3.1. Mathematical Modelling of the Integrated MST2-ERK Network Dynamics
To investigate the quantitative and dynamic properties of the integrated MST2-ERK network, we first extended a previously published model [
6]. In this model, Akt and Ras were considered as direct inputs into the model as time-dependent functions for simplicity, where the forms of these functions were informed by experimental data. In this study, we extended the network model by introducing the detailed regulatory mechanisms of Ras and Akt activation, and EGFR as a new input node. This extension was necessary to directly calibrate the model against dynamic data of Akt, and enable a systematic investigation of potentially different roles of crosstalk/feedback mechanisms between the ERK and MST2 pathways. The resulting model structure (i.e., reaction scheme) is given in
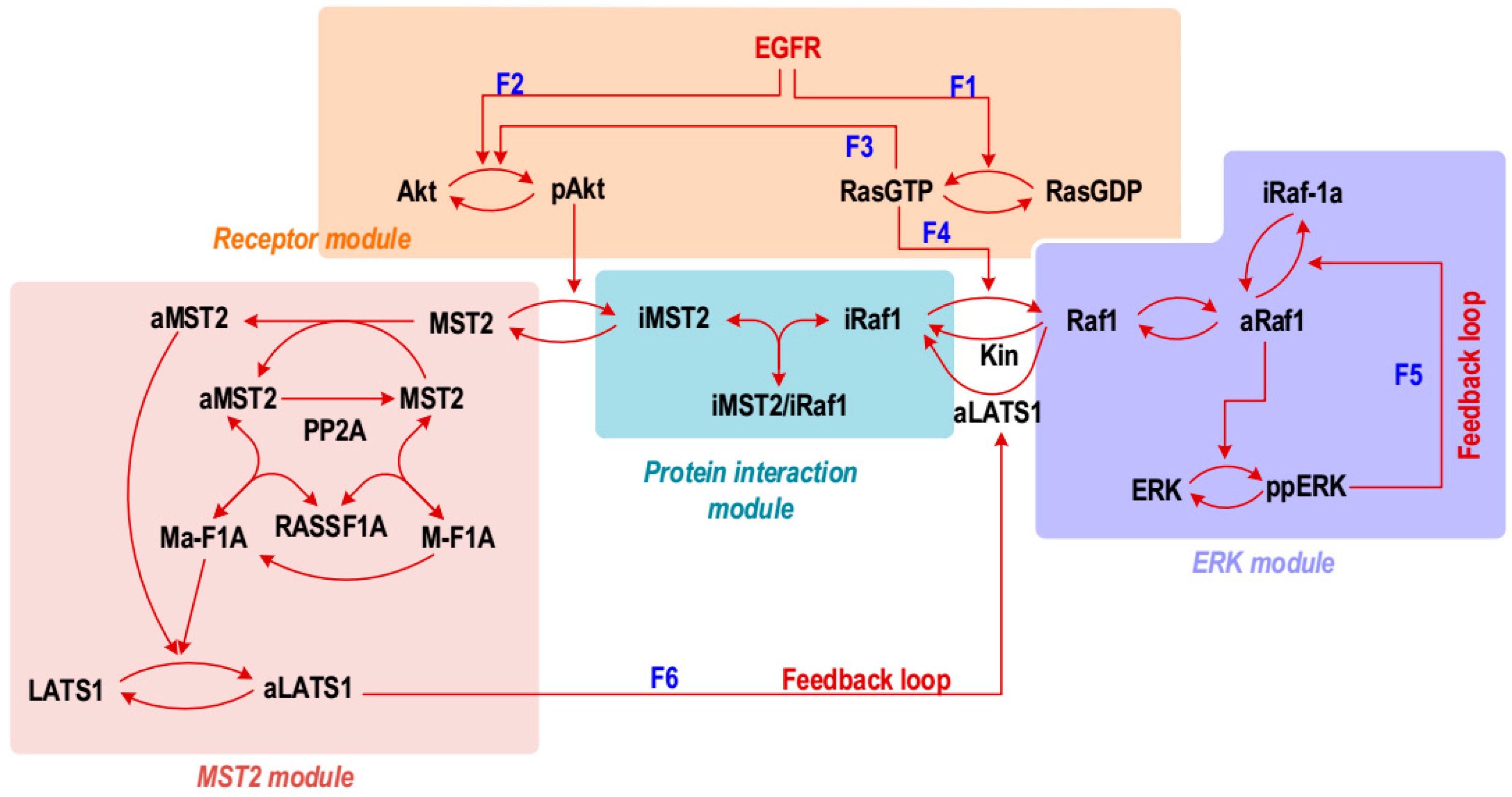
Figure 1, which explicitly describe the mechanistic activation of Akt and Ras following activation of the EGF receptor (EGFR). As such, the new model consists of four sub-modules, as highlighted in
Figure 1. (i) The
receptor module includes Ras and Akt activation triggered by activated EGFR, as well as Ras-induced Akt activation [
13,
14,
15]; (ii) The
ERK module describes activation of the Raf-1/MEK/ERK MAPK cascade initiated by Ras mediated Raf-1 activation, including the negative feedback loop from ERK to Raf-1 [
16]. To reduce the number of model parameters without significantly compromising the network dynamics properties, the MEK and ERK activation steps are lumped into a single step; (iii) The
MST2 module describes activation of the MST2/LATS signalling cascade, which is mediated by the scaffold protein Ras association domain family 1 isoform A (RASSF1A). Importantly, this module contains the cross-pathway feedback from LATS1 to Raf-1, where active LATS phosphorylates Raf-1 and promotes its complexes formation with MST2, thereby inhibiting Raf-1 activity [
6,
8]; (iv) The
protein interaction module describes the non-catalytic binding between MST2 and Raf-1, and that this binding is promoted when MST2 and Raf-1 are phosphorylated by Akt and LATS1, respectively [
6,
7,
8]. This module together with the LATS1 mediated feedback loops constitute critical mechanisms that link the Hippo and ERK pathways in mammalian cells.
