Horizontal Transfer of Symbiosis Genes within and Between Rhizobial Genera: Occurrence and Importance

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Framework and Assumptions of the Study
3. Lateral Transfer of Symbiosis Genes
3.1. Rhizobia Associated with the Caesalpiniodeae
3.2. Rhizobia Associated with the Papilionoideae
3.2.1. The Inverted Repeat-Lacking Clade (IRLC)
3.2.2. Papilionoideae with Indeterminate Nodules Excluding the IRLC
3.2.3. Papilionoideae with Determinate Nodules
4. Recombination of Symbiotic Islands
5. Occurrence and Importance of Horizontal Transfer of Rhizobial Symbiosis Genes
Author Contributions
Funding
Conflicts of Interest
References
- The Legume Phylogeny Working Group (LPWG). A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny. Taxon 2017, 66, 44–77. [Google Scholar]
- Sprent, J.I.; Ardley, J.K.; James, E.K. From North to South: A latitudinal look at legume nodulation processes. S. Afr. J. Bot. 2013, 89, 31–41. [Google Scholar] [CrossRef]
- Betti, M.; Garcia-Calderón, M.; Perez-Delgado, C.M.; Credali, A.; Estivill, G.; Galván, F.; Vega, J.M.; Márquez, A.J. Glutamine synthetase in legumes: Recent advances in enzyme structure and functional genomics. Int. J. Mol. Sci. 2012, 13, 7994–8024. [Google Scholar] [CrossRef] [PubMed]
- Seabra, A.R.; Pereira, P.A.; Becker, J.D.; Carvalho, H.G. Inhibition of glutamine synthetase by phosphinothricin leads to transcriptome reprogramming in root nodules of Medicago truncatula. Mol. Plant–Microbe Interact. 2012, 25, 976–992. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A.; Andrews, M. Evolution of tree nutrition. Tree Physiol. 2010, 30, 1050–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, M.; Raven, J.A.; Lea, P.J. Do plants need nitrate? The mechanisms by which nitrogen form affects plants. Ann. Appl. Biol. 2013, 163, 174–199. [Google Scholar] [CrossRef]
- Andrews, M.; Scholefield, D.; Abberton, M.T.; McKenzie, B.A.; Hodge, S.; Raven, J.A. Use of white clover as an alternative to nitrogen fertilizer for dairy pastures in nitrate vulnerable zones in the UK: Productivity, environmental impact and economic considerations. Ann. Appl. Biol. 2007, 151, 11–23. [Google Scholar] [CrossRef]
- Jackson, L.E.; Burger, M.; Cavagnaro, T.R. Roots, nitrogen transformations, and ecosystem services. Ann. Rev. Plant Biol. 2008, 59, 341–363. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; James, E.K.; Sprent, J.I.; Boddey, R.M.; Gross, E.; Dos Reis, F.B., Jr. Nitrogen fixation in legumes and actinorhizal plants in natural ecosystems: Values obtained using 15N natural abundance. Plant Ecol. Divers. 2011, 4, 131–140. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Menge, D.N.L.; Reed, S.C.; Cleveland, C.C. Biological nitrogen fixation: Rates, patterns and ecological controls in terrestrial ecosystems. Philos. Trans. R. Soc. B 2013, 368, 20130119. [Google Scholar] [CrossRef] [PubMed]
- Peck, M.C.; Fischer, R.F.; Long, S.R. Diverse flavonoids stimulate NodD1 binding to nod gene promoters in Simorhizobium meliloti. J. Bacteriol. 2006, 188, 5417–5427. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yang, S.; Tang, F.; Zhu, H. Symbiosis specificity in the legume-rhizobial mutualism. Cell Microbiol. 2012, 14, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-W.; Murray, J.D. The role of flavonoids in nodulation host-range specificity: An update. Plants 2016, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.; Downie, J.A. Coordinating nodule morphogenesis with rhizobial infection in legumes. Ann. Rev. Plant Biol. 2008, 59, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Roche, P.; Maillet, F.; Plazanet, C.; Debellé, F.; Ferro, M.; Truchet, G.; Promé, J.-C.; Dénarié, J. The common nodABC genes of Rhizobium meliloti are host-range determinants. Proc. Natl. Acad. Sci. USA 1996, 93, 15305–15310. [Google Scholar] [CrossRef] [PubMed]
- Perret, X.; Staehelin, C.; Broughton, W.J. Molecular basis of symbiotic promiscuity. Microbiol. Mol. Biol. Rev. 2000, 64, 180–201. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; Andrews, M.E. Specificity in legume rhizobia symbioses. Int. J. Mol. Sci. 2017, 18, 705. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.Y.Y.; Ridgway, H.J.; James, T.K.; James, E.K.; Chen, W.-M.; Sprent, J.I.; Young, J.P.W.; Andrews, M. Burkholderia sp. induces functional nodules on the South African invasive legume Dipogon lignosus (Phaseoleae) in New Zealand soils. Microb. Ecol. 2014, 68, 542–555. [Google Scholar] [PubMed]
- Sullivan, J.T.; Patrick, H.N.; Lowther, W.L.; Scott, D.B.; Ronson, C.W. Nodulating strains of Rhizobium loti arise through chromosomal gene transfer in the environment. Proc. Natl. Acad. Sci. USA 1995, 92, 8985–8989. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.T.; Ronson, C.W. Evolution of rhizobia by acquisition of a 500-kb symbiosis island that integrates into a phe-tRNA gene. Proc. Natl. Acad. Sci. USA 1998, 95, 5145–5149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, S.P.; Gyaneshwar, P.; Vinuesa, P.; Farruggia, F.T.; Andrews, M.; Humphry, D.; Elliott, G.N.; Nelson, A.; Orr, C.; Pettitt, D.; et al. Nodulation of Sesbania species by Rhizobium (Agrobacterium) strain IRBG74 and other rhizobia. Environ. Microbiol. 2009, 11, 2510–2525. [Google Scholar] [CrossRef] [PubMed]
- Remigi, P.; Zhu, J.; Young, J.P.W.; Masson-Boivin, C. Symbiosis within symbiosis: Evolving nitrogen-fixing legume symbionts. Trends Microbiol. 2016, 24, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Mergaert, P.; Uchiumi, T.; Alunni, B.; Evanno, G.; Cheron, A.; Catrice, O.; Mausset, A.-E.; Barloy-Hubler, F.; Galibert, F.; Kondorosi, A.; et al. Eukaryotic control on bacterial cell cycle and differentiation in the Rhizobium-legume symbiosis. Proc. Natl. Acad. Sci. USA 2006, 103, 5230–5235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA 7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 37, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP: Phylogenetic Analysis Using Parsimony; Version 3.1; Smithsonian Institute: Washington, DC, USA, 1991. [Google Scholar]
- Farris, J.S.; Källersjö, M.; Kluge, A.G.; Bult, C. Testing significance of incongruence. Cladistics 1994, 10, 315–319. [Google Scholar] [CrossRef]
