
Genetic Variants of Retinoic Acid Receptor-Related Orphan Receptor Alpha Determine Susceptibility to Type 2 Diabetes Mellitus in Han Chinese

Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Biochemical Measurements
2.3. SNP Selection and Genotyping
2.4. Statistical Analyses
3. Results
3.1. General Characteristics of the Subjects
3.2. Distribution of the SNPs in RORA between T2DM Patients and Controls
3.3. Association of Genotypes with T2DM under Different Genetic Models
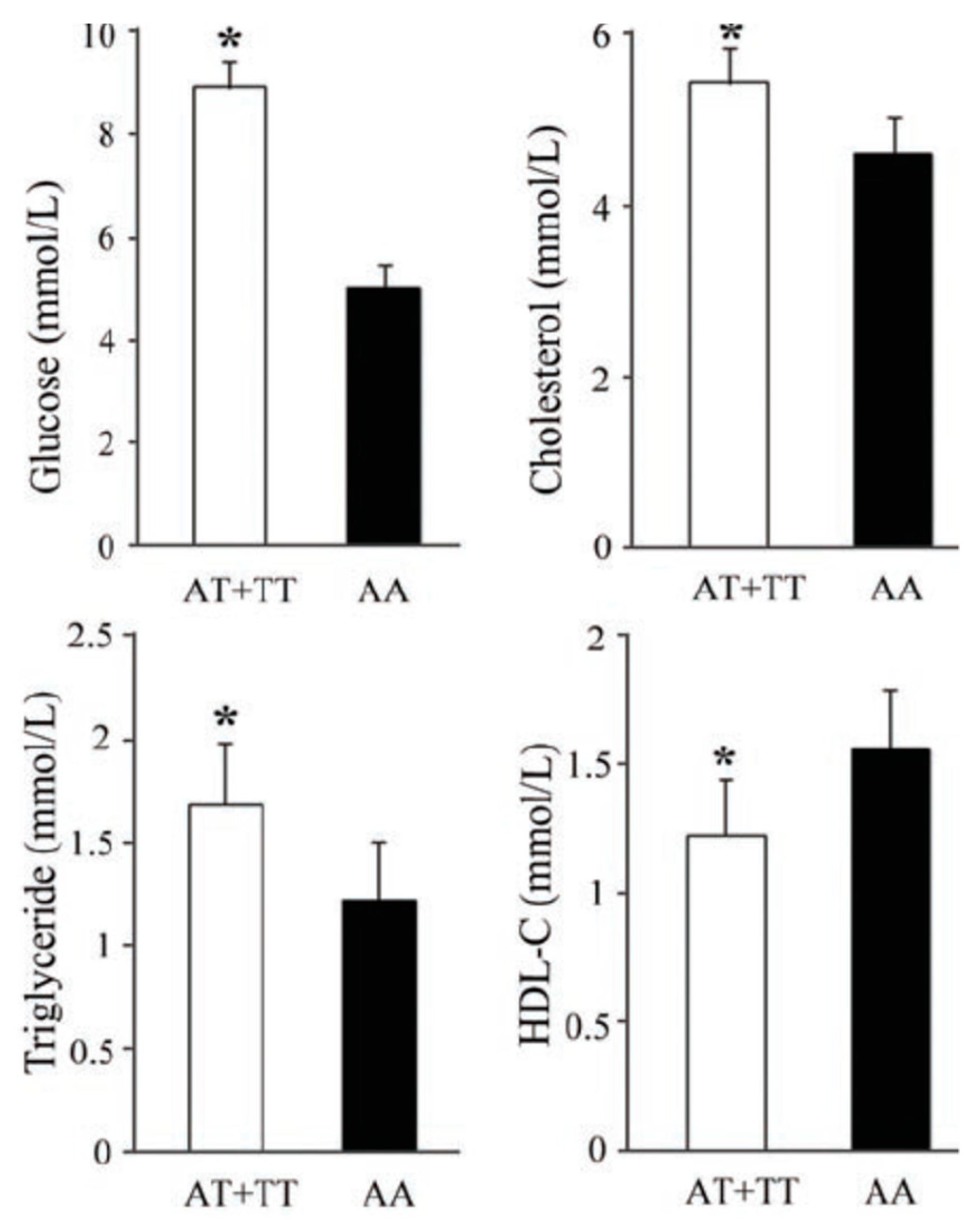
3.4. Effects of rs10851685 on the Different Metabolic Parameters
3.5. RORA Haplotypes and T2DM
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Chen, L.; Magliano, D.J.; Zimmet, P.Z. The worldwide epidemiology of type 2 diabetes mellitus—Present and future perspectives. Nat. Rev. Endocrinol. 2011, 8, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Lu, J.; Weng, J.; Jia, W.; Ji, L.; Xiao, J.; Shan, Z.; Liu, J.; Tian, H.; Ji, Q.; et al. Prevalence of diabetes among men and women in China. N. Engl. J. Med. 2010, 362, 1090–1101. [Google Scholar] [CrossRef] [PubMed]
- Stumvoll, M.; Goldstein, B.J.; van Haeften, T.W. Type 2 diabetes: Pathogenesis and treatment. Lancet 2008, 371, 2153–2156. [Google Scholar] [CrossRef]
- Mohlke, K.L.; Boehnke, M. Recent advances in understanding the genetic architecture of type 2 diabetes. Hum. Mol. Genet. 2015, 24, R85–R92. [Google Scholar] [CrossRef] [PubMed]
- Giguere, V.; Tini, M.; Flock, G.; Ong, E.; Evans, R.M.; Otulakowski, G. Isoform-specific amino-terminal domains dictate DNA-binding properties of RORα, a novel family of orphan hormone nuclear receptors. Genes Dev. 1994, 8, 538–553. [Google Scholar] [CrossRef] [PubMed]
- Vu-Dac, N.; Gervois, P.; Grotzinger, T.; De Vos, P.; Schoonjans, K.; Fruchart, J.C.; Auwerx, J.; Mariani, J.; Tedgui, A.; Staels, B. Transcriptional regulation of apolipoprotein A-I gene expression by the nuclear receptor RORA. J. Biol. Chem. 1997, 272, 22401–22404. [Google Scholar] [CrossRef] [PubMed]
- Raspé, E.; Duez, H.; Gervois, P.; Fiévet, C.; Fruchart, J.-C.; Besnard, S.; Mariani, J.; Tedgui, A.; Staels, B. Transcriptional regulation of apolipoprotein C-III gene expression by the orphan nuclear receptor RORA. J. Biol. Chem. 2001, 276, 2865–2871. [Google Scholar] [CrossRef] [PubMed]
- Genoux, A.; Dehondt, H.; Helleboid-Chapman, A.; Duhem, C.; Hum, D.W.; Martin, G.; Pennacchio, L.A.; Staels, B.; Fruchart-Najib, J.; Fruchart, J.C. Transcriptional regulation of apolipoprotein A5 gene expression by the nuclear receptor RORA. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1186–1192. [Google Scholar] [CrossRef] [PubMed]