Thus, the developed model incorporates protein interactions, phosphorylation reactions and feedback loops, which was described by ordinary differential equations (ODEs). The resulting model comprises 20 ODEs and a total of 50 kinetic parameters, described using a combination of mass-action and Michaelis-Menten kinetics (see
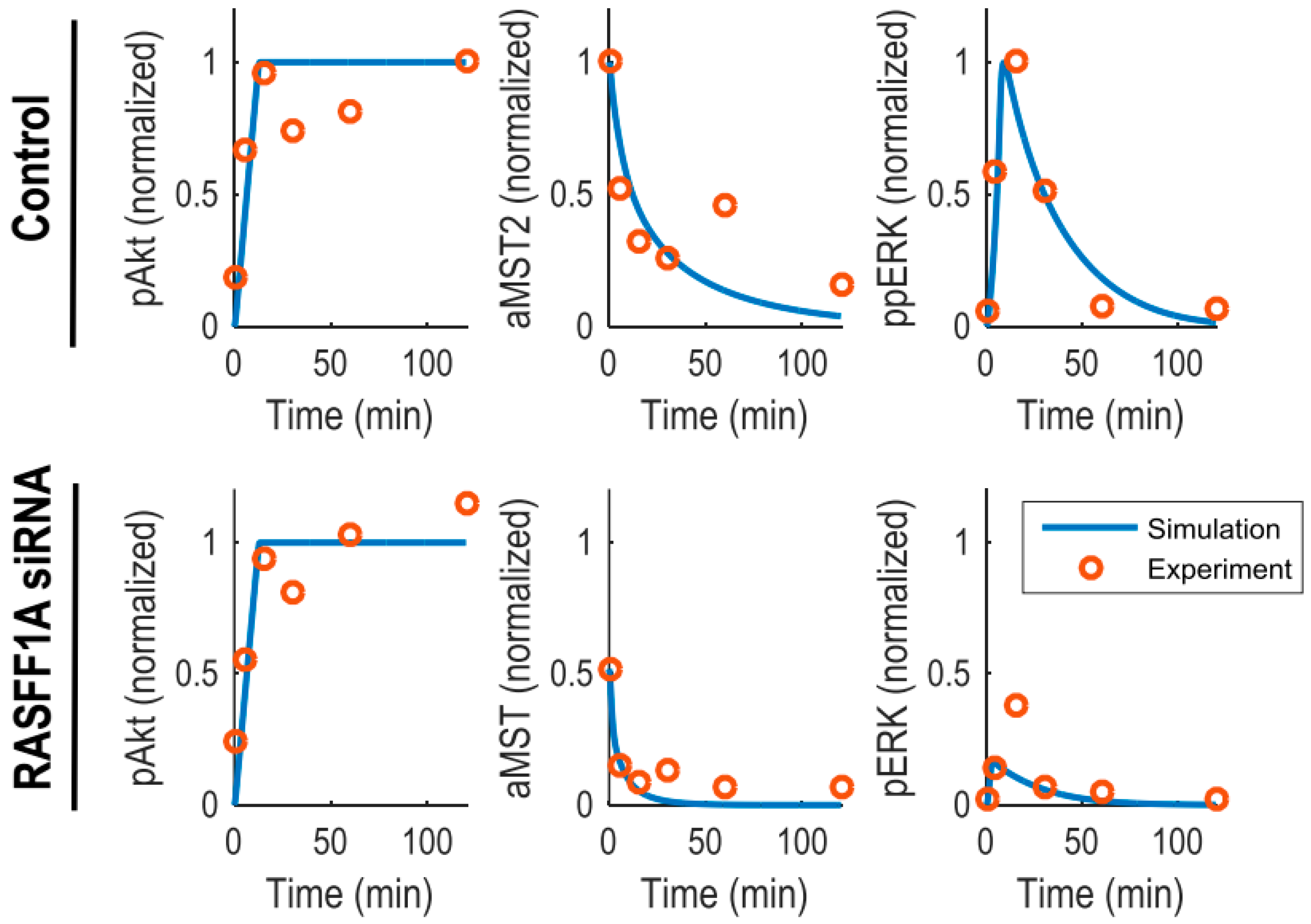
Supplementary Materials Tables S1 and S2). To generate quantitative predictions, we calibrated the model and estimated kinetic parameters using a set of time-course data obtained from HeLa cells [
6]. These include the levels of phosphorylated (active) Akt (pAkt), ERK (ppERK) and MST2 (aMST2) measured at 6 time points over 2 h serum stimulation under untreated and RASSF1A knockdown conditions (
Figure 2). The data show that while RASSF1A knockdown did not affect the kinetics of Akt activation, it significantly downregulated the basal and temporal levels of MST2 activity. Similarly, ERK activity were remarkably reduced by downregulation of RASSF1A, however its transient dynamics was retained albeit with a lower peak. For parameter estimation, we used genetic algorithm, a computational method that was inspired by evolutionary biology concepts such as selection, crossover, mutation, etc. [
17,
18,
19]. In engineering fields, this algorithm is widely exploited to find approximate solutions of optimization problems [
20]. To ensure the biological relevance of the kinetic parameters during, we constrained the parameters within their respective physiologically plausible ranges before fitting. These ranges were informed by previous studies [
16,
21,
22,
23], and also used in our previous work [
24,
25]. For examples, k
a (association rate) = [1 × 10
−4, 1 × 10
4] (nM
−1·min
−1); k
d (dissociation rate) = [1 × 10
−4, 1 × 10
1] (min
−1); k
c (catalytic rate) = [1 × 10
−4, 1 × 10
3] (min
−1); V
max (max velocity) = [1 × 10
−4, 1 × 10
3] (nM·min
−1) and K
m (Michaelis-Menten constant) = [1 × 10
−4, 1 × 10
4] (nM). Model simulations of the best fitted parameter set showed that the model quantitatively recapitulates all the observed dynamics of the MST2-ERK network very well (
Figure 2).
