Heterologous Production of Flavour and Aroma Compounds in Saccharomyces cerevisiae


The Australian Wine Research Institute, P.O. Box 197, Glen Osmond, SA 5064, Australia
*
Author to whom correspondence should be addressed.
Genes 2018, 9(7), 326; https://doi.org/10.3390/genes9070326
Submission received: 28 May 2018
/
Revised: 21 June 2018
/
Accepted: 25 June 2018
/
Published: 28 June 2018
(This article belongs to the Special Issue Emerging Applications in Synthetic Biology)
{kind=link}
Abstract
:Over the last two decades, rapid progress in the field of synthetic biology has opened several avenues for the heterologous de novo production of complex biological compounds, such as biofuels, pharmaceuticals, and food additives in microbial hosts. This minireview addresses the usage of the yeast Saccharomyces cerevisiae as a microbial cell factory for the production of flavour and aroma compounds, thereby providing a path towards a sustainable and efficient means of producing what are normally rare, and often expensive plant-derived chemicals.
1. Introduction
Synthetic biology is one of the most rapidly evolving branches of the biological sciences. It allows the introduction of custom made genetic pathways into organisms that provide them the ability to display biological and/or biochemical properties over-and-above the original, wild type background. Numerous examples have been recently reported of the successful implementation of synthetic biology in microorganisms such as Saccharomyces cerevisiae, such as for the production of valuable biomedical compounds [1] or biofuels [2,3]. Many different microorganisms, like Yarowia lippolitica or Escherichia coli, have also been utilized as hosts for various synthetic biology applications [4,5]. However, because of the availability of a vast array of well-established molecular and genetic techniques, baker’s yeast S. cerevisiae continuously remains one of the most frequently used microorganisms in this rapidly growing field. In this minireview, attention will be focused on the utilization of S. cerevisiae for the heterologous production of flavour and aroma compounds, and their potential applications in food and beverage industries.
2. Heterologous Compounds Derived from Intermediates of the Shikimate Pathway
Vanillin, (4-hydroxy-3-methoxybenzaldehyde), is a plant secondary metabolite commonly extracted from the seed pods of the vanilla orchids Vanilla planifolia, Vanilla tahitensis, or Vanilla pompona. It is one of the most commonly used food and beverage flavouring additives, with over 16,000 tonnes of vanillin consumed worldwide each year [6]. However, the slow growth of the vanilla orchid combined with the relatively low content of vanillin in the mature seed pods, means that only a minor fraction (~0.25%) of the total production of this compound is actually derived from vanilla orchids [7]. Market demand for this flavouring agent is therefore mostly fulfilled through chemical synthesis from lignin and/or fossil hydrocarbons, processes that are regarded as environmentally unfriendly and not sustainable. Alternative means of non-synthetic vanillin production are therefore highly desirable [8].
Heterologous biosynthetic production of vanillin by S. cerevisiae is one of the most widely recognized examples of applying a synthetic biology system for the industrial production of a high value compound via a microbial cell factory. The metabolic pathway that allows yeast to produce vanillin was first developed nearly a decade ago, requiring the expression of four heterologous enzymes; 3-dehydroshikimate dehydratase (3DSD) from the dung mold Podospora pauciseta, an aromatic carboxylic acid reductase (ACAR) from a bacterium of the Nocardia genus, an O-methyltransferase from Homo sapiens (HsOMT), and a phosphopantetheinyl transferase from Corynebacterium glutamicum (EntD); the latter enzyme was required for activation of ACAR via phosphopantetheinylation [9] (Figure 1). Introduction of these foreign genes provided S. cerevisiae with the ability to convert the endogenous metabolic precursor 3-dehydroshikimate (3-DHS), an intermediate of the shikimate pathway for aromatic amino acid biosynthesis, into vanillin at concentrations approaching 45 mg/L.
However, these initial attempts at optimizing the production of vanillin were complicated by the reported toxicity of vanillin against many microorganisms, including yeast, where concentrations exceeding 50 mg/L can significantly inhibit growth [9], thereby imposing a ceiling on maximum vanillin production levels. The problem of vanillin toxicity was solved via the heterologous expression of a glycosyltransferase (UGT) from Arabidopsis thaliana, which converted vanillin into its less toxic and more soluble form, vanillin-b-d-glucoside (VG). This strategy, along with flux engineering of the native yeast metabolism, involving removal of the pyruvate decarboxylase (PDC1) and glutamate dehydrogenase (GDH1) genes, as well as overexpression of glutamate dehydrogenase (GDH2), resulted in a vanillin production level of 500 mg/L. However, despite this significant increase, these levels were still far below the theoretical yields predicted through metabolic modelling analysis [7]. To remove further metabolic bottlenecks, two potentially rate limiting heterologous steps in the vanillin synthesis pathway were addressed by additional gene overexpression; ACAR, which converts protocatechuic acid (PAC) to protocatechuic aldehyde (PAL), and HsOMT which catalyses the conversion of (PAL) to vanillin (Figure 1). While overexpression of ACAR had no effect on final VG production, a 30% improvement was observed in the HsOMT overexpressing strain [10].
In an alternative study, Strucko and colleagues [11] investigated the impact of different yeast strains on the VG production. By reconstituting heterologous VG biosynthesis pathway in two commonly used yeast strains, S. cerevisiae CEN.PK and S288c, it was observed that the S288c background was able to produce up to a 10-fold greater concentration of VG under the same fermentation conditions. As there are hundreds of different strains of S. cerevisiae available, this work clearly demonstrates that the original genetic background is highly important in regards of choosing a yeast strain for an efficient microbial cell factory.
Raspberry ketone is the primary compound that contributes to the aroma of raspberries (Rubus idaeus) [12,13], although it is also produced by many other plants, including cranberries, blackberries, and rhubarb [14]. Raspberry ketone extracted from natural sources is an expensive flavoring agent, costing as much as US $3000/kg. However, as seen for vanillin, raspberry ketone can also be derived utilizing chemical synthesis, with the synthetic version costing far less than the naturally-derived version (US $58/kg) [15]. In R. idaeus, raspberry ketone is produced via the diketide pathway, starting with condensation of 4-coumaryl-CoA and malonyl-CoA by benzalacetone synthase (BAS) to form 4-(4-hydroxyphenyl)but-3-en-2-one (benzalacetone) [16,17]. In a subsequent reaction, benzalacetone is reduced by NADPH-dependent benzalactone reductase (BAR), also known as raspberry ketone/zingerone synthase 1 (RiZS1), to form raspberry ketone [18].