- Rincón-Rosales, R.; Lloret, L.; Ponce, E.; Martínez-Romero, E. Rhizobia with different symbiotic efficiencies nodulate Acaciella angustissima in Mexico, including Sinorhizobium chiapanecum sp. nov. which has common symbiotic genes with Sinorhizobium mexicanum. FEMS Microbiol. Ecol. 2009, 67, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.W.; Penttinen, P.; Chen, Y.X.; Chen, Q.; Zhang, X. Symbiotic efficiency and phylogeny of the rhizobia isolated from Leucaena leucocephala in arid-hot river valley area in Panxi, Sichuan, China. Appl. Microbiol. Biotchnol. 2013, 97, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Platero, R.; James, E.K.; Rios, C.; Iriarte, A.; Sandes, L.; Zabaleta, M.; Battistoni, F.; Fabiano, E. Novel Cupriavidus strains isolated from root nodules of native Uruguayan Mimosa species. Appl. Environ. Microbiol. 2016, 82, 3150–3164. [Google Scholar] [CrossRef] [PubMed]
- Elliott, G.N.; Chou, J.-H.; Chen, W.-M.; Bloemberg, G.V.; Bontemps, C.; Martínez-Romero, E.; Velázquez, E.; Young, J.P.W.; Sprent, J.I.; James, E.K. Burkholderia spp. are the most competitive symbionts of Mimosa, particularly under N-limited conditions. Environ. Microbiol. 2009, 11, 762–778. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Wei, S.; Wang, F.; James, E.K.; Guo, X.Y.; Zagar, C.; Xia, L.G.; Dong, X.; Wang, Y.P. Burkholderia and Cupriavidus spp. are the preferred symbionts of Mimosa spp. in Southern China. FEMS Microbiol. Ecol. 2012, 80, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Baraúna, A.C.; Rouws, L.F.M.; Simoes-Araujo, J.L.; dos Reis, F.B., Jr.; Iannetta, P.P.M.; Maluk, M.; Goi, S.R.; Reis, V.M.; James, E.K.; Zilli, J.E. Rhizobium altiplani sp. nov., isolated from effective nodules on Mimosa pudica growing in untypically alkaline soil in Central Brazil. Int. J. Syst. Evol. Microbiol. 2016, 66, 1–7. [Google Scholar]
- Rivas, R.; Velázquez, E.; Willems, A.; Vizcaíno, N.; Subba-Rao, N.S.; Mateos, P.F.; Gillis, M.; Dazzo, F.B.; Martínez-Molina, E. A new species of Devosia that forms a unique nitrogen-fixing root-nodule symbiosis with the aquatic legume Neptunia natans (L.f.) Druce. Appl. Environ. Microbiol. 2002, 68, 5217–5222. [Google Scholar] [CrossRef] [PubMed]
- Sankhla, I.S.; Tak, N.; Meghwal, R.R.; Choudhary, S.; Tak, A.; Rathi, S.; Sprent, J.I.; James, E.K.; Gehlot, H.S. Molecular characterization of nitrogen fixing microsymbionts from root nodules of Vachellia (Acacia) jacquemontii, a native legume from the Thar Desert of India. Plant Soil 2017, 410, 21–40. [Google Scholar] [CrossRef]
- Cordero, I.; Ruiz-Díez, B.; de la Peña, T.C.; Balaguer, L.; Lucas, M.M.; Rincón, A.; Pueyo, J.J. Rhizobial diversity, symbiotic effectiveness and structure of nodules of Vachellia macracantha. Soil Biol. Biochem. 2016, 96, 39–54. [Google Scholar] [CrossRef]
- Degefu, T.; Wolde-meskel, E.; Frostegård, Å. Phylogenetic multilocus sequence analysis identifies seven novel Ensifer genospecies isolated from a less-well-explored biogeographical region in East Africa. Int. J. Syst. Evol. Microbiol. 2012, 62, 2286–2295. [Google Scholar] [CrossRef] [PubMed]
- Bontemps, C.; Elliott, G.N.; Simon, M.F.; dos Reis, F.B., Jr.; Gross, E.; Lawton, R.C.; Neto, N.E.; de Fátima Loureiro, M.; de Faria, S.M.; Sprent, J.I.; et al. Burkholderia species are ancient symbionts of legumes. Mol. Ecol. 2010, 19, 44–52. [Google Scholar] [PubMed]
- Bontemps, C.; Rogel, M.A.; Wiechmann, A.; Mussabekova, A.; Moody, S.; Simon, M.F.; Moulin, L.; Elliott, G.N.; Lacercat-Didier, L.; Dasilva, C.; et al. Endemic Mimosa species from Mexico prefer alphaproteobacterial rhizobial symbionts. New Phytol. 2016, 209, 319–333. [Google Scholar] [CrossRef] [PubMed]
- De Castro Pires, R.; dos Reis Júnior, F.B.; Zilli, J.E.; Fisher, D.; Hoffmann, A.; James, E.K.; Simon, M.F. Soil characteristics determine the rhizobia in association with different species of Mimosa in central Brazil. Plant Soil 2018, 423, 411–428. [Google Scholar] [CrossRef]
- Bournaud, C.; de Faria, S.M.; dos Santos, J.M.F.; Tisseyre, P.; Silva, M.; Chaintreuil, C.; Gross, E.; James, E.K.; Prin, Y.; Moulin, L. Burkholderia species are the most common and preferred nodulating symbionts of the Piptadenia group (Tribe Mimoseae). PLoS ONE 2013, 8, e63478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-de los Santos, P.; Palmer, M.; Chávez-Ramírez, B.; Beukes, C.; Steenkamp, E.T.; Briscoe, L.; Khan, N.; Maluk, M.; Lafos, M.; Humm, E.; et al. Whole genome analyses suggest that Burkholderia sensu lato contains two additional novel genera (Mycetohabitans gen. nov., and Trinickia gen. nov.): Implications for the evolution of diazotrophy and nodulation in the Burkholderiaceae. Genes 2018, 9, 268. [Google Scholar]
- Rivas, R.; Laranjo, M.; Mateos, P.F.; Oliveira, S.; Martinez-Molina, E.; Velázquez, E. Strains of Mesorhizobium amorphae and Mesorhizobium tianshanense, carrying symbiotic genes of common chickpea endosymbiotic species, constitute a novel biovar (ciceri) capable of nodulating Cicer arietinum. Lett. Appl. Microbiol. 2007, 44, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Laranjo, M.; Alexandre, A.; Rivas, R.; Velázquez, E.; Young, J.P.W.; Oliveira, S. Chickpea rhizobia symbiosis genes are highly conserved across multiple Mesorhizobium species. FEMS Microbiol. Ecol. 2008, 66, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Laranjo, M.; Young, J.P.W.; Oliveira, S. Multilocus sequence analysis reveals multiple symbiovars within Mesorhizobium species. Syst. Appl. Microbiol. 2012, 35, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Lou, K.; Jin, X.; Mao, P.H.; Wang, E.T.; Tian, C.F.; Sui, X.H.; Chen, W.F.; Chen, W.X. Distinctive Mesorhizobium populations associated with Cicer arietinum L. in alkaline soils of Xinjiang, China. Plant Soil 2012, 353, 123–134. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, X.; Guo, C.; de Lajudie, P.; Singh, R.P.; Wang, E.; Chen, W. Mesorhizobium muleiense and Mesorhizobium gsp. nov. are symbionts of Cicer arietinum L. in alkaline soils of Gansu, Northwest China. Plant Soil 2017, 410, 103–112. [Google Scholar] [CrossRef]
- Tena, W.; Wolde-Meskel, E.; Degefu, T.; Walley, F. Genetic and phenotypic diversity of rhizobia nodulating chickpea (Cicer arietinum L.) in soils from southern and central Ethiopia. Can. J. Microbiol. 2017, 63, 690–707. [Google Scholar] [CrossRef] [PubMed]
- Armas-Carpote, N.; Pérez-Yépez, J.; Martínez-Hidalgo, P.; Garzón-Machado, V.; del Arco-Aguilar, M.; Velázquez, E.; Léon-Barrios, M. Core and symbiotic genes reveal nine Mesorhizobium genospecies and three symbiotic lineages among the rhizobia nodulating Cicer canariense in its natural habitat (La Palma, Canary Islands). Syst. Appl. Microbiol. 2014, 37, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Mutch, L.A.; Young, J.P.W. Diversity and specificity of Rhizobium leguminosarum biovar viciae on wild and cultivated legumes. Mol. Ecol. 2004, 13, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.F.; Wang, E.T.; Wu, L.J.; Han, T.X.; Chen, W.F.; Gu, C.T.; Gu, J.G.; Chen, W.X. Rhizobium fabae sp. nov., a bacterium that nodulates Vicia faba. Int. J. Syst. Evol. Microbiol. 2008, 58, 2871–2875. [Google Scholar] [CrossRef] [PubMed]
