UBE2C Is a Transcriptional Target of the Cell Cycle Regulator FOXM1

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Ethics
2.3. RNA Extraction, Reverse Transcription and qRT-PCR
2.4. Cell Line and Transfections
2.5. Chromatin Immunoprecipitation (ChIP) Assay
2.6. UBE2C Promoter Cloning and Dual-Luciferase Assay
2.7. Protein Extraction and Western Blot
2.8. In Silico Analysis
2.9. Statistical Analyses
3. Results
3.1. FOXM1 and UBE2C Are Co-Expressed in Healthy Tissues
3.2. FOXM1 and UBE2C Co-Overexpression Is a Common Event in Cancer, Including ESCC
3.3. UBE2C Figures as a Potential FOXM1 Transcriptional Target
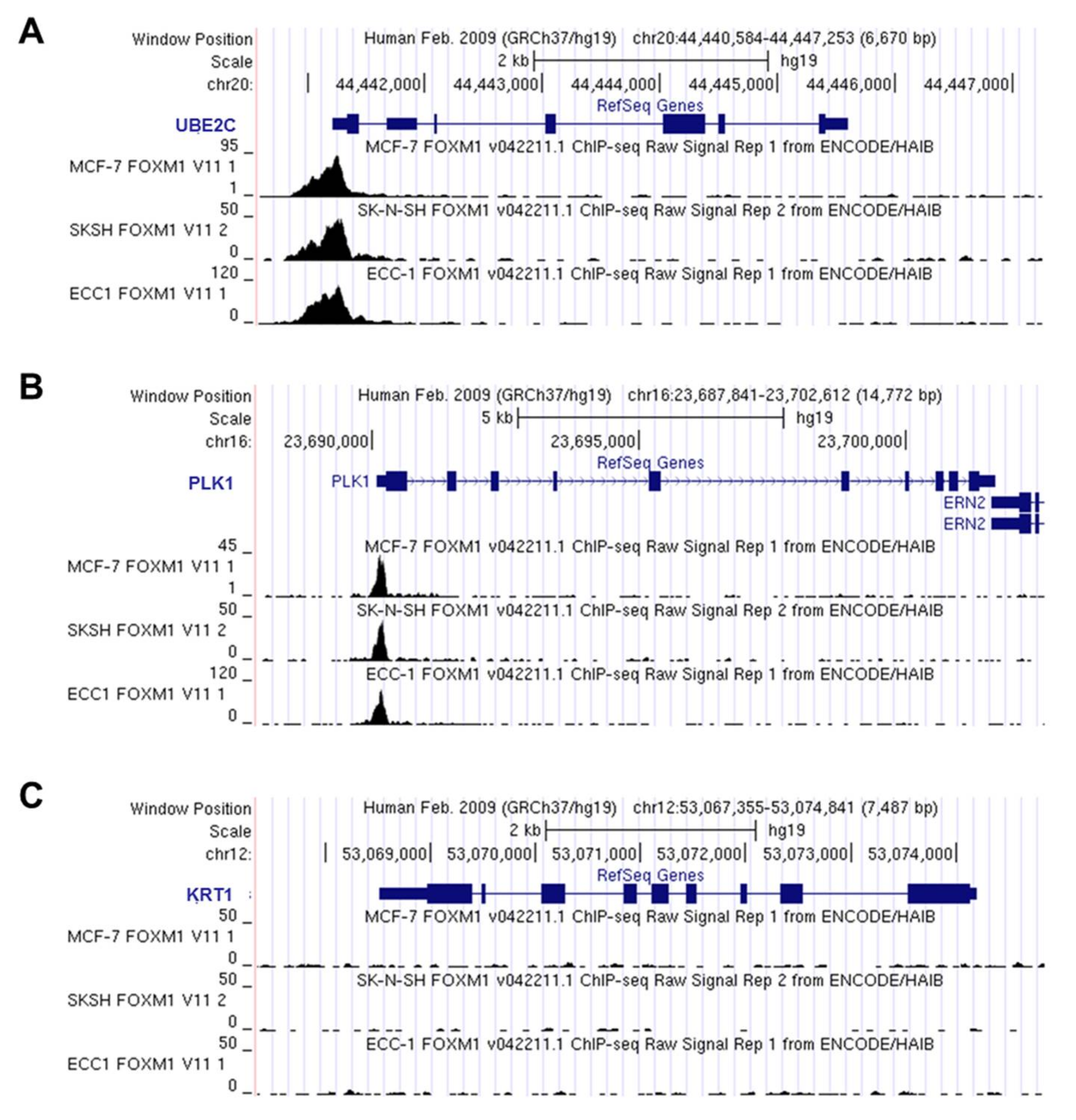
3.4. FOXM1 Binds onto UBE2C Promoter and Transcriptionally Activates It in an ESCC-Derived Cell Line
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Calnan, D.R.; Brunet, A. The FoxO code. Oncogene 2008, 27, 2276–2288. [Google Scholar] [CrossRef] [PubMed]
- Myatt, S.S.; Lam, E.W. Targeting FOXM1. Nat. Rev. Cancer 2008, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- Pilarsky, C.; Wenzig, M.; Specht, T.; Saeger, H.D.; Grützmann, R. Identification and validation of commonly overexpressed genes in solid tumors by comparison of microarray data. Neoplasia 2004, 6, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Uddin, S.; Ahmed, M.; Hussain, A.; Abubaker, J.; Al-Sanea, N.; AbdulJabbar, A.; Ashari, L.H.; Alhomoud, S.; Al-Dayel, F.; Jehan, Z.; et al. Genome-wide expression analysis of middle eastern colorectal cancer reveals FOXM1 as a novel target for cancer therapy. Am. J. Pathol. 2011, 178, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Hui, M.K.; Chan, K.W.; Luk, J.M.; Lee, N.P.; Chung, Y.; Cheung, L.C.; Srivastava, G.; Tsao, S.W.; Tang, J.C.; Law, S. Cytoplasmic forkhead box M1 (FOXM1) in esophageal squamous cell carcinoma significantly correlates with pathological disease stage. World J. Surg. 2012, 36, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takata, A.; Takiguchi, S.; Okada, K.; Takahashi, T.; Kurokawa, Y.; Yamasaki, M.; Miyata, H.; Nakajima, K.; Mori, M.; Doki, Y. Clinicopathological and prognostic significance of FOXM1 expression in esophageal squamous cell carcinoma. Anticancer Res. 2014, 34, 2427–2432. [Google Scholar] [PubMed]
- Koo, C.Y.; Muir, K.W.; Lam, E.W. Foxm1: From cancer initiation to progression and treatment. Biochim. Biophys. Acta 2012, 1819, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Townsley, F.M.; Aristarkhov, A.; Beck, S.; Hershko, A.; Ruderman, J.V. Dominant-negative cyclin-selective ubiquitin carrier protein E2-C/UbcH10 blocks cells in metaphase. Proc. Natl. Acad. Sci. USA 1997, 94, 2362–2367. [Google Scholar] [CrossRef] [PubMed]