S. cerevisiae lacks the diketide pathway, with phenylalanine and tyrosine representing the metabolic branch points for heterologous de novo production of RK (Figure 1). Primary attempts of heterologous expression of plant genes, 4-coumarate-coenzyme A ligase (4CL) and chalcone synthase (CHS), in this yeast for raspberry ketone production were not entirely successful. The recombinant yeast grown in media containing p-coumaric acid produced only very low levels of raspberry ketone [19]. However, in a recent study, Lee et al. [14] engineered S. cerevisiae to express the complete de novo pathway for heterologous production of raspberry ketone, without necessity of supplementation with the precursor; p-coumaric acid. The recombinant yeast strain was genetically modified to produce p-coumaric acid through the expression of the heterologous enzymes phenylalanine/tyrosine ammonia lyase from Rhodosporidium toruloides (RtPAL) (which provided both phenylalanine and tyrosine ammonia lyase activists), and cinnamate-4-hydroxylase from A. thaliana (AtC4H), a combination shown previously to drive p-coumaric acid production in S. cerevisiae [20,21]. Additional expression of a synthetic fusion protein that linked coumarate-CoA ligase 2 from Petroselinum crispum and benzalacetone synthase from Rheum palmatum (Pc4CL2-RpBAS), yielded a strain able to produce 3.5 mg/L of raspberry ketone [14]. Although levels of this highly valuable flavouring compound produced by the above recombinant strain are not yet adequate for its industrial application, this study paves the way for future development of S. cerevisiae cell factories for commercial raspberry ketone production.
Cinnamaldehyde is the main impact flavonoid found in the bark, lives and fruit of the cinnamon tree Cinnamomum zeylanicum [22,23]. It is the main organic compound that contributes the flavour and aroma of cinnamon and is commonly used as a flavouring compound in the food and beverage industries [24,25]. Many chemical methods exist to produce this flavonoid synthetically, however steam distillation of the oil derived from the cinnamon bark remains the most economical and commonly used method [26,27]. Cinnamaldehyde can be heterologously produced in S. cerevisiae through an extension of the shikimate pathway (via phenylalanine), through the activity of two heterologous enzymes; phenylalanine ammonia lyase 2 (PAL2) which converts phenylalanine into trans-cinnamic acid, and aryl carboxylic acid reductase (ACAR) which converts trans-cinnamic acid into cinnamaldehyde (Figure 1). As also seen for the production of vanillin, activation of ACAR was achieved by overexpression of a phosphopantetheinyl transferase [28]. Although successfully achieving a proof of concept for de novo production of cinnamaldehyde, only negligible amounts of this compound were produced, primarily due to a combination of cinnamaldehyde toxicity (~0.68 mM in S. cerevisiae) and off-target metabolism towards cinnamyl alcohol and hydrocinnamyl alcohol [28,29]. Thus, while heterologous production of cinnamaldehyde has been achieved, the transition to an industrial scale will require further metabolic optimization.
3. Heterologous Compounds Derived from Intermediates of the Mevalonate Pathway
3.1. Isoprenoids
Isoprenoids (also known as terpenes or terpenoids) are a large group of diverse organic compounds that encompass thousands of different chemical forms and which are mainly observed as the products of plants. Despite their chemical diversity, all isoprenoids are biosynthesized from the basic unit isoprene (C5H8), with different subclasses of compounds classified according to the number of isoprene units (hemiterpenes, monoterpenes, sesquiterpenes, and diterpenes with 1, 2, 3, and 4 isoprene units, respectively). Many of the compounds belonging to the terpene group are toxic and/or display unpleasant, irritant odours and are used by host plants to deter predatorial foraging and pathogen invasion [30]. However, a small selection of specific isoprenoinds have found application in variety of industries, including food and beverage as attractive flavouring additives.
3.1.1. Monoterpenes
Monoterpenes are all produced via the action of monoterpene synthases acting on geranyl pyrophosphate (GPP) as a common substrate. Each individual monoterpene synthase is therefore responsible for producing a specific monoterpene (or a combination of monoterpenes) from this common metabolic precursor [31].
Geraniol is one of the most in-demand monoterpenes, primarily due to its pleasant rose-like aroma. It is now an established additive in the food and beverage industries, where it is commonly used to enhance the flavours of a broad range of beverages, ice cream, candy, chewing gum, and many other products [32].
Due to its high sensory impact, geraniol is produced in negligible concentrations in most plants, such that the yields of final extracted compounds are very low [33]. While plant sources of geraniol are generally limiting, yeast has proven to be a very efficient cell factory for the heterologous production of this compound. S. cerevisiae lacks the ability to produce monoterpenes, however it produces the penultimate monoterpene precursor, GPP, as part of the mevalonate/ergosterol pathway [34,35] (Figure 1). Interestingly, despite lacking recognised monoterpene synthase activity, S. cerevisiae strains carrying mutations in farnesyl diphosphate synthase gene (ERG20) were shown to produce low levels of geraniol (1.3 mg/L), presumably through non-specific production due to high GPP levels [36]. Later, Iijima and colleagues isolated and characterized the specific geraniol synthase (GES) from sweet basil (Ocimum basilicum) that catalyses conversion of GPP to geraniol [37], unlocking the possibility of geraniol production in microbial hosts (Figure 1). Heterologous expression of this enzyme in S. cerevisiae allowed production of geraniol at levels up to 500 µg/L in the growth medium [34]. Following on from this, engineering of ERG20p via amino acid substitutions surrounding the catalytic site resulted in a strain able to produce around 5 mg/L of geraniol [38].
Geraniol synthase enzymes have now been identified and characterized from a large number of different plant sources [39,40]. This expanded catalogue of natural variation, combined with additional metabolic engineering, has been used to further improve heterologous geraniol production in S. cerevisiae. By introducing a fusion protein composed of geraniol synthase from Valeriana officinalis (tVoGES) that lacks plastid targeting signal and a mutant farnesyl diphosphate synthase ERG20p(F96W-N127W), in a strain with a metabolically-engineered high-flux mevalonate pathway, 239 mg/L of geraniol was achieved in fed-batch cultures [41,42,43]. However, this concentration was still deemed inadequate for industrialization of the system. In an elegant study Jiang and colleagues [44] have now achieved much higher levels of geraniol production through a combination bioprospecting for highly active GES proteins, site directed mutagenesis and enzyme structural analysis, which ultimately resulted in a strain able to produce over 0.5 g/L of geraniol. When this strain was further tested in fed-batch fermentation under carbon restricted conditions, it produced 1.68 g/L of geraniol, the highest level of this monoterpene produced by any eukaryotic organism reported to date [44].
In parallel to approaches that seek to produce the highest amounts of geraniol possible for purification as an additive, alternative studies have been conducted to investigate the in situ production of geraniol during wine fermentation [45]. Expression of GES from sweet basil resulted in the accumulation of more than 750 µg/L of geraniol in finished wine that was made from aromatically neutral grapes. Surprisingly, in additional to geraniol, the final wine also contained significant amounts of additional monoterpenes, including citronellol, linalool, and nerol, which were likely derived from indigenous enzymatic activities in the industrial wine yeast background. Although the amounts of geraniol produced by this recombinant industrial yeast were much lower compared to the heterologous systems described above, the levels of this, as well as other monoterpenes, were way above the olfactory sensory thresholds, positively contributing to overall sensory characteristics of wines by providing enhanced floral and fruity attributes [45].
