Portrait of Matrix Gene Expression in Candida glabrata Biofilms with Stress Induced by Different Drugs


Laboratório de Investigação em Biofilmes Rosário Oliveira (LIBRO), Centre of Biological Engineering, University of Minho, 4710-057 Braga, Portugal
*
Author to whom correspondence should be addressed.
Genes 2018, 9(4), 205; https://doi.org/10.3390/genes9040205
Submission received: 6 March 2018
/
Revised: 29 March 2018
/
Accepted: 5 April 2018
/
Published: 10 April 2018
(This article belongs to the Special Issue Fungal Pathogenesis in Humans: The Growing Threat)
Abstract
:(1) Background: Candida glabrata is one of the most significant Candida species associated with severe cases of candidiasis. Biofilm formation is an important feature, closely associated with antifungal resistance, involving alterations of gene expression or mutations, which can result in the failure of antifungal treatments. Hence, the main goal of this work was to evaluate the role of a set of genes, associated with matrix production, in the resistance of C. glabrata biofilms to antifungal drugs. (2) Methods: the determination of the expression of BGL2, XOG1, FKS1, FKS2, GAS2, KNH1, UGP1, and MNN2 genes in 48-h biofilm’s cells of three C. glabrata strains was performed through quantitative real-time PCR (RT-qPCR), after contact with Fluconazole (Flu), Amphotericin B (AmB), Caspofungin (Csf), or Micafungin (Mcf). (3) Results: Mcf induced a general overexpression of the selected genes. It was verified that the genes related to the production of β-1,3-glucans (BGL2, XOG1, GAS2) had the highest expressions. (4) Conclusion: though β-1,6-glucans and mannans are an essential part of the cell and biofilm matrix, C. glabrata biofilm cells seem to contribute more to the replacement of β-1,3-glucans. Thus, these biopolymers seem to have a greater impact on the biofilm matrix composition and, consequently, a role in the biofilm resistance to antifungal drugs.
1. Introduction
Fungal infections continue to increase worldwide, particularly among immunosuppressed patients, individuals under prolonged hospitalization, catheterization, or continued antimicrobial treatments [1,2,3]. Candida spp. are the commonest fungal species involved in these diseases. Candida albicans is the most isolated species, but Candida glabrata and Candida parapsilosis are the second most isolated species in the United States of America and Europe, respectively [1,4,5]. Though C. glabrata does not have the capacity to form hyphae and pseudohyphae or to secret proteases, this species has other virulence factors, such as the ability to secrete phospholipases, lipases, and haemolysins and, importantly, the capacity to form biofilms [6,7,8]. These factors highly contribute to a high aggressiveness, resulting in a low therapeutic response and severe cases of recurrent candidiasis [8,9]. Biofilms are communities of microorganisms that colonize tissues and indwelling medical devices, embedded in an extracellular matrix [10,11]. These heterogeneous structures provide high resistance to antifungal therapy and strong host immune responses [7,8,12]. C. glabrata has shown to form a compact biofilm structure in different multilayers [6,7], with proteins, carbohydrates, and ergosterol into their matrices [6,7,13].
Various reports have shown the presence of β-1,3 glucans in the biofilm matrices of C. albicans [14,15,16,17]. Interestingly, it has been demonstrated that an increase in cell wall glucan was associated with biofilm growth [14] and, more recently, β-1,3 glucans were shown to be also present in the matrices of C. glabrata biofilms [13,18,19]. This specific carbohydrate has been associated with a general increase of extracellular matrix delivery, which is critical for securing biofilm cells to a surface and crucial to develop an antifungal drug resistance phenotype [14,19,20,21,22,23]. Several genes are involved in the delivery and accumulation of extracellular matrix. It is recognized that, in C. albicans, the major β-1,3 glucan synthases are encoded mainly by FKS1 but also by FKS2 [24]. The BGL2 and XOG1 genes also have important roles in glucan matrix delivery by encoding glucanosyltransferases and β-1,3 exoglucanase, respectively [25,26]. These genes play an important part in cell wall remodeling, however, the influence of the corresponding enzymes in matrix glucan delivery does not appear to affect cell wall ultrastructure or β-1,3 glucan concentration, suggesting that these enzymes function specifically for matrix delivery [17,19,26,27,28]. Identical to Saccharomyces cerevisiae, in C. glabrata, the GAS gene family is a regulator of the production of β-1,3 glucan [29]. Gas2, a glycosylphosphatidylinositol (GPI)-anchored cell surface protein [30,31], is a putative carbohydrate-active enzyme that may change cell wall polysaccharides [29,32].
Another carbohydrate of C. glabrata cell wall is β-1,6-glucan, present as a polymer covalently attached to glycoproteins [33,34,35,36], β-1,3-glucan, and chitin [37]. Nagahashi et al. [36] reported the isolation of KNH1 homologs (genes encoding cell surface O-glycoproteins), suggesting the evolutionary conservation of these molecules as essential components of β-1,6-glucan synthesis in C. glabrata, which was also discussed before [35,38]. Additionally, the UGP1 gene is a putative uridine diphosphate (UDP)-glucose pyrophosphorylase related to the general β-1,6-d-glucan biosynthetic process [39,40]. During stress conditions, several S. cerevisiae orthologous genes are induced in C. glabrata. In glucose starvation stress, UGP1 is induced [39].
The external layer cell wall of Candida spp. also consists of highly glycosylated mannoproteins [41,42,43], which play a major role in host recognition, adhesion, and cell wall integrity [44,45,46,47,48,49,50,51,52,53,54,55,56]. These proteins have both N- and O-linked sugars, predominantly mannans, which are also known to be present in the biofilm matrices of C. albicans [57,58,59]. The MNN2 gene is one putative element of N-linked glycosylation, directly responsible for mannans production for both cell and biofilm matrices of C. glabrata [57,58,59].
The goal of this work was to determine the expression profile of selected genes (Table 1 and Table 2) related to the production of biofilm matrix components in response to stress caused by drugs from the most important antifungal classes: azoles (Fluconazole, Flu), polyenes (Amphotericin B, AmB), and echinocandins (Caspofungin, Csf, and Micafungin, Mcf).
2. Materials and Methods
2.1. Organisms
Three strains of C. glabrata were used in the course of this study: One reference strain (C. glabrata ATCC 2001) from the American Type Culture Collection (Manassas, VA, USA), one strain recovered from the urinary tract (C. glabrata 562123) of a patient, and one strain recovered from the vaginal tract of a patient (C. glabrata 534784) in the Hospital Escala, Braga, Portugal. The identity of all isolates was confirmed using CHROMagarTM Candida (CHROMagarTM, Paris, France) and by PCR-based sequencing using specific primers (ITS1 and ITS4) against the 5.8 s subunit gene reference [60]. The PCR products were sequenced using the ABI-PRISM Big Dye terminator cycle sequencing kit (Perkin Elmer, Applied Biosystems, Warrington, UK).
2.2. Growth Conditions
For each experiment, C. glabrata ATCC2001, C. glabrata 534784, and C. glabrata 562123 strains were subcultured on Sabouraud dextrose agar (SDA) (Merck, Darmstadt, Germany) for 24 h at 37 °C. The cells were then inoculated in Sabouraud dextrose broth (SDB) (Merck) and incubated for 18 h at 37 °C under agitation at 120 rpm. After incubation, the cells were harvested by centrifugation at 3000× g (Thermo Scientific, CL10, Hampton, NH, USA) for 10 min at 4 °C and washed twice with phosphate buffered saline (PBS, pH = 7.5). The cell pellets were then suspended in Roswell Park memorial institute (RPMI), and the cellular density was adjusted to 1 × 105 cells/mL, using a Neubauer counting chamber.
2.3. Antifungal Drugs
Flu, Csf, and Mcf were kindly provided by Pfizer® (New York, NY, USA), MSD® (Kenilworth, NJ, USA) and Astellas Pharma, Ltd., (Tokyo, Japan), respectively, in their pure form. AmB was purchased from Sigma® (Sigma-Aldrich, Buffalo, NY, USA). Aliquots of 5000 mg/L were prepared using dimethyl sulfoxide (DMSO). The final concentrations used were prepared in RPMI-1640 (Sigma-Aldrich).
2.4. Biofilm Formation
The minimum biofilm eradicatory concentration (MBEC) values were previously determined by the group, according to the European committee on antimicrobial susceptibility testing (EUCAST) guidelines [61,62]. For biofilm formation, standardized cell suspensions (1000 μL) were placed into selected wells of 24-wells polystyrene microtiter plates (Orange Scientific, Braine-l’Alleud, Belgium). At 24 h, 500 μL of RPMI-1640 was removed, and an equal volume of fresh RPMI-1640 plus the antifungal solution was added, on the basis of the MBEC values determined and indicated in bold in Table 1 (2× concentrated). The plates were incubated at 37 °C for additional 24 h at 120 rpm. RPMI-1640 containing only the antifungal agent was used as a negative control. As a positive control, cell suspensions were tested in the absence of the antifungal agent [18].