- Santillana, N.; Ramírez-Bahena, M.H.; García-Fraile, P.; Velázquez, E.; Zúñiga, D. Phylogenetic diversity based on rrs, atpD, recA genes and 16S–23S intergenic sequence analyses of rhizobial strains isolated from Vicia faba and Pisum sativum in Peru. Arch. Microbiol. 2008, 189, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Saidi, S.; Ramírez-Bahena, M.H.; Santillana, N.; Zúñiga, D.; Álvarez-Martínez, E.R.; Peix, A.; Mhamdi, R.; Velázquez, E. Rhizobium laguerreae sp. nov. nodulates Vicia faba on several continents. Int. J. Syst. Evol. Microbiol. 2014, 64, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Zheng, W.T.; Everall, I.; Young, J.P.W.; Zhang, X.X.; Tian, C.F.; Sui, X.H.; Wang, E.T.; Chen, W.X. Rhizobium anhuiense sp. nov., isolated from effective nodules of Vicia faba and Pisum sativum. Int. J. Syst. Evol. Microbiol. 2015, 65, 2960–2967. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Young, J.P.W.; Everall, I.; Clercx, P.; Willems, A.; Braun, M.S.; Wink, M. Average nucleotide identity of genome sequences supports the description of Rhizobium lentis sp. nov., Rhizobium bangladeshense sp. nov. and Rhizobium binae sp.nov from lentil (Lens culinaris) nodules. Int. J. Syst. Evol. Microbiol. 2015, 65, 3037–3045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, T.X.; Wang, E.T.; Wu, L.J.; Chen, W.F.; Gu, J.G.; Gu, C.T.; Tian, C.F.; Chen, W.X. Rhizobium multihospitium sp. nov., isolated from multiple legume species native of Xinjiang, China. Int. J. Syst. Evol. Microbiol. 2008, 58, 1693–1699. [Google Scholar] [CrossRef] [PubMed]
- Gnat, S.; Malek, W.; Oleńska, E.; Wdowiak-Wróbel, S.; Kalita, M.; Lotocka, B.; Wójcik, M. Phylogeny of symbiotic genes and the symbiotic properties of rhizobia specific to Astragalus glycyphyllos L. PLoS ONE 2015, 10, e0141504. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.A.; Li, L.; Wei, G.; Räsänen, L.; Lindström, K. Evolution and taxonomy of native mesorhizobia nodulating medicinal Glycyrrhiza species in China. Syst. Appl. Microbiol. 2016, 39, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.S.; Zhao, L.F.; Kong, Z.Y.; Yang, W.Q.; Lindström, K.; Wang, E.T.; Wei, G.H. Diversity of endophytic bacteria within nodules of the Sphaerophysa salsula in different regions of Loess Plateau in China. FEMS Microbiol. Ecol. 2011, 76, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Chen, W.; Young, J.P.W.; Bontemps, C. A new clade of Mesorhizobium nodulating Alhagi sparsifolia. Syst. Appl. Microbiol. 2009, 32, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.L.; Chen, W.F.; Wang, E.T.; Guan, S.H.; Yan, X.R.; Chen, W.X. Genetic diversity and biogeography of rhizobia associated with Caragana species in three ecological regions of China. Syst. Appl. Microbiol. 2009, 32, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Marek-Kozaczuk, M.; Leszcz, A.; Wielbo, J.; Wdowiak-Wróbel, S.; Skorupska, A. Rhizobium pisi sv. trifolii K3.22 harboring nod genes of the Rhizobium leguminosarum sv. trifolii cluster. Syst. Appl. Microbiol. 2013, 36, 252–258. [Google Scholar] [PubMed]
- Zhang, J.J.; Jing, X.Y.; de Lajudie, P.; Ma, C.; He, P.X.; Singh, R.P.; Chen, W.F.; Wang, E.T. Association of white clover (Trifolium repens L.) with rhizobia of sv. trifolii belonging to three genomic species in alkaline soils in North and East China. Plant Soil 2016, 407, 417–427. [Google Scholar]
- Nandaseena, K.G.; O’Hara, G.W.; Tiwari, R.P.; Howieson, J.G. Rapid in situ evolution of nodulating strains for Biserrula pelecinus L. through lateral transfer of a symbiosis island from the original mesorhizobial inoculant. Appl. Environ. Microbiol. 2006, 72, 7365–7367. [Google Scholar] [CrossRef] [PubMed]
- Nandaseena, K.G.; O’Hara, G.W.; Tiwari, R.P.; Sezmiş, E.; Howieson, J.G. In situ lateral transfer of symbiosis islands results in rapid evolution of diverse competitive strains of mesorhizobia suboptimal in symbiotic nitrogen fixation on the pasture legume Biserrula pelecinus L. Environ. Microbiol. 2007, 9, 2496–2511. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, B.; Van Cauwenberghe, J.; Chimphango, S.; Stirton, C.; Honnay, O.; Smets, E.; Muasya, A.M. Recombination and horizontal transfer of nodulation and ACC deaminase (acdS) genes within Alpha- and Betaproreobacteria nodulating legumes of the Cape Fynbos biome. FEMS Microbiol. Ecol. 2015, 91, fiv118. [Google Scholar] [CrossRef] [PubMed]
- Sy, A.; Giraud, E.; Jourand, P.; Garcia, N.; Willems, A.; de Lajudie, P.; Prin, Y.; Neyra, M.; Gillis, M.; Boivin-Masson, C.; et al. Methylotrophic Methylobacterium bacteria nodulate and fix nitrogen in symbiosis with legumes. J. Bacteriol. 2001, 183, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Ardley, J.K.; Parker, M.A.; De Meyer, S.E.; Trengove, R.D.; O’Hara, G.W.; Reeve, W.G.; Yates, R.J.; Dilworth, M.J.; Willems, A.; Howieson, J.G. Microvirga lupini sp. nov., Microvirga lotononidis sp. nov. and Microvirga zambiensis sp. nov. are alphaproteobacterial root-nodule bacteria that specifically nodulate and fix nitrogen with geographically and taxonomically separate legume hosts. Int. J. Syst. Evol. Microbiol. 2012, 62, 2579–2588. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, B.; Dlodlo, O.; Chimphango, S.; Stirton, C.; Schrire, B.; Boatwright, J.S.; Honnay, O.; Smets, E.; Sprent, J.; James, E.K.; et al. Symbiotic diversity, specificity and distribution of rhizobia in native legumes of the Core Cape Subregion (South Africa). FEMS Microbiol. Ecol. 2015, 91, fiv118. [Google Scholar] [CrossRef] [PubMed]
- Vinuesa, P.; Léon-Barrios, M.; Silva, C.; Willems, A.; Jarabo-Lorenzo, A.; Pérez-Galdona, R.; Werner, D.; Martínez-Romero, E. Bradyrhizobium canariense sp. nov., an acid-tolerant endosymbiont that nodulates endemic genistoid legumes (Papilionoideae: Genisteae) from the Canary Islands, along with Bradyrhizobium japonicum bv. genistearum, Bradyrhizobium genospecies alpha and Bradyrhizobium genospecies beta. Int. J. Syst. Evol. Microbiol. 2005, 55, 569–575. [Google Scholar] [PubMed]