Linalool can be naturally found in many fruits and flowers and is responsible for pleasant floral aromas. Heterologous production of this flavour compound in yeast was originally achieved via the expression of a linalool synthase (LIS) from Clarkia breweri, one of the earliest identified monoterpene synthases, and which specifically converts GPP to linalool [46,47] (Figure 1). While initial concentrations of linalool were very low (~22 µg/L), engineering of flux through the mevalonate pathway via overexpression of catalytic domain of Hmg1p (3-hydroxy-3-methylglutaryl coenzyme A reductase), combined with specific yeast strain selection, resulted in a 6-fold increase in linalool concentrations [48]. A subsequent study that used LIS from Lavandula angustifolia, combined with overexpression of HMG1 and downregulation of ERG9, resulted in levels of up to 95 µg/L of linalool [49]. However, yields remain well behind those observed for geraniol and further optimization of this system is required if industrially-relevant production levels are to be achieved.
As for geraniol, there have also been attempts to produce linalool in situ during fermentation instead of as a purified additive. Linalool and geraniol are the main impact molecules that confer dry-hopped flavour and aroma in specific styles of beer. In classical brewing, these monoterpenes originate from the flowers of the hop plant, which are added to the wort during fermentation. In a recent study, Denby et al. [50] screened for combination of linalool and geraniol synthases and gene promoters, that could be combined in brewing yeast to produce these monoterpenes at levels comparable to those achieved by standard dry-hopping during the beer fermentation process. A brewing strain of S. cerevisiae, that was metabolically engineered to produce higher levels of GPP, was equipped with a truncated LIS from Mentha citrata and full-length GES from Ocimum basilicum, yielding a self-flavouring brewing yeast able to produce beer with basic dry-hopped flavour profiles, without the necessity of adding hop flowers.
Limonene is another example of terpenoid that is commonly used as a flavouring agent [25]. It is mainly found in citrus fruit peels and used in food and beverage products such as candy and soft drinks [51]. With an annual production of over 60,000 tonnes [52], it is primarily obtained from citrus essential oils, which are byproducts from orange juice production, and may contain over 90% of this compound in the total pool of the compounds present [53,54]. As the price of natural limonene is relatively high, its production in microbial hosts would reduce the dependence on its industrial production from plant sources, such as citrus fruits.
The potential for heterologous production of limonene in S. cerevisiae has recently been reported, where it is based on the same metabolic principals as those for the production of geraniol, except for the final step in which GPP is converted to limonene by a limonene synthase (LMS) (Figure 1). However, the reported yields of limonene remain far lower than those of either geraniol or linanool, with a maximum of 0.12 mg/L (+)-limonene and 0.49 mg/L (−)-limonene produced in S. cerevisiae [55,56]. These low production levels suggest that major metabolic redesign and optimization are required for limonene production to be industrially applicable. However, as 27 limonene synthases, derived from 9 different plant families, have recently been characterized [57], this opens new possibilities for metabolic engineering and/or enzyme optimization in yeast, with potential to improve the yields of this valuable monoterpene.
Sabinene is another example of monoterpene that was reported to be produced in S. cerevisiae by means of heterologous gene expression. This monoterpene can be naturally found and extracted from the essential oils of many plants, including Hyptis pectinata Poit. (Lamiaceae) [58], or Zornia diphylla (L.) Pers, where it can reach more than 40% of the total oil extracted from whole plant [59]. In S. cerevisiae production of sabinene was achieved by heterologous expression of highly specific sabinene synthase (SAS) from Salvia pomifera [60] (Figure 1). By introduction of fusion protein ERG20p(F96W-N127W)-SpSASp nearly 1.9 mg/L of sabinene was produced by this recombinant yeast. Subsequent deletion of one copy of the wild type ERG20, and introduction of the second copy of fusion protein ERG20p(F96W-N127W)-SpSabS1 further improved sabinene production to 17.5 mg/L.
3.1.2. Sesquiterpenes
Sesquiterpenes are a diverse group of plant secondary metabolites which are generally derived from C-15 precursor; farnesyl pyrophosphate (FPP).
Valencene can be found in many citrus fruits [61,62] and is commonly and inexpensively extracted from the essential oil of Valencia oranges. Heterologous production of this sesquiterpene in S. cerevisiae was achieved by identification and expression of valencene synthase (VS), with variants of this enzyme having been characterized from many plants including Citrus paradisi (grapefruit), Citrus sinensis or Nootka cypress (Callitropsis nootkatensis) [63,64]. Expression of the C. paradisi VS in a S. cerevisiae strain (Figure 1), which was metabolically engineered to produce elevated levels of FPP [65], was shown to produce low levels of valencene (5.3 µg/L) [66].
Later, Farhi and colleagues [63] have altered metabolic flux through the mevalonate pathway for enhanced farnesyl pyrophosphate (FPP) accumulation by overexpression of tHMG in C. sinensis valencene synthase CsTPS1 expressing yeast strain, which led to 1.5-fold (ca. 40 to ca. 60 µg/L) increase in valencene production, compared to the expression of CsTPS1 alone. Introduction either A. thaliana short isoform of farnesyl diphosphate synthase (AtFDPS), or H. sapiens farnesyl diphosphate synthase (HsFDPS), further increased valencene production; the final strain carrying CsTPS1, tHMG and AtFDPS was able to produce 370 µg/L of valencene.
Subsequent investigation of the effects of cellular compartmentalization have led to identification of the mitochondrion as a potential sub-cellular location for terpene production. To achieve this, the authors equipped CsTPS1 with mitochondrial targeting peptide from the COX4 gene, studied earlier by Hurt et al. [67], to test whether mitochondrial pool of FPP can be utilized for valencene production. Expression of mitochondrial CsTPS1 (mtCsTPS1) by itself resulted in nearly 3-fold increase in valencene production, followed by further 50% increase when co-expressed with tHMG, and additional 40% increase while mitochondria targeted FDPS (mtFDPS) was used, yielding ca. 1.2 mg/L of valencene. Ultimately, the expression of an additional cytosolic copy of CsTPS1 resulted in production levels of approximately 1.5 mg/L of this sesquiterpene, suggesting that both mitochondrial and cytosolic pools of FPP might be used for the production of terpenes.
In a recent study, the utility of exploring natural protein variation was highlighted by the discovery of an alternative valencene synthase derived from the heartwood of the Nootka cypress (CnVS) [64]. In comparison to valencene synthase obtained from C. paradisi, expression of CnVS in a non-optimized S. cerevisiae strain produced 1.36 mg/L of valencene, an almost 450-fold increase in production compared to the citrus equivalent and a level which approximates those achieved via extensive metabolic optimization of strains containing the citrus enzymes. This new valencene synthase may therefore represent a valuable resource for further optimization and industrial scale production of this sesquiterpene.
(+)-nootkatone is an oxidized form of the sesquiterpene (+)-valencene, with a characteristic grapefruit flavour and relatively low sensory threshold [68]. It is found naturally in (and can be extracted from) plants including Citrus (Rutaceae spp.), Java (Cyperus rotundus), and Vetiver grass (Vetiveria spp.) [69,70]. Due to the very low concentrations of nootkatone found in natural sources, a synthetic compound is currently used for industrial applications [71]. However, as seen for many other flavour and aroma compounds, this synthetic production usually involves environmentally unfriendly processes and chemicals so heterologous production sources are being investigated [72,73].
