B Chromosomes of the Asian Seabass (Lates calcarifer) Contribute to Genome Variations at the Level of Individuals and Populations


 ,
, 
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Material and DNA Extraction
2.2. Estimation of DNA Content in Different Tissues by Flow Cytometry
2.3. Isolation and Sequencing of B Chromosomes
2.4. Fluorescence In Situ Hybridization (FISH) and Immunohistochemical Analysis
2.5. Annotation of B Chromosome-Derived Pseudo-Scaffolds
2.6. Data Availability
2.7. Population-Level Analyses of Asian Seabass B Chromosomes from Re-Sequencing Data
2.8. Estimation of the Density of Single Nucleotide Variants and Polymorphisms
2.9. Expression of Predicted B-Chromosomal Genes
3. Results
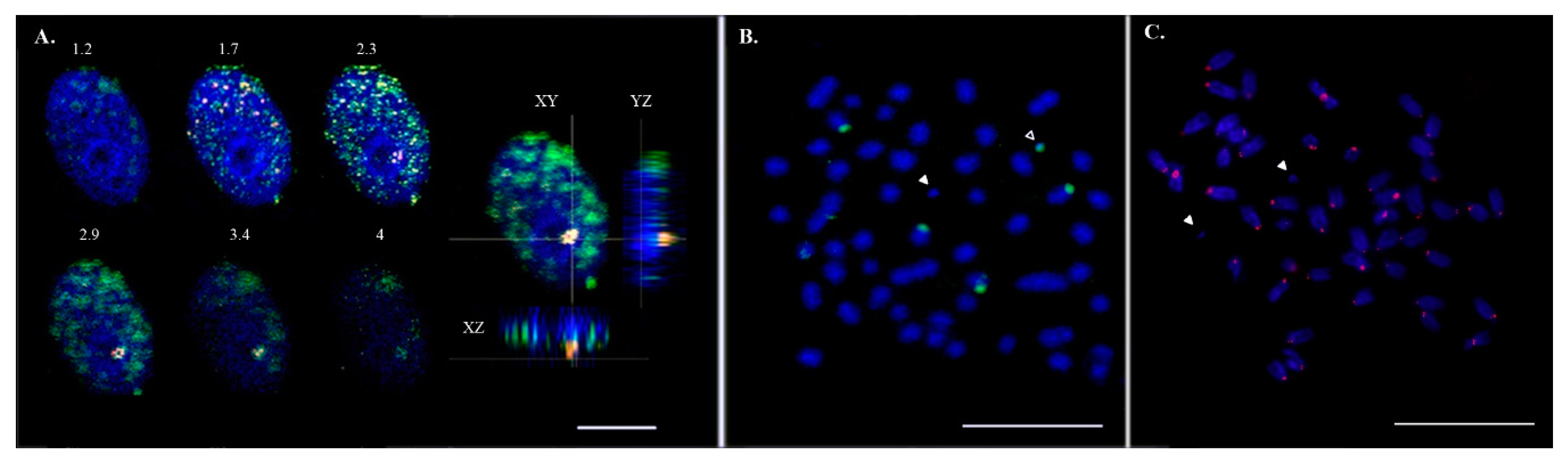
3.1. Asian Seabass B Chromosomes are Repeat-Rich and Associated with Nucleolus
3.2. B Chromosomal Repeats
3.3. B chromosomal Genes
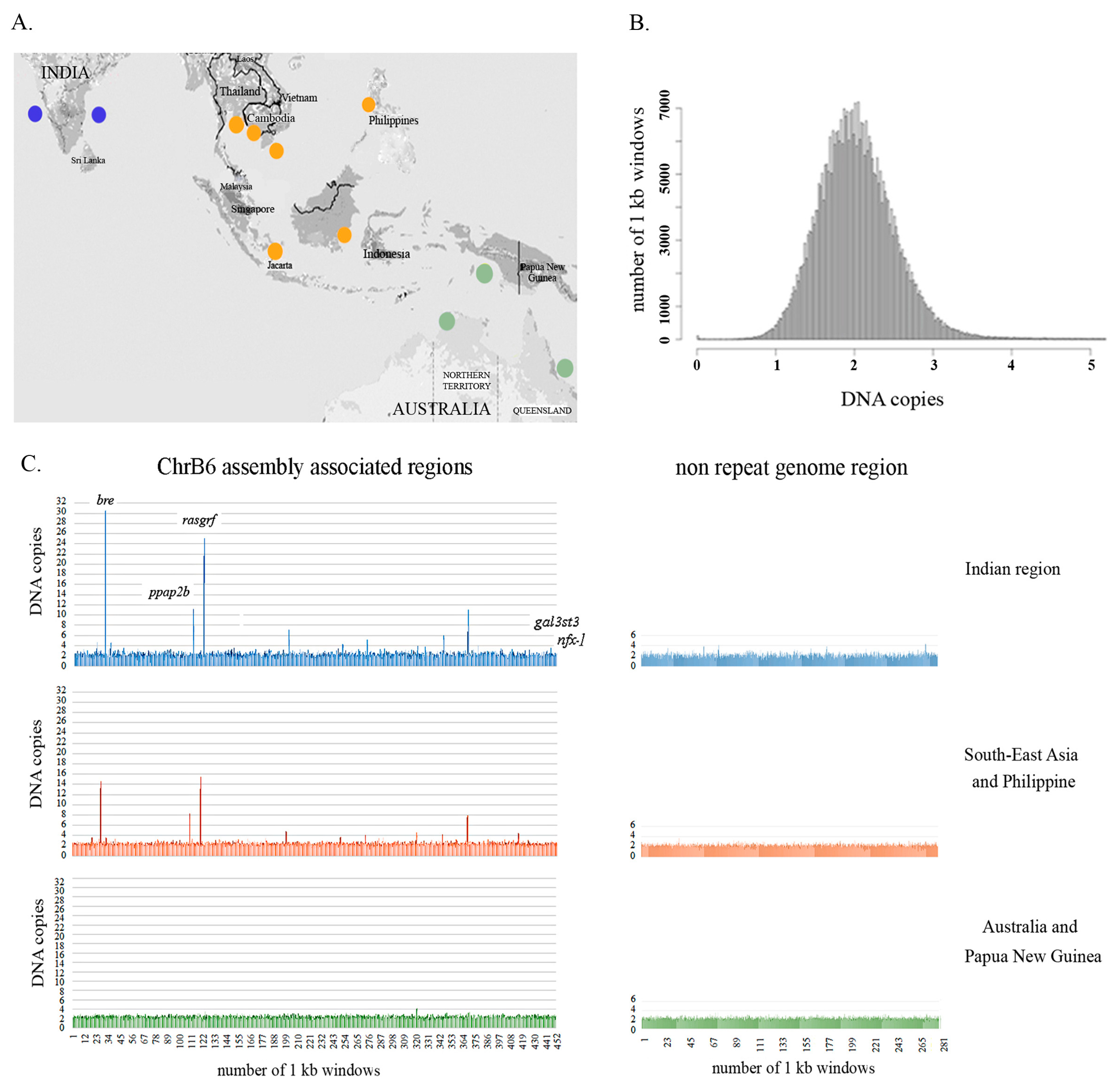
3.4. Population-Level Analysis of B Chromosome-Associated Regions Supports the Existence of an Asian Seabass Species Complex
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chou, R.; Lee, H.B. Commercial marine fish farming in Singapore. Aquacult. Res. 1997, 28, 767–776. [Google Scholar] [CrossRef]
- Frost, L.A.; Evans, B.S.; Jerry, D.R. Loss of genetic diversity due to hatchery culture practices in barramundi (Lates calcarifer). Aquaculture 2006, 261, 1056–1064. [Google Scholar] [CrossRef]
- Pethiyagoda, R.; Gill, A.C. Description of two new species of sea bass (Teleostei: Latidae: Lates) from Myanmar and Sri Lanka. Zootaxa 2012, 3314, 1–16. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- Vij, S.; Purushothaman, K.; Gopikrishna, G.; Lau, D.; Saju, J.M.; Shamsudheen, K.V.; Kumar, K.V.; Basheer, V.S.; Gopalakrishnan, A.; Hossain, M.S.; et al. Barcoding of Asian seabass across its geographic range provides evidence for its bifurcation into two distinct species. Front. Mar. Sci. 2014, 1, 30. [Google Scholar] [CrossRef]
- Vij, S.; Kuhl, H.; Kuznetsova, I.S.; Komissarov, A.; Yurchenko, A.A.; Van Heusden, P.; Singh, S.; Thevasagayam, N.M.; Prakki, S.R.S.; Purushothaman, K.; et al. Chromosomal-level assembly of the Asian seabass genome using long sequence reads and multi-layered scaffolding. PLoS Genet. 2016, 12, e1005954. [Google Scholar] [CrossRef] [Green Version]
- Russell, D.; Garrett, R. Early life history of barramundi, Lates calcarifer (Bloch), in north-eastern Queensland. Mar. Freshw. Res. 1985, 36, 191–201. [Google Scholar] [CrossRef]
