
Genome-Wide Identification, Characterization and Expression Profiling of ADF Family Genes in Solanum lycopersicum L.


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Analysis of ADF Family Genes
2.2. Analysis of Gene Duplication and Chromosomal Localization
2.3. Preparation of Plant Materials and Sample Collection
2.4. RNA Extraction and cDNA Synthesis
2.5. qPCR Expression Analysis
3. Results
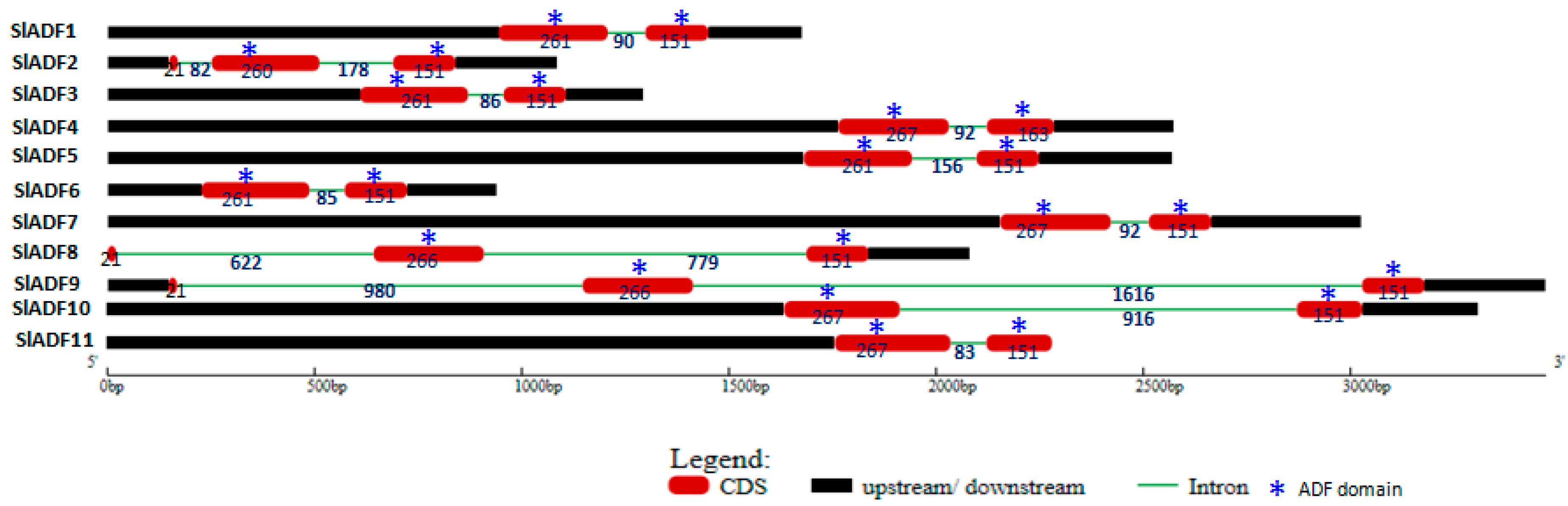
3.1. Sequence Analysis and Genomic Organization of Tomato ADF Genes and Corresponding Proteins
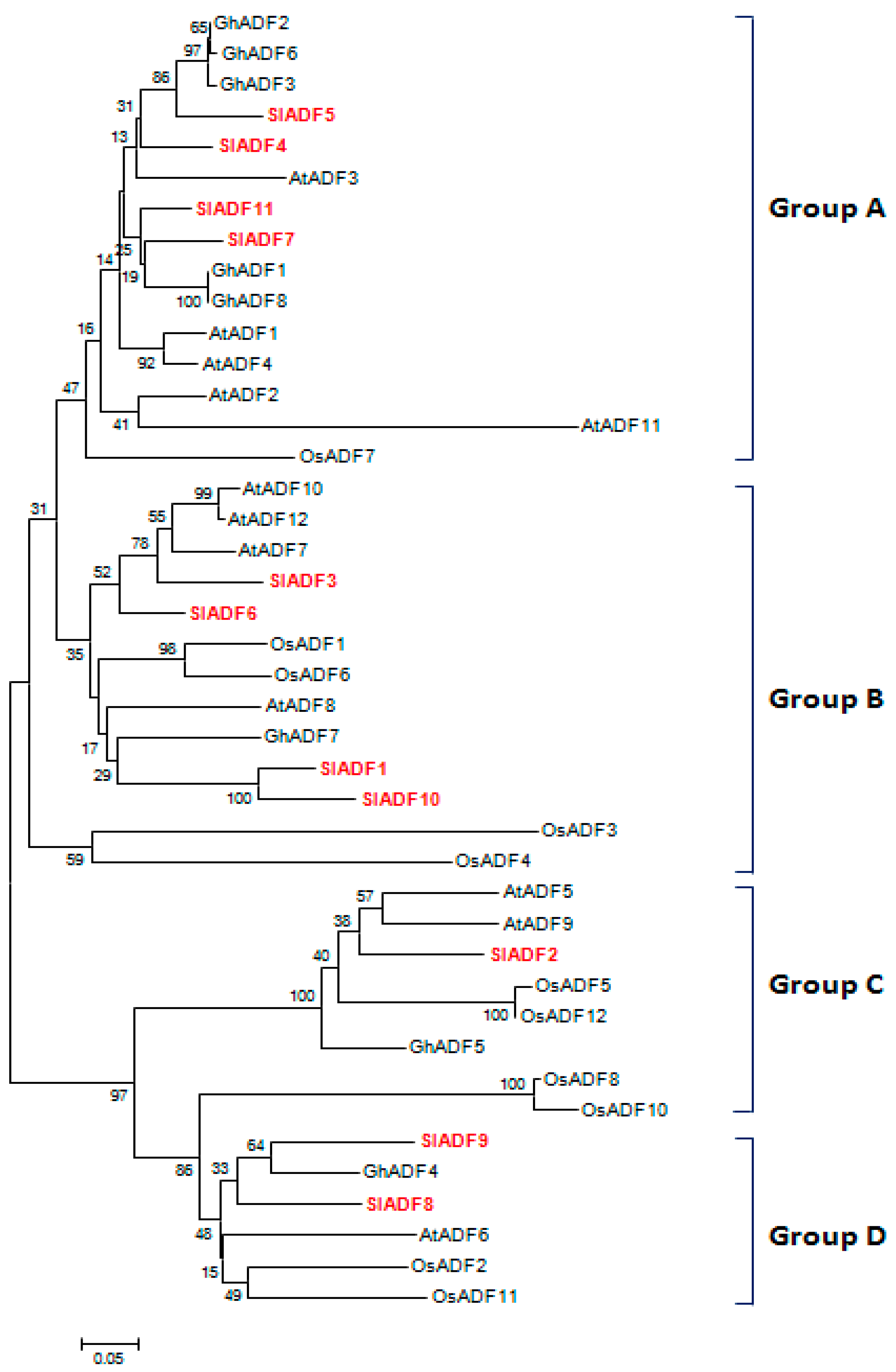
3.2. Phylogenetic Analysis of SlADF Proteins
3.3. Chromosomal Position and Duplication of Tomato ADF Genes
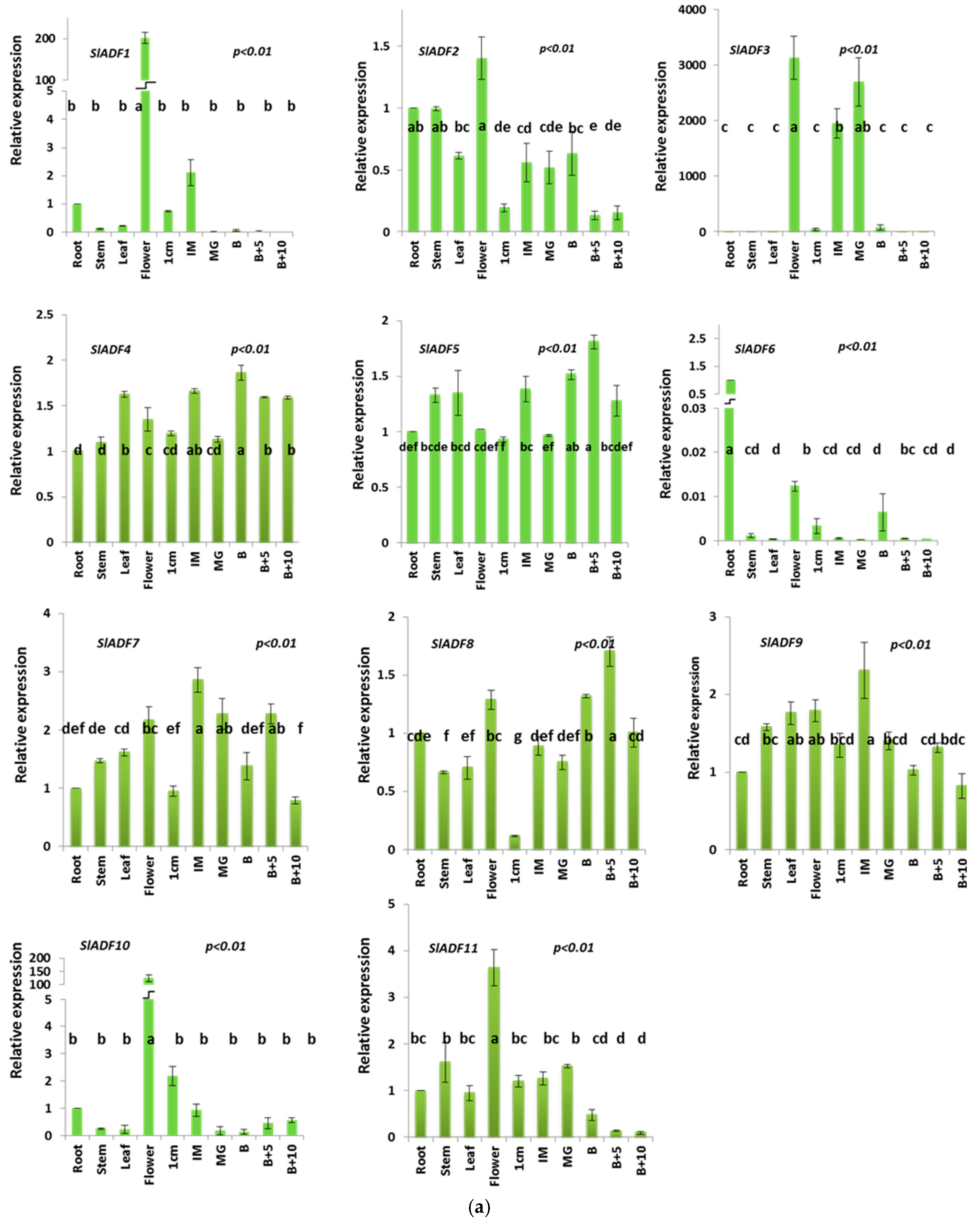
3.4. Expression Analysis of Tomato ADF Genes in Different Organs
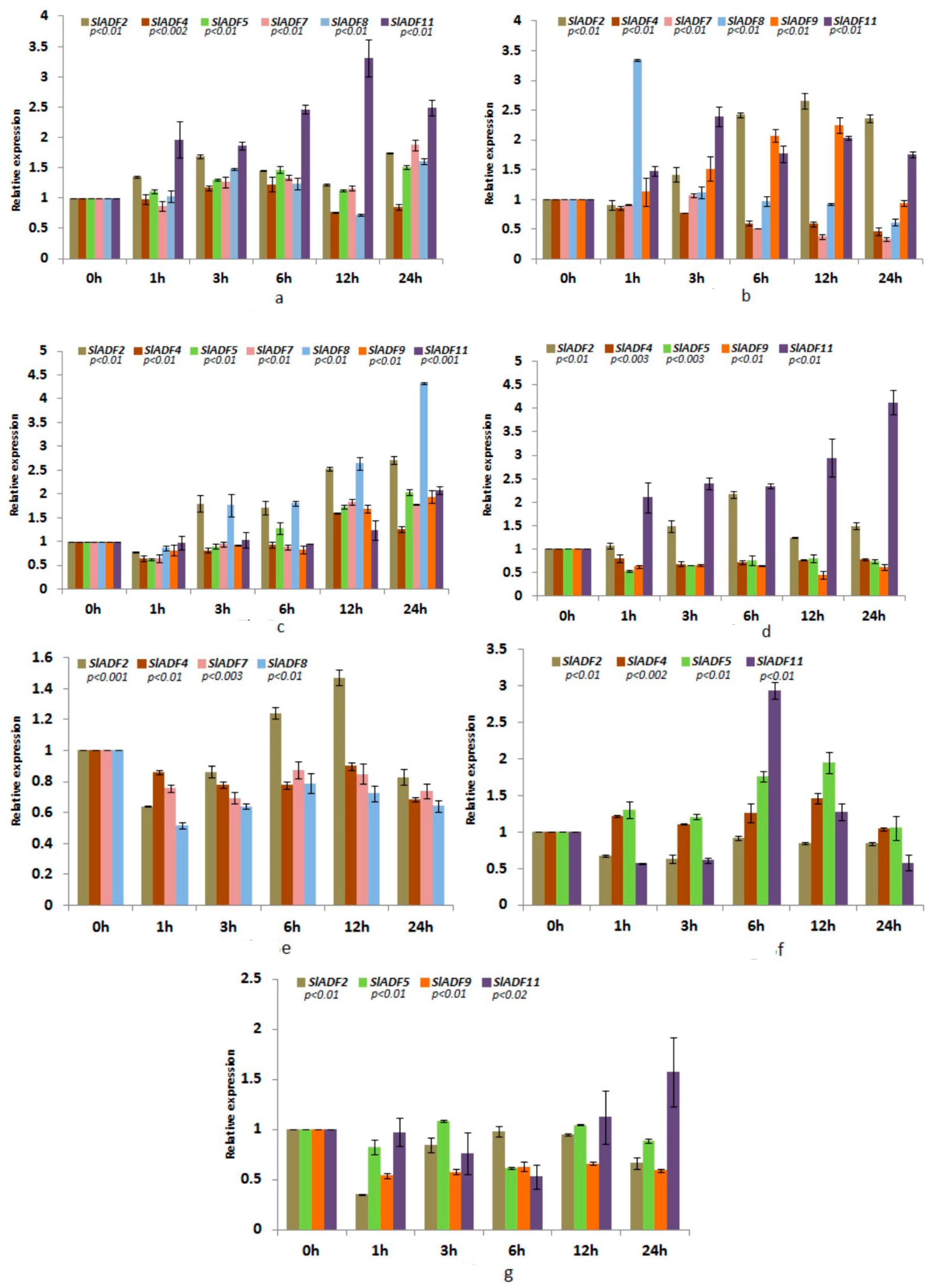
3.5. Expression Analysis of Tomato ADF Genes under Different Abiotic Stresses
3.5.1. Cold Stress
3.5.2. Heat Stress
3.5.3. Drought Stress
3.5.4. NaCl Stress
3.5.5. ABA Treatment
3.5.6. JA Treatment
3.5.7. Wounding
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ouellet, F.; Carpentier, E.; Cope, M.J.T.V.; Monroy, A.F.; Sarhan, F. Regulation of a wheat actin-depolymerizing factor during cold acclimation. Plant Physiol. 2001, 125, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Hepler, P.K.; Vidali, L.; Cheung, A.Y. Polarized cell growth in higher plants. Annu. Rev. Cell Dev. Biol. 2001, 17, 159–187. [Google Scholar] [CrossRef] [PubMed]
- Drobak, B.K.; Franklin-Tong, V.E.; Staiger, C.T. The role of the actin cytoskeleton in plant cell signaling. New Phytol. 2004, 163, 13–30. [Google Scholar] [CrossRef]
- Bamburg, J.R.; Harris, H.E.; Weeds, A.G. Partial purification and characterization of an actin depolymerizing factor from brain. FEBS Lett. 1980, 121, 178–182. [Google Scholar] [CrossRef]