- Chauvet, C.; Vanhoutteghem, A.; Duhem, C.; Saint-Auret, G.; Bois-Joyeux, B.; Djian, P.; Staels, B.; Danan, J.-L. Control of gene expression by the retinoic acid-related orphan receptor alpha in HepG2 human hepatoma cells. PLoS ONE 2011, 6, e22545. [Google Scholar] [CrossRef] [PubMed]
- Kuang, J.; Hou, X.; Zhang, J.; Chen, Y.; Su, Z. Identification of insulin as a novel retinoic acid receptor-related orphan receptor alpha target gene. FEBS Lett. 2014, 588, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Gold, D.A.; Gent, P.M.; Hamilton, B.A. ROR alpha in genetic control of cerebellum development: 50 staggering years. Brain Res. 2007, 1140, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Lau, P.; Fitzsimmons, R.; Pearen, M.; Watt, M.; Muscat, G.E. Homozygous staggerer (sg/sg) mice display improved insulin sensitivity and enhanced glucose uptake in skeletal muscle. Diabetologia 2011, 54, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Lau, P.; Fitzsimmons, R.L.; Raichur, S.; Wang, S.-C.M.; Lechtken, A.; Muscat, G.E. The orphan nuclear receptor, RORA, regulates gene expression that controls lipid metabolism. J. Biol. Chem. 2008, 283, 18411–18421. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Li, Y.; James, J.C.; Matsumoto, A.H.; Helm, G.A.; Lusis, A.J.; Shi, W. Genetic linkage of hyperglycemia, body weight and serum amyloid-P in an intercross between C57BL/6 and C3H apolipoprotein E-deficient mice. Hum. Mol. Genet. 2006, 15, 1650–1658. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.G.; Pluzhnikov, A.; Miyake, K.; Sun, Y.; Ng, M.C.Y.; Roe, C.A.; Below, J.E.; Nicolae, R.I.; Konkashbaev, A.; Bell, G.I. Identification of type 2 diabetes genes in Mexican Americans through genome-wide association studies. Diabetes 2007, 56, 3033–3044. [Google Scholar] [CrossRef] [PubMed]
- Rampersaud, E.; Damcott, C.M.; Fu, M.; Shen, H.; McArdle, P.; Shi, X.; Shelton, J.; Yin, J.; Chang, Y.P.; Ott, S.H.; et al. Identification of novel candidate genes for type 2 diabetes from a genome-wide association scan in the Old Order Amish: Evidence for replication from diabetes-related quantitative traits and from independent populations. Diabetes 2007, 56, 3053–3062. [Google Scholar] [CrossRef] [PubMed]
- Solt, L.A.; Burris, T.P. Action of RORs and their ligands in (patho)physiology. Trends Endocrinol. Metab. 2012, 23, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Solt, L.A.; Griffin, P.R.; Burris, T.P. Ligand regulation of retinoic acid receptor-related orphan receptors: Implications for development of novel therapeutics. Curr. Opin. Lipidol. 2010, 21, 204–211. [Google Scholar] [CrossRef] [PubMed]
