmpF porin from Escherichia coli. Proc. Natl. Acad. Sci. USA 1994, 91, 10675–10679. [Google Scholar] [CrossRef] [PubMed]
- Benz, R. Structure and function of porins from gram-negative bacteria. Annu. Rev. Microbiol. 1988, 42, 359–393. [Google Scholar] [CrossRef] [PubMed]
- Ferenci, T. Bacterial physiology, regulation and mutational adaptation in a chemostat environment. Adv. Microb. Physiol. 2008, 53, 169–229. [Google Scholar] [PubMed]
- Levins, R. Evolution in Changing Environments, Some Theoretical Exploration; Princeton University Press: Princeton, NJ, USA, 1968. [Google Scholar]
- Stearns, S.C. Trade-offs in life-history evolution. Funct. Ecol. 1989, 3, 259–268. [Google Scholar] [CrossRef]
- Maharjan, R.; Nilsson, S.; Sung, J.; Haynes, K.; Beardmore, R.E.; Hurst, L.D.; Ferenci, T.; Gudelj, I. The form of a trade-off determines the response to competition. Ecol. Lett. 2013, 16, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.R.; Gudelj, I.; Beardmore, R. Biophysical mechanisms that maintain biodiversity through trade-offs. Nat. Commun. 2015, 6, 6278–6278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard, B.; Garcia, J.S.; Gouriou, S.; Duriez, P.; Brahimi, N.; Bingen, E.; Denamur, E. The link between phylogeny and virulence in Escherichia coli extraintestinal infection. Infect. Immun. 1999, 67, 546–553. [Google Scholar] [PubMed]
- Phan, K.; Ferenci, T. A design-constraint trade-off underpins the diversity in ecologically important traits in species Escherichia coli. ISME J. 2013, 7, 2034–2043. [Google Scholar] [CrossRef] [PubMed]
- Donaldson-Matasci, M.C.; Lachmann, M.; Bergstrom, C.T. Phenotypic diversity as an adaptation to environmental uncertainty. Evol. Ecol. Res. 2008, 10, 493–515. [Google Scholar]
- Day, T.; Huijben, S.; Read, A.F. Is selection relevant in the evolutionary emergence of drug resistance? Trends Microbiol. 2015, 23, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Schwan, W.R. Survival of uropathogenic Escherichia coli in the murine urinary tract is dependent on OmpR. Microbiol. Sgm 2009, 155, 1832–1839. [Google Scholar] [CrossRef] [PubMed]
- Toprak, E.; Veres, A.; Michel, J.B.; Chait, R.; Hartl, D.L.; Kishony, R. Evolutionary paths to antibiotic resistance under dynamically sustained drug selection. Nat. Genet. 2012, 44, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Munck, C.; Gumpert, H.K.; Wallin, A.I.N.; Wang, H.H.; Sommer, M.O.A. Prediction of resistance development against drug combinations by collateral responses to component drugs. Sci. Transl. Med. 2014. [Google Scholar] [CrossRef] [PubMed]
- Hajjar, E.; Bessonov, A.; Molitor, A.; Kumar, A.; Mahendran, K.R.; Winterhalter, M.; Pages, J.M.; Ruggerone, P.; Ceccarelli, M. Toward screening for antibiotics with enhanced permeation properties through bacterial porins. Biochemistry 2010, 49, 6928–6935. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferenci, T.; Phan, K. How Porin Heterogeneity and Trade-Offs Affect the Antibiotic Susceptibility of Gram-Negative Bacteria. Genes 2015, 6, 1113-1124. https://doi.org/10.3390/genes6041113
Ferenci T, Phan K. How Porin Heterogeneity and Trade-Offs Affect the Antibiotic Susceptibility of Gram-Negative Bacteria. Genes. 2015; 6(4):1113-1124. https://doi.org/10.3390/genes6041113
Chicago/Turabian StyleFerenci, Thomas, and Katherine Phan. 2015. "How Porin Heterogeneity and Trade-Offs Affect the Antibiotic Susceptibility of Gram-Negative Bacteria" Genes 6, no. 4: 1113-1124. https://doi.org/10.3390/genes6041113
APA StyleFerenci, T., & Phan, K. (2015). How Porin Heterogeneity and Trade-Offs Affect the Antibiotic Susceptibility of Gram-Negative Bacteria. Genes, 6(4), 1113-1124. https://doi.org/10.3390/genes6041113