- Moore, R. Spawning and early life history of barramundi, Lates calcarifer (Bloch), in Papua New Guinea. Mar. Freshw. Res. 1982, 33, 647–661. [Google Scholar] [CrossRef]
- Jerry, D.R. (Ed.) Biology and Culture of Asian Seabass; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2014; ISBN 978-1-4822-0807-8. [Google Scholar]
- Wang, L.; Wan, Z.Y.; Lim, H.S.; Yue, G.H. Genetic variability, local selection and demographic history: Genomic evidence of evolving towards allopatric speciation in Asian seabass. Mol. Ecol. 2016, 25, 3605–3621. [Google Scholar] [CrossRef] [PubMed]
- Zohar, Y.; Abraham, M.; Gordin, H. The gonadal cycle of the captivity-reared hermaphroditic teleost Sparus aurata (L.) during the first two years of life. Ann. Biol. Anim. Biochim. Biophys. 1978, 18, 877–882. [Google Scholar] [CrossRef]
- Sadovy, Y.; Shapiro, D.Y. Criteria for the diagnosis of hermaphroditism in fishes. Copeia 1987, 1987, 136. [Google Scholar] [CrossRef]
- Ravi, P.; Jiang, J.; Liew, W.C.; Orbán, L. Small-scale transcriptomics reveals differences among gonadal stages in Asian seabass (Lates calcarifer). Reprod. Biol. Endocrinol. 2014, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Guiguen, Y.; Cauty, C.; Fostier, A.; Fuchs, J.; Jalabert, B. Reproductive cycle and sex inversion of the seabass, Lates calcarifer, reared in sea cages in French Polynesia: Histological and morphometric description. Environ. Biol. Fishes 1994, 39, 231–247. [Google Scholar] [CrossRef]
- Camacho, J.P.M.; Schmid, M.; Cabrero, J. B chromosomes and sex in animals. Sex. Dev. 2011, 5, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Devlin, R.H.; Nagahama, Y. Sex determination and sex differentiation in fish: An overview of genetic, physiological, and environmental influences. Aquaculture 2002, 208, 191–364. [Google Scholar] [CrossRef]
- Houben, A. B Chromosomes—A matter of chromosome drive. Front. Plant Sci. 2017, 8, 210. [Google Scholar] [CrossRef] [PubMed]
- Makunin, A.I.; Dementyeva, P.V.; Graphodatsky, A.S.; Volobouev, V.T.; Kukekova, A.V.; Trifonov, V.A. Genes on B chromosomes of vertebrates. Mol. Cytogenet. 2014, 7, 99. [Google Scholar] [CrossRef] [PubMed]