- HapMap. Available online: http://www.hapmap.org/ (accessed on 1 January 2015).
- Yuan, Y.; Jiang, H.; Kuang, J.; Hou, X.; Feng, Y.; Su, Z. Genetic variations in ADIPOQ gene are associated with chronic obstructive pulmonary disease. PLoS ONE 2012, 7, e50848. [Google Scholar] [CrossRef] [PubMed]
- Bland, J.M.; Altman, D.G. Statistics notes. The odds ratio. BMJ 2000, 320, 1468. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G.; Bland, J.M. How to obtain the confidence interval from a p value. BMJ 2011, 343, d2090. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar]
- Chopra, A.R.; Louet, J.F.; Saha, P.; An, J.; Demayo, F.; Xu, J.; York, B.; Karpen, S.; Finegold, M.; Moore, D.; et al. Absence of the SRC-2 coactivator results in a glycogenopathy resembling Von Gierke’s disease. Science 2008, 322, 1395–1399. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Solt, L.A.; Burris, T.P. Regulation of FGF21 expression and secretion by retinoic acid receptor-related orphan receptor alpha. J. Biol. Chem. 2010, 285, 15668–15673. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.M.; Rimm, E.B.; Colditz, G.A.; Stampfer, M.J.; Willett, W.C. Obesity, fat distribution, and weight gain as risk factors for clinical diabetes in men. Diabetes Care 1994, 17, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.S.; Okamoto, K.; Takeda, Y.; Beak, J.Y.; Gerrish, K.; Bortner, C.D.; DeGraff, L.M.; Wada, T.; Xie, W.; Jetten, A.M. Transcriptional profiling reveals a role for RORA in regulating gene expression in obesity-associated inflammation and hepatic steatosis. Physiol. Genom. 2011, 43, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Parhofer, K.G. Diabetic dyslipidemia. Metabolism 2014, 63, 1469–1479. [Google Scholar] [CrossRef] [PubMed]
- Mamontova, A.; Seguret-Mace, S.; Esposito, B.; Chaniale, C.; Bouly, M.; Delhaye-Bouchaud, N.; Luc, G.; Staels, B.; Duverger, N.; Mariani, J.; et al. Severe atherosclerosis and hypoalphalipoproteinemia in the staggerer mouse, a mutant of the nuclear receptor RORα. Circulation 1998, 98, 2738–2743. [Google Scholar] [CrossRef] [PubMed]
- Korb, M.; Ke, Y.B.; Johnson, L.F. Stimulation of gene expression by introns: Conversion of an inhibitory intron to a stimulatory intron by alteration of the splice donor sequence. Nucl. Acids Res. 1993, 21, 5901–5908. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.R.; Wegmann, D.; Ehm, M.G.; Kessner, D.; St Jean, P.; Verzilli, C.; Shen, J.; Tang, Z.; Bacanu, S.A.; Fraser, D.; et al. An abundance of rare functional variants in 202 drug target genes sequenced in 14,002 people. Science 2012, 337, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Gan, W.; Walters, R.G.; Holmes, M.V.; Bragg, F.; Millwood, I.Y.; Banasik, K.; Chen, Y.; Du, H.; Iona, A.; Mahajan, A.; et al. Evaluation of type 2 diabetes genetic risk variants in Chinese adults: Findings from 93,000 individuals from the China Kadoorie Biobank. Diabetologia 2016, 59, 1446–1457. [Google Scholar] [CrossRef] [PubMed]
- Kuo, J.Z.; Sheu, W.H.; Assimes, T.L.; Hung, Y.J.; Absher, D.; Chiu, Y.F.; Mak, J.; Wang, J.S.; Kwon, S.; Hsu, C.C.; et al. Trans-ethnic fine mapping identifies a novel independent locus at the 3′ end of CDKAL1 and novel variants of several susceptibility loci for type 2 diabetes in a Han Chinese population. Diabetologia 2013, 56, 2619–2628. [Google Scholar] [CrossRef] [PubMed]
- Tsai, F.J.; Yang, C.F.; Chen, C.C.; Chuang, L.M.; Lu, C.H.; Chang, C.T.; Wang, T.Y.; Chen, R.H.; Shiu, C.F.; Liu, Y.M.; et al. A genome-wide association study identifies susceptibility variants for type 2 diabetes in Han Chinese. PLoS Genet. 2010, 6, e1000847. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.O.; Long, J.; Cai, Q.; Qi, L.; Xiang, Y.B.; Cho, Y.S.; Tai, E.S.; Li, X.; Lin, X.; Chow, W.H.; et al. Identification of new genetic risk variants for type 2 diabetes. PLoS Genet. 2010, 6, e1001127. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Zhu, X.; Xu, M.; Guo, T.; Zhu, D.; Chen, G.; Li, X.; Xu, L.; Bi, Y.; Chen, Y.; et al. A genome-wide association study confirms previously reported loci for type 2 diabetes in Han Chinese. PLoS ONE 2011, 6, e22353. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gan, W.; Lu, L.; Dong, X.; Han, X.; Hu, C.; Yang, Z.; Sun, L.; Bao, W.; Li, P.; He, M.; et al. A genome-wide association study identifies GRK5 and RASGRP1 as type 2 diabetes loci in Chinese Hans. Diabetes 2013, 62, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Yin, X.; Li, S.; Jin, W.; Lou, H.; Yang, L.; Gong, X.; Wang, H.; Shen, Y.; Pan, X.; et al. Genomic dissection of population substructure of Han Chinese and its implication in association studies. Am. J. Hum. Genet. 2009, 85, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.C.; Hu, C.; Tam, C.H.; Zhang, R.; Kwan, P.; Leung, T.F.; Thomas, G.N.; Go, M.J.; Hara, K.; Sim, X.; et al. Genome-wide association study in a Chinese population identifies a susceptibility locus for type 2 diabetes at 7q32 near PAX4. Diabetologia 2013, 56, 1291–1305. [Google Scholar] [CrossRef]