Conversion of (+)-valencene to its oxidation products; (+)-nootkatone and α- and β-nootkatole (all having pleasant grapefruit-like aromas) are thought to be catalysed by members of the cytochrome P450 monoxygenase superfamily. Co-expression of the P450 enzyme CYP71AV8 from chicory root (Cichorium intybus), also known as (+)-valencene oxidase (VOX), which was reported to convert valencene to nootkatone via its oxidation at the C2 positions, along with valencene synthase from C. sinensis, allowed a recombinant S. cerevisiae strain to convert 68% of total pool of (+)-valencene to trans-nootkatol (0.92 mg/L), 8% to cis-nootkatol (0.11 mg/L), and 3% to (+)-nootkatone (0.04 mg/L) [74] (Figure 1).
In a later study, Gaviera et al. [75] used an in silico gene mining approach to identify and characterize novel cytochrome P450 valencene 2-oxygenases; CYP71D4 from Solanum tuberosum, CYP71D51v2 from Nicotiana tabacum, CYP71D1 from Catharanthus roseus, and CYP71D326 from Ricinus communis. Subsequently, they tested each of these oxygenases by introducing them into WAT11 yeast strain, which was genetically modified for stable expression of A. thaliana P450 reductase ATR1 [76]. The tobacco (Nicotiana tabacum) enzyme, CYP71D51v2, displayed the highest expression levels in this recombinant yeast, as well as efficient conversion of (+)-valencene to β-nootkatol and low levels of undesired byproducts. Incubation of this yeast strain with increasing (+)-valencene concentrations resulted in maximum combined yield of 6 mg/L of β-nootkatol and (+)-nootkatone, while 200 mg/L (+)-valencene was used. Further increase of (+)-valencene feed concentrations resulted in lower conversion rates, likely due to increased substrate and/or product toxicity.
Production of trans-nootkatol was further improved by overexpression of the native yeast type III membrane associate protein Ice2p [77]. Introduction of this genetic modification into recombinant strain engineered earlier to produce (+)-valencene via expression of premnaspirodiene oxygenase (HPO; CYP) from Hyoscyamus muticus and cytochrome P450 from A. thaliana (AtCPR) [64], resulted in stabilizing the levels and enzymatic activity of the latter heterologous enzyme. This modified recombinant strain was able to synthesize approximately 30 mg/L of trans-nootkatol. Although promising, the heterologous nootkatone production system would need to be further improved for its potential application on the industrial scale.
Nerolidol occurs in a broad spectrum of plants and, in its purified form, displays wood- and fresh bark-like aromas. It is officially permitted to be used as food flavouring additive by the U.S. Food and Drug Administration (FDA) and is currently used in industrial processing [25].
Recently, a study has explored the heterologous production of this sesquiterpene by S. cerevisiae via the expression of a nerolidol synthase (NES) from Actinida chinensis in an ERG20 overexpression background, resulting in the production of nerolidol in the growth media at 5 mg/L [78]. Further pathway optimization of this strain, which included replacing the initial three enzymes of “upper” MEV pathway (ERG10, ERG13, and tHMG1) with codon optimized heterologous counterparts and co-expressing HMG-CoA synthase and a bi-functional acetoacetyl-CoA thiolase/HMG-CoA reductase from Enterococcus faecalis resulting in over a ten-fold increase in nerolidol (56 mg/L). The authors also addressed the issue of undesired production of high levels of squalene, resulting from enzymatic activity of squalene epoxidase (ERG9), which competes with the heterologous nerolidol synthase (AcNES1) for the cellular pools of FPP. As production of squalene is essential for the survival of yeast, they implemented elegant system in which the degradation of the native Erg9 protein was enhanced, thereby decreasing its cellular half-life and enzymatic activity and increasing the production of trans-nerolidol by 86% (105 mg/L) [79,80].
3.1.3. Norisoprenoids
Norisoprenoids are a group of C-13-carotenoid-derived aromatic compounds, produced by many fruit and flowers, such as grapes, peaches, blackberries, and which are greatly valued as flavouring additives [81]. In plants, norisoprenoids are produced by the enzymatic activity of carotenoid cleavage oxygenases (CCO), which cleave carotenoids at positions C9 through C13 and thereby yielding different classes of these compounds [82].
β-ionone is one of the most common of this group of aromatic compounds, displaying floral, violet-like characteristics and exhibiting an extremally low odour threshold of 0.007 ppb (in water) [81]. Due to its desirable properties β-ionone has been used as a food and beverage additive to enhance the flavour of ice cream, candy, baked goods, gelatines, puddings, chewing gum, and non-alcoholic beverages [83].
While β-ionone can be extracted from plants, where it is naturally produced by the enzymatic cleavage of β-carotene by carotenoid cleavage dioxygenase (CCD1), the overall process is tedious and expensive, and highly depends on availability of agricultural resources. Thus, microbial heterologous production of this valuable compound is an interesting alternative. De novo biosynthesis of β-carotene in S. cerevisiae, which is a precursor for β-ionone production, were successfully achieved by expression of three heterologous genes from the ascomycete Xanthophyllomyces dendrorhous; phytoene desaturase (crtI), GGPP synthase (crtE), and phytoene synthase/lycopene cyclase (crtYB) [84] (Figure 1). Later, Beekwilder and colleagues [85] used the same enzymes, along with expression of carotenoid-cleavage dioxygenase from raspberry (RiCCD1), demonstrating the possibility of heterologous production of β-ionone in S. cerevisiae. Relatively low amounts (0.22 mg/L) of this compound were produced by the recombinant yeast, which was presumably associated with the low efficiency of translation of the single polycistronic episomal construct carrying crtYB/crtI/crtE genes. Recently, Lopes et al. [86] reported engineering a more efficient heterologous strain of S. cerevisiae that was able to produce more than 5 mg/L of β-ionone. It was achieved by genetically modifying previously reported, FPP overproducing strain (SCGIS22) [87], via overexpression of tHMG1 and GGPP synthase gene BTS1, along with expression of heterologous crtI and crtYB from X. dendrorhous, and CCD1 from Petunia hybrida (PhCCD1) (Figure 1)
4. Next Generation Sweeteners
Excessive consumption levels of high-caloric sweeteners, such as sugars, has been proven to have multiple negative impacts on human health, resulting in gain of weight, high blood pressure, or type 2 diabetes [88]. Thus, there is growing demand in food and beverage industries for a ‘new generation’ sweetening agents, which would provide to the consumers similar to common sugars perception of sweetness, while significantly restricting the high calorie intake. Natural plant derived compounds, such as stevia glucosides, has proven to be an attractive solution.
Steviol Glycosides
Steviol glycosides (SGs) are a group of active compounds found in the leaves of Stevia rebaudiana Bertoni (also known as “sweet herb”), which has been used for centuries as a natural sweetener by native people of South America [89]. The SGs present in stevia plants; stevioside (Ste), rebaudioside- (Reb-) A, B, C, D, E, F, and M, dulcoside A, steviolbioside, and rubusoside, have distinctive sweet tastes that are estimated to range from 30 to 250 times more potent than sucrose [90]. Given these properties, it is not surprising that several SGs are now being used in the food and beverages industries as alternative, low-caloric sweetening agents [91].