- Stegmeier, F.; Rape, M.; Draviam, V.M.; Nalepa, G.; Sowa, M.E.; Ang, X.L.; McDonald, E.R.; Li, M.Z.; Hannon, G.J.; Sorger, P.K.; et al. Anaphase initiation is regulated by antagonistic ubiquitination and deubiquitination activities. Nature 2007, 446, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, A.; Da Costa, N.M.; De Martino, M.; Sepe, R.; Pellecchia, S.; de Sousa, V.P.; Nicolau Neto, P.; Kruel, C.D.; Bergman, A.; Nasciutti, L.E.; et al. UBE2C is overexpressed in ESCC tissues and its abrogation attenuates the malignant phenotype of ESCC cell lines. Oncotarget 2016, 7, 65876–65887. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, E.F.; Chen, X.; Han, N.; Webber, A.; Ji, Z.; Sharrocks, A.D.; Ang, Y.S. Deregulation of the FOXM1 target gene network and its coregulatory partners in oesophageal adenocarcinoma. Mol. Cancer 2015, 14, 69. [Google Scholar] [CrossRef] [PubMed]
- Nestal de Moraes, G.; Delbue, D.; Silva, K.L.; Robaina, M.C.; Khongkow, P.; Gomes, A.R.; Zona, S.; Crocamo, S.; Mencalha, A.L.; Magalhães, L.M.; et al. FOXM1 targets XIAP and Survivin to modulate breast cancer survival and chemoresistance. Cell. Signal. 2015, 27, 2496–2505. [Google Scholar] [CrossRef] [PubMed]
- Mathelier, A.; Fornes, O.; Arenillas, D.J.; Chen, CY.; Denay, G.; Lee, J.; Shi, W.; Shyr, C.; Tan, G.; Worsley-Hunt, R.; et al. JASPAR 2016: A major expansion and update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2016, 4, D110–D115. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 6, 57–74. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, A.; Kalinichenko, V.V.; Yutzey, K.E. FoxO1 and FOXM1 transcription factors have antagonistic functions in neonatal cardiomyocyte cell-cycle withdrawal and IGF1 gene regulation. Circ. Res. 2013, 112, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Lam, E.W. Role of the forkhead transcription factor FoxO-FOXM1 axis in cancer and drug resistance. Front. Med. 2012, 6, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Lam, E.W.; Brosens, J.J.; Gomes, A.R.; Koo, C.Y. Forkhead box proteins: Tuning forks for transcriptional harmony. Nat. Rev. Cancer 2013, 13, 482–495. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Zhang, H.; Cowell, J. Ubiquitin-conjugating enzyme UBE2C: Molecular biology, role in tumorigenesis, and potential as a biomarker. Tumour. Biol. 2012, 33, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, C.; Rorick, A.; Wu, D.; Chiu, M.; Thomas-Ahner, J.; Chen, Z.; Chen, H.; Clinton, S.K.; Chan, K.K.; et al. CCI-779 inhibits cell-cycle G2-M progression and invasion of castration-resistant prostate cancer via attenuation of UBE2C transcription and mRNA stability. Cancer Res. 2011, 71, 4866–4876. [Google Scholar] [CrossRef] [PubMed]