- Puertas, M.J. Nature and evolution of B chromosomes in plants: A non-coding but information-rich part of plant genomes. Cytogenet. Genome Res. 2002, 96, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Valente, G.T.; Conte, M.A.; Fantinatti, B.E.A.; Cabral-de-Mello, D.C.; Carvalho, R.F.; Vicari, M.R.; Kocher, T.D.; Martins, C. Origin and evolution of B chromosomes in the cichlid fish Astatotilapia latifasciata based on integrated genomic analyses. Mol. Biol. Evol. 2014, 31, 2061–2072. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.; Fuchs, J.; Marques, A.; Schubert, V.; Soloviev, A.; Raskina, O.; Badaeva, E.; Houben, A. B Chromosomes of Aegilops speltoides are enriched in organelle genome-derived sequences. PLoS ONE 2014, 9, e90214. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.N.; Viegas, W.; Houben, A.A. Century of B chromosomes in plants: So what? Ann. Bot. 2008, 101, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Lamb, J.C.; Riddle, N.C.; Cheng, Y.-M.; Theuri, J.; Birchler, J.A. Localization and transcription of a retrotransposon-derived element on the maize B chromosome. Chromosom. Res. 2007, 15, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Montiel, E.E.; Cabrero, J.; Ruiz-Estévez, M.; Burke, W.D.; Eickbush, T.H.; Camacho, J.P.M.; López-León, M.D. Preferential occupancy of R2 retroelements on the B chromosomes of the grasshopper Eyprepocnemis plorans. PLoS ONE 2014, 9, e91820. [Google Scholar] [CrossRef] [PubMed]
- Montiel, E.E.; Cabrero, J.; Camacho, J.P.M.; López-León, M.D. Gypsy, RTE and Mariner transposable elements populate Eyprepocnemis plorans genome. Genetica 2012, 140, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, C.G.; Lamatsch, D.K.; Steinlein, C.; Engel, W.; Schartl, M.; Schmid, M. The giant B chromosome of the cyprinid fish Alburnus alburnus harbours a retrotransposon-derived repetitive DNA sequence. Chromosom. Res. 2003, 11, 23–35. [Google Scholar] [CrossRef]
- Makunin, A.I.; Kichigin, I.G.; Larkin, D.M.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Yang, F.; Trifonov, V.A. Contrasting origin of B chromosomes in two cervids (Siberian roe deer and grey brocket deer) unravelled by chromosome-specific DNA sequencing. BMC Genome 2016, 17, 618. [Google Scholar] [CrossRef] [PubMed]
- Makunin, A.I.; Rajičić, M.; Karamysheva, T.V.; Romanenko, S.A.; Druzhkova, A.S.; Blagojević, J.; Vujošević, M.; Rubtsov, N.B.; Graphodatsky, A.S.; Trifonov, V.A. Low-pass single-chromosome sequencing of human small supernumerary marker chromosomes (sSMCs) and Apodemus B chromosomes. Chromosoma 2018, 127, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, I.S.; Thevasagayam, N.M.; Sridatta, P.S.R.; Komissarov, A.S.; Saju, J.M.; Ngoh, S.Y.; Jiang, J.; Shen, X.; Orbán, L. Primary analysis of repeat elements of the Asian seabass (Lates calcarifer) transcriptome and genome. Front. Genet. 2014, 5, 223. [Google Scholar] [CrossRef] [PubMed]