3.2. Diverse Dose-Response Profiles Generated by the Interlinked Hippo-ERK Network
Feedback loops have been long known to generate complex and non-linear systems dynamics [
16,
22,
26,
27,
28,
29]. Our recent work further demonstrates that reversible, non-catalytic protein-protein interaction motifs could also bring about rich repertoire of complex dynamics features to signalling cascades by generating hidden feedback and feed-forward loops [
7,
30]. Given that the Hippo-ERK network features both these types of design in its wiring, we hypothesize that they can provoke diverse systems dynamics patterns under different cellular contexts. Nevertheless, what these patterns are and when or how they may arise are questions not yet examined.
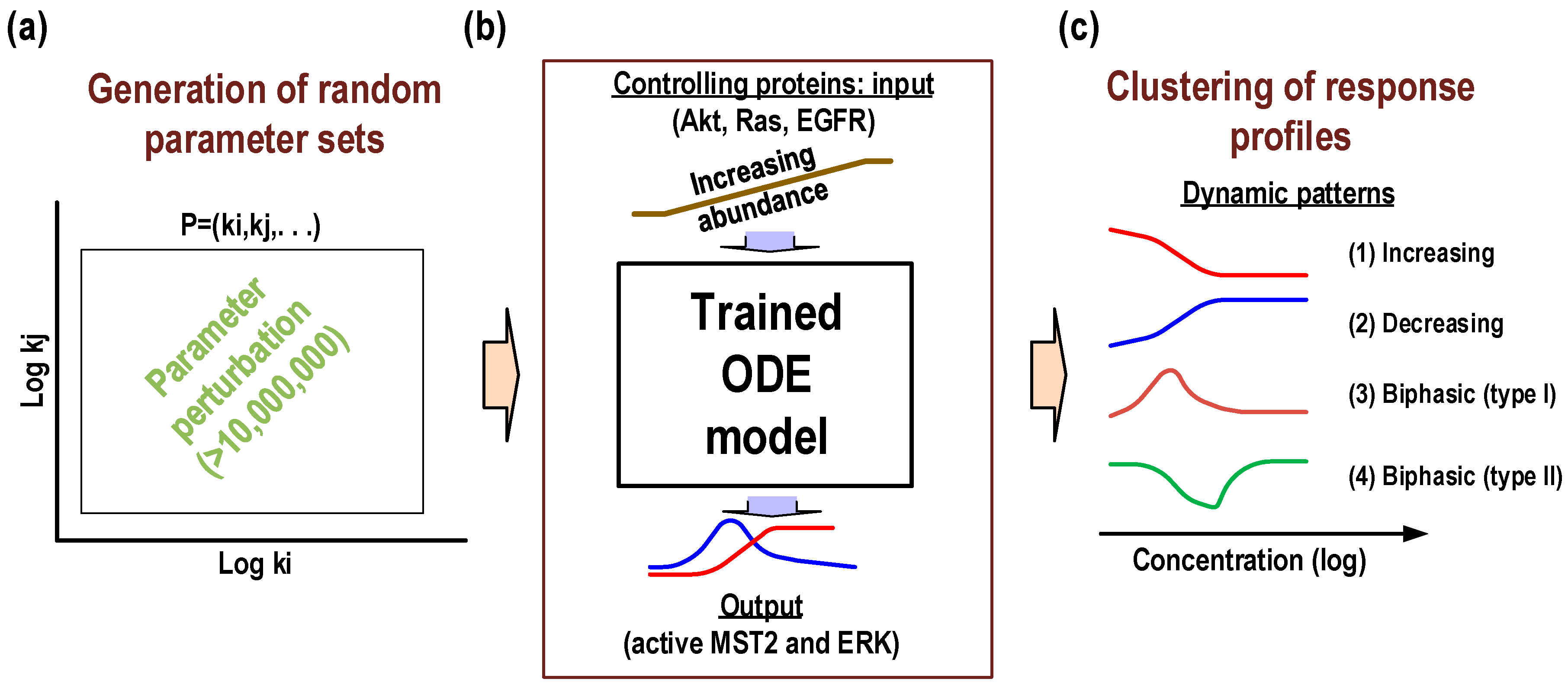
To address these questions, we carried out extensive computational simulations and analyses using the calibrated model. We focus on the steady-state behaviours of the system by simulating the dose-response profiles (i.e., input-output relationships) of active MST2 (aMST2) and ERK (ppERK), considered as the network’s two major outputs, in response to increasing abundance of specific controlling inputs, in this case of Akt, Ras and EGFR, referred to as the “controlling proteins” (
Figure 3). The crosstalk and feedback interactions between MST2 and ERK pathways are expected to play important roles in determining the dynamic behaviours of the overall network. Thus, to explore possible dose-response patterns we selected six critical regulatory links (denoted F
1 to F
6 in
Figure 1) that mediate the crosstalk and feedback regulations between the pathways, and varied the kinetic parameters underlying these six links randomly and in combination within large ranges around their nominal values (i.e., between 10
−3 and 10
3 fold of the fitted values,
Figure 3). Note that the remaining parameters were fixed at their fitted values. A total of >10 million parameter sets were generated, for each set the described dose-response profiles were simulated. While network topology is likely conserved between cell types or tissues, the strength of network connections may strongly vary between different molecular contexts [
11,
12,
31]. The extensive parameter variation introduced in our simulations is thus essential to ensure we fully capture the dynamics repertoire of the network that may occur under various molecular backgrounds.