2.5. Gene Expression Analysis
2.5.1. Gene Selection and Primer Design for Quantitative Real-Time PCR
Genes related to the production of biofilm matrix components (β-1,3, β-1,6 glucans, and mannans)—BGL2, FKS1, FKS2, GAS2, KNH1, UGP1, XOG1 and MNN2—were selected for this study. The gene sequences of interest were obtained from Candida Genome Database [63] and the primers for quantitative real-time PCR (RT-qPCR) were designed using Primer 3 [64] web-based software and are listed in Table 2. ACT1 was chosen as a housekeeping gene. In order to verify the specificity of each primer pair for its corresponding target gene, the PCR products were first amplified from C. glabrata ATCC2001.
2.5.2. Preparation of Biofilm Cells for RNA Extraction
After biofilm formation, the medium was eliminated, and the wells were washed with sterile water to remove non-adherent cells. The biofilms were scraped from the wells with 1 mL of sterile water and sonicated (Ultrasonic Processor, Cole-Parmer, IL, USA) for 30 s at 30 W to separate the cells from the biofilm matrix. The cells were harvested by centrifugation at 8000× g for 5 min at 4 °C [18].
2.5.3. RNA Extraction
RNA extraction was performed using PureLink RNA Mini Kit (Invitrogen, Carlsbad, CA, USA). Prior to RNA extraction, a lysis buffer from PureLink RNA Mini kit was prepared by adding 1% of ß-mercaptoethanol to the supplied buffer solution. Then, 500 µL of lysis buffer containing glass beads (0.5 mm diameter) was added to each pellet. The cell suspensions were homogenized twice for 30 s using a Mini-Bead-Beater-8 (Stratech Scientific, Soham, UK). After cell disruption, the PureLink RNA Mini Kit (Invitrogen) was used for total RNA extraction according to the manufacturer’s recommended protocol. To avoid potential DNA contamination, the samples were treated with RNase-Free DNase I (Invitrogen) [18].
2.5.4. Synthesis of Complementary DNA
To synthesize complementary DNA (cDNA), the iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA, USA) was used according to the manufacturer’s instructions. For each sample, 10 μL of the extracted RNA was used in a final reaction volume of 50 μL. cDNA synthesis was performed firstly at 70 °C for 5 min and then at 42 °C for 1 h. The reaction was stopped by heating for 5 min at 95 °C [18].
2.5.5. Quantitative Real-Time PCR
RT-qPCR (CFX96 Real-Time PCR System, Bio-Rad) was performed to determine the relative levels of all genes mRNA transcripts in the RNA samples, with ACT1 used as a reference Candida housekeeping gene. Each reaction mixture consisted of a working concentration of SoFast EvaGreen Supermix (Bio-Rad), 50 µM forward and reverse primers, and 4 µL cDNA, in a final reaction volume of 20 µL. Negative controls (water) as well as non-transcriptase reverse controls (NRT) were included in each run [18]. The relative quantification of gene expression was performed by the 2−ΔCT method [65]. Each reaction was performed in triplicate, and mean values of relative expression were determined for each gene. The results are presented after calculation of 2−ΔCT.
2.6. Statistical Analysis
All experiments were repeated three times in independent assays. The results were compared using one-way analysis of variance (ANOVA), Dunnett’s multiple comparisons tests, using GraphPadTM PrismTM 7 software (GraphPad Software, San Diego, CA, USA). All tests were performed with a confidence level of 95%. In order to determine the similarity of the strains’ gene profiles, the Pearson Correlation Coefficient (r) was also applied.
3. Results and Discussion
Candidaemia related to C. glabrata has been increasing in the last years in parallel with its high drug resistance, particularly to the azole antifungal class [1,20,66]. Biofilms of C. glabrata are highly recalcitrant to treatments with antifungal agents as a consequence of multiple resistance mechanisms, such as those linked to the presence of a strong net of exopolysaccharydes and other biopolymers that protect the cells and hinder the diffusion of the drugs [1,15,67,68,69]. In order to stress C. glabrata biofilm cells, four antifungals were applied (at concentrations based on MBECs values, Table 1) in pre-formed biofilms, and then an evaluation of biofilms’ matrix gene expression was performed and compared with the expression of a housekeeping gene.
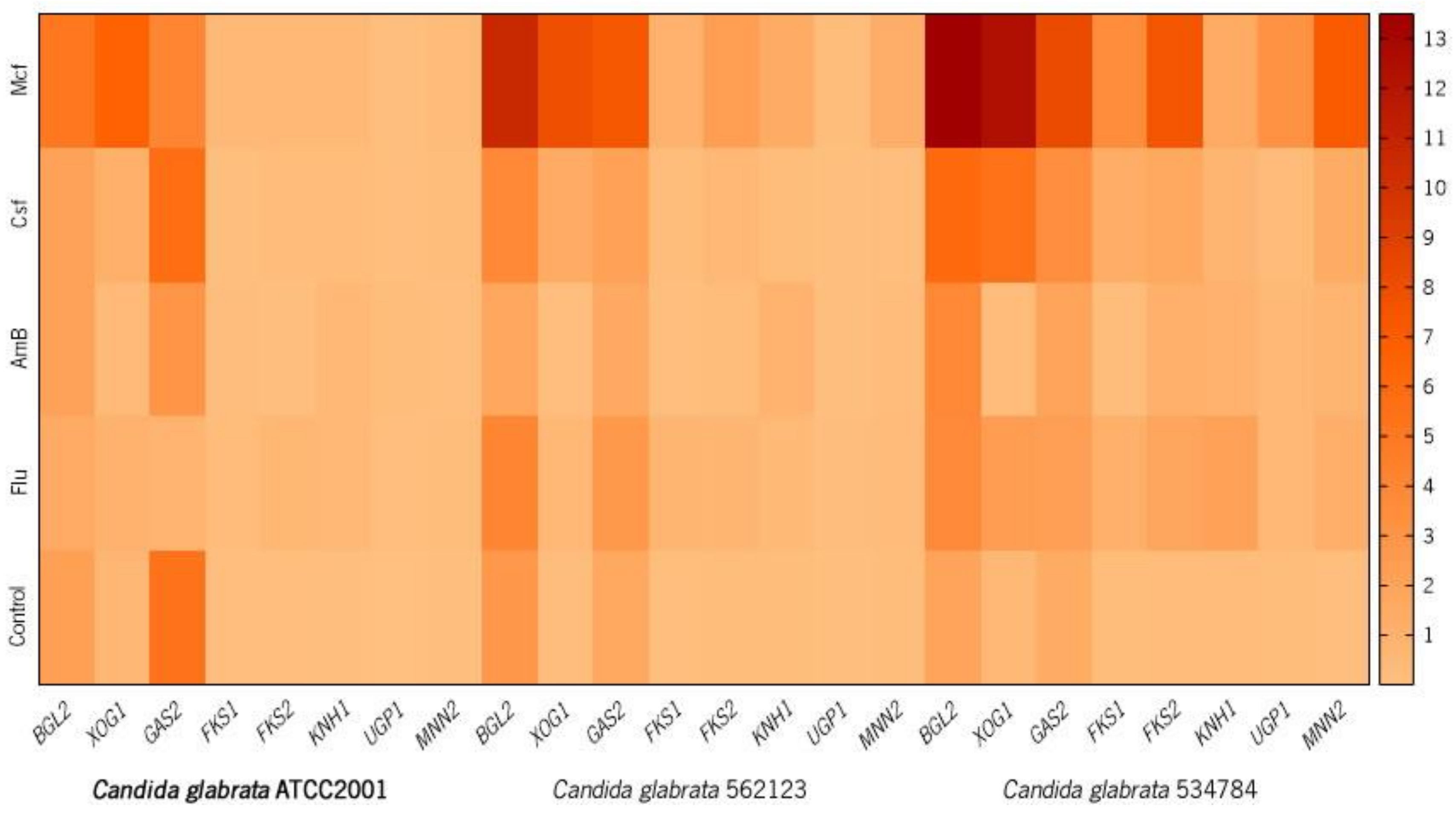
Figure 1 shows the heatmap with the results of the RT-qPCR expression profiling of biofilm cells of C. glabrata ATCC2001 (A), C. glabrata 562123 (B), and C. glabrata 534784 (C) in the presence of antifungal drugs. The final data are presented in fold-change (FC) in comparison to the expression of the housekeeping gene (2−ΔCT) [70].
Generally, BGL2, FKS1, FKS2, GAS2, and XOG1 displayed higher expression levels in biofilm cells in response to the drugs and, by contrast, KNH1, UPG1, and MNN2 displayed minor expression changes (Figure 1 and Table 3).