- Cunha, I.W.; Carvalho, K.C.; Martins, W.K.; Marques, S.M.; Muto, N.H.; Falzoni, R.; Rocha, R.M.; Aguiar, S.; Simoes, A.C.; Fahham, L.; et al. Identification of genes associated with local aggressiveness and metastatic behavior in soft tissue tumors. Transl. Oncol. 2010, 3, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Jiang, X.; Zeng, C.; Zheng, S.; Luo, B.; Zeng, Y.; Ding, R.; Jiang, H.; He, Q.; Guo, J.; et al. High expression of ubiquitin-conjugating enzyme 2c (UBE2C) correlates with nasopharyngeal carcinoma progression. BMC Cancer 2013, 13, 192. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wen, M.; Huang, Y.; He, X.; Wang, Y.; Wu, Q.; Li, Z.; Castellanos-Martin, A.; Abad, M.; Cruz-Hernandez, J.J.; et al. C2ORF40 suppresses breast cancer cell proliferation and invasion through modulating expression of M phase cell cycle genes. Epigenetics 2013, 8, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Ikeda, H.; Taira, N.; Hatoh, S.; Naito, M.; Doihara, H. Overexpression of UbcH10 alternates the cell cycle profile and accelerate the tumor proliferation in colon cancer. BMC Cancer 2009, 9, 87. [Google Scholar] [CrossRef] [PubMed]
- Delpuech, O.; Griffiths, B.; East, P.; Essafi, A.; Lam, E.W.; Burgering, B.; Downward, J.; Schulze, A. Induction of Mxi1-SRα by FoxO3a contributes to repression of Myc-dependent gene expression. Mol. Cell. Biol. 2007, 27, 4917–4930. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Ding, Z.; Huang, N.; Huang, Z.; Zhang, N.; Xia, Z. Forkhead box m1 positively regulates UBE2C and protects glioma cells from autophagic death. Cell Cycle 2017, 16, 1705–1718. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.; Dumon, S.; Ward, C.; Jäger, R.; Freeman, S.; Dawood, B.; Sheriff, L.; Lorvellec, M.; Kralovics, R.; Frampton, J.; et al. MYBL2 haploinsufficiency increases susceptibility to age-related haematopoietic neoplasia. Leukemia 2013, 27, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, S.; Alam, S.K.; Roy, K.S.; Datta, A.; Nath, S.; Roychoudhury, S. E2 ubiquitin-conjugating enzyme, UBE2C gene, is reciprocally regulated by wild-type and gain-of-function mutant p53. J. Biol. Chem. 2016, 291, 14231–14247. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Sprenger, C.; Wu, P.J.; Sun, S.; Uo, T.; Haugk, K.; Epilepsia, K.S.; Plymate, S. MED1 mediates androgen receptor splice variant induced gene expression in the absence of ligand. Oncotarget 2015, 6, 288–304. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Ishii, Y.; Nishida, Y.; Ikarashi, M.; Nagata, T.; Nakamura, T.; Yamamori, S.; Asai, S. Detection of aberrations of ubiquitin-conjugating enzyme E2C gene (UBE2C) in advanced colon cancer with liver metastases by DNA microarray and two-color fish. Cancer Genet. Cytogenet. 