- Jetten, A.M.; Kang, H.S.; Takeda, Y. Retinoic acid-related orphan receptors α and γ: Key regulators of lipid/glucose metabolism, inflammation, and insulin sensitivity. Front. Endocrinol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Variable | Cases (n = 427) | Controls (n = 408) | p |
|---|---|---|---|
| Age (years) | 57.37 ± 11.28 | 58.26 ± 10.51 | 0.062 b |
| Sex (Men/Women) | 219/208 | 209/199 | 0.075 b |
| BMI (kg/m2) c | 24.16 ± 2.25 | 23.28 ± 2.13 | 0.086 b |
| SBP (mmHg) c | 140.16 ± 18.21 | 124.41 ± 14.18 | 0.007 d |
| DBP (mmHg) c | 83.47 ± 8.68 | 77.63 ± 9.32 | 0.008 d |
| FPG (mmol/L) c | 9.77 ± 1.15 | 4.86 ± 0.58 | 0.001 d |
| TC (mmol/L) c | 5.11 ± 0.92 | 4.87 ± 0.88 | 0.009 d |
| HDL-C (mmol/L) c | 1.22 ± 0.33 | 1.38 ± 0.37 | 0.008 d |
| LDL-C (mmol/L) c | 2.77 ± 0.82 | 2.69 ± 0.91 | 0.073 b |
| TG (mmol/L) c | 1.72 ± 0.53 | 1.21 ± 0.34 | 0.003 d |
| SNP | Group | Genotype | HWE a | Allele | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Number | p | FDR q b | p | Freq. % | p | FDR q b | OR [95% CI] c | |||
| rs17270188 | AA/AG/GG | A | G | |||||||
| T2DM | 94/195/138 | 0.869 | 0.871 | 0.112 | 44.8 | 55.2 | 0.614 | 0.658 | 0.95 [0.78–1.15] | |
| Control | 93/190/125 | 0.204 | 46.1 | 53.9 | ||||||
| rs1898413 | AA/AG/GG | A | G | |||||||
| T2DM | 14/112/301 | 0.772 | 0.793 | 0.373 | 16.4 | 83.6 | 0.644 | 0.681 | 1.06 [0.82–1.38] | |
| Control | 10/107/291 | 0.965 | 15.6 | 84.4 | ||||||
| rs11638541 | TT/TC/CC | T | C | |||||||
| T2DM | 336/84/7 | 0.809 | 0.822 | 0.512 | 88.5 | 11.5 | 0.541 | 0.592 | 1.10 [0.81–1.50] | |
| Control | 327/76/5 | 0.806 | 89.5 | 10.5 | ||||||
| rs8033552 | AA/AG/GG | A | G | |||||||
| T2DM | 17/117/293 | 0.801 | 0.814 | 0.225 | 17.7 | 82.3 | 0.494 | 0.521 | 1.09 [0.85–1.41] | |
| Control | 14/106/288 | 0.280 | 16.4 | 83.6 | ||||||
| rs10851685 | TT/TA/AA | T | A | |||||||
| T2DM | 26/156/245 | 0.029 | 0.038 | 0.860 | 24.4 | 75.6 | 0.009 | 0.015 | 1.36 [1.08–1.72] | |
| Control | 13/130/265 | 0.540 | 19.1 | 80.9 | ||||||
| rs8041381 | AA/AG/GG | A | G | |||||||
| T2DM | 310/111/6 | 0.847 | 0.864 | 0.262 | 85.6 | 14.4 | 0.745 | 0.776 | 1.05 [0.80–1.38] | |
| Control | 299/105/4 | 0.113 | 86.2 | 13.8 | ||||||
| rs340002 | AA/AG/GG | A | G | |||||||
| T2DM | 51/204/172 | 0.804 | 0.818 | 0.421 | 35.8 | 64.2 | 0.586 | 0.601 | 1.06 [0.871–1.29] | |
| Control | 43/196/169 | 0.210 | 34.6 | 65.4 | ||||||
| rs340023 | CC/CT/TT | C | T | |||||||
| T2DM | 68/189/170 | 0.661 | 0.692 | 0.206 | 38.1 | 61.9 | 0.421 | 0.479 | 1.09 [0.89–1.32] | |
| Control | 56/183/169 | 0.566 | 36.2 | 63.8 | ||||||
| rs28724570 | CC/CT/TT | C | T | |||||||
| T2DM | 107/215/105 | 0.311 | 0.358 | 0.884 | 50.2 | 49.8 | 0.134 | 0.142 | 1.16 [0.96–1.40] | |
| Control | 90/200/118 | 0.762 | 46.6 | 53.4 | ||||||