BGL2 showed similar expression in the control groups, and its FC expression decreased in the reference strain when Flu and Csf were present (FC: 1.40 and 2.00 respectively) and in the urinary strain when AmB was present (FC: 1.71). In all the other cases, BGL2 FC expression increased, particularly when the biofilms were treated with Mcf (FC: C. glabrata ATCC2001: 5.13; C. glabrata 562123: 10.58; C. glabrata 534784: 13.49). All changes in BGL2 expression were statistically significant compared to the untreated cells (p < 0.0001). Compared to the controls, XOG1 gene revealed a statistically significant downregulation after contact with AmB (FC: C. glabrata ATCC2001 0.37; C. glabrata 562123 0.10; C. glabrata 534784: 0.27) and overexpression in the presence of all the other antifungals, in all strains. For this gene, the most noteworthy overexpression was observed in Flu-treated C. glabrata 534784 (FC: 2.35, p < 0.0001), Csf-treated C. glabrata 562123 and C. glabrata 534784 (FC: 1.42 and 5.45, p < 0.0001, respectively), and in all strains after contact with Mcf (FC: C. glabrata ATCC2001: 6.54, C. glabrata 562123: 7.89, C. glabrata 534784: 12.38, all p < 0.0001).
In an important report, Taff et al. [19] concluded that mutants of C. albicans unable to produce Bgl2 and Xog1 enzymes did not show perturbations in the cell wall glucan composition of biofilm cells, and that these enzymes were not necessary for filamentation or biofilm formation. However, the biofilms had a reduced matrix glucan content, reduced total matrix biomass accumulation, and improved susceptibility to antifungal drug therapy [19]. Similarly, Li et al. [71] showed that, in C. albicans’ persister cells (frequent in biofilms [72,73]), there was an increased expression of cell wall integrity proteins such as Xog1 and Bgl2. These studies recognized a biofilm-specific pathway involving Bgl2 and Xog1 (and Phr1) enzymes and affecting matrix delivery, by which these enzymes release and modify cell wall glucan for deposition in the extracellular space; however, an alternative explanation is that these enzymes act in the extracellular space, being crucial for mature matrix organization and function [19]. These enzymes have been localized in the cell wall, supporting the hypothesis of their activity in the cell wall, but have also secretion sequences that support an extracellular function. As seen earlier, BGL2 is one of the glucan modifying genes for glucan delivery, and XOG1 is a glucanase [19], necessary for modification and delivery of carbohydrates to the mature biofilm matrix. Without delivery and accumulation of matrix glucan, the biofilms exhibit enhanced susceptibility to antifungal drugs [19]. The change in the regulation of BGL2 and XOG1 in the biofilm cells of C. glabrata after drug treatment that we observed is interpreted as a response of the biofilm cells to the reduction of biofilm matrix, specifically of β-1,3-glucans, and it has been described before [7,13,18].
The GAS gene family is also a regulator in the production of β-1,3-glucan, and Gas2 is a glycosylphosphatidylinositol (GPI)-anchored cell surface protein [31] involved in the production of β-1,3-glucan in C. glabrata [29,30]. Gas2 is a documented putative carbohydrate-active enzyme and consequently it can alter the cell wall polysaccharides in order to build and remodel the cell wall glycan network during growth in C. glabrata [29]. In C. glabrata ATCC2001, GAS2 was highly expressed in the non-treated group and after Csf contact (FC: 5.72), while Flu, AmB, and Mcf led to its downregulation. The clinical isolates upregulated the gene in all conditions, except for AmB treatment of C. glabrata 562123 (Figure 1 and Table 3, all p < 0.0001). Hence, when analyzing the results of C. glabrata 562123 and C. glabrata 534784, the GAS2 network seems to be activated also after glycan’s loss following drug treatment, in order to replace the lack of 1,3-β-glucans and re-establish biofilm cells’ homeostasis. All results compared with those from untreated cells (controls) were statistically significant (p < 0.0001).
The resistance to echinocandins increased from 4.9% to 12.3% between 2001 and 2010 [74] with a rapid development of FKS mutations in Candida spp., especially in C. glabrata [75,76]. The amino acid substitutions occurring in FKS1 [30,77,78,79] and FKS2 [30,80] are directly related to the resistance to this class of drugs: acquired FKS mutations [81] are reported to confer low β-(1,3)-d-glucan synthase sensitivity and to increase the minimum inhibitory concentration (MIC) values, which are related to clinical failure [82]; intrinsic FKS mutations, also lead to elevated MIC levels but have a weaker effect on the reduction of β-(1,3)-d-glucan synthase sensitivity [82,83,84,85]. Generally, in the C. glabrata strains, a 24 h contact with both echinocandins upregulated FKS1 and FKS2 genes and, in the reference strain, the presence of Mcf upregulated FKS1. More specifically, the results showed that all strains upregulated the expression of FKS1 after drug exposure (statistically significant), with the exception of Csf in C. glabrata ATCC2001 (FC: 0.07 p < 0.001) and Csf in C. glabrata 562123 (FC: 0.08; p < 0.0001). For FKS2, its overexpression was observed for almost all treatments in the three strains, excluding following AmB treatment in C. glabrata ATCC2001 (FC: 0.28; p < 0.001) and C. glabrata 562123 (FC: 0.55; p < 0.05). C. glabrata 534784 revealed, again, to have the highest capacity to overexpress both genes in response to drug stress. These differences among the strains may be related to the described Candida spp. intra-strains variations [62]. Bizerra et al. [76] reported the occurrence of a mutation associated with the resistance phenotype against echinocandins in C. glabrata isolated from a single cancer patient with candidemia exposed to antifungal prophylaxis with Mcf. Arendrup et al. [86] revealed that Mcf MICs of C. glabrata FKS hot spot mutant isolates were less raised than those obtained for the other echinocandins, showing that the efficacy of Mcf could be differentially dependent on specific FKS genes mutations. These reports mention singularities regarding the FKS gene and Mcf, which can also be observed in our results (Figure 1 and Table 3). Interestingly, up and downregulations of FKS1 and FKS2 were similar in the clinical isolates and parallel to those observed for BGL2, which makes sense, since this gene has shown to perform, with XOG1 (and PHR1), in a complementary manner in order to distribute the matrix downstream of the primary β-1,3 glucan synthase encoded by FKS1 [19]. Previous investigations also found elevated transcript levels of FKS1, BGL2, and XOG1 during in vivo C. albicans biofilm growth when compared to planktonic growth, which is consistent with our results and with a role in a biofilm-specific function, such as matrix formation [87,88].
The overexpressed values obtained for BGL1, XOG1, FKS1, FKS2, and GAS2 after the stress conditions induced by most antifungals endorse the impact of β-1,3-glucans in the maintenance of the cell and biofilm matrix structure.
Sequencing studies have shown that C. glabrata is more closely related to S. cerevisiae than to C. albicans [89], with some genes functionally interchangeable among the two species [90,91]. An important component of the cell wall and the biofilm matrix is β-1,6-glucan, which is regulated by several genes, such as KNH1. Preceding studies have demonstrated that the KNH1 homologs are essential components of β-1,6-glucan synthesis in C. glabrata [35,36,38]. In S. cerevisiae, many genes involved in β-1,6-glucan synthesis were isolated through mutations (kre [killer resistant] mutations) that are responsible for the resistance to the K1 killer toxin, which kills sensitive yeast cells after binding to β-1,6-glucan [35,38,92]. Dijkgraaf and colleagues [35] reported that the disruption of both KRE9 and KNH1 was synthetically lethal for C. glabrata, demonstrating the importance of these genes in the maintenance of cell structure. In the present study, after a drug stress, all C. glabrata strains upregulated this gene (Figure 1 and Table 3), indicating an effort to replace these β-1,6-glucans after losses due to the aggression of the antifungals, confirming also a certain degree of relevance of these elements in the cell wall and biofilm matrix of C. glabrata [35,36,38]. C. glabrata ATCC2001 showed to upregulate the KNH1 gene in the presence of antifungal drugs, and C. glabrata 562123 indicated an identical pattern by marginally increasing KNH1 gene expression in these conditions. Compared to the other two strains, with the exception of Flu treatment (FC: 2.08; p < 0.0001), KNH1 showed a different regulation in the vaginal tract strain (Figure 1 and Table 3). C. glabrata 534784 demonstrated to have the highest upregulation capacity, presenting overexpression almost for all genes (Figure 1 and Table 3).
During glucose starvation, a set of genes orthologous to S. cerevisiae is induced in C. glabrata, including UGP1, related to the β-1,6-d-glucan biosynthetic process [39,40], which shows that the environmental stress response is conserved between S. cerevisiae and C. glabrata [39]. UPG1 showed to have the lowest expression, compared with other genes and controls. Nonetheless, except for one condition and strain, in the presence of antifungal drugs, several overexpression states were observed (Figure 1 and Table 3). The reference strain displayed overexpression in all conditions, with the highest gene upregulation occurring in the presence of AmB (FC: 0.17; p < 0.0001) and the lowest in the presence of Csf (FC: 0.01; non-significant); the urinary tract strain also revealed limited gene upregulation in the presence of Csf and AmB and the highest expression in the presence of Mcf (0.15; p < 0.0001). Generally, C. glabrata 534784 demonstrated the highest FC expression in all conditions. The lowest gene upregulation was observed in biofilm cells stressed by Csf (FC: 0.33; p < 0.0001). Srikantha and colleagues [91] identified a set of genes that are upregulated by the transcription factor Bcr1, involved in impermeability, impenetrability, and drug resistance of C. albicans’ biofilms. The authors concluded that the induction of Bcr1 overexpression in weak biofilms of C. albicans conferred those three characteristics and, in these cases, UGP1 gene was downregulated [91]. This result supports the FC expression we obtained: since C. glabrata biofilms were weakened by the drugs, UGP1 expression was increased in order to balance this defect (as seen with KNH1). The overexpression values we obtained for both KNH1 and UPG1 point to the relevance of β-1,6-glucans in the maintenance of a good cell and matrix structure.