- Horn, K.; Parker, I.M.; Malek, W.; Rodríguez-Echeverria, S.; Matthew, A.P. Disparate origins of Bradyrhizobium symbionts for invasive populations of Cytisus scoparius (Leguminosae) in North America. FEMS Microbiol. Ecol. 2014, 89, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Chahboune, R.; Barrijal, S.; Moreno, S.; Bedmar, E.J. Characterization of Bradyrhizobium species isolated from root nodules of Cytisus villosus grown in Morocco. Syst. Appl. Microbiol. 2011, 34, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Cobo-Díaz, J.F.; Martínez-Hidalgo, P.; Fernández-González, A.J.; Martínez-Molina, E.; Toro, N.; Velázquez, E.; Fernández-López, M. The endemic Genista versicolor from Sierra Nevada National Park in Spain is nodulated by putative new Bradyrhizobium species and a novel symbiovar (sierranevadense). Syst. Appl. Microbiol. 2014, 37, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, E.; Valverde, A.; Rivas, R.; Gomis, V.; Peix, A.; Gantois, I.; Igual, J.M.; León-Barrios, M.; Willems, A.; Mateos, P.F.; et al. Strains nodulating Lupinus albus on different continents belong to several new chromosomal and symbiotic lineages within Bradyrhizobium. Antonie Van Leeuwenhoek 2010, 97, 363–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarabo-Lorenzo, A.; Pérez-Galdona, R.; Donate-Correa, J.; Rivas, R.; Velázquez, E.; Hernández, M.; Temprano, F.; Martínez-Molina, E.; Ruiz-Argüeso, T.; Léon-Barrios, M. Genetic diversity of Bradyrhizobial populations from diverse geographic origins that nodulate Lupinus spp. and Ornithopus spp. Syst. Appl. Microbiol. 2003, 26, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Stępkowski, T.; Żak, M.; Moulin, L.; Króliczak, J.; Golińska, B.; Narożna, D.; Safronova, V.I.; Mądrzak, C.J. Bradyrhizobium canariense and Bradyrhizobium japonicum are the two dominant rhizobium species in root nodules of lupin and serradella plants growing in Europe. Syst. Appl. Microbiol. 2011, 34, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, M.E.; Willems, A.; Abril, A.; Planchuelo, A.-M.; Rivas, R.; Ludeña, D.; Mateos, P.F.; Martínez-Molina, E.; Velázquez, E. Nodulation of Lupinus albus by strains of Ochrobactrum lupini sp. nov. Appl. Environ. Microbiol. 2005, 71, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- Durán, D.; Rey, L.; Sánchez-Cañizares, C.; Navarro, A.; Imperial, J.; Ruiz-Argueso, T. Genetic diversity of indigenous rhizobial symbionts of the Lupinus mariae-josephae endemism form alkaline-limed soils within its area of distribution in Eastern Spain. Syst. Appl. Microbiol. 2013, 36, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Bourebaba, Y.; Durán, D.; Boulila, F.; Ahnia, H.; Boulila, A.; Temprano, F.; Palacios, J.M.; Imperial, J.; Ruiz-Argüeso, T.; Rey, L. Diversity of Bradyrhizobium strains nodulating Lupinus micranthus on both sides of the Western Mediterranean: Algeria and Spain. Syst. Appl. Microbiol. 2016, 39, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Beukes, C.W.; Venter, S.N.; Law, I.J.; Phalane, F.L.; Steenkamp, E.T. South African papilionoid legumes are nodulated by diverse Burkholderia with unique nodulation and nitrogen-fixation loci. PLoS ONE 2013, 8, e68406. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Kong, Z.; Chen, W.; Wei, G. Genetic diversity and symbiotic evolution of rhizobia from root nodules of Coronilla varia. Syst. Appl. Microbiol. 2013, 36, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Stępkowski, T.; Moulin, L.; Krzyżańska, A.; McInnes, A.; Law, I.J.; Howieson, J. European origin of Bradyrhizobium populations infecting lupins and serradella in soils of Western Australia and South Africa. Appl. Environ. Microbiol. 2005, 71, 7041–7052. [Google Scholar] [CrossRef] [PubMed]
- Tak, N.; Awasthi, E.; Bissa, G.; Meghwal, R.R.; James, E.K.; Sprent, J.S.; Gehlot, H.S. Multi locus sequence analysis and symbiotic characterization of novel Ensifer strains nodulating Tephrosia spp. in the Indian Thar Desert. Syst. Appl. Microbiol. 2016, 39, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, B.; Van Cauwenberghe, J.; Verstraete, B.; Chimphango, S.; Stirton, C.; Honnay, O.; Smets, E.; Sprent, J.; James, E.K.; Muasya, A.M. Characterization of the papilionoid-Burkholderia interaction in the Fynbos biome: The diversity and distribution of beta-rhizobia nodulating Podalyria calyptrata (Fabaceae, Podalyrieae). Syst. Appl. Microbiol. 2016, 39, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, X.; Liu, Y.; Wang, E.T.; Ren, C.; Liu, W.; Xu, H.; Wu, H.; Jiang, N.; Li, Y.; et al. Genetic diversity and community structure of rhizobia nodulating Sesbania cannabina in saline-alkaline soils. Syst. Appl. Microbiol. 2016, 39, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.W.; Heenan, P.B.; De Meyer, S.E.; Willems, A.; Andrews, M. Diverse novel mesorhizobia nodulate New Zealand native Sophora species. Syst. Appl. Microbiol. 2015, 38, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.D.; Heenan, P.B.; De Meyer, S.E.; James, T.K.; Chen, W.-M.; Morton, J.D.; Andrews, M. Genetic diversity and nitrogen fixation of mesorhizobia symbionts of New Zealand endemic Sophora species. N. Z. J. Bot. 2017, 55. [Google Scholar] [CrossRef]
- Jiao, Y.S.; Liu, Y.H.; Yan, H.; Wang, E.T.; Tian, C.F.; Chen, W.X.; Guo, B.L.; Chen, W.F. Rhizobial diversity and nodulation characteristics of the extremely promiscuous legume Sophora flavescens. Mol. Plant Microbe Interact. 2015, 28, 1338–1352. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, X.; Huo, H.; Yuan, G.; Sun, Y.; Zhang, D.; Cao, Y.; Xu, L.; Wei, G. Phylogenetic diversity of Ammopiptanthus rhizobia and distribution of rhizobia associated with Ammopiptanthus mongolicus in diverse regions of Northwest China. Microb. Ecol. 2016, 72, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Donate-Correa, J.; Léon-Barrios, M.; Hernández, M.; Pérez-Galdona, R.; del Arco-Aguilar, M. Different Mesorhizobium species sharing the same symbiotic genes nodulate the shrub legume Anagyris latifolia. Syst. Appl. Microbiol. 2007, 30, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Stepkowski, T.; Banasiewicz, J.; Granada, C.E.; Andrews, M.; Passaglia, L.M.P. Phylogeny and phylogeography of rhizobial symbionts nodulating legumes of the tribe Genisteae. Genes 2018, 9, 163. [Google Scholar] [CrossRef] [PubMed]
- De Meyer, S.E.; Tan, H.W.; Andrews, M.; Heenan, P.B.; Willems, A. Mesorhizobium calcicola sp. nov., Msorhizobium waitakense sp. nov., Mesorhizobium sophorae sp. nov., Mesorhizobium newzealandense sp. nov. and Mesorhizobium kowhaii sp. nov. isolated from Sophora root nodules in New Zealand. Int. J. Syst. Evol. Microbiol. 2016, 66, 786–795. [Google Scholar] [CrossRef] [PubMed]