- Heinlein, C.; Speidel, D. High-resolution cell cycle and DNA ploidy analysis in tissue samples. Curr. Protoc. Cytom. 2011. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.R.; Clarindo, W.R.; Praça, M.M.; Araújo, F.S.; Carels, N. Genome size, base composition and karyotype of Jatropha curcas L., an important biofuel plant. Plant Sci. 2008, 174, 613–617. [Google Scholar] [CrossRef]
- Dolezel, J.; Bartos, J.; Voglmayr, H.; Greilhuber, J. Nuclear DNA content and genome size of trout and human. Cytom. A 2003, 51, 127–128. [Google Scholar] [CrossRef]
- Mantovani, M.; Dos Santos Abel, L.D.; Moreira-Filho, O. Conserved 5S and variable 45S rDNA chromosomal localisation revealed by FISH in Astyanax scabripinnis (Pisces, Characidae). Genetica 2005, 123, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Howe, D.G.; Bradford, Y.M.; Conlin, T.; Eagle, A.E.; Fashena, D.; Frazer, K.; Knight, J.; Mani, P.; Martin, R.; Moxon, S.A.T.; et al. ZFIN, the zebrafish model organism database: Increased support for mutants and transgenics. Nucleic Acids Res. 2013, 41, D854–D860. [Google Scholar] [CrossRef] [PubMed]
- Alkan, C.; Kidd, J.M.; Marques-Bonet, T.; Aksay, G.; Antonacci, F.; Hormozdiari, F.; Kitzman, J.O.; Baker, C.; Malig, M.; Mutlu, O.; et al. Personalized copy number and segmental duplication maps using next-generation sequencing. Nat. Genet. 2009, 41, 1061–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Ortí, G.; Zhang, G.; Lu, G. A practical approach to phylogenomics: The phylogeny of ray-finned fish (Actinopterygii) as a case study. BMC Evol. Biol. 2007, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Platt, A.; Gugger, P.F.; Pellegrini, M.; Sork, V.L. Genome-wide signature of local adaptation linked to variable CpG methylation in oak populations. Mol. Ecol. 2015, 24, 3823–3830. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J. Functional Genomic Analysis of Gonad Development in the Protandrous Asian Seabass. Ph.D. Thesis, National University of Singapore, Singapore, 2014. [Google Scholar]
- Wang, C.M.; Bai, Z.Y.; He, X.P.; Lin, G.; Xia, J.H.; Sun, F.; Lo, L.C.; Feng, F.; Zhu, Z.Y.; Yue, G.H. A high-resolution linkage map for comparative genome analysis and QTL fine mapping in Asian seabass, Lates calcarifer. BMC Genom. 2011, 12, 174. [Google Scholar] [CrossRef] [PubMed]
- Chumová, Z.; Mandáková, T.; Trávníček, P. Are B-chromosomes responsible for the extraordinary genome size variation in selected Anthoxanthum annuals? Plant Syst. Evol. 2016, 302, 731–738. [Google Scholar] [CrossRef]