Regarding mannans regulation, all strains showed a low or moderate expression of MNN2 in the controls (non-treated cells), but relevant expression changes arose in the presence of all drugs, particularly when Mcf was added (Figure 1 and Table 3). C. glabrata ATCC2001 demonstrated the lowest expression in the control group, among all strains (Figure 1 and Table 3). The urinary strain presented the lowest gene expression, when compared to the untreated group (Figure 1 and Table 3). Flu and Mcf induced the highest MNN2 values (FC: 0.34 and 1.21, respectively, both p < 0.0001), while AmB and Csf were associated with the lowest expressions (FC: 0.24, p < 0.0001 and 0.13, p < 0.0005). For C. glabrata ATCC2001 and C. glabrata 534784, the weakest effects were associated with the biofilm cells that were stressed by AmB (FC: 0.13 and 0.71 respectively, both p < 0.0001). When Mcf was applied, the biofilm cells of the vaginal strain showed a strong response to the stress, compared to the other two strains (FC: 7.03; p < 0.0001). Our team has also found that C. glabrata ATCC2001 increased the amounts of mannans on its cell walls in the presence of these drugs (data not shown), revealing a possible adaptation of the cells to the stress caused by the antifungal drugs. Other studies reported analogous adjustments of the cell walls after environmental drug stress, which has been related to high antifungal resistance events [1,2,19,92,93,94], supporting these results.
Interestingly, and when compared to the rest of the genes, the present results demonstrate that KNH1, UGP1, and MNN2 had the lowest values of FC expression. This seems to indicate that, although β-1,6-glucans are an important part of the cell and biofilm matrix, the cells appear to invest more in replacing the lost β-1,3-glucans, leading to consider that these components have a greater significance in the maintenance of the homeostasis of the biofilm matrix and the biofilm cells. In fact, in studies developed in our group [13,18], the total polysaccharides and β-1,3-glucans concentrations increased significantly in C. glabrata biofilm matrices after Flu, AmB, and Mcf contact. These higher concentrations in β-1,3 glucans content might explain part of the main biofilm resistance to the drugs that was formerly described [7,95,96,97].
Finally, downregulation of most genes and strains happened in the presence of AmB whereas, in opposition, Mcf induced the main overexpression alterations (Figure 1 and Table 3). AmB is a fungicidal drug and the most important antifungal polyene used for the treatment of systemic fungal infections [98,99]. This drug binds to the ergosterol of the cell membrane but also induces oxidative stress. This explains the existence of a still low reported rate of resistance and the good effectiveness of AmB [1,13,100,101,102,103,104]. Also, this low resistance may be associated with the lower gene expression effects that we detected after AmB exposure in C. glabrata (Figure 1 and Table 3). In opposition, the most acute upregulations occurred in the presence of echinocandins and, particularly, when Mcf was applied. This class of antifungals act by inhibiting β-1,3-glucan synthesis [1,100,105], which affects cell wall and matrix composition. By overexpressing the genes related to β-1,3-glucan synthesis (BGL2, FKS1, FKS2, GAS2, XOG1), the cells were attempting to compensate and replace the β-1,3-glucan losses in their matrices induced by the drugs and, thus, protect and decrease their susceptibility to the antifungals [19]. This general increase in total carbohydrates and specifically in β-1,3-glucans in Candida spp. biofilm matrices has already been described [7,13,18].
Regarding the correlation between the gene expression profiles in C. glabrata, the results based on the r are displayed in Table 4.
The results showed a strong positive correlation (r near 1) between the response profiles of BGL2, XOG1, FKS1, and MNN2 gene expression in the three strains, which means that up and downregulation had a high tendency to occur similarly in all strains. The scores of the r for the profile of the FKS2 gene revealed a moderate positive correlation between the reference strain (C. glabrata ATCC2001) and the isolates (C. glabrata 562123 and C. glabrata 534784). This indicates that, although the correlation was positive, it was weak, and the profiles of the gene response were variable in the three strains. On the other hand, the clinical isolates showed strong positive correlation between the expression profiles of this gene. C. glabrata ATCC2001 demonstrated a moderate positive correlation between the expression profiles of the GAS2 gene. C. glabrata 562123 and C. glabrata 534784 had a strong positive correlation between the expression profiles of the GAS2 gene. KNH1 gene was the most variable and difficult gene to correlate between the strains. The reference and the 562123 strain showed a moderate correlation, whereas the reference and the 534784 strain showed a strong correlation, and the clinical isolates showed the only weak correlation detected in this study. As for the UPG1 gene, although there was a negative correlation, the association between its expression in ATCC2001 and in the clinical isolates can be considered weak. Between the isolates, it was determined that UPG1 up and downregulation had a high tendency to occur similarly in all strains (thus showing strong correlation). In summary, BGL2, XOG1, FKS1, and MNN2 appeared to be the genes presenting the most similar responses to antifungal drugs within the transcriptome of the three strains; also, the clinical isolates appeared to be nearer each other than to the reference strain. Once more, β-1,3-glucan synthesis was identified as important in C. glabrata (three of the four genes affected are responsible for β-1,3-glucan production). These similarities among the two clinical strains may be due to the fact that both were derived from a hospital environment, and it is probable that they had already been challenged by several drugs, so their responses were prompter compared to the reference strain that is a wild type strain.
4. Conclusions
The in vitro high-dose paradox associated with Candida spp. isolates is being increasingly reported and connected to slightly elevated MICs, potentially contributing to clinical resistance and failure of antifungal treatments. These drug tolerance and adaptive mechanisms are highly related to Candida spp. biofilm forms. C. glabrata extracellular matrix is crucial for mature biofilm formation, not only contributing to the adhesive nature of the biofilm cells, but also protecting the cells from antifungal agents and from the host immune system. Understanding the production of the biofilm matrix components and the associated delivery processes is important for the development of effective biofilm therapies. All stakeholders in this process represent potentially attractive targets for detection of and therapeutic interventions against candidiasis.
Acknowledgments
This study was supported by the Portuguese Foundation for Science and Technology (FCT) under the scope of the strategic funding of UID/BIO/04469/2013 unit and COMPETE 2020 (POCI-01-0145-FEDER-006684) and BioTecNorte operation (NORTE-01-0145-FEDER-000004) funded by the European Regional Development Fund under the scope of Norte2020—Programa Operacional Regional do Norte and Célia F. Rodrigues’ [SFRH/BD/93078/2013] Ph.D. grant. We also would like to acknowledge MSD® and Astellas® for the kind donation of Caspofungin and Micafungin, respectively.