- De Meyer, S.E.; Tan, H.W.; Heenan, P.B.; Andrews, M.; Willems, A. Mesorhizobium waimense sp. nov. isolated from Sophora longicarinata root nodules and Mesorhizobium cantuariense sp. nov. isolated from Sophora microphylla root nodules in New Zealand. Int. J. Syst. Evol. Microbiol. 2015, 65, 3419–3426. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Deng, Z.; Yang, W.; Cao, Y.; Wang, E.; Wei, G. Diverse rhizobia associated with Sophora alopecuroides grown in different regions of Loess Plateau in China. Syst. Appl. Microbiol. 2010, 33, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Wang, R.; Zhang, X.X.; Young, J.P.W.; Wang, E.T.; Sui, X.H.; Chen, W.X. Bradyrhizobium guangdongense sp. nov. and Bradyrhizobium guangxiense sp. nov., isolated from effective nodules of peanut. Int. J. Syst. Evol. Microbiol. 2015, 65, 4655–4661. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hu, M.; Ma, H.; Wang, Y.; Wang, E.T.; Zhou, Z.; Gu, J. Genetic diversity and distribution of bradyrhizobia nodulating peanut in acid-neutral soils in Guangdong Province. Syst. Appl. Microbiol. 2016, 39, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.W.; Zou, L.; Penttinen, P.; Zeng, X.; Liu, M.; Zhao, K.; Chen, C.; Chen, Y.X.; Zhang, X. Diversity and phylogeny of rhizobia associated with Desmodium spp. in Panxi, Sichuan, China. Syst. Appl. Microbiol. 2016, 39, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Barcellos, F.G.; Menna, P.; da Silva Batista, J.S.; Hungria, M. Evidence of horizontal transfer of symbiotic genes from a Bradyrhizobium japonicum inoculant strain to indigenous diazotrophs Sinorhizobium (Ensifer) fredii and Bradyrhizobium elkanii in a Brazilian savannah soil. Appl. Environ. Microbiol. 2007, 73, 2635–2643. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Li, Y., Jr.; Chen, W.F.; Wang, E.T.; Tian, C.F.; Li, Q.Q.; Zhang, Y.Z.; Sui, X.H.; Chen, W.X. Biodiversity and biogeography of rhizobia associated with soybean plants grown in the North China Plain. Appl. Environ. Microbiol. 2011, 77, 6331–6342. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Wang, E.T.; Chang, Y.L.; Zhang, Y.Z.; Zhang, Y.M.; Sui, X.H.; Chen, W.F.; Chen, W.X. Ensifer sojae sp. nov., isolated from root nodules of Glycine max grown in saline-alkaline soils. Int. J. Syst. Evol. Microbiol. 2011, 61, 1981–1988. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Wang, R.; Zhang, Y.M.; Liu, H.C.; Chen, W.F.; Wang, E.T.; Sui, X.H.; Chen, W.X. Bradyrhizobium daqingense sp. nov., isolated from soybean nodules. Int. J. Syst. Evol. Microbiol. 2013, 63, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Habibi, S.; Ayubi, A.G.; Ohkama-Ohtsu, N.; Sekimoto, H.; Yokoyama, T. Genetic characterization of soybean rhizobia isolated from different ecological zones in north-eastern Afghanistan. Microbes Environ. 2017, 32, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Fan, M.; Zhang, D.; Yang, R.; Zhang, F.; Xu, L.; Wei, X.; Shen, Y.; Wei, G. Distribution and diversity of rhizobia associated with wild soybean (Glycine soja Sieb. & Zucc.) in Northwest China. Syst. Appl. Microbiol. 2014, 37, 449–456. [Google Scholar] [PubMed]
- Laguerre, G.; Nour, S.M.; Macheret, V.; Sanjuan, J.; Drouin, P.; Amarger, N. Classification of rhizobia based on nodC and nifH gene analysis reveals a close phylogenetic relationship among Phaseolus vulgaris symbionts. Microbiology 2001, 147, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Valverde, A.; Igual, J.M.; Peix, A.; Cervantes, E.; Velázquez, E. Rhizobium lusitanum sp. nov. a bacterium that nodulates Phaseolus vulgaris. Int. J. Syst. Evol. Microbiol. 2006, 56, 2631–2637. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Man, C.X.; Wang, E.T.; Chen, W.X. Diversity of rhizobia and interactions among the host legumes and rhizobial genotypes in an agricultural-forestry ecosystem. Plant Soil 2009, 314, 169–182. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, E.-T.; Zhao, L.; Chen, W.-M.; Wei, G.-H. Diversity and distribution of rhizobia nodulated with Phaseolus vulgaris in two ecoregions of China. Soil Biol. Biochem. 2014, 78, 128–137. [Google Scholar] [CrossRef]
- Wang, L.; Cao, Y.; Wang, E.T.; Qiao, Y.J.; Jiao, S.; Liu, Z.S.; Zhao, L.; Wei, G.H. Biodiversity and biogeography of rhizobia associated with common bean (Phaseolus vulgaris L.) in Shaanxi Province. Syst. Appl. Microbiol. 2016, 39, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Han, L.L.; Wang, E.T.; Lu, Y.L.; Zhang, Y.F.; Sui, X.H.; Chen, W.F.; Chen, W.X. Bradyrhizobium spp. and Sinorhizobium fredii are predominant in root nodules of Vigna angularis, a native legume crop in the subtropical region of China. J. Microbiol. 2009, 47, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Risal, C.P.; Djedidi, S.; Dhakal, D.; Ohkama-Ohtsu, N.; Sekimoto, H.; Yokoyama, T. Phylogenetic diversity and symbiotic functioning in mungbean (Vigna radiata L. Wilczek) bradyrhizobia from contrast agro-ecological regions of Nepal. Syst. Appl. Microbiol. 2012, 35, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Steenkamp, E.T.; Stępkowski, T.; Przymusiak, A.; Botha, W.J.; Law, I.J. Cowpea and peanut in southern Africa are nodulated by diverse Bradyrhizobium strains harboring nodulation genes that belong to the large pantropical clade common in Africa. Mol. Phylogenet. Evol. 2008, 48, 1131–1144. [Google Scholar] [CrossRef] [PubMed]
- Radl, V.; Simões-Araújo, J.L.; Leite, J.; Passos, S.R.; Martins, L.M.V.; Xavier, G.R.; Rumjanek, N.G.; Baldani, J.I.; Zilli, J.E. Microvirga vignae sp. nov., a root nodule symbiotic bacterium isolated from cowpea grown in semi-arid Brazil. Int. J. Syst. Evol. Microbiol. 2014, 64, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Tampakaki, A.P.; Fotiadis, C.T.; Ntatsi, G.; Savvas, D. A novel symbiovar (aegeanense) of the genus Ensifer nodulates Vigna unguiculata. J. Sci. Food Agric. 2017, 97, 4314–4325. [Google Scholar] [CrossRef] [PubMed]
- Lorite, M.J.; Donate-Correa, J.; del Arco-Aguilar, M.; Galdona, R.P.; Sanjuán, J.; Léon-Barrios, M. Lotus endemic to the Canary Islands are nodulated by diverse and novel rhizobial species and symbiotypes. Syst. Appl. Microbiol. 2010, 33, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Lorite, M.J.; Muñoz, S.; Olivares, J.; Soto, M.J.; Sanjuán, J. Characterization of strains unlike Mesorhizobium loti that nodulate Lotus spp. in saline soils of Granada, Spain. Appl. Environ. Microbiol. 2010, 76, 4019–4026. [Google Scholar] [CrossRef] [PubMed]
- Estrella, M.J.; Muñoz, S.; Soto, M.J.; Ruiz, O.; Sanjuán, J. Genetic diversity and host range of rhizobia nodulating Lotus tenuis in typical soils of the Salado River basin (Argentina). Appl. Environ. Microbiol. 2009, 75, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Wang, E.T.; Tian, C.F.; Wang, F.Q.; Han, L.L.; Chen, W.F.; Chen, W.X. Bradyrhizobium elkanii, Bradyrhizobium yuanmingense and Bradyrhizobium japonicum are the main rhizobia associated with Vigna unguiculata and Vigna radiata in the subtropical region of China. FEMS Microbiol. Lett. 2008, 285, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Ampomah, O.Y.; Huss-Danell, K. Genetic diversity of root nodule bacteria nodulating Lotus corniculatus and Anthyllis vulneraria in Sweden. Syst. Appl. Microbiol. 2011, 34, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Cañizares, C.; Jorrín, B.; Durán, D.; Nadendla, S.; Albareda, M.; Rubio-Sanz, L.; Lanza, M.; González-Guerrero, M.; Prieto, R.I.; Brito, B.; et al. Genomic diversity in the endosymbiotic bacterium Rhizobium leguminosarum. Genes 2018, 9, 60. [Google Scholar] [CrossRef] [PubMed]