The natural biosynthesis of SGs involves the isoprenoid pathway, with GGPP representing the metabolic branch point from this core metabolic pathway [92]. Four novel enzymatic steps then lead to the production of kaurenoic acid, which is further converted to steviol by kaurenoic acid hydroxylase (KAH). From this point, the activity of a variety of specific UDP-glycosyltransferases (UGTs) on the basic steviol molecule gives rise to the various SGs that are observed in plant extracts. The above metabolic steps were used to introduce de novo SGs production pathway into S. cerevisiae with five heterologous enzymes driving production of steviol from GGPP (Figure 1).
Fortunately, the heterologous production of SGs from steviol is aided in yeast by the presence of naturally high levels of cytosolic UDP-glucose [93]. This intracellular UDP-glucose pool can therefore be exploited as a sugar-donor by various heterologously-expressed UDP-glucosyltransferases for the synthesis of specific SGs. Efficient bioconversion (60%) of supplemented stevioside (relative sweetness of ~210) to rebaudioside A (relative sweetness of ~200), has been recently achieved by expressing the UDP-glycosyltransferase, UGT76G1, from S. rebaudiana in S. cerevisiae [94].
However, complicating the production of specific SGs is the fact that many glucosyltransferases display loosely-constrained substrate requirements and have the ability to catalyze multiple different glycosylation reactions. This leads to the co-synthesis of mixtures of different SGs, rather than the production of a single pure compound. For example, UGT76G1, which catalyzes the formation of highly desirable steviol glucosides, such as rebaudioside M and D, is also shown to produce undesirable byproducts, including rebaudioside G, Q, I and 1,3-bioside [90]. To address this enzymatic promiscuity, Olsson and colleagues [95] used protein site saturation mutagenesis to produce variants in both the predicted substrate-binding pocket, and enzyme active site UGT76G1. A library of 1748 mutants were screened using a strain of S. cerevisiae that carried the set of heterologous genes required for generation of rebaudioside, but lucking a glycosyltransferase [96]. Several mutants, such as UGT76G1Thr146Gly and UGT76G1His155Leu, displayed the desired increase in rebaudioside M and/or D production with minimal byproduct formation. The study brings important insights into the possibility of altering the enzymatic specificity of steviol glucosyltransferases towards more efficient production of desired SGs.
5. Conclusions
The estimated turnover of the flavour and fragrance industries has been estimated to reach over US $30 billion in 2017, with a compounding annual growth rate of 5.6% between 2012 and 2017 [97]. To fulfil the demands of this constantly growing market, chemical synthesis of flavour and aroma compounds has routinely been used. However, the processes involved are generally not sustainable and the final products are considered of synthetic origin, which dramatically decreases their market value. While extraction of flavours and fragrances from natural sources provides a more valuable, natural product, there are number of limiting factors that make these processes inefficient and relatively expensive, including the availability of the source plant materials, and the low levels of the desired compounds in planta. Thus, there is a constantly growing demand for new, improved biotechnologies which allow more efficient, cheaper, and environmentally friendly ways to produce aroma and flavour compounds.
Over the last two decades, the rapidly growing field of synthetic biology has provided the possibility for the heterologous production of specific flavours and aromas in microorganisms, a unique resource that could be used to help fulfil these global demands. The biotechnological company Evolva [98] has taken advantage of synthetic biology and yeast for the industrial scale heterologous production of natural flavour and aroma compounds, such as nootkatone, valencene, vanillin, and next-generation sweeteners. However, from the thousands of flavour and aroma compounds that have been identified to date, only a limited number have been successfully produced by heterologous means. Along with new advances in metabolic engineering of S. cerevisiae [99], this presents an opportunity for development and potential industrial applications of new heterologous pathways to produce novel valuable compounds in yeast.
Funding
The AWRI, a member of the Wine Innovation Cluster in Adelaide, is supported by Australia’s grape growers and winemakers through their investment body Wine Australia with matching funds from the Australian Government.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Galanie, S.; Thodey, K.; Trenchard, I.J.; Filsinger Interrante, M.; Smolke, C.D. Complete biosynthesis of opioids in yeast. Science 2015, 349, 1095–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, T.L.K.H.; Kong, I.I.; Liu, J.J.; Zhang, G.C.; Jin, Y.S. Engineering and Evolution of Saccharomyces Cerevisiae to Produce Biofuels and Chemicals; Springer International Publishing: Cham, Switzerland, 2018; pp. 175–215. [Google Scholar]
- Teo, W.S.; Ling, H.; Yu, A.-Q.; Chang, M.W. Metabolic engineering of Saccharomyces cerevisiae for production of fatty acid short- and branched-chain alkyl esters biodiesel. Biotechnol. Biofuels 2015, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Ledesma-Amaro, R.; Nicaud, J.-M. Metabolic engineering for expanding the substrate range of Yarrowia lipolytica. Trends Biotechnol. 2016, 34, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.R.; Shin, J.H.; Cho, J.S.; Yang, D.; Lee, S.Y. Systems metabolic engineering of Escherichia coli. EcoSal Plus 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Priefert, H.; Rabenhorst, J.; Steinbuchel, A. Biotechnological production of vanillin. Appl. Microbiol. Biotechnol. 2001, 56, 296–314. [Google Scholar] [CrossRef] [PubMed]
- Brochado, A.R.; Matos, C.; Moller, B.L.; Hansen, J.; Mortensen, U.H.; Patil, K.R. Improved vanillin production in baker’s yeast through in silico design. Microb. Cell Factories 2010, 9, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walton, N.J.; Narbad, A.; Faulds, C.; Williamson, G. Novel approaches to the biosynthesis of vanillin. Curr. Opin. Biotechnol. 2000, 11, 490–496. [Google Scholar] [CrossRef]
- Hansen, E.H.; Moller, B.L.; Kock, G.R.; Bunner, C.M.; Kristensen, C.; Jensen, O.R.; Okkels, F.T.; Olsen, C.E.; Motawia, M.S.; Hansen, J. De novo biosynthesis of vanillin in fission yeast (Schizosaccharomyces pombe) and baker’s yeast (Saccharomyces cerevisiae). Appl. Environ. Microbiol. 2009, 75, 2765–2774. [Google Scholar] [CrossRef] [PubMed]