Author Contributions
Célia F. Rodrigues conceived, designed, performed the experiments, analyzed the data, and wrote the paper; Mariana Henriques conceived, designed the experiments, analyzed the data, and wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rodrigues, C.F.; Rodrigues, M.E.; Silva, S.; Henriques, M. Candida glabrata biofilms: How far have we come? J. Fungi 2017, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Rodrigues, C.F.; Araújo, D.; Rodrigues, M.E.; Henriques, M. Candida species biofilms’ antifungal resistance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Costa-Orlandi, C.; Sardi, J.; Pitangui, N.; de Oliveira, H.; Scorzoni, L.; Galeane, M.; Medina-Alarcón, K.; Melo, W.; Marcelino, M.; Braz, J.; et al. Fungal biofilms and polymicrobial diseases. J. Fungi 2017, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- McCall, A.; Edgerton, M. Real-time approach to flow cell imaging of Candida albicans biofilm development. J. Fungi 2017, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Johnson, A.D. Candida albicans biofilms and human disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, E.; Silva, S.; Rodrigues, C.F.; Alves, C.T.; Azeredo, J.; Henriques, M. Effects of fluconazole on Candida glabrata biofilms and its relationship with ABC transporter gene expression. Biofouling 2014, 30, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.; Silva, S.; Henriques, M. Candida glabrata: A review of its features and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 673–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negri, M.; Silva, S.; Henriques, M.; Oliveira, R. Insights into Candida tropicalis nosocomial infections and virulence factors. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1399–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial Biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.; Costerton, J. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Candida glabrata, Candida parapsilosis and Candida tropicalis: Biology, epidemiology, pathogenicity and antifungal resistance. FEMS Microbiol. Rev. 2012, 36, 288–305. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.; Silva, S.; Azeredo, J.; Henriques, M. Candida glabrata’s recurrent infections: Biofilm formation during Amphotericin B treatment. Lett. Appl. Microbiol. 2016, 63, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.; Lincoln, L.; Marchillo, K.; Massey, R.; Holoyda, K.; Hoff, B.; VanHandel, M.; Andes, D. Putative role of β-1,3 glucans in Candida albicans biofilm resistance. Antimicrob. Agents Chemother. 2007, 51, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.E.; Sanchez, H.; Cain, M.T.; Andes, D.R. Genetic basis of Candida biofilm resistance due to drug-sequestering matrix glucan. J. Infect. Dis. 2010, 202, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.E.; Crawford, K.; Marchillo, K.; Andes, D.R.; Nett, J.E.; Crawford, K.; Marchillo, K.; Andes, D.R. Role of Fks1p and matrix glucan in Candida albicans biofilm resistance to an echinocandin, pyrimidine, and polyene. Antimicrob. Agents Chemother. 2010, 54, 3505–3508. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.E.; Sanchez, H.; Cain, M.T.; Ross, K.M.; Andes, D.R. Interface of Candida albicans biofilm matrix-associated drug resistance and cell wall integrity regulation. Eukaryot. Cell 2011, 10, 1660–1669. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.; Gonçalves, B.; Rodrigues, M.E.; Silva, S.; Azeredo, J.; Henriques, M. The effectiveness of voriconazole in therapy of Candida glabrata’s biofilms oral infections and its influence on the matrix composition and gene expression. Mycopathologia 2017, 182, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Taff, H.T.; Nett, J.E.; Zarnowski, R.; Ross, K.M.; Sanchez, H.; Cain, M.T.; Hamaker, J.; Mitchell, A.P.; Andes, D.R. A Candida biofilm-induced pathway for matrix glucan delivery: Implications for drug resistance. PLoS Pathog. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.F.; Henriques, M. Oral mucositis caused by Candida glabrata biofilms: Failure of the concomitant use of fluconazole and ascorbic acid. Ther. Adv. Infect. Dis. 2017, 1. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Ribot, J.L. Large-scale biochemical profiling of the Candida albicans biofilm matrix: New compositional, structural, and functional insights. MBio 2014, 5, e01314–e01333. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C. Fungal biofilm resistance. Int. J. Microbiol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.F.; Zarnowski, R.; Andes, D.R. Fungal super glue: The biofilm matrix and its composition, assembly, and functions. PLoS Pathog. 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Douglas, C. Fungal beta (1,3)-d-glucan synthesis. Med. Mycol. 2001, 39, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Del Mar Gonzalez, M.; Diez-Orejas, R.; Molero, G.; Alvarez, A.M.; Pla, J.; Nombela, C.; Sanchez-Perez, M. Phenotypic characterization of a Candida albicans strain deficient in its major exoglucanase. Microbiology 1997, 143, 3023–3032. [Google Scholar] [CrossRef] [PubMed]
- Sarthy, A.V.; McGonigal, T.; Coen, M.; Frost, D.J.; Meulbroek, J.A.; Goldman, R.C. Phenotype in Candida albicans of a disruption of the BGL2 gene encoding a 1,3-β-glucosyltransferase. Microbiology 1997, 143, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Mouyna, I.; Fontaine, T.; Vai, M.; Monod, M.; Fonzi, W.A.; Diaquin, M.; Popolo, L.; Hartland, R.P.; Latgé, J.P. Glycosylphosphatidylinositol-anchored glucanosyltransferases play an active role in the biosynthesis of the fungal cell wall. J. Biol. Chem. 2000, 275, 14882–14889. [Google Scholar] [CrossRef] [PubMed]
- Goldman, R.C.; Sullivan, P.A.; Zakula, D.; Capobianco, J.O. Kinetics of β-1,3 glucan interaction at the donor and acceptor sites of the fungal glucosyltransferase encoded by the BGL2 gene. Eur. J. Biochem. 1995, 227, 372–378. [Google Scholar] [CrossRef] [PubMed]
- De Groot, P.W.J.; Kraneveld, E.A.; Yin, Q.Y.; Dekker, H.L.; Gross, U.; Crielaard, W.; de Koster, C.G.; Bader, O.; Klis, F.M.; Weig, M. The cell wall of the human pathogen Candida glabrata: Differential incorporation of novel adhesin-like wall proteins. Eukaryot. Cell 2008, 7, 1951–1964. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Effron, G.; Lee, S.; Park, S.; Cleary, J.D.; Perlin, D.S. Effect of Candida glabrata FKS1 and FKS2 mutations on echinocandin sensitivity and kinetics of 1,3-β-d-glucan synthase: Implication for the existing susceptibility breakpoint. Antimicrob. Agents Chemother. 2009, 53, 3690–3699. [Google Scholar] [CrossRef] [PubMed]
- Vai, M.; Orlandi, I.; Cavadini, P.; Alberghina, L.; Popolo, L. Candida albicans homologue of GGP1/GAS1 gene is functional in Saccharomyces cerevisiae and contains the determinants for glycosylphosphatidylinositol attachment. Yeast 1996, 12, 361–368. [Google Scholar] [CrossRef]
- Miyazaki, T.; Nakayama, H.; Nagayoshi, Y.; Kakeya, H.; Kohno, S. Dissection of Ire1 functions reveals stress response mechanisms uniquely evolved in Candida glabrata. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Montijn, R.C.; van Rinsum, J.; van Schagen, F.A.; Klis, F.M. Glucomannoproteins in the cell wall of Saccharomyces cerevisiae contain a novel type of carbohydrate side chain. J. Biol. Chem. 1994, 269, 19338–19342. [Google Scholar] [PubMed]
- Kapteyn, J.C.; Montijn, R.C.; Vink, E.; de la Cruz, J.; Llobell, A.; Douwes, J.E.; Shimoi, H.; Lipke, P.N.; Klis, F.M. Retention of Saccharomyces cerevisiae cell wall proteins through a phosphodiester-linked β-1,3-/β-1,6-glucan heteropolymer. Glycobiology 1996, 6, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Dijkgraaf, G.J.; Brown, J.L.; Bussey, H. The KNH1 gene of Saccharomyces cerevisiae is a functional homolog of KRE9. Yeast 1996, 12, 683–692. [Google Scholar] [CrossRef]
- Nagahashi, S.; Lussier, M.; Bussey, H. Isolation of Candida glabrata homologs of the Saccharomyces cerevisiae KRE9 and KNH1 genes and their involvement in cell wall β-1,6-glucan synthesis. J. Bacteriol. 1998, 180, 5020–5029. [Google Scholar] [PubMed]
- Cid, V.J.; Durán, A.; del Rey, F.; Snyder, M.P.; Nombela, C.; Sánchez, M. Molecular basis of cell integrity and morphogenesis in Saccharomyces cerevisiae. Microbiol. Rev. 1995, 59, 345–386. [Google Scholar] [PubMed]
- Brown, J.L.; Kossaczka, Z.; Jiang, B.; Bussey, H. A mutational analysis of killer toxin resistance in Saccharomyces cerevisiae identifies new genes involved in cell wall (1–6)-β-glucan synthesis. Genetics 1993, 133, 837–849. [Google Scholar] [PubMed]
- Roetzer, A.; Gregori, C.; Jennings, A.M.; Quintin, J.; Ferrandon, D.; Butler, G.; Kuchler, K.; Ammerer, G.; Schüller, C. Candida glabrata environmental stress response involves Saccharomyces cerevisiae Msn2/4 orthologous transcription factors. Mol. Microbiol. 2008, 69, 603–620. [Google Scholar] [CrossRef] [PubMed]