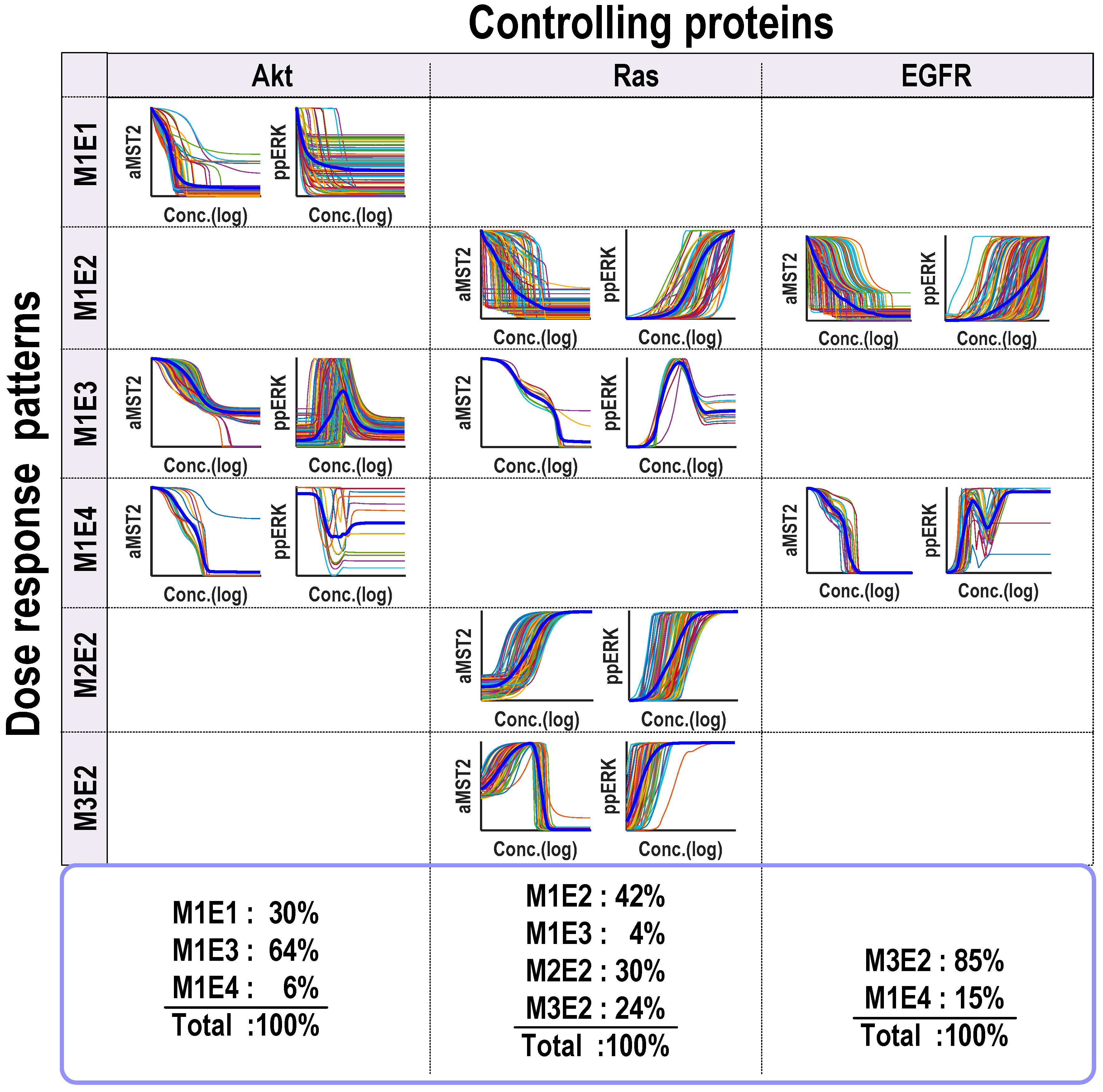
Model simulations showed that the Hippo-ERK network could generate diverse response profiles, which can be clustered into several distinct dose-response patterns for both the outputs, active MST2 and ERK (
Figure 4). Specifically, four different dose-response patterns that occur frequently were identified: (1) increasing; (2) decreasing; (3) biphasic type I and (4) biphasic type II. Under these definition, patterns 1 (or 2) represents a monotonic increase (or decrease) of active levels of the output to increasing abundance of the controlling input protein; while patterns 3 (or 4) describes a biphasic response of the output to increasing abundance of the controlling protein in a concave-down (or concave-up) manner. Combining both network outputs (active MST2 and ERK), we found that the Hippo-ERK network can generate a total of 8 different patterns for the three controlling proteins (Akt, Ras and EGFR). Shown in
Figure 4, Akt can trigger 3 distinct combined patterns (denoted M
1E
1, M
1E
3 and M
1E
4) as the regulating protein; while Ras can generate 4 patterns (M
1E
2, M
1E
3, M
2E
2 and M
3E
2) and EGFR produces 2 patterns (M
1E
2 and M
1E
4). Here, the notation “M
iE
j” means active MST2 (“M”) follows pattern
i and active ERK (“E”) follows pattern
j as defined previously (subscripts
i and
j can range between 1 and 4).
Although in principle each controlling protein can generate up to 16 combinatorial dose-response patterns for MST2 and ERK on the basis of the 4 defined patterns, the fact that only a limited subset of these are actually found indicates that the systems dynamics are well constrained by the network wiring. At the same time, since more than one patterns are observed for each controlling proteins, the Hippo-ERK systems dynamics also display robust plasticity determined by the network’s kinetic parameters. These observations confirm the view that systems dynamics are a product of both network topology and network parameterization [
32,
33].