- Brochado, A.R.; Patil, K.R. Overexpression of O-methyltransferase leads to improved vanillin production in baker’s yeast only when complemented with model-guided network engineering. Biotechnol. Bioeng. 2013, 110, 656–659. [Google Scholar] [CrossRef] [PubMed]
- Strucko, T.; Magdenoska, O.; Mortensen, U.H. Benchmarking two commonly used Saccharomyces cerevisiae strains for heterologous vanillin-β-glucoside production. Metab. Eng. Commun. 2015, 2, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.; Poll, L. Odour thresholds of some important aroma compounds in raspberries. Z. Lebensm.-Unters. Forsch. 1990, 191, 129–131. [Google Scholar] [CrossRef]
- Larsen, M.; Poll, L.; Callesen, O.; Lewis, M. Relations between the content of aroma compounds and the sensory evaluation of 10 raspberry varieties (Rubus idaeus L.). Acta Agric. Scand. 1991, 41, 447–454. [Google Scholar] [CrossRef]
- Lee, D.; Lloyd, N.D.; Pretorius, I.S.; Borneman, A.R. Heterologous production of raspberry ketone in the wine yeast Saccharomyces cerevisiae via pathway engineering and synthetic enzyme fusion. Microb. Cell Factories 2016, 15, 49. [Google Scholar] [CrossRef] [PubMed]
- Stabnikova, O.; Wang, J.-Y.; Ivanov, V. Value-added biotechnological products from organic wastes. In Environment Biotechnology; Humana Press: Totowa, NJ, USA, 2010; pp. 343–394. [Google Scholar]
- Abe, I.; Takahashi, Y.; Morita, H.; Noguchi, H. Benzalacetone synthase. A novel polyketide synthase that plays a crucial role in the biosynthesis of phenylbutanones in Rheum palmatum. Eur. J. Biochem. 2001, 268, 3354–3359. [Google Scholar] [CrossRef] [PubMed]
- Borejsza-Wysocki, W.; Hrazdina, G. Biosynthesis of p-hydroxyphenylbutan-2-one in raspberry fruits and tissue cultures. Phytochemistry 1994, 35, 623–628. [Google Scholar] [CrossRef]
- Koeduka, T.; Watanabe, B.; Suzuki, S.; Hiratake, J.; Mano, J.; Yazaki, K. Characterization of raspberry ketone/zingerone synthase, catalyzing the alpha, beta-hydrogenation of phenylbutenones in raspberry fruits. Biochem. Biophys. Res. Commun. 2011, 412, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.; van der Meer, I.M.; Sibbesen, O.; Broekgaarden, M.; Qvist, I.; Mikkelsen, J.D.; Hall, R.D. Microbial production of natural raspberry ketone. Biotechnol. J. 2007, 2, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Vannelli, T.; Qi, W.W.; Sweigard, J.; Gatenby, A.A.; Sariaslani, F.S. Production of p-hydroxycinnamic acid from glucose in Saccharomyces cerevisiae and Escherichia coli by expression of heterologous genes from plants and fungi. Metab. Eng. 2007, 9, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.Y.; Jung, S.M.; Kim, M.D.; Han, N.S.; Seo, J.H. Production of resveratrol from tyrosine in metabolically engineered Saccharomyces cerevisiae. Enzyme. Microb. Technol. 2012, 51, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Raina, V.K.; Srivastava, S.K.; Aggarwal, K.K.; Ramesh, S.; Kumar, S. Essential oil composition of Cinnamomum zeylanicum Blume leaves from Little Andaman, India. Flavour Fragr. J. 2001, 16, 374–376. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Rao, L.J.; Sakariah, K.K. Chemical composition of the volatile oil from the fruits of Cinnamomum zeylanicum blume. Flavour Fragr. J. 1997, 12, 331–333. [Google Scholar] [CrossRef]
- Friedman, M.; Kozukue, N.; Harden, L.A. Cinnamaldehyde content in foods determined by gas chromatography-mass spectrometry. J. Agric. Food. Chem. 2000, 48, 5702–5709. [Google Scholar] [CrossRef] [PubMed]
- Official Journal of the European Union: Commission Implementing Regulation (EU) No 872/2012, 1st October 2012. Available online: https://publications.europa.eu/en/publication-detail/-/publication/9d00bfce-0bc9-11e2-8e28-01aa75ed71a1/language-en (accessed on 11 May 2018).
- Tajkarimi, M.M.; Ibrahim, S.A.; Cliver, D.O. Antimicrobial herb and spice compounds in food. Food Control 2010, 21, 1199–1218. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Gottardi, M.; Knudsen, J.D.; Prado, L.; Oreb, M.; Branduardi, P.; Boles, E. De novo biosynthesis of trans-cinnamic acid derivatives in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 4883–4893. [Google Scholar] [CrossRef] [PubMed]
- Larroy, C.; Fernandez, M.R.; Gonzalez, E.; Pares, X.; Biosca, J.A. Characterization of the Saccharomyces cerevisiae YMR318C (ADH6) gene product as a broad specificity NADPH-dependent alcohol dehydrogenase: Relevance in aldehyde reduction. Biochem. J. 2002, 361, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.A.; Croteau, R.B. Resin-based defenses in conifers. Trends Plant Sci. 1999, 4, 184–190. [Google Scholar] [CrossRef]
- Loza-Tavera, H. Monoterpenes in essential oils. Biosynthesis and properties. Adv. Exp. Med. Biol. 1999, 464, 49–62. [Google Scholar] [PubMed]
- Winter, R. A Consumer’s Dictionary of Food Additives, 7th ed.; Random House USA Inc.: New York, NY, USA, 2009; p. 592. [Google Scholar]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Oswald, M.; Fischer, M.; Dirninger, N.; Karst, F. Monoterpenoid biosynthesis in Saccharomyces cerevisiae. FEMS Yeast Res. 2007, 7, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Withers, S.T.; Keasling, J.D. Biosynthesis and engineering of isoprenoid small molecules. Appl. Microbiol. Biotechnol. 2007, 73, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Chambon, C.; Ladeveze, V.; Oulmouden, A.; Servouse, M.; Karst, F. Isolation and properties of yeast mutants affected in farnesyl diphosphate synthetase. Curr. Genet. 1990, 18, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Iijima, Y.; Gang, D.R.; Fridman, E.; Lewinsohn, E.; Pichersky, E. Characterization of geraniol synthase from the peltate glands of sweet basil. Plant Physiol. 2004, 134, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.J.; Meyer, S.; Claudel, P.; Bergdoll, M.; Karst, F. Metabolic engineering of monoterpene synthesis in yeast. Biotechnol. Bioeng. 2011, 108, 1883–1892. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Li, J.; Wang, H.X.; Zeng, Y. A geraniol-synthase gene from cinnamomum tenuipilum. Phytochemistry 2005, 66, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Miettinen, K.; Goedbloed, M.; Verstappen, F.W.; Voster, A.; Jongsma, M.A.; Memelink, J.; van der Krol, S.; Bouwmeester, H.J. Characterization of two geraniol synthases from Valeriana officinalis and Lippia dulcis: Similar activity but difference in subcellular localization. Metab. Eng. 2013, 20, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Polakowski, T.; Stahl, U.; Lang, C. Overexpression of a cytosolic hydroxymethylglutaryl-coa reductase leads to squalene accumulation in yeast. Appl. Microbiol. Biotechnol. 1998, 49, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, W.; Du, G.; Chen, J.; Zhou, J. Overproduction of geraniol by enhanced precursor supply in Saccharomyces cerevisiae. J. Biotechnol. 