- Dodgson, A.R.; Pujol, C.; Denning, D.W.; Soll, D.R.; Fox, A.J. Multilocus sequence typing of Candida glabrata reveals geographically enriched clades. J. Clin. Microbiol. 2003, 41, 5709–5717. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, J.; Victoria Elorza, M.; Valentin, E.; Sentandreu, R. Molecular organization of the cell wall of Candida albicans and its relation to pathogenicity. FEMS Yeast Res. 2006, 6, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Latgé, J.-P. The cell wall: A carbohydrate armour for the fungal cell. Mol. Microbiol. 2007, 66, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Brown, G.D.; Kullberg, B.J.; Gow, N.A.R. An integrated model of the recognition of Candida albicans by the innate immune system. Nat. Rev. Microbiol. 2008, 6, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Mardi, L. Innate immune mechanisms for recognition and uptake of Candida species. Trends Immunol. 2010, 31, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.R.; Netea, M.G.; Munro, C.A.; Ferwerda, G.; Bates, S.; Mora-Montes, H.M.; Walker, L.; Jansen, T.; Jacobs, L.; Tsoni, V.; et al. Immune recognition of Candida albicans β-glucan by Dectin-1. J. Infect. Dis. 2007, 196, 1565–1571. [Google Scholar] [CrossRef] [PubMed]
- Van de Veerdonk, F.L.; Kullberg, B.J.; van der Meer, J.W.; Gow, N.A.; Netea, M.G. Host-microbe interactions: Innate pattern recognition of fungal pathogens. Curr. Opin. Microbiol. 2008, 11, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.M.; Gow, N.A.; Brown, G.D. Pattern recognition: Recent insights from Dectin-1. Curr. Opin. Immunol. 2009, 21, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Mora-Montes, H.M.; Bates, S.; Netea, M.G.; Castillo, L.; Brand, A.; Buurman, E.T.; Diaz-Jimenez, D.F.; Jan Kullberg, B.; Brown, A.J.P.; Odds, F.C.; et al. A multifunctional mannosyltransferase family in Candida albicans determines cell wall mannan structure and host-fungus interactions. J. Biol. Chem. 2010, 285, 12087–12095. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.R.; Tsoni, S.V.; Willment, J.A.; Dennehy, K.M.; Rosas, M.; Findon, H.; Haynes, K.; Steele, C.; Botto, M.; Gordon, S.; et al. Dectin-1 is required for β-glucan recognition and control of fungal infection. Nat. Immunol. 2007, 8, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Murciano, C.; Moyes, D.L.; Runglall, M.; Islam, A.; Mille, C.; Fradin, C.; Poulain, D.; Gow, N.A.R.; Naglik, J.R. Candida albicans cell wall glycosylation may be indirectly required for activation of epithelial cell proinflammatory responses. Infect. Immun. 2011, 79, 4902–4911. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.; Hube, B. Importance of the Candida albicans cell wall during commensalism and infection. Curr. Opin. Microbiol. 2012, 15, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.; Hughes, H.B.; Munro, C.A.; Thomas, W.P.H.; MacCallum, D.M.; Bertram, G.; Atrih, A.; Ferguson, M.A.J.; Brown, A.J.P.; Odds, F.C.; et al. Outer chain N-glycans are required for cell wall integrity and virulence of Candida albicans. J. Biol. Chem. 2006, 281, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.; MacCallum, D.M.; Bertram, G.; Munro, C.A.; Hughes, H.B.; Buurman, E.T.; Brown, A.J.P.; Odds, F.C.; Gow, N.A.R. Candida albicans Pmr1p, a secretory pathway P-type Ca2+/Mn2+-ATPase, is required for glycosylation and virulence. J. Biol. Chem. 2005, 280, 23408–23415. [Google Scholar] [CrossRef] [PubMed]
- Mora-Montes, H.M.; Bates, S.; Netea, M.G.; Diaz-Jimenez, D.F.; Lopez-Romero, E.; Zinker, S.; Ponce-Noyola, P.; Kullberg, B.J.; Brown, A.J.P.; Odds, F.C.; et al. Endoplasmic reticulum -glycosidases of Candida albicans are required for N glycosylation, cell wall integrity, and normal host-fungus interaction. Eukaryot. Cell 2007, 6, 2184–2193. [Google Scholar] [CrossRef] [PubMed]
- Munro, C.A.; Bates, S.; Buurman, E.T.; Hughes, H.B.; MacCallum, D.M.; Bertram, G.; Atrih, A.; Ferguson, M.A.J.; Bain, J.M.; Brand, A.; et al. Mnt1p and Mnt2p of Candida albicans are partially redundant α-1,2-mannosyltransferases that participate in O-linked mannosylation and are required for adhesion and virulence. J. Biol. Chem. 2005, 280, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Saijo, S.; Ikeda, S.; Yamabe, K.; Kakuta, S.; Ishigame, H.; Akitsu, A.; Fujikado, N.; Kusaka, T.; Kubo, S.; Chung, S.; et al. Dectin-2 recognition of α-mannans and induction of Th17 cell differentiation is essential for host defense against Candida albicans. Immunity 2010, 32, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Lal, P.; Sharma, D.; Pruthi, P.; Pruthi, V. Exopolysaccharide analysis of biofilm-forming Candida albicans. J. Appl. Microbiol. 2010, 109, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Correia, I.A. Role of Secreted Aspartyl Proteases in Candida Albicans Virulence, Host Immune Response and Immunoprotection in Murine Disseminated Candidiasis; Universidade do Minho: Braga, Portugal, 2012. [Google Scholar]
- Johnson, C.J.; Cabezas-Olcoz, J.; Kernien, J.F.; Wang, S.X.; Beebe, D.J.; Huttenlocher, A.; Ansari, H.; Nett, J.E. The extracellular matrix of Candida albicans biofilms impairs formation of neutrophil extracellular traps. PLoS Pathog. 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.W.; Wilson, M.J.; Lewis, M.A.O.; Potts, A.J.C. Identification of Candida species by PCR and restriction fragment length polymorphism analysis of intergenic spacer regions of ribosomal DNA. J. Clin. Microbiol. 1995, 33, 2476–2479. [Google Scholar] [PubMed]
- Arendrup, M.C.; Arikan, S.; Barchiesi, F.; Bille, J.; Dannaoui, E.; Denning, D.W.; Donnelly, J.P.; Fegeler, W.; Moore, C.; Richardson, M.; et al. EUCAST Technical note on the method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for conidia—Forming moulds. ESCMID Tech. Notes 2008, 14, 982–984. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing, EUCAST Breakpoint Tables for Interpretation of MICs, Version 8.1; Available online: http://www.eucast.org.
- Skrzypek, M.S.; Binkley, J.; Binkley, G.; Miyasato, S.R.; Simison, M.; Sherlock, G. The Candida Genome Database (CGD): Incorporation of Assembly 22, systematic identifiers and visualization of high throughput sequencing data. Nucleic Acids Res. 2017, 45, D592–D596. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.-J.; Shin, J.H.; Kim, S.H.; Kim, H.-K.; Park, J.S.; Sung, H.; Kim, M.N.; Im, H.J. Emergence of multiple resistance profiles involving azoles, echinocandins and Amphotericin B in Candida glabrata isolates from a neutropenia patient with prolonged fungaemia. J. Antimicrob. Chemother. 2015, 70, 1268–1270. [Google Scholar] [CrossRef] [PubMed]
- Douglas, L.J. Candida biofilms and their role in infection. Trends Microbiol. 2003, 11, 30–36. [Google Scholar] [CrossRef]
- Zarnowski, R.; Westler, W.M.; Lacmbouh, G.A.; Marita, J.M.; Bothe, J.R.; Bernhardt, J.; Sahraoui, A.L.H.; Fontainei, J.; Sanchez, H.; Hatfeld, R.D.; et al. Novel entries in a fungal biofilm matrix encyclopedia. MBio 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Chandra, J. Candida biofilm resistance. Drug. Resist. Updat. 2004, 7, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J. User Bulletin #2 ABI P RISM 7700 Sequence Detection System SUBJECT: Relative Quantitation of Gene Expression—Updated 2001; Applied Biosystems: Foster City, CA, USA, 1997. [Google Scholar]
- Li, P.; Seneviratne, C.; Alpi, E.; Vizcaino, J.; Jin, L. Delicate metabolic control and coordinated stress response critically determine antifungal tolerance of Candida albicans biofilm persisters. Antimicrob. Agents Chemother. 2015, 59, 6101–6112. [Google Scholar] [CrossRef] [PubMed]
- Al-Dhaheri, R.S.; Douglas, L.J. Absence of Amphotericin B-tolerant persister cells in biofilms of some Candida species. Antimicrob. Agents Chemother. 2008, 52, 1884–1887. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, Z.; Chu, H.; Guo, J.; Jiang, G.; Qi, Q. Candida albicans Amphotericin B-tolerant persister formation is closely related to surface adhesion. Mycopathologia 2016, 181, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.D.; Johnson, M.D.; Pfeiffer, C.D.; Jimenez-Ortigosa, C.; Catania, J.; Booker, R.; Castanheira, M.; Messer, S.A.; Perlin, D.S.; Pfaller, M.A. Increasing echinocandin resistance in Candida glabrata: Clinical failure correlates with presence of FKS mutations and elevated minimum inhibitory concentrations. Clin. Infect. Dis. 2013, 56, 1724–1732. [Google Scholar] [CrossRef] [PubMed]
- Pinhati, H.M.S.; Casulari, L.A.; Souza, A.C.R.; Siqueira, R.A.; Damasceno, C.M.G.; Colombo, A.L. Outbreak of candidemia caused by fluconazole resistant Candida parapsilosis strains in an intensive care unit. BMC Infect. Dis. 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Bizerra, F.C.; Jimenez-Ortigosa, C.; Souza, A.C.R.; Breda, G.L.; Queiroz-Telles, F.; Perlin, D.S.; Colombo, A.L. Breakthrough candidemia due to multidrug-resistant Candida glabrata during prophylaxis with a low dose of micafungin. Antimicrob. Agents Chemother. 2014, 58, 2438–2440. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kelly, R.; Kahn, J.N.N.; Robles, J.; Hsu, M.-J.; Register, E.; Li, W.; Vyas, V.; Fan, H.; Abruzzo, G.; et al. Specific substitutions in the echinocandin target Fks1p account for reduced susceptibility of rare laboratory and clinical Candida sp. isolates. Antimicrob. Agents Chemother. 2005, 49, 3264–3273. [Google Scholar] [CrossRef] [PubMed]