2006, 168, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Au, A.Y.; Foukakis, T.; Barbaro, M.; Kiss, N.; Clifton-Bligh, R.; Staaf, J.; Borg, A.; Delbridge, L.; Robinson, B.G.; et al. Array-CGH identifies cyclin D1 and UBCH10 amplicons in anaplastic thyroid carcinoma. Endocr. Relat. Cancer 2008, 15, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Tzelepi, V.; Zhang, J.; Lu, J.F.; Kleb, B.; Wu, G.; Wan, X.; Hoang, A.; Efstathiou, E.; Sircar, K.; Navone, N.M.; et al. Modeling a lethal prostate cancer variant with small-cell carcinoma features. Clin. Cancer Res. 2012, 18, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Sakai, N.; Kajiyama, Y.; Iwanuma, Y.; Tomita, N.; Amano, T.; Isayama, F.; Ouchi, K.; Tsurumaru, M. Study of abnormal chromosome regions in esophageal squamous cell carcinoma by comparative genomic hybridization: Relationship of lymph node metastasis and distant metastasis to selected abnormal regions. Dis. Esophagus 2010, 23, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.Y.; Zhu, Z.M.; Chen, L.B.; Wang, J.H.; Su, Q.S.; Yang, J.R.; Lin, Y.; Xue, L.J.; Liu, X.B.; Mo, X.B. FOXM1 expression correlates with tumor invasion and a poor prognosis of colorectal cancer. Acta Histochem. 2012, 114, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Bellelli, R.; Castellone, M.D.; Garcia-Rostan, G.; Ugolini, C.; Nucera, C.; Sadow, P.M.; Nappi, T.C.; Salerno, P.; Cantisani, M.C.; Basolo, F.; et al. FOXM1 is a molecular determinant of the mitogenic and invasive phenotype of anaplastic thyroid carcinoma. Endocr. Relat. Cancer 2012, 19, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Balli, D.; Ustiyan, V.; Fulford, L.; Hiller, A.; Misetic, V.; Zhang, Y.; Paluch, A.M.; Waltz, S.E.; Kasper, S.; et al. FOXM1 expression in prostate epithelial cells is essential for prostate carcinogenesis. J. Biol. Chem. 2013, 288, 22527–22541. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, P.; Chen, H. Overexpression of FOXM1 is associated with metastases of nasopharyngeal carcinoma. Ups. J. Med. Sci. 2014, 119, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.E.; Myatt, S.S.; Krol, J.; Hartman, J.; Peck, B.; McGovern, U.B.; Wang, J.; Guest, S.K.; Filipovic, A.; Gojis, O.; et al. FOXM1 is a downstream target and marker of HER2 overexpression in breast cancer. Int. J. Oncol. 2009, 35, 57–68. [Google Scholar] [PubMed]
- Berlingieri, M.T.; Pallante, P.; Guida, M.; Nappi, C.; Masciullo, V.; Scambia, G.; Ferraro, A.; Leone, V.; Sboner, A.; Barbareschi, M.; et al. UbcH10 expression may be a useful tool in the prognosis of ovarian carcinomas. Oncogene 2007, 26, 2136–2140. [Google Scholar] [CrossRef] [PubMed]
- Millour, J.; de Olano, N.; Horimoto, Y.; Monteiro, L.J.; Langer, J.K.; Aligue, R.; Hajji, N.; Lam, E.W. ATM and p53 regulate FOXM1 expression via E2F in breast cancer epirubicin treatment and resistance. Mol. Cancer Ther. 2011, 10, 1046–1058. [Google Scholar] [CrossRef] [PubMed]
- Olivier, M.; Eeles, R.; Hollstein, M.; Khan, M.A.; Harris, C.C.; Hainaut, P. The IARC TP53 database: New online mutation analysis and recommendations to users. Hum. Mutat. 2002, 19, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network; Analysis Working Group: Asan University; BC Cancer Agency; Brigham and Women’s Hospital; Broad Institute; Brown University; Case Western Reserve University; Dana-Farber Cancer Institute; Duke University; Greater Poland Cancer Centre; et al. Integrated genomic characterization of oesophageal carcinoma. Nature 2017, 541, 169–175. [Google Scholar] [CrossRef]