- Chi, J.; Wang, X.; Zhou, H.; Zhang, G.; Sun, Y.; Li, Z.; Ma, Z. Molecular cloning and characterization of the actin-depolymerizing factor gene in Gossypium barbadense. Genes Genet. Syst. 2008, 83, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Maciver, S.K.; Hussey, P.J. The ADF/cofilin family: Actin-remodeling proteins. Genome Biol. 2002. [Google Scholar] [CrossRef]
- Ruzicka, D.R.; Kandasamy, M.K.; McKinney, E.C.; Burgos-Rivera, B.; Meagher, R.B. The ancient subclasses of Arabidopsis Actin Depolymerizing Factor genes exhibit novel and differential expression. Plant J. 2007, 52, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Liu, Q.; Xue, Q. Comparative study of rice and Arabidopsis actin-depolymerizing factors gene families. J. Plant Physiol. 2006, 163, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Poukkula, M.; Kremneva, E.; Serlachius, M.; Lappalainen, P. Actin-depolymerizing factor homology domain: A conserved fold performing diverse roles in cytoskeletal dynamics. Cytoskeleton 2011, 68, 471–490. [Google Scholar] [CrossRef] [PubMed]
- Allwood, E.G.; Smertenko, A.P.; Hussey, P.J. Phosphorylation of plant actin-depolymerising factor by calmodulin-like domain protein kinase. FEBS Lett. 2001, 499, 97–100. [Google Scholar] [CrossRef]
- Augustine, R.C.; Vidali, L.; Kleinman, K.P.; Bezanilla, M. Actin depolymerizing factor is essential for viability in plants, and its phosphoregulation is important for tip growth. Plant J. 2008, 54, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Maciver, S.K. How ADF/cofilin depolymerizes actin filaments: Commentary. Curr. Opin. Cell Biol. 1998, 10, 140–144. [Google Scholar] [CrossRef]
- Dong, C.H.; Xia, G.X.; Hong, Y.; Ramachandran, S.; Kost, B.; Chua, N.H. ADF proteins are involved in the control of flowering and regulate F-actin organization, cell expansion, and organ growth in Arabidopsis. Plant Cell 2001, 13, 1333–1346. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Schiefelbein, J. Cloning and characterization of an actin depolymerizing factor gene from grape (Vitis vinifera L.) expressed during rooting in stem cuttings. Plant Sci. 2002, 162, 283–288. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chung, A.Y.; Wu, H. Actin-depolymerizing factor mediates Rac/Rop GTPase-regulated pollen tube growth. Plant Cell 2003, 15, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Lopez, I.; Anthony, R.G.; Maciver, S.K.; Jiang, C.J.; Khan, S.; Weeds, A.G.; Hussey, P.J. Pollen specific expression of maize genes encoding actin depolymerizing factor-like proteins. Proc. Natl. Acad. Sci. USA 1996, 93, 7415–7420. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Liu, X.; Chen, C.; Tian, W.; Xia, Z.; Li, D. Molecular cloning and characterization of an actin-depolymerizing factor gene in Hevea brasiliensis. Afr. J. Biotechnol. 2010, 9, 7603–7610. [Google Scholar]
- Clément, M.; Ketelaar, T.; Rodiuc, N.; Banora, M.Y.; Smertenko, A.; Engler, G.; Abad, P.; Hussey, P.J.; de Almeida Engler, J. Actin-depolymerizing factor2-mediated actin dynamics are essential for root-knot nematode infection of Arabidopsis. Plant Cell 2009, 21, 2963–2979. [Google Scholar] [CrossRef] [PubMed]
- Miklis, M.; Consonni, C.; Bhat, R.A.; Lipka, V.; Schulze-Lefert, P.; Panstruga, R. Barley MLO modulates actin-dependent and actin-independent antifungal defense pathways at the cell periphery. Plant Physiol. 2007, 144, 1132–1143. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Chaudhry, F.; Ruzicka, D.R.; Meagher, R.B.; Staiger, C.J.; Day, B. Arabidopsis actin-depolymerizing factor AtADF4 mediates defense signal transduction triggered by the Pseudomonas syringae effector AvrPphB. Plant Physiol. 2009, 150, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Porter, K.; Shimono, M.; Tian, M.; Day, B. Arabidopsis actin-depolymerizing factor-4 links pathogen perception, defense activation and transcription to cytoskeletal dynamics. PLoS Pathog. 2012, 8, e1003006. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Lv, W.; Jiang, S.; Zhang, D.; Cai, G.; Pan, J.; Li, D. Genome-wide identification and expression analysis of calcium-dependent protein kinase in maize. BMC Genom. 2013. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Yano, K.; Suzuki, A.; Kawamura, S.; Sakurai, N.; Suda, K.; Ooga, K. Large-scale analysis of full length cDNAs from the tomato (Solanum lycopersicum) cultivar Micro-Tom, a reference system for the Solanaceae genomics. BMC Genom. 2010, 11, 210. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, H.; Fukui, Y.; Yahara, I. Live dynamics of Dictyostelium cofilin suggests a role in remodeling actin lattice work into bundles. J. Cell Sci. 1997, 110, 2333–2344. [Google Scholar] [PubMed]
- Agnew, B.J.; Minamide, L.S.; Bamburg, J.R. Reactivation of phosphorylated actin depolymerizing factor and identification of the regulatory site. J. Biol. Chem. 1995, 270, 17582–17587. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, K.; Iida, K.; Yahara, I. Phosphorylation of Ser-3 of cofilin regulates its essential function on actin. Genes Cells 1996, 1, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Smertenko, A.P.; Jiang, C.J.; Simmons, N.J.; Weeds, A.G.; Davies, D.R.; Hussey, P.J. Ser6 in the maize actindepolymerizing factor, ZmADF3, is phosphorylated by calcium stimulated protein kinase and is essential for the control of functional activity. Plant J. 1998, 14, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Guo, L.; Wang, X.; Zhang, H.; Shi, H.; Xu, W.; Li, X. Molecular characterization of four ADF genes differentially expressed in cotton. J. Genet. Genom. 2007, 34, 347–354. [Google Scholar] [CrossRef]
- Dong, C.H.; Kost, B.; Xia, G.; Chua, N.H. Molecular identification and characterization of the Arabidopsis AtADF1, AtADF5 and AtADF6 genes. Plant Mol. Biol. 2001, 45, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Carlier, M.F.; Laurent, V.; Santolini, J.; Melki, R.; Didry, D.; Xia, G.X.; Hong, Y.; Chua, N.H.; Pantaloni, D. Actin depolymerizing factor (ADF/cofilin) enhances the rate of filament turnover: Implication in actin-based motility. J. Cell Biol. 1997, 136, 1307–1322. [Google Scholar] [CrossRef] [PubMed]