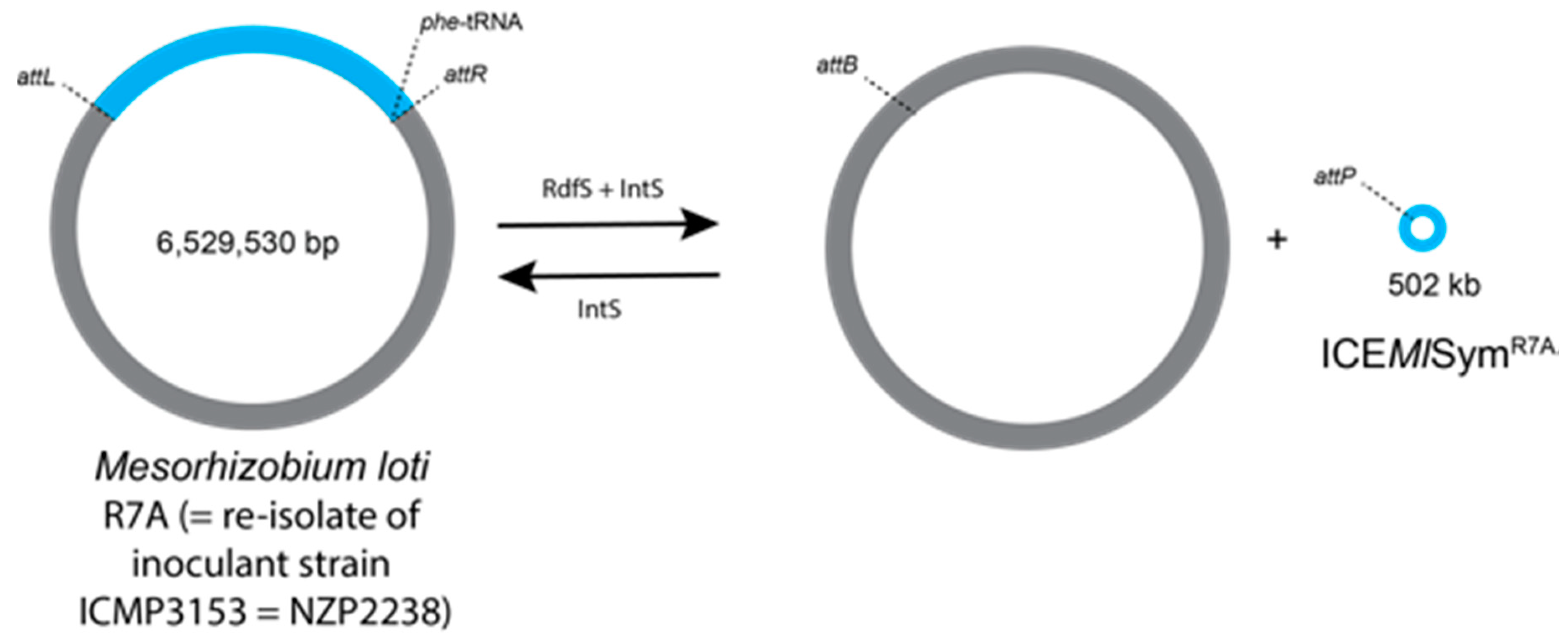
- Ramsay, J.P.; Sullivan, J.T.; Stuart, G.S.; Lamont, I.L.; Ronson, C.W. Excision and transfer of the Mesorhizobium loti R7A symbiosis island requires an integrase IntS, a novel recombination directionality factor RdfS, and a putative relaxase RlxS. Mol. Microbiol. 2006, 62, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.; Sullivan, J.; Ronson, C.; Tian, R.; Brau, L.; Munk, C.; Goodwin, L.; Han, C.; Woyke, T.; Reddy, T.; et al. Genome sequence of the Lotus spp. microsymbiont Mesorhizobium loti strain R7A. Stand. Genom. Sci. 2014, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Haskett, T.L.; Ramsay, J.P.; Bekuma, A.A.; Sullivan, J.T.; O’Hara, G.W.; Terpolilli, J.J. Evolutionary persistence of tripartite integrative and conjugative elements. Plasmid 2017, 92, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Haskett, T.; Wang, P.; Ramsay, J.; O’Hara, G.; Reeve, W.; Howieson, J.; Terpolilli, J. Complete genome sequence of Mesorhizobium ciceri strain CC1192, an efficient nitrogen-fixing microsymbiont of Cicer arietinum. Genome Announc. 2016, 4, e00516-16. [Google Scholar] [PubMed]
- Haskett, T.L.; Terpolilli, J.J.; Bekuma, A.; O’Hara, G.W.; Sullivan, J.T.; Wang, P.; Ronson, C.W.; Ramsay, J.P. Assembly and transfer of tripartite integrative and conjugative genetic elements. Proc. Natl. Acad. Sci. USA 2016, 113, 12268–12273. [Google Scholar] [CrossRef] [PubMed]
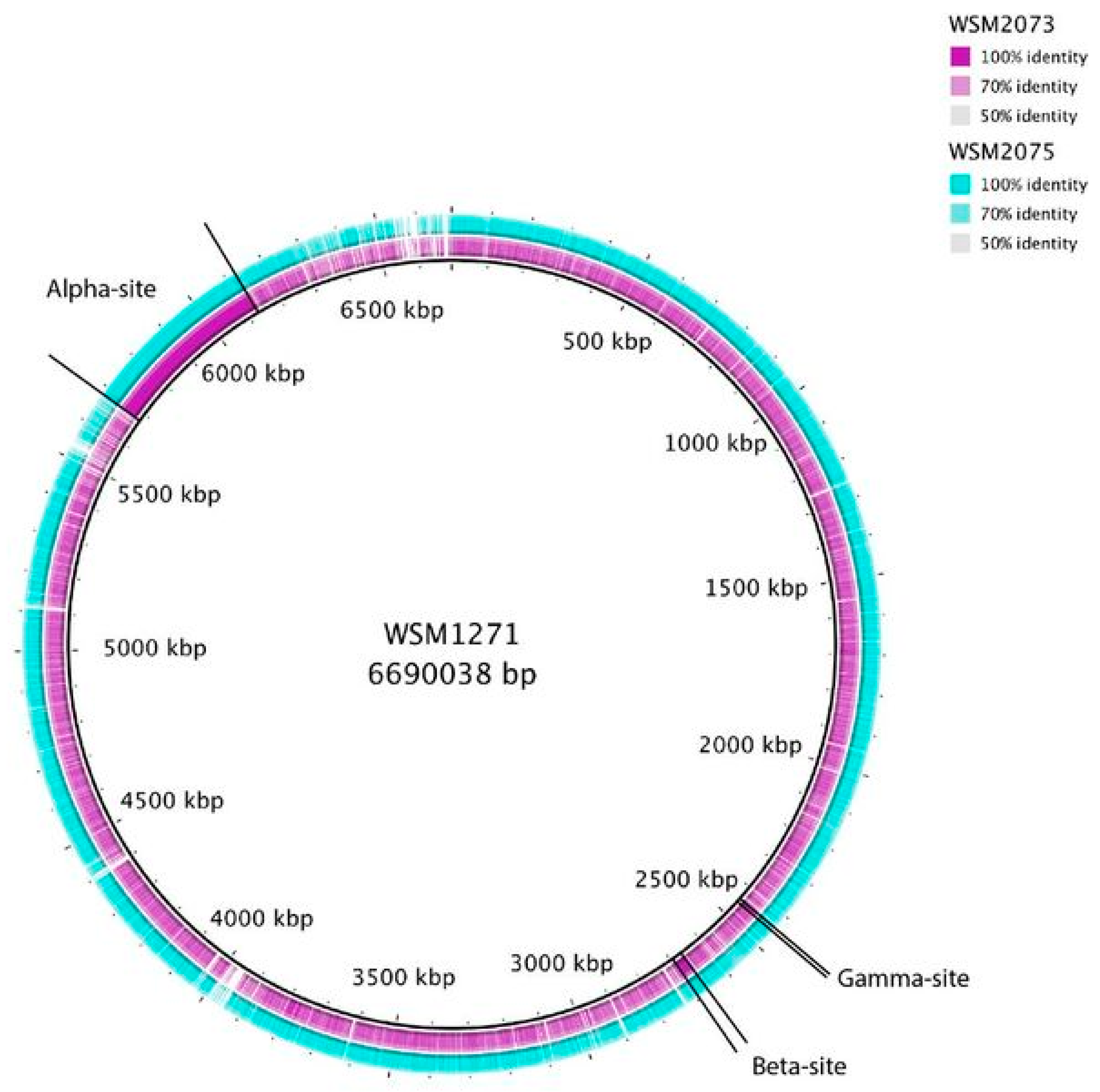
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryotic genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Caesalpinioideae Mimosoid Clade | Rhizobia |
|---|---|
| Acaciella angustissima | Ensifer chiapanecum ITTG S70T and Ensifer mexicanum ITTG R7T had different gyrA, nolR, recA. rpoB, and rrs gene sequences but similar nifH and nodA sequences [27] |
| Leucaena leucocephala | Ensifer isolates formed three clades in both 16S rRNA and recA phylogenetic trees but only one clade in both nifH and nodC trees [28] |
| Mimosa cruenta, Mimosa magentea, Mimosa ramulosa, Mimosa reptans, Mimosa schleidenii | Cupriavidus isolates separated into two groups on 16S rRNA, recA and gyrB sequences but grouped together on nifH and nodA sequences [29] |
| Mimosa diplotricha | Burkholderia caribensis TS182 characterized on 16S rRNA sequence grouped with Cupriavidus strains on nodA sequence [30] |
| M. diplotricha, Mimosa pudica | Burkholderia strains with diverse 16S rRNA gene sequences grouped together along with B. phymatum STM815T on nodA sequence [31] |
| M. pudica | Rhizobium altiplani BR 10423T had nifH and nodC sequences closely related (identical for nodC) to those of Rhizobium mesoamericanum CCGE 501T [32] |
| Neptunia natans | Devosia isolates characterized on 16S rRNA sequences had nifH and nodD sequences closely related to those of Rhizobium tropici CIAT899T [33] |
| Vachellia jacquemontii | Ensifer showed incongruence across all three of concatenated rrs-glnII-atpD-recA-dnaK, nifH, and nodA gene sequences [34] |
| Vachellia macracantha | Ensifer sequences for nifH and nodC were incongruent with those for 16S rRNA [35] |
| Vachellia seyal, Vachellia tortilis | Ensifer isolates separated into seven groups on the basis of 16S rRNA, recA, gyrB, rpoB, atpD, gap and pnp gene sequences but were closely related with respect to their nifH and nodC gene sequences [36]. |
| Papilionoideae Tribes and Genera | Rhizobia |
|---|---|
| Cicereae | |
| Cicer arietinum | Mesorhizobium ciceri, Mesorhizobium mediterraneum, Mesorhizobium muluense and Mesorhizobium spp. with diverse 16S rDNA, recA, atpD, glnII and gyrB sequences had similar nifH, nodA and nodC sequences [42,43,44,45,46,47] |
| Cicer canariense | Mesorhizobium with diverse 16S rRNA, recA and glnII sequences had similar nodC gene sequences [48] |
| Fabeae | |
| Lathyrus spp., Lens culinaris, Pisum sativum and Vicia spp. | Rhizobium fabeae, Rhizobium pisi, Rhizobium laguerreae, Rhizobium anhuiense, Rhizobium bangladeshense, Rhizobium binae, Rhizobium lentis and Rhizobium spp. with diverse 16S rRNA and recA, atpD and glnII sequences had similar nifH, nodA and nodC sequences [49,50,51,52,53,54] |
| Lathyrus odoratus, Vicia hirsuta | Rhizobium multihospitium isolates had nifH and nodD sequences 100% similar to those of Rhizobium lusitanum P1–7T and Devosia neptuniae J1T [55] |
| Galegeae | |
| Astragalus aksuensis, Astragalus sp., Oxytropis glabra, Oxytropis meinshausenii | R. multihospitium isolates had nifH and nodD sequences 100% similar to those of R. lusitanum P1–7T and D. neptuniae J1T [55] |
| Astragalus glycyphyllos | Mesorhizobium isolates showing 16S rRNA sequences similar to M. ciceri, Mesorhizobium amorphae or Mesorhizobium septentrionale formed one clearly separated, closely related cluster for nodA, nodC, nodH and nifH sequences [56] |
| Glycyrrhiza uralensis | Mesorhizobium concatenated rrs-recA-rpoB, nifH, nodA and nodC sequences were not congruent [57] |
| Sphaerophysa salsula | Mesorhizobium with diverse 16S rRNA sequences showed similar nifH sequences [58]Mesorhizobium and Rhizobium identified on 16S rRNA sequences showed similar nifH sequences [58] |
| Hedysareae | |
| Alhagi sparsifolia | Mesorhizobium isolates separated into three groups on the basis of their rrs, dnaK and dnaJ sequences but their nodA and nodC sequences were closely related [59] |
| Alhagi sp., Caragana jubata, Halimodendron halodendron, | R. multihospitium isolates had nifH and nodD sequences 100% similar to those of R. lusitanum P1-7T and D. neptuniae J1T [55] |
| Caragana bicolor, Caragana erinacea, Caragana franchetiana, Caragana intermedia | Mesorhizobium isolates with diverse 16S–23S IGS 16S rRNA sequences and one Bradyrhizobium isolate (16S rRNA) from C. intermedia had similar nodC sequences [60] |
| Trifolium | |
| Trifolium repens | R. pisi sv. trifolii K3.22 characterised on the basis of 16S rRNA, atpD, dnaK, glnA, gyrB, recA and rpoB sequences had nodA, nodB, nodC and nodD sequences with high similarity to those of Rhizobium leguminosarum sv. trifolii [61].Rhizobium spp. with diverse 16S rRNA and concatenated atpD-recA-glnII sequences had similar nifH and nodC sequences [62] |
| Papilionoideae Tribes (Genera) | Rhizobia |
|---|---|
| Crotalarieae | |