3.3. Akt Generates Diverse ERK Response Patterns
Our model simulations previously showed that Akt can generate three distinct response patterns, namely M
1E
1, M
1E
3 and M
1E
4 (
Figure 4, first column). Interestingly, while Akt always decreases MST2 activity, it induces diverse response profiles of ERK activity, provoking a decreasing ERK activity response in some case, and triggering biphasic ERK activity responses in others. In fact, M
1E
3 is the most frequently observed pattern, accounting for 64% of the total sampled parameter sets, followed by M
1E
1 (30%) and M
1E
4 (4%). Here, we seek to understand how Akt can generate such different dynamic behaviours and aim to provide a mechanistic explanation of the underlying mechanisms in each case.
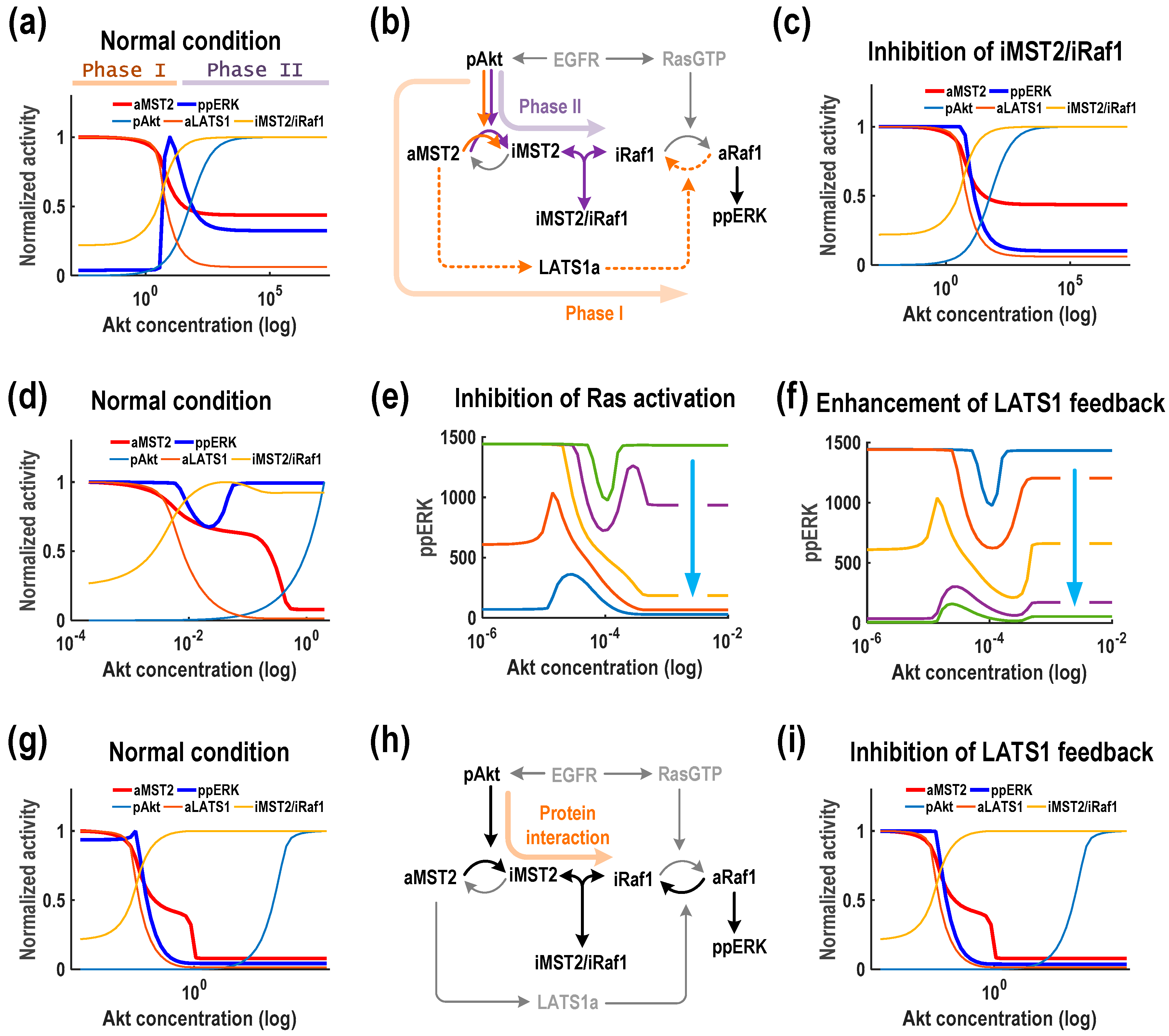
We first consider the most common pattern M
1E
3, where Akt induces a biphasic (concave-down) response of ERK activity. This pattern indicates that Akt could play opposing roles in regulating ERK activity which switch in a dose-dependent manner: Akt functions as a promoter when its levels are low, but as an inhibitor when its levels are high. To investigate the underlying mechanisms of this biphasic profile, we simulated the steady-state levels of key network components in response to increasing Akt, as shown in
Figure 5a. Analysing these curves and the network wiring suggests that ERK’s biphasic profile can be generated mechanistically through two different phases as the balance of network regulation mediated by the LATS1 mediated feedback and MST2-Raf-1 interaction tips. Specifically, increasing Akt starting at a low level phosphorylates and inhibits MST2, thus relieving the LATS1-to-Raf-1 negative feedback, leading to increased Raf-1/ERK activity (Phase I). However, further increase in Akt promotes accumulation of the inactive form of MST2 which sequesters Raf-1 from being activated by Ras via formation of the MST2-ERK complexes, thus resulting in suppressed ERK activity at high Akt levels (Phase II, indicated in
Figure 5b). To confirm that this mechanism indeed underlies M
1E
3, we first blocked the protein interaction module by setting the association rate between MST2 and Raf-1 to null. Consequently, the biphasic ERK activity profile was abolished and replaced by a monotonic decreasing pattern; whereas the activity profile MST2 was not affected (
Figure 5c). Next, we inhibited the LATS1 feedback loop and found that the ERK activity profile was replaced by a monotonic increasing pattern (
Supplementary Materials Figure S1a). Together, these simulations support that the Raf-1/MST2 interaction module interplays with the LATS1 mediated feedback to bring about the dual, opposite roles of Akt in ERK regulation.