- Desnos-Ollivier, M.; Moquet, O.; Chouaki, T.; Guerin, A.M.; Dromer, F. Development of echinocandin resistance in Clavispora lusitaniae during caspofungin treatment. J. Clin. Microbiol. 2011, 49, 2304–2306. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.H.; Johansen, H.K.; Arendrup, M.C. Stepwise development of a homozygous S80P substitution in Fks1p, conferring echinocandin resistance in Candida tropicalis. Antimicrob. Agents Chemother. 2013, 57, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.S.; Wiederhold, N.P.; Wickes, B.L.; Patterson, T.F.; Jorgensen, J.H. Rapid emergence of echinocandin resistance in Candida glabrata resulting in clinical and microbiologic failure. Antimicrob. Agents Chemother. 2013, 57, 4559–4561. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.K.; Nguyen, M.H.; Press, E.G.; Kwa, A.L.; Cheng, S.; Du, C.; Clancy, C.J. The presence of an FKS mutation rather than MIC is an independent risk factor for failure of echinocandin therapy among patients with invasive candidiasis due to Candida glabrata. Antimicrob. Agents Chemother. 2012, 56, 4862–4869. [Google Scholar] [CrossRef] [PubMed]
- Beyda, N.D.; Lewis, R.E.; Garey, K.W. Echinocandin resistance in Candida species: Mechanisms of reduced susceptibility and therapeutic approaches. Ann. Pharmacother. 2012, 46, 1086–1096. [Google Scholar] [CrossRef] [PubMed]
- Barchiesi, F.; Spreghini, E.; Tomassetti, S.; Della Vittoria, A.; Arzeni, D.; Manso, E.; Scalise, G. Effects of caspofungin against Candida guilliermondii and Candida parapsilosis. Antimicrob. Agents Chemother. 2006, 50, 2719–2727. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Effron, G.; Katiyar, S.K.; Park, S.; Edlind, T.D.; Perlin, D.S. A naturally occurring proline-to-alanine amino acid change in Fks1p in Candida parapsilosis, Candida orthopsilosis, and Candida metapsilosis accounts for reduced echinocandin susceptibility. Antimicrob. Agents Chemother. 2008, 52, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Forastiero, A.; Garcia-Gil, V.; Rivero-Menendez, O.; Garcia-Rubio, R.; Monteiro, M.C.; Alastruey-Izquierdo, A.; Jordan, R.; Agorio, I.; Mellado, E. Rapid development of Candida krusei echinocandin resistance during caspofungin therapy. Antimicrob. Agents Chemother. 2015, 59, 6975–6982. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.; Perlin, D.; Jensen, R.; Howard, S.; Goodwin, J.; Hopec, W. Differential in vivo activities of anidulafungin, caspofungin, and micafungin against Candida glabrata isolates with and without FSK resistance mutations. Antim Agents Chemoter. 2012, 56, 2435–2442. [Google Scholar] [CrossRef] [PubMed]
- Fanning, S.; Mitchell, A.P. Fungal Biofilms. PLoS Pathog. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.; Lepak, A.; Marchillo, K.; Andes, D. Time course global gene expression analysis of an in vivo Candida biofilm. J. Infect. Dis. 2009, 200, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Barns, S.M.; Lane, D.J.; Sogin, M.L.; Bibeau, C.; Weisburg, W.G. Evolutionary relationships among pathogenic Candida species and relatives. J. Bacteriol. 1991, 173, 2250–2255. [Google Scholar] [CrossRef] [PubMed]
- Kitada, K.; Yamaguchi, E.; Arisawa, M. Cloning of the Candida glabrata TRP1 and HIS3 genes, and construction of their disruptant strains by sequential integrative transformation. Gene 1995, 165, 203–206. [Google Scholar] [CrossRef]
- Nakayama, H.; Ueno, K.; Uno, J.; Nagi, M.; Tanabe, K.; Aoyama, T.; Chibana, H.; Bard, M. Growth defects resulting from inhibiting ERG20 and RAM2 in Candida glabrata. FEMS Microbiol. Lett. 2011, 317, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Boone, C.; Sommer, S.S.; Hensel, A.; Bussey, H. Yeast KRE genes provide evidence for a pathway of cell wall beta-glucan assembly. J. Cell Biol. 1990, 110, 1833–1843. [Google Scholar] [CrossRef] [PubMed]
- Srikantha, T.; Daniels, K.J.; Pujol, C.; Kim, E.; Soll, D.R. Identification of genes upregulated by the transcription factor Bcr1 that are involved in impermeability, impenetrability, and drug resistance of Candida albicans a/α biofilms. Eukaryot. Cell 2013, 12, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-H.; Miyazaki, T.; Tsai, H.-F.; Bennett, J.E. The bZip transcription factor Cgap1p is involved in multidrug resistance and required for activation of multidrug transporter gene CgFLR1 in Candida glabrata. Gene 2007, 386, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Sanguinetti, M.; De Bernardis, F.; Torelli, R.; Posteraro, B.; Vandeputte, P.; Sanglard, D. Loss of mitochondrial functions associated with azole resistance in Candida glabrata results in enhanced virulence in mice. Antimicrob. Agents Chemother. 2011, 55, 1852–1860. [Google Scholar] [CrossRef] [PubMed]
- Mathé, L.; Van Dijck, P. Recent insights into Candida albicans biofilm resistance mechanisms. Curr. Genet. 2013, 59, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Marco, F.; Pfaller, M.A.; Messer, S.A.; Jones, R.N. Activity of MK-0991 (L-743,872), a new echinocandin, compared with those of LY303366 and four other antifungal agents tested against blood stream isolates of Candida spp. Diagn. Microbiol. Infect. Dis. 1998, 32, 33–37. [Google Scholar] [CrossRef]
- Kuhn, D.M.; George, T.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Antifungal susceptibility of Candida biofilms: Unique efficacy of amphotericin B lipid formulations and echinocandins. Antimicrob. Agents Chemother. 2002, 46, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Scorzoni, L.; de Paula e Silva, A.C.A.; Marcos, C.M.; Assato, P.A.; de Melo, W.C.; de Oliveira, H.C.; Costa-Orlandi, C.B.; Mendes-Giannini, M.J.; Fusco-Almeida, A.M. Antifungal therapy: New advances in the understanding and treatment of mycosis. Front. Microbiol. 2017, 8, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Pierce, C.G.; Srinivasan, A.; Uppuluri, P.; Ramasubramanian, A.K.; López-Ribot, J.L. Antifungal therapy with an emphasis on biofilms. Curr. Opin. Pharmacol. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical practice guideline for the management of Candidiasis: 2016 update by the infectious diseases society of America. Clin. Infect. Dis. 2015, 62, e1–e50. [Google Scholar] [CrossRef] [PubMed]
- Canuto, M.M.; Rodero, F.G. Antifungal drug resistance to azoles and polyenes. Lancet Infect. Dis. 2002, 2, 550–563. [Google Scholar] [CrossRef]
- Rex, J.H.J.; Walsh, T.J.; Sobel, J.D.J.; Filler, S.G.; Pappas, P.G.; Dismukes, W.E.; Edwards, J.E. Practice guidelines for the treatment of candidiasis. Clin. Infect. Dis. 2000, 30, 662–678. [Google Scholar] [CrossRef] [PubMed]
- Schmalreck, A.F.; Willinger, B.; Haase, G.; Blum, G.; Lass-Flörl, C.; Fegeler, W.; Becker, K.; Antifungal susceptibility testing-AFST study group. Species and susceptibility distribution of 1062 clinical yeast isolates to azoles, echinocandins, flucytosine and amphotericin B from a multi-centre study. Mycoses 2012, 55, e124–e137. [Google Scholar] [CrossRef] [PubMed]
- Perlin, D.S. Mechanisms of echinocandin antifungal drug resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 1–11. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Real-time PCR expression profiling of BGL2, XOG1, GAS2, FKS1, FKS2, KNH1, UGP1, and MNN2 genes in biofilm cells of Candida glabrata ATCC2001 (reference strain, in bold) (a), C. glabrata 562123 (b), and C. glabrata 534784 (c) in the presence of antifungal drugs. The heatmap was generated by a log transformation of the RT-qPCR data and the fold-change (FC) expression determined through 2−ΔCT. The numerical scale on the right represents the FC. (Control: non-treated cells).
Figure 1.
Real-time PCR expression profiling of BGL2, XOG1, GAS2, FKS1, FKS2, KNH1, UGP1, and MNN2 genes in biofilm cells of Candida glabrata ATCC2001 (reference strain, in bold) (a), C. glabrata 562123 (b), and C. glabrata 534784 (c) in the presence of antifungal drugs. The heatmap was generated by a log transformation of the RT-qPCR data and the fold-change (FC) expression determined through 2−ΔCT. The numerical scale on the right represents the FC. (Control: non-treated cells).


{kind=link}
Table 1.
Minimum biofilm eradicatory concentrations (MBEC) for the Candida glabrata strains of fluconazole (Flu), amphotericin B (AmB), caspofungin (Csf), and micafungin (Mcf) (mg/L).
Table 1.
Minimum biofilm eradicatory concentrations (MBEC) for the Candida glabrata strains of fluconazole (Flu), amphotericin B (AmB), caspofungin (Csf), and micafungin (Mcf) (mg/L).
| Origin | Strain | Flu | AmB | Csf | Mcf |
|---|---|---|---|---|---|
| Reference (Wild Type) | ATCC2001 | >1250 | 4 | 2.5–3 | 16–17 |
| Urinary Tract | 562123 | 625 | 2 | 0.5–1 | 16–17 |
| Vaginal Tract | 534784 | >1250 | 2 | 2.5–3 | 5.5–6 |
Bold: concentrations applied to the pre-formed biofilms.