- Bamburg, J.R. Proteins of the ADF/cofilin family: Essential regulators of actin dynamics. Annu. Rev. Cell Dev. Biol. 1999, 15, 185–230. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Bernstein, B.W.; Bamburg, J.R. Regulating actin-filament dynamics in vivo. Trends Biochem. Sci. 2000, 25, 19–23. [Google Scholar] [CrossRef]
- Yeoh, S.; Pope, B.; Mannherz, H.G.; Weeds, A. Determining the differences in actin binding by human ADF and cofilin. J. Mol. Biol. 2002, 315, 911–925. [Google Scholar] [CrossRef] [PubMed]
- Allwood, E.G.; Anthony, R.G.; Smertenko, A.P.; Reichelt, S.; Drobak, B.K.; Doonan, J.H.; Weeds, A.G.; Hussey, P.J. Regulation of the pollen-specific actin-depolymerizing factor LlADF1. Plant Cell 2002, 14, 2915–2927. [Google Scholar] [PubMed]
- Jiang, C.J.; Weeds, A.G; Hussey, P.J. The maize actin-depolymerizing factor, ZmADF3, redistributes to the growing tip of elongating root hairs and can be induced to translocate into the nucleus with actin. Plant J. 1997, 12, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- McLean, B.G.; Hempel, F.D.; Zambryski, P.C. Plant intercellular communication via plasmodesmata. Plant Cell 1997, 9, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Wang, J.; Gao, P.; Jiao, G.L.; Zhao, P.M.; Li, Y.; Wang, G.L.; Xia, G.X. Down-regulation of GhADF1 gene expression affects cotton fibre properties. Plant Biotechnol. J. 2009, 7, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.; Han, Y.; Wang, X.; Wu, L.; Zhang, G.; Ma, Z. Overexpression of the Gossypium barbadense actin-depolymerizing factor 1 gene mediates biological changes in transgenic tobacco. Plant Mol. Biol. Rep. 2013, 31, 833–839. [Google Scholar] [CrossRef]