Another intriguing pattern identified in our simulations was M
1E
4 where ERK activity displays the type II biphasic (concave-up) response against increasing Akt. In contrast to the previous case, here Akt acts as an inhibitor of ERK at low levels, but as a facilitator at high levels (
Figure 5d). ERK activity is high at both low and high Akt levels, which suggests that Raf-1 is also highly active at these concentrations. Raf-1 activation is controlled by Ras and LATS1 as opposing regulators: Ras-induced phosphorylation at Serine 338 activates Raf-1 but LATS1-induced phosphorylation at Serine 259 inhibits it. Thus, at a lower concentration, Akt can inhibit Raf-1 activation through promoting the inhibitory iMST2/iRaf-1 complex formation and thus, Raf-1 activation was reduced with increasing Akt amount. On the other hand, at higher concentrations Akt phosphorylates and inhibits MST2 activity toward LATS1, which releases the LATS1 inhibition of Raf-1 through the feedback loop and thereby increasing ERK activity. Therefore, we asked how breaking the Ras/LATS1 balance may influence the response pattern of ERK. Interestingly, graded suppression of Ras activity transforms ERK activity from a concave-up biphasic to a monotonic decreasing pattern to a concave-down biphasic pattern (
Figure 5e). Exactly the opposite observation was made when we strengthened the LATS1 feedback loop (
Figure 5f). Taken together, these results suggest that which function Akt plays in regulating ERK activity is critically dependent on the balance between Ras and LATS1.
The last pattern found for Akt as the controlling protein was M1E1 in which Akt consistently inhibits both ERK and MST2 activation (
Figure 5g). In this case, the protein interaction module plays a major role in driving these effects, as MST2-Raf-1 binding acts a strong negative regulator of ERK activation while the effect from LATS1 feedback and Ras are both relatively weak (
Figure 5h). Indeed, blocking the LATS1 feedback (and Ras activation) did not really affect the network responses (
Figure 5i and
Supplementary Materials Figure S1b). An intuitive explanation is that increasing Akt depletes active MST2, thereby weakening the LATS feedback. Concurrently, accumulated inactive MST2 sequesters Raf-1 through their binding, leading to downregulated Raf-1 and thus ERK activity (
Figure 5h).
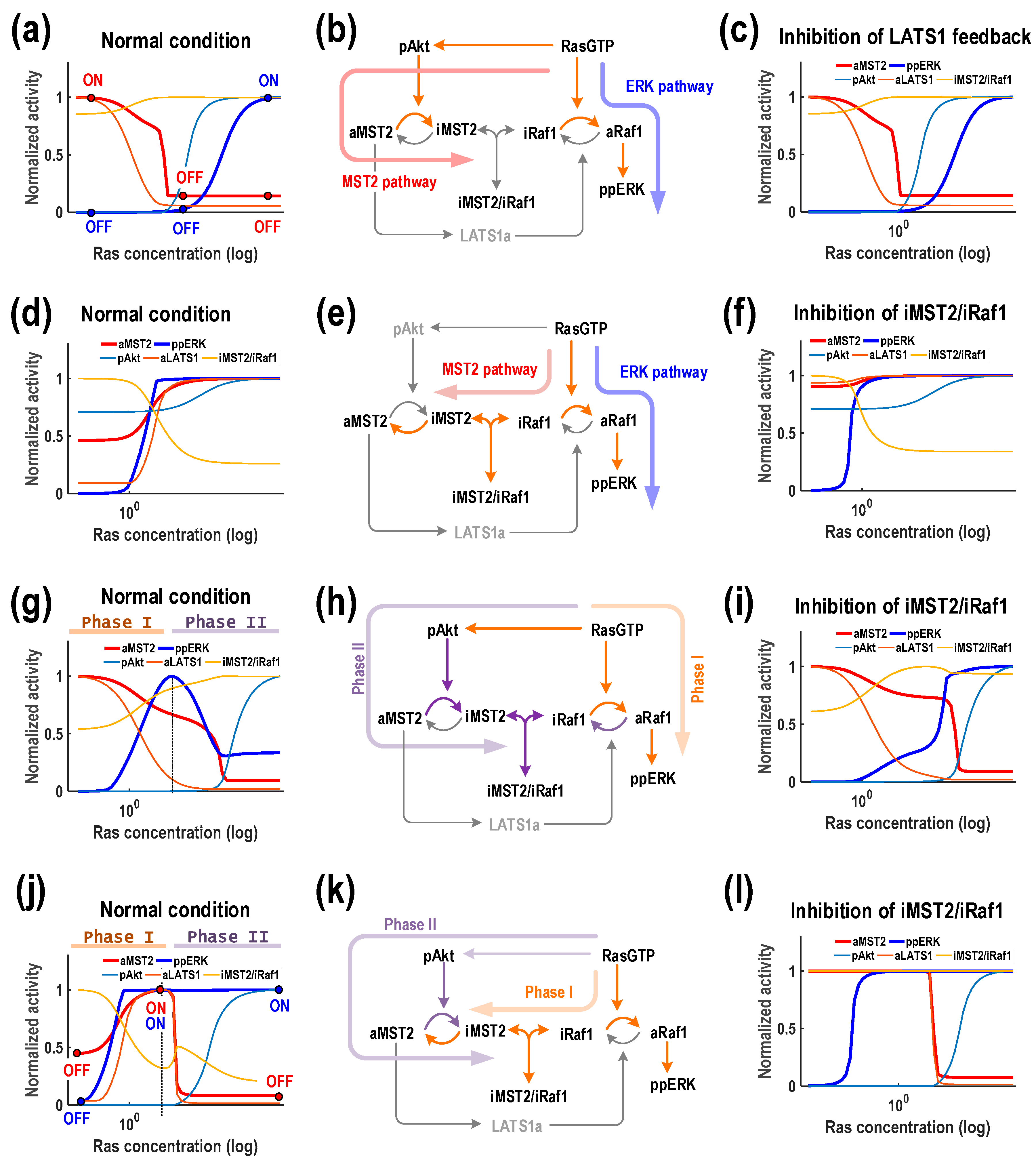
3.4. Ras Displays Multi-Functional Role and Drives Biphasic Responses of both MST2 and ERK
Unlike Akt, our simulations revealed that Ras can generate multiple response patterns for both MST2 and ERK activities. Specifically, Ras induces four different patterns, M
1E
2, M
1E
3, M
2E
2 and M
3E
2, showing that it can either promote or inhibit MST2 and ERK under specific conditions, and even display both roles in some cases (
Figure 4, the second column). Since Ras is directly upstream of Raf-1, it is natural to expect increasing Ras abundance leads to increased ERK activity, and this was indeed seen in 3 observed patterns (M
1E
2, M
2E
2 and M
3E
2) which together account for the majority of the total simulated parameter sets (96%). What was not expected is that the Ras-induced MST2 activity profile is distinct in each case. We first consider pattern M
1E
2 where increasing Ras robustly downregulates MST2 activity (
Figure 6a). This downregulation can be attributed to the link between Ras and Akt, as Ras promotes Akt activity to inhibit MST2. We observed that the switches regarding MST2 (“off”) and ERK activity (“on”) occurred at different Ras thresholds, which interestingly suggests that depending on Ras concentration, the system can turn ON only MST2 (low Ras), only ERK (high Ras), or shut down both (intermediate Ras). Ras thus serves multi-functional effect depending on its abundance. To evaluate whether the MST2/Raf-1 interaction module and/or the LATS1 feedback contribute to induction of the M