| Aspalathus sp. | Rhizobium isolate characterized on 16S rRNA and concatenated recA-atpD-gyrB-glnA sequences had nifH and concatenated nodA-B-C sequences closely related to those of Mesorhizobium [65] |
| Aspalathus astroites, Aspalathus aurantiaca, Aspalathus bracteata, Aspalathus ciliaris, Aspalathus cordata, Aspalathus ericifolia, Aspalathus spicata | Mesorhizobium phylogenetic relationships between concatenated recA-atpD-gyrB-glnA and nodA-B-C sequences were incongruent [65] |
| Crotalaria podocarpa | Methylobacterium nodulans ORS2060T nodA sequence groups with nodA sequences for Bradyrhizobium spp. [66] |
| Listia angolensis | Microvirga lotonidis WSM3557T and Microvirga zambiensis WSM3693T nodA sequences were identical and clustered with Bradyrhizobium, Burkholderia and Methylobacterium nodA sequences [67] |
| Rafnia triflora | Burkholderia isolate characterized on concatenated 16S rRNA-recA-atpD sequences had a nifH sequence closely related to those of Ensifer spp. [68] |
| Genisteae | |
| Argyrolobium lunare, Argyrolobium velutinum | Mesorhizobium phylogenetic relationships between concatenated recA-atpD-gyrB-glnA and nodA-nodB-nodC sequences were incongruent [65] |
| Cytisus proliferus | Bradyrhizobium with diverse 16S–23S rRNA, atpD, glnII and recA sequences showed similar nifH and nodC sequences [69] |
| Cytisus scoparius | Bradyrhizobium 16S rRNA, 23S rRNA, dnaK, gyrB, rplC, rpoB, nifD, nifH and nodC sequences indicated a highly heterogeneous ancestry [70] |
| Cytisus villosus | Bradyrhizobium with diverse 16S rRNA and concatenated glnII-recA sequences showed similar nifH and nodC sequences [71] |
| Genista versicolor | Bradyrhizobium with diverse 16S–23S ITS and atpD sequences showed similar nifH and nodC sequences for almost all strains [72] |
| Lupinus albus | Bradyrhizobium with diverse 16S–23S ITS and rrs and atpD sequences clustered together on nodC sequences [73] |
| L. albus, Lupinus angustifolius, Lupinus luteus, Lupinus sp. | Bradyrhizobium with diverse 16S–23S ITS and 16S rRNA sequences clustered together on nodC sequences [74] |
| L. albus, L. angustifolius, L. luteus | Bradyrhizobium with diverse concatenated atpD-glnII-recA sequences clustered together on nodA sequences [75] |
| Lupinus honoratus | Ochrobactrum lupini LUP21T nifH sequence showed 99.6% similarity to M. ciceri strains; its nodD sequence showed 86.4% similarity to Rhizobium etli CFN42T [76] |
| Lupinus mariae-josephae | Bradyrhizobium with diverse concatenated atpD-glnII-recA sequences separated into two distinct clusters on nodA and nodC sequences [77] |
| Lupinus micranthus | Bradyrhizobium with diverse concatenated 16S rRNA and concatenated atpD-gln11-recA sequences showed similar nodC gene sequences [78] |
| Lupinus texensis | Microvirga lupini Lut6T concatenated nifD-nifH sequence aligned close to R. etli CFN42T; its nodA sequence was placed in a clade that contained strains of Rhizobium, Mesorhizobium and Ensifer [67] |
| Hypocalypteae | |
| Hypocalyptus sophoroides, Hypocalyptus oxalidifolius, Hypocalyptus colutoides | Burkholderia phylogenies inferred from nifH and nodA sequences were incongruent; Burkholderia phylogenies inferred from nifH and nodA sequences were incongruent with those from 16S rRNA and recA sequences [79] |
| Loteae | |
| Coronilla varia | Mesorhizobium phylogenies for 16S rRNA, nifH and nodC sequences were incongruent [80] |
| Ornithopus compressus, Ornithopus sativus | Bradyrhizobium with diverse 16S–23S rRNA ITS and dnaK, atpD, glnII and recA sequences clustered together on nodA, nodZ and nolL sequences [75,81] |
| Millettieae | |
| Tephrosia falciformis, Tephrosia leptostachya, Tephrosia purpurea, Tephrosia villosa, Tephrosia wallichii | Ensifer 16S rRNA and concatenated recA-atpD-glnII-dnaK sequences grouped with Ensifer saheli LMG 7837T and Ensifer kotiensis LMG 19225T but nifH, nodA and nodC sequences clustered with Ensifer fredii USDA 205T [82] |
| Podalyrieae | |
| Cyclopia buxifolia, Cyclopia genistoides, Cyclopia glabra, Cyclopia intemedia, Cyclopia longifolia, Cyclopia maculata, Cyclopia meyeriana, Cyclopia pubescens, Cyclopia sessiflora, Cyclopia subternata | Burkholderia phylogenies inferred from nifH and nodA sequences were incongruent; phylogenies inferred from nifH and nodA sequences were incongruent with those from 16S rRNA and recA sequences [79] |
| Podalyria calyptrata | Burkholderia phylogenetic relationships between concatenated recA-atpD-gyrB-glnA and nodA-B-C sequences were largely incongruent [65]Burkholderia phylogenies inferred from nifH and nodA sequences were incongruent; phylogenies inferred from nifH and nodA sequences were incongruent with those from 16S rRNA and recA sequences [79]Burkholderia phylogenetic relationships between recA and nodA sequences were largely congruent but different branching patterns were observed among numerous isolates [83] |
| Virgilia oroboides | Burkholderia phylogenies inferred from nifH and nodA sequences were incongruent; phylogenies inferred from nifH and nodA sequences were incongruent with those from 16S rRNA and recA sequences [79] |
| Robineae | |
| R. pseudoacacia | R. multihospitium isolates had nifH and nodD sequences 100% similar to those of R. lusitanum P1–7T and D. neptuniae J1T [55] |
| Sesbanieae | |
| Sesbania cannabina | Rhizobium strain IRBG74 characterised on concatenated 16S rRNA–rpoB-fusA sequence harboured a sym-plasmid containing nifH and nodA genes similar to those of Ensifer strains that nodulate this legume [21]Rhizobium/Agrobacterium and Ensifer characterized on concatenated recA-atpD-glnII sequences had similar Ensifer nifH and nodA sequences [84] |
| Sesbania sesban | Ensifer isolates separated into three groups on the basis of concatenated 16S rRNA-recA-gyrB-rpoB-atpD-gap-pnp sequences but were closely related with respect to their nifH and nodC sequences [36] |
| Sophoreae | |
| Sophora alopecuroides | R. multihospitium isolates had nifH and nodD sequences 100% similar to those of R. lusitanum P1–7T and D. neptuniae J1T [55] |
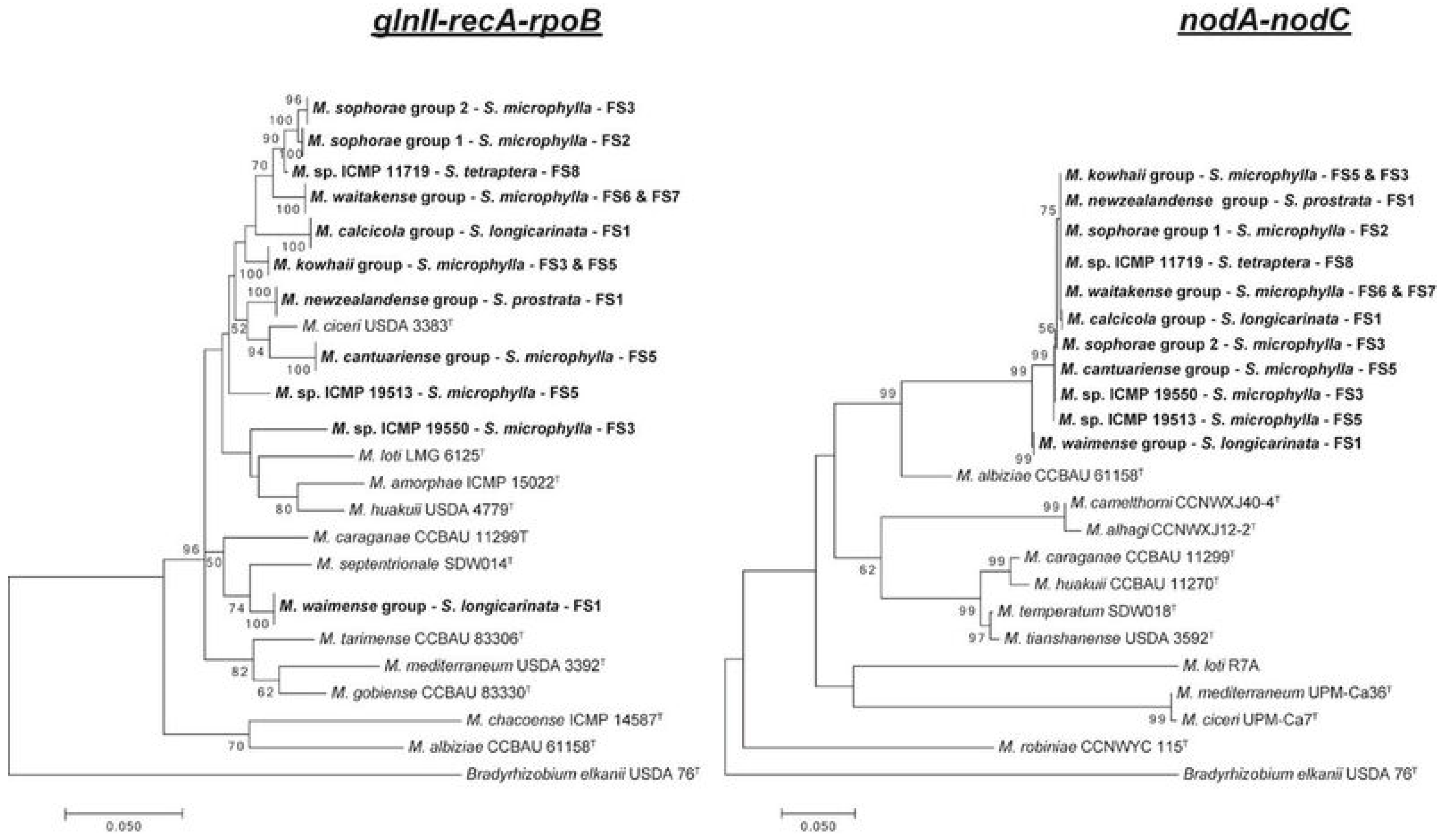
| Sophora chathamica, Sophora fulvida, Sophora godleyi, Sophora longicarinata, Sophora microphylla, Sophora prostrata, Sophora tetraptera | Mesorhizobium with diverse concatenated recA-glnII-rpoB sequences had similar nifH, nodA and nodC sequences [85,86] |
| Sophora flavescens | Rhizobium mongolense isolate characterized on concatenated atpD-glnII-recA sequences had nodC sequence similar to isolates characterized as M. septentrionale [87]. E. fredii isolate characterized on concatenated atpD-glnII-recA sequences had nodC sequence identical to Mesorhizobium temperatumT [87]. Phyllobacterium sophoraeT isolate characterized on concatenated atpD-glnII-recA sequences had nodC sequence closely related to M. septentrionaleT [87]. Mesorhizobium and Rhizobium phylogenetic relationships between concatenated atpD-glnII-recA and nodC sequences were incongruent [87] |