- Durst, S.; Nick, P.; Maisch, J. Nicotiana tabacum actin-depolymerizing factor 2 is involved in actin-driven, auxin-dependent patterning. J. Plant Physiol. 2013, 170, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.J. The plant hormones: Their nature, occurrence, and functions. In Plant Hormones; Springer: Dordrecht, The Netherlands, 2010; pp. 1–15. [Google Scholar]
- Campanoni, P.; Blasius, B.; Nick, P. Auxin transport synchronizes the pattern of cell division in a tobacco cell line. Plant Physiol. 2003, 133, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Maisch, J.; Nick, P. Actin is involved in auxin-dependent patterning. Plant Physiol. 2007, 143, 1695–1704. [Google Scholar] [CrossRef] [PubMed]
- Lemaire-Chamley, M.; Petit, J.; Garcia, V.; Just, D.; Baldet, P.; Germain, V.; Rothan, C. Changes in transcriptional profiles are associated with early fruit tissue specialization in tomato. Plant Physiol. 2005, 139, 750–769. [Google Scholar] [CrossRef] [PubMed]
- Gillaspy, G.; Ben-David, H.; Gruissem, W. Fruits: A developmental perspective. Plant Cell 1993, 5, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, J.J. Genetic regulation of fruit development and ripening. Plant Cell 2004, 16, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Qiao, D.; Dong, Y.; Zhang, L.; Zhou, Q.; Hu, C.; Ren, Y.; Li, Y. Ectopic expression of the maize ZmADF3 gene in Arabidopsis revealing its functions in kernel development. PCTOC 2016, 126, 239–253. [Google Scholar] [CrossRef]
- Abdrakhamanova, A.; Wang, Q.Y.; Khokhlova, L.; Nick, P. Is microtubule disassembly a trigger for cold acclimation? Plant Cell Physiol. 2003, 44, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Nick, P. Microtubules as sensors for abiotic stimuli. In Plant Microtubules; Springer: Berlin, Germany, 2008; pp. 175–203. [Google Scholar]