- Mandard, A.M.; Hainaut, P.; Hollstein, M. Genetic steps in the development of squamous cell carcinoma of the esophagus. Mutat. Res. 2000, 462, 335–342. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor | Number of Samples | Spearman r | p Value |
|---|---|---|---|
| Acute myeloid Leukemia | 197 | 0.7 | <0.0001 |
| Adrenocortical carcinoma | 92 | 0.87 | <0.0001 |
| Bladder urothelial carcinoma | 413 | 0.63 | <0.0001 |
| Brain low grade glioma | 530 | 0.94 | <0.0001 |
| Breast cancer invasive | 1100 | 0.85 | <0.0001 |
| Cholangiocarcinoma | 35 | 0.79 | <0.0001 |
| Colorectal adenocarcinoma | 382 | 0.47 | <0.0001 |
| Glioblastoma multiforme | 166 | 0.71 | <0.0001 |
| Hepatocellular carcinoma | 373 | 0.82 | <0.0001 |
| Head and Neck Squamous cell carcinoma | 522 | 0.46 | <0.0001 |
| Kidney renal clear cell carcinoma | 534 | 0.84 | <0.0001 |
| Lung adenocarcinoma | 517 | 0.78 | <0.0001 |
| Lung squamous cell carcinoma | 177 | 0.38 | <0.0001 |
| Lymphoid neoplasm diffuse large B-cell lymphoma | 28 | 0.29 * | 0.0389 * |
| Mesothelioma | 86 | 0.83 | <0.0001 |
| Ovarian serous cystadenocarcinoma | 307 | 0.58 | <0.0001 |
| Pancreatic adenocarcinoma | 179 | 0.81 | <0.0001 |
| Pheochromocytoma and paraganglioma | 184 | 0.8 | <0.0001 |
| Sarcoma | 262 | 0.77 | <0.0001 |
| Skin cutaneous melanoma | 471 | 0.74 | <0.0001 |
| Stomach adenocarcinoma | 415 | 0.6 | <0.0001 |
| Testicular germ cell cancer | 156 | 0.62 | <0.0001 |
| Thymoma | 120 | 0.77 | <0.0001 |
| Thyroid carcinoma | 509 | 0.79 | <0.0001 |
| Uterine corpus endometrial carcinoma | 177 | 0.52 | <0.0001 |
| Uveal melanoma | 80 | 0.9 | <0.0001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicolau-Neto, P.; Palumbo, A.; De Martino, M.; Esposito, F.; De Almeida Simão, T.; Fusco, A.; Nasciutti, L.E.; Meireles Da Costa, N.; Ribeiro Pinto, L.F. UBE2C Is a Transcriptional Target of the Cell Cycle Regulator FOXM1. Genes 2018, 9, 188. https://doi.org/10.3390/genes9040188
Nicolau-Neto P, Palumbo A, De Martino M, Esposito F, De Almeida Simão T, Fusco A, Nasciutti LE, Meireles Da Costa N, Ribeiro Pinto LF. UBE2C Is a Transcriptional Target of the Cell Cycle Regulator FOXM1. Genes. 2018; 9(4):188. https://doi.org/10.3390/genes9040188
Chicago/Turabian StyleNicolau-Neto, Pedro, Antonio Palumbo, Marco De Martino, Francesco Esposito, Tatiana De Almeida Simão, Alfredo Fusco, Luiz Eurico Nasciutti, Nathalia Meireles Da Costa, and Luis Felipe Ribeiro Pinto. 2018. "UBE2C Is a Transcriptional Target of the Cell Cycle Regulator FOXM1" Genes 9, no. 4: 188. https://doi.org/10.3390/genes9040188