1E
2 pattern, we simulated the effect of blocking either the formation of Raf-1/MST2 complexes or the LATS1 feedback loop and found that these did not really influence response patterns of the system (
Figure 6c and
Supplementary Materials Figure S2a). This suggests that the M
1E
2 pattern emerged primarily through the Ras/Akt link but not the protein interaction module nor the LATS1 mediated feedback mechanism (
Figure 6b).
Ras, in contrast, promotes MST2 signalling in M2E2 (
Figure 6d). To tease out possible mechanisms governing this rather counterintuitive monotonic increase of MST2, we first perturbed the LATS1 feedback loop, which showed that varying the strength of this feedback has no effect on MST2 activity (
Supplementary Materials Figure S2b). Perturbation of the protein interaction module however dramatically abolished the steep Ras-dependent increase of MST2 activation, turning it into an almost constant response (
Figure 6f). These simulations lead to an explanation that Ras induced activation of Raf-1 steers it away from the MST2/Raf-1 complexes, leaving more available MST2 free for dimerization, resulting in increased MST2 activity (
Figure 6e). This is supported by the steep depletion of the MST2/Raf-1 complexes as Ras levels increases (
Figure 6d). In contrast to M
1E
2, the Ras/Akt link is necessarily weak here, which may be the case in specific cell types.
This leaves the remaining patterns M
1E
3 and M
3E
2, where Ras triggers biphasic response profiles in ERK and MST2, respectively (
Figure 6g,j). Like M
1E
2, the decreasing pattern for MST2 in M
1E
3 is underlined by a strong Ras-to-Akt link (
Figure 6g). However, the biphasic nature of ERK activity in this pattern was caused by presence of a strong protein interaction module featuring tight association between MST2 and Raf-1 (
Figure 6h), which was lacking in M
1E
2. Indeed, relaxing this association completely eliminated ERK biphasic profile, while largely maintained other components’ behaviours (
Figure 6i). Thus, the biphasic response of ERK can be explained by splitting it into 2 phases as depicted in
Figure 6g: low Ras upregulates ERK (Phase I), but as Ras level continues to rise, Akt is activated leading to accumulated inactive MST2 which subsequently sequesters Raf-1 into the complexes and inhibits its activation (Phase II). Of note, blocking the LATS1 feedback did not affect the overall systems responses, implying that this feedback is not functional in this pattern (
Supplementary Materials Figure S2c). Finally, we examine M
3E
2 where Ras provokes essentially opposite dynamic behaviours: a biphasic response for MST2 and increasing response for ERK. Under this regime, simulations showed that Ras can turn off (or on) both MST2 and ERK, or selectively only turn on ERK activity (
Figure 6j). The MST2/Raf-1 interaction module was again found critical in generating a biphasic MST2, as a weakened interaction completely removed this pattern (
Figure 6l). Note that blocking the LATS1 feedback did not affect the overall systems responses (
Supplementary Materials Figure S2d). Intuitively, the increase in MST2 activity at low Ras is due to more MST2 being available for activation as Raf-1 is sequestered away from its complexes with MST2 by Ras (Phase I). As Ras level rises, Akt becomes activated and starts to suppress MST2 by promoting the complexes formation (Phase II,
Figure 6k).