- Engler, J.A.; Rodiuc, N.; Smertenko, A.; Abad, P. Plant actin cytoskeleton remodeling by plant parasitic nematodes. Plant Signal. Behav. 2010, 5, 213–217. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, L.; Huang, R.D. Cytoskelton and plant salt stress tolerance. Plant Signal. Behav. 2011, 6, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhao, F.A.; Fang, W.; Xie, D.; Hou, J.; Yang, X.; Lv, S. Identification of early salt stress responsive proteins in seedling roots of upland cotton (Gossypium hirsutum L.) employing iTRAQ-based proteomic technique. Front. Plant Sci. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, L.; Yuan, M.; Ge, Y.; Liu, Y.; Fan, J.; Ruan, Y.; Cui, Z.; Tong, S.; Zhang, S. The microfilament cytoskeleton plays a vital role in salt and osmotic stress tolerance in Arabidopsis. Plant Biol. 2010, 12, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Zheng, B.; Mao, C.; Yi, K.; Liu, F.; Wu, Y.; Tao, Q.; Wu, P. cDNA-AFLP analysis of inducible gene expression in rice seminal root tips under a water deficit. Gene 2003, 314, 141–148. [Google Scholar] [CrossRef]
- Chen, C.W.; Yang, Y.W.; Lur, H.S.; Tsai, Y.G.; Chang, M.C. A novel function of abscisic acid in regulation of rice (Oryza sativa L.) roots growth and development. Plant Cell Physiol. 2006, 47, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Huang, W.L.; Hong, C.Y.; Lur, H.S.; Chang, M.C. Comprehensive analysis of differentially expressed rice actin depolymerizing factor gene family and heterologous overexpression of OsADF3 confers Arabidopsis Thaliana drought tolerance. Rice 2012. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Locus Name | ORF (bp) | Chromosomal Location | Protein | Subcellular Localization | No. of Introns | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Length (aa) | ADF Domain Start–End (aa) | MW (KDa) | pI | GRAVY | ||||||
| SlADF1 | Solyc01g094400 | 414 | 1 | 137 | 12–137 | 15.63 | 6.13 | −0.437 | Extracellular | 1 |
| SlADF2 | Solyc01g111380 | 432 | 1 | 143 | 16–143 | 16.44 | 8.69 | −0.397 | Extracellular | 2 |
| SlADF3 | Solyc03g025750 | 414 | 3 | 137 | 12–137 | 15.80 | 5.12 | −0.442 | Extracellular | 1 |
| SlADF4 | Solyc04g011370 | 432 | 4 | 143 | 12–139 | 16.49 | 5.33 | −0.469 | Extracellular | 1 |