| SNP | Genetic Model a | p Value | FDR q | OR [95% CI] | |
|---|---|---|---|---|---|
| rs17270181 | Dominant | (AG + AA) vs. GG | 0.183 | 0.231 | 1.32 [0.88–2.01] |
| rs1898413 | Additive | AG vs. GG | 0.918 | 0.921 | 1.01 [0.81–1.26] |
| AA vs. GG | 0.384 | 0.413 | 1.30 [0.72–2.34] | ||
| rs11638541 | Dominant | (AG + GG) vs. AA | 0.222 | 0.298 | 1.15 [0.92–1.44] |
| rs8033552 | Recessive | AA vs. (AG + GG) | 0.178 | 0.202 | 1.34 [0.87–2.05] |
| rs10851685 | Dominant | (TA + TT) vs. AA | 0.025 | 0.032 | 1.38 [1.04–1.82] |
| rs8041381 | Dominant | (AG + GG) vs. AA | 0.333 | 0.402 | 0.88 [0.58–1.33] |
| rs340002 | Dominant | (AG + AA) vs. GG | 0.699 | 0.748 | 1.04 [0.86–1.26] |
| rs340023 | Recessive | CC vs. (CT + TT) | 0.195 | 0.243 | 1.21 [0.91–1.62] |
| rs28724570 | Recessive | CC vs. (CT + TT) | 0.057 | 0.103 | 1.79 [0.98–3.23] |
| Haplotype a | Freq. (case) | Freq. | χ2 | Fisher’s p | OR [95% CI] b |
|---|---|---|---|---|---|
| (control) | |||||
| GGTGTAGTT | 0.044 | 0.031 | 1.03 | 0.417 | |
| GGTGTAGTT | 0.072 | 0.056 | 2.37 | 0.097 | |
| GGTGTAACT | 0.102 | 0.058 | 8.38 | 0.004 | 2.11 [1.31–3.42] |
| GGTGTAACC | 0.093 | 0.024 | 17.21 | 5.27 × 10−5 | 3.26[1.78–5.95] |
| GATATAGCT | 0.061 | 0.06 | 3.01 | 0.086 | |
| GATGAGGTT | 0.038 | 0.029 | 0.08 | 0.862 | |
| GATGAAGTT | 0.179 | 0.234 | 13.46 | 0.0001 | 0.54 [0.39–0.81] |
| AGTGAAGTT | 0.033 | 0.054 | 5.36 | 0.018 | 0.49 [0.29–0.91] |
| GATATAACT | 0.046 | 0 | 19.51 | 1.72 × 10−6 | |
| AATGAAATT | 0 | 0.034 | 18.79 | 1.16 × 10−5 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liu, Y.; Liu, Y.; Zhang, Y.; Su, Z. Genetic Variants of Retinoic Acid Receptor-Related Orphan Receptor Alpha Determine Susceptibility to Type 2 Diabetes Mellitus in Han Chinese. Genes 2016, 7, 54. https://doi.org/10.3390/genes7080054
Zhang Y, Liu Y, Liu Y, Zhang Y, Su Z. Genetic Variants of Retinoic Acid Receptor-Related Orphan Receptor Alpha Determine Susceptibility to Type 2 Diabetes Mellitus in Han Chinese. Genes. 2016; 7(8):54. https://doi.org/10.3390/genes7080054
Chicago/Turabian StyleZhang, Yuwei, Yulan Liu, Yin Liu, Yanjie Zhang, and Zhiguang Su. 2016. "Genetic Variants of Retinoic Acid Receptor-Related Orphan Receptor Alpha Determine Susceptibility to Type 2 Diabetes Mellitus in Han Chinese" Genes 7, no. 8: 54. https://doi.org/10.3390/genes7080054
APA StyleZhang, Y., Liu, Y., Liu, Y., Zhang, Y., & Su, Z. (2016). Genetic Variants of Retinoic Acid Receptor-Related Orphan Receptor Alpha Determine Susceptibility to Type 2 Diabetes Mellitus in Han Chinese. Genes, 7(8), 54. https://doi.org/10.3390/genes7080054