- Fantinatti, B.E.A.; Mazzuchelli, J.; Valente, G.T.; Cabral-de-Mello, D.C.; Martins, C. Genomic content and new insights on the origin of the B chromosome of the cichlid fish Astatotilapia latifasciata. Genetica 2011, 139, 1273–1282. [Google Scholar] [CrossRef] [PubMed]
- Shirak, A.; Grabherr, M.; Di Palma, F.; Lindblad-Toh, K.; Hulata, G.; Ron, M.; Kocher, T.D.; Seroussi, E. Identification of repetitive elements in the genome of Oreochromis niloticus: Tilapia repeat masker. Mar. Biotechnol. 2010, 12, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Abdurashitov, M.A.; Gonchar, D.A.; Chernukhin, V.A.; Tomilov, V.N.; Tomilova, J.E.; Schostak, N.G.; Zatsepina, O.G.; Zelentsova, E.S.; Evgen’ev, M.B.; Degtyarev, S. Medium-sized tandem repeats represent an abundant component of the Drosophila virilis genome. BMC Genom. 2013, 14, 771. [Google Scholar] [CrossRef] [PubMed]
- López-Flores, I.; Garrido-Ramos, M.A. The Repetitive DNA Content of Eukaryotic Genomes. Genome Dyn. 2012, 7, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Gornung, E. Twenty years of physical mapping of major ribosomal RNA genes across the teleosts: A review of research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, V.A.; Dementyeva, P.V.; Larkin, D.M.; O’Brien, P.C.M.; Perelman, P.L.; Yang, F.; Ferguson-Smith, M.A.; Graphodatsky, A.S. Transcription of a protein-coding gene on B chromosomes of the Siberian roe deer (Capreolus pygargus). BMC Biol. 2013, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Leach, C.R.; Houben, A.; Field, B.; Pistrick, K.; Demidov, D.; Timmis, J.N. Molecular evidence for transcription of genes on a B chromosome in Crepis capillaris. Genetics 2005, 171, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Y.; Liu, X.-L.; Ding, M.; Li, Z.; Zhou, L.; Zhang, X.-J.; Gui, J.-F. A novel male-specific SET domain-containing gene setdm identified from extra microchromosomes of gibel carp males. Sci. Bull. 2017, 62, 528–536. [Google Scholar] [CrossRef]
- Nakai, Y.; Kubota, S.; Kohno, S. Chromatin diminution and chromosome elimination in four Japanese hagfish species. Cytogenet. Cell Genet. 1991, 56, 196–198. [Google Scholar] [CrossRef] [PubMed]