| SlADF5 | Solyc06g005360 | 414 | 6 | 137 | 12–137 | 15.84 | 6.15 | −0.350 | Extracellular | 1 |
| SlADF6 | Solyc06g035980 | 414 | 6 | 137 | 12–137 | 15.79 | 5.50 | −0.293 | Extracellular | 1 |
| SlADF7 | Solyc09g010440 | 420 | 9 | 139 | 12–139 | 16.00 | 6.73 | −0.465 | Extracellular | 1 |
| SlADF8 | Solyc09g072590 | 450 | 9 | 145 | 18–145 | 16.69 | 7.77 | −0.594 | Extracellular | 2 |
| SlADF9 | Solyc09g090110 | 456 | 9 | 145 | 18–145 | 16.84 | 6.74 | −0.598 | Extracellular | 2 |
| SlADF10 | Solyc10g017550 | 420 | 10 | 139 | 12–139 | 15.98 | 5.63 | −0.524 | Extracellular | 1 |
| SlADF11 | Solyc10g084660 | 420 | 10 | 139 | 12–139 | 16.11 | 5.29 | −0.523 | Extracellular | 1 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatun, K.; Robin, A.H.K.; Park, J.-I.; Kim, C.K.; Lim, K.-B.; Kim, M.-B.; Lee, D.-J.; Nou, I.S.; Chung, M.-Y. Genome-Wide Identification, Characterization and Expression Profiling of ADF Family Genes in Solanum lycopersicum L. Genes 2016, 7, 79. https://doi.org/10.3390/genes7100079
Khatun K, Robin AHK, Park J-I, Kim CK, Lim K-B, Kim M-B, Lee D-J, Nou IS, Chung M-Y. Genome-Wide Identification, Characterization and Expression Profiling of ADF Family Genes in Solanum lycopersicum L. Genes. 2016; 7(10):79. https://doi.org/10.3390/genes7100079
Chicago/Turabian StyleKhatun, Khadiza, Arif Hasan Khan Robin, Jong-In Park, Chang Kil Kim, Ki-Byung Lim, Min-Bae Kim, Do-Jin Lee, Ill Sup Nou, and Mi-Young Chung. 2016. "Genome-Wide Identification, Characterization and Expression Profiling of ADF Family Genes in Solanum lycopersicum L." Genes 7, no. 10: 79. https://doi.org/10.3390/genes7100079
APA StyleKhatun, K., Robin, A. H. K., Park, J.-I., Kim, C. K., Lim, K.-B., Kim, M.-B., Lee, D.-J., Nou, I. S., & Chung, M.-Y. (2016). Genome-Wide Identification, Characterization and Expression Profiling of ADF Family Genes in Solanum lycopersicum L. Genes, 7(10), 79. https://doi.org/10.3390/genes7100079