2013, 168, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Bao, X.; Li, C.; Shen, Y.; Hou, J. Improving monoterpene geraniol production through geranyl diphosphate synthesis regulation in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2016, 100, 4561–4571. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.Z.; Yao, M.D.; Wang, Y.; Zhou, L.; Song, T.Q.; Liu, H.; Xiao, W.H.; Yuan, Y.J. Manipulation of GES and ERG20 for geraniol overproduction in Saccharomyces cerevisiae. Metab. Eng. 2017, 41, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Pardo, E.; Rico, J.; Gil, J.V.; Orejas, M. De novo production of six key grape aroma monoterpenes by a geraniol synthase-engineered S. cerevisiae wine strain. Microb. Cell Factories 2015, 14, 136. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Cseke, L.; Blanc, V.M.; Pichersky, E. Evolution of floral scent in clarkia: Novel patterns of s-linalool synthase gene expression in the C. breweri flower. Plant Cell 1996, 8, 1137–1148. [Google Scholar] [CrossRef] [PubMed]
- Herrero, O.; Ramon, D.; Orejas, M. Engineering the Saccharomyces cerevisiae isoprenoid pathway for de novo production of aromatic monoterpenes in wine. Metab. Eng. 2008, 10, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Rico, J.; Pardo, E.; Orejas, M. Enhanced production of a plant monoterpene by overexpression of the 3-hydroxy-3-methylglutaryl coenzyme a reductase catalytic domain in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2010, 76, 6449–6454. [Google Scholar] [CrossRef] [PubMed]
- Amiri, P.; Shahpiri, A.; Asadollahi, M.A.; Momenbeik, F.; Partow, S. Metabolic engineering of Saccharomyces cerevisiae for linalool production. Biotechnol. Lett. 2016, 38, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Denby, C.M.; Li, R.A.; Vu, V.T.; Costello, Z.; Lin, W.; Chan, L.J.G.; Williams, J.; Donaldson, B.; Bamforth, C.W.; Petzold, C.J.; et al. Industrial brewing yeast engineered for the production of primary flavor determinants in hopped beer. Nat. Commun. 2018, 9, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duetz, W.A.; Bouwmeester, H.; van Beilen, J.B.; Witholt, B. Biotransformation of limonene by bacteria, fungi, yeasts, and plants. Appl. Microbiol. Biotechnol. 2003, 61, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, B.M. Biosynthesis and biotechnology of high-value p-menthane monoterpenes, including menthol, carvone, and limonene. Adv. Biochem. Eng. Biotechnol. 2015, 148, 319–353. [Google Scholar] [CrossRef] [PubMed]
- Tranchida, P.Q.; Zoccali, M.; Bonaccorsi, I.; Dugo, P.; Mondello, L.; Dugo, G. The off-line combination of high performance liquid chromatography and comprehensive two-dimensional gas chromatography-mass spectrometry: A powerful approach for highly detailed essential oil analysis. J. Chromatogr. A 2013, 1305, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Soković, M.; Glamočlija, J.; Marin, P.D.; Brkić, D.; van Griensven, L.J. Antibacterial effects of the essential oils of commonly consumed medicinal herbs using an in vitro model. Molecules 2010, 15, 7532–7546. [Google Scholar] [CrossRef] [PubMed]
- Behrendorff, J.B.; Vickers, C.E.; Chrysanthopoulos, P.; Nielsen, L.K. 2,2-diphenyl-1-picrylhydrazyl as a screening tool for recombinant monoterpene biosynthesis. Microb. Cell Factories 2013, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Jongedijk, E.; Cankar, K.; Ranzijn, J.; van der Krol, S.; Bouwmeester, H.; Beekwilder, J. Capturing of the monoterpene olefin limonene produced in Saccharomyces cerevisiae. Yeast 2015, 32, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Jongedijk, E.; Cankar, K.; Buchhaupt, M.; Schrader, J.; Bouwmeester, H.; Beekwilder, J. Biotechnological production of limonene in microorganisms. Appl. Microbiol. Biotechnol. 2016, 100, 2927–2938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietschmann, M.; Vostrowsky, O.; Bestmann, H.J.; Pant, A.K.; Mathela, C.S. Volatile constituents of Hyptis pectinata Poit. (lamiaceae). J. Essent. Oil Res. 1998, 10, 550–552. [Google Scholar] [CrossRef]
- Arunkumar, R.; Nair, S.A.; Rameshkumar, K.B.; Subramoniam, A. The essential oil constituents of Zornia diphylla (L.) Pers, and anti-inflammatory and antimicrobial activities of the oil. Rec. Nat. Prod. 2014, 8, 385–393. [Google Scholar]
- Kampranis, S.C.; Ioannidis, D.; Purvis, A.; Mahrez, W.; Ninga, E.; Katerelos, N.A.; Anssour, S.; Dunwell, J.M.; Degenhardt, J.; Makris, A.M.; et al. Rational conversion of substrate and product specificity in a salvia monoterpene synthase: Structural insights into the evolution of terpene synthase function. Plant Cell 2007, 19, 1994–2005. [Google Scholar] [CrossRef] [PubMed]
- Sharon-Asa, L.; Shalit, M.; Frydman, A.; Bar, E.; Holland, D.; Or, E.; Lavi, U.; Lewinsohn, E.; Eyal, Y. Citrus fruit flavor and aroma biosynthesis: Isolation, functional characterization, and developmental regulation of Cstps1, a key gene in the production of the sesquiterpene aroma compound valencene. Plant J. 2003, 36, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, M.; Hashimoto, T.; Noma, Y.; Asakawa, Y. Highly efficient production of nootkatone, the grapefruit aroma from valencene, by biotransformation. Chem. Pharm. Bull. 2005, 53, 1513–1514. [Google Scholar] [CrossRef] [PubMed]
- Farhi, M.; Marhevka, E.; Masci, T.; Marcos, E.; Eyal, Y.; Ovadis, M.; Abeliovich, H.; Vainstein, A. Harnessing yeast subcellular compartments for the production of plant terpenoids. Metab. Eng. 2011, 13, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.; van Houwelingen, A.; Cankar, K.; van Dijk, A.D.; de Jong, R.M.; Stoopen, G.; Bouwmeester, H.; Achkar, J.; Sonke, T.; Bosch, D. Valencene synthase from the heartwood of nootka cypress (Callitropsis nootkatensis) for biotechnological production of valencene. Plant Biotechnol. J. 2014, 12, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Ro, D.K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 2006, 440, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.D.; MacNevin, G.; Ro, D.K. De novo synthesis of high-value plant sesquiterpenoids in yeast. Methods Enzymol. 2012, 517, 261–278. [Google Scholar] [CrossRef] [PubMed]
- Hurt, E.C.; Pesold-Hurt, B.; Suda, K.; Oppliger, W.; Schatz, G. The first twelve amino acids (less than half of the pre-sequence) of an imported mitochondrial protein can direct mouse cytosolic dihydrofolate reductase into the yeast mitochondrial matrix. EMBO J. 1985, 4, 2061–2068. [Google Scholar] [PubMed]
- Haring, H.G.; Rijkens, F.; Boelens, H.; Van der Gen, A. Olfactory studies on enantiomeric eremophilane sesquiterpenoids. J. Agric. Food Chem. 1972, 20, 1018–1021. [Google Scholar] [CrossRef]