Table 2.
Primers, targets used, and specific function of the genes used for the expression analysis.
Table 2.
Primers, targets used, and specific function of the genes used for the expression analysis.
| Sequence (5′ → 3′) | Primer | Target | Properties and Proposed Function a |
|---|---|---|---|
| 5′-GGC AAG AAA CTG GAC AGA GC-3′ | F | BGL2 | β-1,3-glucanosyltransferase activity; glucan endo-β-1,3-d-glucosidase activity |
| 5′-GGA AAA CTT GGG TCC TGC TG-3′ | R | ||
| 5′-GTC CTA ACC TTG CAC ACC AG-3′ | F | FKS1 | β-1,3-d-glucan synthase activity |
| 5′-CTA CGC CCA AAC ATC AGC-3′ | R | ||
| 5′-GGG TCA CTG TGA AAT GTT-3 | F | FKS2 | β-1,3-d-glucan synthase activity |
| 5′-GTA GAC GGG TTC GGA TT-3 | R | ||
| 5′-ACC AGT CGT ACC ATT ACC GG-3′ | F | GAS2 | β-1,3-glucanosyltransferase activity |
| 5′-CCT GCC CAA CTT CTA ACA GC-3′ | R | ||
| 5′-CGG TGC CAA CGG TTA CTA-3′ | F | KNH1 | β-1,6-d-glucan biosynthetic process |
| 5′-GTG ACA CGG GTT TCA GGA-3′ | R | ||
| 5′-AAT CGC ACA AGG CAG AGA-3′ | F | UGP1 | β-1,6-d-glucan biosynthetic process |
| 5′-ACT TGG GCG ACT TCC AAT-3′ | R | ||
| 5′-GGT GAG TTG CAA CGT GAC AT-3′ | F | XOG1 | Glucan endo-β-1,6 and 1,3-glucosidase activity |
| 5′-ATT CGG TTA AAG CGG CAC TC-3′ | R | ||
| 5′-GAA GCC TGA TGG TGG TGA-3′ | F | MNN2 | α-mannosyltransferase biosynthetic process |
| 5′-ATT GGG CGA TGA CCT TCT-3′ | R | ||
| 5′-GTT GAC CGA GGC TCC AAT GA-3′ | F | ACT1 | Housekeeping gene |
| 5′-CAC CGT CAC CAG AGT CCA AA-3′ | R |
a CGD: Candida Genome Database [63]; F: forward; R: reverse.
Table 3.
Real-time PCR expression profiling of BGL2, FKS1, FKS2, GAS2, KNH1, UGP1, XOG1, and MNN2 genes in biofilm cells of C. glabrata ATCC2001, C. glabrata 562123, C. glabrata 534784 with and without antifungal treatment (FC: 2−ΔCT). The significance of the FC results was determined by comparing the treated groups with the non-treated ones (controls) (* p < 0.05; ** p < 0.001; *** p < 0.0005; **** p < 0.0001).
Table 3.
Real-time PCR expression profiling of BGL2, FKS1, FKS2, GAS2, KNH1, UGP1, XOG1, and MNN2 genes in biofilm cells of C. glabrata ATCC2001, C. glabrata 562123, C. glabrata 534784 with and without antifungal treatment (FC: 2−ΔCT). The significance of the FC results was determined by comparing the treated groups with the non-treated ones (controls) (* p < 0.05; ** p < 0.001; *** p < 0.0005; **** p < 0.0001).
| Gene | Candida glabrata ATCC2001 | Candida glabrata 562123 | Candida glabrata 534784 | |
|---|---|---|---|---|
| Fold-Change | ||||
| BGL2 | Non treated | 2.23 | 2.77 | 1.90 |
| Flu | 1.41 **** | 4.01 **** | 3.66 **** | |
| AmB | 2.04 **** | 1.71 **** | 3.85 **** | |
| Csf | 2.00 **** | 3.82 **** | 5.98 **** | |
| Mcf | 5.13 **** | 10.58 **** | 13.49 **** | |
| XOG1 | Non treated | 0.57 | 0.15 | 0.46 |
| Flu | 0.96 **** | 0.54 **** | 2.35 **** | |
| AmB | 0.37 **** | 0.10 *** | 0.27 **** | |
| Csf | 1.08 **** | 1.42 **** | 5.45 **** | |
| Mcf | 6.54 **** | 7.89 **** | 12.38 **** | |
| GAS2 | Non treated | 5.34 | 1.67 | 1.39 |
| Flu | 0.76 **** | 2.73 **** | 2.26 **** | |
| AmB | 3.02 **** | 1.56 **** | 1.95 **** | |
| Csf | 5.72 **** | 2.08 **** | 3.39 **** | |
| Mcf | 3.99 **** | 7.43 **** | 8.18 **** | |
| FKS1 | Non treated | 0.11 | 0.11 | 0.17 |
| Flu | 0.22 **** | 0.65 **** | 1.07 **** | |
| AmB | 0.16 **** | 0.08 **** | 0.19 (ns) | |
| Csf | 0.07 *** | 0.20 **** | 1.27 **** | |
| Mcf | 0.49 **** | 0.94 **** | 3.55 **** | |
| FKS2 | Non treated | 0.14 | 0.20 | 0.27 |
| Flu | 0.61 **** | 0.64 **** | 1.77 **** | |
| AmB | 0.06 *** | 0.19 * | 1.05 **** | |
| Csf | 0.28 **** | 0.55 **** | 1.66 **** | |
| Mcf | 0.43 **** | 2.29 **** | 7.50 **** | |
| KNH1 | Non treated | 0.06 | 0.20 | 0.22 |
| Flu | 0.50 **** | 0.41 **** | 2.08 **** | |
| AmB | 0.51 **** | 0.87 **** | 0.94 *** | |
| Csf | 0.24 **** | 0.21 (ns) | 0.72 **** | |
| Mcf | 0.45 **** | 1.33 **** | 1.43 **** | |
| UGP1 | Non treated | 0.002 | 0.07 | 0.20 |
| Flu | 0.10 **** | 0.15 **** | 0.55 **** | |
| AmB | 0.17 **** | 0.05 **** | 0.46 **** | |
| Csf | 0.01 (ns) | 0.06 ** | 0.33 **** | |
| Mcf | 0.04 ** | 0.21 **** | 3.17 **** | |
| MNN2 | Non treated | 0.02 | 0.18 | 0.13 |
| Flu | 0.19 **** | 0.34 **** | 1.17 **** | |
| AmB | 0.13 **** | 0.24 **** | 0.71 **** | |
| Csf | 0.18 **** | 0.13 *** | 1.40 **** | |
| Mcf | 0.31 **** | 1.21 **** | 7.03 **** | |
(ns, non-significant; Non-treated, controls).
Table 4.
Pearson Correlation Coefficient (r) determined for the expression profiles of BGL2, FKS1, FKS2, GAS2, KNH1, UGP1, XOG1, and MNN2 genes in biofilm cells of C. glabrata ATCC2001, C. glabrata 562123, C. glabrata 534784, in the presence or absence of antifungal drugs.
Table 4.
Pearson Correlation Coefficient (r) determined for the expression profiles of BGL2, FKS1, FKS2, GAS2, KNH1, UGP1, XOG1, and MNN2 genes in biofilm cells of C. glabrata ATCC2001, C. glabrata 562123, C. glabrata 534784, in the presence or absence of antifungal drugs.
| Gene | ATCC2001 vs. 562123 | ATCC2001 vs. 534784 | 562123 vs. 534784 |
|---|---|---|---|
| BGL2 | 0.9100 | 0.9145 | 0.946 |
| XOG1 | 0.9965 | 0.9459 | 0.9646 |
| FKS1 | 0.8947 | 0.8723 | 0.8778 |
| FKS2 | 0.5074 | 0.4481 | 0.9937 |
| GAS2 | −0.0514 | 0.1091 | 0.9663 |
| KNH1 | 0.6427 | 0.8107 | 0.3697 |
| UGP1 | −0.1091 | -0.1122 | 0.8519 |
| MNN2 | 0.7924 | 0.8618 | 0.9728 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rodrigues, C.F.; Henriques, M. Portrait of Matrix Gene Expression in Candida glabrata Biofilms with Stress Induced by Different Drugs. Genes 2018, 9, 205. https://doi.org/10.3390/genes9040205
AMA Style
Rodrigues CF, Henriques M. Portrait of Matrix Gene Expression in Candida glabrata Biofilms with Stress Induced by Different Drugs. Genes. 2018; 9(4):205. https://doi.org/10.3390/genes9040205
Chicago/Turabian StyleRodrigues, Célia F., and Mariana Henriques. 2018. "Portrait of Matrix Gene Expression in Candida glabrata Biofilms with Stress Induced by Different Drugs" Genes 9, no. 4: 205. https://doi.org/10.3390/genes9040205
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.