Taken together, the above analyses revealed Ras as a highly flexible and dynamic regulator of both MST2 and ERK activity. Although a Ras-induced biphasic response of MST2 seems to be a rare event (only 4% of the total simulations) compared to other patterns, it nevertheless could occur given the right conditions. Which role Ras exactly assumes is strongly dependent on its abundance in cells, and is not trivially recognised without detailed analysis of the network structure.
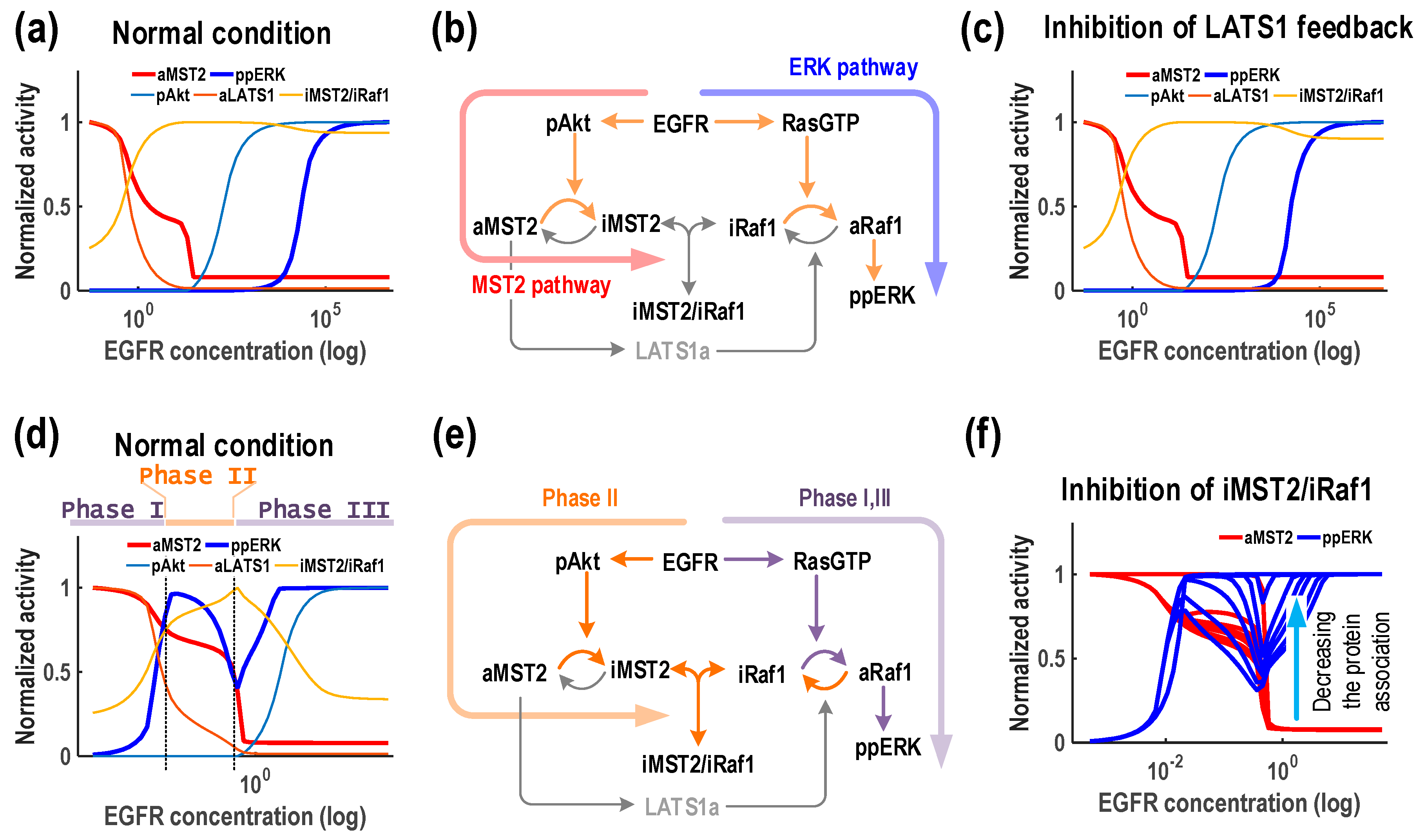
3.5. EGFR Generates Multi-Phasic ERK Response
We also examined possible dynamics generated by EGFR and detected only two different dynamic patterns (M
1E
2 and M
1E
4), in both cases MST2 showed a monotonic decrease trend against rising EGFR expression. ERK activity displayed a monotonic increase profile in M
1E
2, (accounting for most of the simulations at 85%) but showed a combined pattern of both type I and type II biphasic response in M1E4 (15%,
Figure 4, right column).
The M
1E
2 pattern and its explanation were similar to that induced by Ras (
Figure 6a) since EGFR is direct upstream of Ras which can also directly promotes Akt signalling. M
1E
4 was more curious as the activation of ERK showed an initial increase (Phase I) at low EGFR, followed by a decrease (Phase II) at intermediate EGFR levels, but bounced back (Phase III) again at high EGFR (
Figure 7d). ERK activation in Phase I can be explained by the direct activation of Raf-1/ERK via Ras. Phase II was due to accumulation of inactive MST2 by EGFR and Ras induced Akt, which binds to and inhibits Raf-1 and thus ERK. Phase III can be brought about by further activation of Ras that overrides the inhibitory effect of MST2 through the interaction with Raf-1. We confirmed this mechanism by blocking the protein association between Raf-1 and MST2 and found that the activation of ppERK did not display the temporal decrease pattern (
Figure 7f).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