- Kloc, M.; Zagrodzinska, B. Chromatin elimination—An oddity or a common mechanism in differentiation and development? Differentiation 2001, 68, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Henning, F.; Trifonov, V.; Ferguson-Smith, M.A.; de Almeida-Toledo, L.F. Non-homologous sex chromosomes in two species of the genus Eigenmannia (Teleostei: Gymnotiformes). Cytogenet. Genome Res. 2008, 121, 55–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, D.S.; Milhomem, S.S.; Pieczarka, J.C.; Nagamachi, C.Y. Cytogenetic studies in Eigenmannia virescens (Sternopygidae, Gymnotiformes) and new inferences on the origin of sex chromosomes in the Eigenmannia genus. BMC Genet. 2009, 10, 74. [Google Scholar] [CrossRef] [PubMed]
- Lamatsch, D.K.; Nanda, I.; Epplen, J.T.; Schmid, M.; Schartl, M. Unusual triploid males in a microchromosome-carrying clone of the Amazon molly, Poecilia formosa. Cytogenet. Cell Genet. 2000, 91, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Terai, Y.; Mizoiri, S.; Aibara, M.; Nishihara, H.; Watanabe, M.; Kuroiwa, A.; Hirai, H.; Hirai, Y.; Matsuda, Y.; et al. B chromosomes have a functional effect on female sex determination in Lake Victoria cichlid fishes. PLoS Genet. 2011, 7, e1002203. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komissarov, A.; Vij, S.; Yurchenko, A.; Trifonov, V.; Thevasagayam, N.; Saju, J.; Sridatta, P.S.R.; Purushothaman, K.; Graphodatsky, A.; Orbán, L.; et al. B Chromosomes of the Asian Seabass (Lates calcarifer) Contribute to Genome Variations at the Level of Individuals and Populations. Genes 2018, 9, 464. https://doi.org/10.3390/genes9100464
Komissarov A, Vij S, Yurchenko A, Trifonov V, Thevasagayam N, Saju J, Sridatta PSR, Purushothaman K, Graphodatsky A, Orbán L, et al. B Chromosomes of the Asian Seabass (Lates calcarifer) Contribute to Genome Variations at the Level of Individuals and Populations. Genes. 2018; 9(10):464. https://doi.org/10.3390/genes9100464
Chicago/Turabian StyleKomissarov, Aleksey, Shubha Vij, Andrey Yurchenko, Vladimir Trifonov, Natascha Thevasagayam, Jolly Saju, Prakki Sai Rama Sridatta, Kathiresan Purushothaman, Alexander Graphodatsky, László Orbán, and et al. 2018. "B Chromosomes of the Asian Seabass (Lates calcarifer) Contribute to Genome Variations at the Level of Individuals and Populations" Genes 9, no. 10: 464. https://doi.org/10.3390/genes9100464
APA StyleKomissarov, A., Vij, S., Yurchenko, A., Trifonov, V., Thevasagayam, N., Saju, J., Sridatta, P. S. R., Purushothaman, K., Graphodatsky, A., Orbán, L., & Kuznetsova, I. (2018). B Chromosomes of the Asian Seabass (Lates calcarifer) Contribute to Genome Variations at the Level of Individuals and Populations. Genes, 9(10), 464. https://doi.org/10.3390/genes9100464