| Thermopsideae | |
| Ammopiptanthus nanus, Ammopiptanthus mongolicus | Ensifer arboris and Neorhizobium galegeae characterized on 16S rRNA and concatenated recA-atpD-rpoB-thrC sequences aligned with Ensifer meliloti ATCC9930T on nifH and nodC sequences [88]. Phyllobacterium giardinii characterized on 16S rRNA and concatenated recA-atpD-rpoB-thrC sequences aligned with R. leguminosarum sv. viciae USDA 2370T on nifH and nodC sequences [88]. Rhizobium/Agrobacterium radiobacter characterized on 16S rRNA and concatenated recA-atpD-rpoB-thrC sequences aligned with E. fredii USDA205T on nifH and nodC sequences [88] |
| Anagyris latifolia | Mesorhizobium isolates with diverse 16S–23S rDNA ITS, 16S rRNA and glnII sequences had identical nodC sequences closely related to Mesorhizobium tianshanense USDA 3592T [89] |
| Papilionoideae Tribes and Genera | Rhizobia |
|---|---|
| Dalbergieae | |
| Arachis hypogaea | Bradyrhizobium guangdongense CCBAU 51649T, Bradyrhizobium guangxiense CCBAU 53363T, Bradyrhizobium sp. P1237 and Bradyrhizobium sp. CH81 had identical nodA sequences [94]; Bradyrhizobium with diverse 16S–23S rRNA ITS and concatenated atpD-recA sequences showed similar nodA sequences [95] |
| Desmodieae | |
| Desmodium oldhami | Rhizobium characterized on 16S rRNA and concatenated recA-atpD-glnII sequences aligned with Ensifer sp. on nodC sequences [96] |
| Desmodium sequax | Rhizobium and Pararhizobium characterized on 16S rRNA and concatenated recA-atpD-glnII sequences aligned with Ensifer sp. on nodC sequences [96] |
| Phaseoleae | |
| Glycine max | Bradyrhizobium strains with clearly separated 16S rRNA sequences showed identical or similar nifH, nodC and nodY-nodA sequences [97]; Ensifer strain characterized on 16S rRNA sequence showed similar nifH, nodC and nodY-nodA sequences to B. japonicumT [97]; Bradyrhizobium with diverse 16S rRNA and concatenated recA-glnII-atpD sequences showed identical nifH and nodC sequences [98]; Ensifer with diverse 16S rRNA and concatenated recA-glnII-atpD sequences showed identical nifH and nodC sequences [98]; Ensifer sojae CCBAU 05684T and E. fredii USDA 205T showed identical nodC sequences [99]; Bradyrhizobium daqingense CCBAU 15774T, Bradyrhizobium liaonginenese USDA 3622T and B. japonicum USDA 6T showed identical nifH and nodC sequences [100]; Ensifer isolate classified on 16S rRNA sequence showed 99% similarity to Bradyrhizobium yuanmingense in nodD1 and nifD sequences [101] |
| Glycine soja | Ensifer and Rhizobium with diverse 16S rRNA and concatenated recA-atpD-glnII sequences formed a single Ensifer lineage on nifH and nodA sequences [102] |
| Phaseolus vulgaris | R. etli, Rhizobium gallicum, R. leguminosarum sv. phaseoli and Pararhizobium giardinii characterized on 16S rRNA sequences had similar nodC sequences and a strain characterized as Rhizobium aligned with E. meliloti on nodC sequence [103]; R. lusitanum P1–7T had nifH and nodC sequences similar to D. neptuniae LMG 21357T and R. tropici CIAT 899T [104]; R. etli and R. leguminosarum characterized on 16S rRNA sequences showed similar nifH and nodC sequences to R. etli CFN 42T [105]; Rhizobium with diverse 16S rRNA and concatenated atpD-glnII-recA sequences clustered together on nifH and nodC sequences [106]; Pararhizobium giardinii characterized on 16S rRNA and concatenated recA-glnII-atpD sequences aligned with Ensifer on nodC sequence [107] |
| Vigna angularis | Rhizobium characterized on 16S rRNA and concatenated atpD-recA sequences had a nodC sequence similar to Ensifer strains [108] |
| Vigna radiata | Bradyrhizobium characterized on sequences of the 16S rRNA, nodD1 and nifD genes and the ITS region aligned with Ensifer on nodA sequences [109] |
| Vigna unguiculata | Bradyrhizobium with diverse concatenated rrs-recA-glnII sequences showed similar nodA sequences [110]; Microvirga vignae BR3299T aligned with Mesorhizobium and Rhizobium on nifH sequence and Microvirga lotononidis, M. zambiensis, Bradyrhizobium, Burkholderia and Methylobacterium on nodA sequences [111]; E. fredii characterized on sequences of 16S rRNA, concatenated recA-glnII-gyrB-truA-thrA-SMc00019 and IGS were substantially diverged from E. fredii on nifH, nodC and rhcRST-1 sequences [112] |
| Psoraleae | |
| Otholobium bracteolatum, Otholobium hirtum, Otholobium virgatum, Otholobium zeyheri | Mesorhizobium phylogenetic relationships between concatenated recA-atpD-gyrB-glnA and nodA-B-C sequences were incongruent [65] |
| Psoralea asarina, Psoralea congesta, Psoralea laxa, Psoralea rigidula | Mesorhizobium phylogenetic relationships between concatenated recA-atpD-gyrB-glnA and nodA-B-C sequences were incongruent [65] |
| P. oligophylla | Mesorhizobium isolate characterized on concatenated 16S rRNA-recA-atpD sequence aligned closely to Burkholderia on nodA sequence [68] |
| Psoralea sp. | Mesorhizobium isolate characterized on concatenated 16S rRNA-recA-atpD sequence aligned closely to Ensifer on nifH sequence [68] |
| Loteae | |
| Lotus bertheloti, Lotus callis-viridis, Lotus corniculatus, Lotus campylocladus, Lotus pyranthus, Lotus sessifolius, L. tenuis | Mesorhizobium with diverse 16S rRNA, atpD and recA sequences clustered together on nodC gene sequences [113,114] |
| L. corniculatus | Transfer of symbiotic island between Mesorhizobium loti inoculum and indigenous Mesorhizobium strains [19,20] |
| L. frondosus, L. tenuis | R. multihospitium isolates had nifH and nodD sequences 100% similar to those of R. lusitanum P1–7T and D. neptuniae J1T [55] |
| L. tenuis | Mesorhizobium with diverse 16S rRNA sequences clustered together on nifH and nodC sequences [115] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrews, M.; De Meyer, S.; James, E.K.; Stępkowski, T.; Hodge, S.; Simon, M.F.; Young, J.P.W. Horizontal Transfer of Symbiosis Genes within and Between Rhizobial Genera: Occurrence and Importance. Genes 2018, 9, 321. https://doi.org/10.3390/genes9070321
Andrews M, De Meyer S, James EK, Stępkowski T, Hodge S, Simon MF, Young JPW. Horizontal Transfer of Symbiosis Genes within and Between Rhizobial Genera: Occurrence and Importance. Genes. 2018; 9(7):321. https://doi.org/10.3390/genes9070321
Chicago/Turabian StyleAndrews, Mitchell, Sofie De Meyer, Euan K. James, Tomasz Stępkowski, Simon Hodge, Marcelo F. Simon, and J. Peter W. Young. 2018. "Horizontal Transfer of Symbiosis Genes within and Between Rhizobial Genera: Occurrence and Importance" Genes 9, no. 7: 321. https://doi.org/10.3390/genes9070321