- Leonhardt, R.-H.; Berger, R.G. Nootkatone. Adv. Biochem. Eng. Biotechnol. 2015, 148, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, J.A.; Ortuno, A.; Garcia-Puig, D.; Porras, I.; Garcia-Lidon, A.; Sabater, F. Variations of nootkatone and valencene levels during the development of grapefruit. J. Agric. Food Chem. 1992, 40, 1488–1490. [Google Scholar] [CrossRef]
- Fraatz, M.A.; Berger, R.G.; Zorn, H. Nootkatone—A biotechnological challenge. Appl. Microbiol. Biotechnol. 2009, 83, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Hunter, G.L.K.; Brogden, W.B., Jr. Conversion of valencene to nootkatone. J. Food Sci. 1965, 30, 876–878. [Google Scholar] [CrossRef]
- Wilson, C.W.; Shaw, P.E. Synthesis of nootkatone from valencene. J. Agric. Food Chem. 1978, 26, 1430–1432. [Google Scholar] [CrossRef]
- Cankar, K.; van Houwelingen, A.; Bosch, D.; Sonke, T.; Bouwmeester, H.; Beekwilder, J. A chicory cytochrome P450 mono-oxygenase CYP71AV8 for the oxidation of (+)-valencene. FEBS Lett. 2011, 585, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Gavira, C.; Hofer, R.; Lesot, A.; Lambert, F.; Zucca, J.; Werck-Reichhart, D. Challenges and pitfalls of P450-dependent (+)-valencene bioconversion by Saccharomyces cerevisiae. Metab. Eng. 2013, 18, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Urban, P.; Mignotte, C.; Kazmaier, M.; Delorme, F.; Pompon, D. Cloning, yeast expression, and characterization of the coupling of two distantly related Arabidopsis thaliana NADPH-cytochrome P450 reductases with P450 CYP73A5. J. Biol. Chem. 1997, 272, 19176–19186. [Google Scholar] [CrossRef] [PubMed]
- Emmerstorfer, A.; Wimmer-Teubenbacher, M.; Wriessnegger, T.; Leitner, E.; Muller, M.; Kaluzna, I.; Schurmann, M.; Mink, D.; Zellnig, G.; Schwab, H.; et al. Over-expression of ice2 stabilizes cytochrome P450 reductase in Saccharomyces cerevisiae and Pichia pastoris. Biotechnol. J. 2015, 10, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Plan, M.R.; Chrysanthopoulos, P.; Hodson, M.P.; Nielsen, L.K.; Vickers, C.E. A squalene synthase protein degradation method for improved sesquiterpene production in Saccharomyces cerevisiae. Metab. Eng. 2017, 39, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Mateus, C.; Avery, S.V. Destabilized green fluorescent protein for monitoring dynamic changes in yeast gene expression with flow cytometry. Yeast 2000, 16, 1313–1323. [Google Scholar] [CrossRef] [Green Version]
- Salama, S.R.; Hendricks, K.B.; Thorner, J. G1 cyclin degradation: The PEST motif of yeast Cln2 is necessary, but not sufficient, for rapid protein turnover. Mol. Cell. Biol. 1994, 14, 7953–7966. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Bustamante, E.; Sanchez, S. Microbial production of C13-norisoprenoids and other aroma compounds via carotenoid cleavage. Crit. Rev. Microbiol. 2007, 33, 211–230. [Google Scholar] [CrossRef] [PubMed]
- Enzell, C.R. Biodegradation of carotenoids—An important route to aroma compounds. Pure Appl. Chem. 1985, 57, 693–700. [Google Scholar] [CrossRef]
- Burdock, G.A. Fenaroli’s Handbook of Flavor Ingredients, 6th ed.; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Verwaal, R.; Wang, J.; Meijnen, J.P.; Visser, H.; Sandmann, G.; van den Berg, J.A.; van Ooyen, A.J. High-level production of beta-carotene in Saccharomyces cerevisiae by successive transformation with carotenogenic genes from Xanthophyllomyces dendrorhous. Appl. Environ. Microbiol. 2007, 73, 4342–4350. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.; van Rossum, H.M.; Koopman, F.; Sonntag, F.; Buchhaupt, M.; Schrader, J.; Hall, R.D.; Bosch, D.; Pronk, J.T.; van Maris, A.J.A.; et al. Polycistronic expression of a β-carotene biosynthetic pathway in Saccharomyces cerevisiae coupled to β-ionone production. J. Biotechnol. 2014, 192, 383–392. [Google Scholar] [CrossRef] [PubMed]
- López, J.; Essus, K.; Kim, I.-K.; Pereira, R.; Herzog, J.; Siewers, V.; Nielsen, J.; Agosin, E. Production of β-ionone by combined expression of carotenogenic and plant Ccd1 genes in Saccharomyces cerevisiae. Microb. Cell Factories 2015, 14, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalcinati, G.; Partow, S.; Siewers, V.; Schalk, M.; Daviet, L.; Nielsen, J. Combined metabolic engineering of precursor and co-factor supply to increase α-santalene production by Saccharomyces cerevisiae. Microb. Cell Factories 2012, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, I.A. A review of recent evidence relating to sugars, insulin resistance and diabetes. Eur. J. Nutr. 2016, 55, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, W.H. Early uses of Stevia rebaudiana (Asteraceae) leaves as a sweetener in paraguay. AGRIS 1992, 46, 336–337. [Google Scholar]
- Prakash, I.; Markosyan, A.; Bunders, C. Development of next generation stevia sweetener: Rebaudioside M. Foods 2014, 3, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Prakash, I.; Dubois, G.E.; Clos, J.F.; Wilkens, K.L.; Fosdick, L.E. Development of rebiana, a natural, non-caloric sweetener. Food. Chem. Toxicol. 2008, 46 (Suppl. 7), S75–S82. [Google Scholar] [CrossRef] [PubMed]
- Wanke, M.; Skorupinska-Tudek, K.; Swiezewska, E. Isoprenoid biosynthesis via 1-deoxy-d-xylulose 5-phosphate/2-C-methyl-d-erythritol 4-phosphate (DOXP/MEP) pathway. Acta Biochim. Pol. 2001, 48, 663–672. [Google Scholar] [PubMed]
- Oka, T.; Jigami, Y. Reconstruction of de novo pathway for synthesis of UDP-glucuronic acid and UDP-xylose from intrinsic UDP-glucose in Saccharomyces cerevisiae. FEBS J. 2006, 273, 2645–2657. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Wang, Y.; Chen, L.; Yan, M.; Chen, K.; Xu, L.; Ouyang, P. Production of rebaudioside a from stevioside catalyzed by the engineered Saccharomyces cerevisiae. Appl. Biochem. Biotechnol. 2016, 178, 1586–1598. [Google Scholar] [CrossRef] [PubMed]
- Olsson, K.; Carlsen, S.; Semmler, A.; Simon, E.; Mikkelsen, M.D.; Moller, B.L. Microbial production of next-generation stevia sweeteners. Microb. Cell Factories 2016, 15, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikkelsen, M.D.; Hansen, J.; Simon, E.; Brianza, F.; Semmler, A.; Olsson, K.; Carlsen, S.; Düring, L.; Ouspensku, A.; Hicks, P. Methods for improved production of rebaudioside D and rebaudioside M. U.S. Patent No. 9,957,540, 1 May 2018. [Google Scholar]
- Global Markets for Flavors and Fragrances. Available online: www.bccresearch.com/market-research/chemicals/flavors-fragrances-global-markets-chm034c.html (accessed on 23 May 2018).
- Evolva, A.O. Available online: https://www.evolva.com/ (accessed on 21 May 2018).
- Lian, J.; Mishra, S.; Zhao, H. Recent advances in metabolic engineering of Saccharomyces cerevisiae: New tools and their applications. Metab. Eng. 2018. [Google Scholar] [CrossRef] [PubMed]
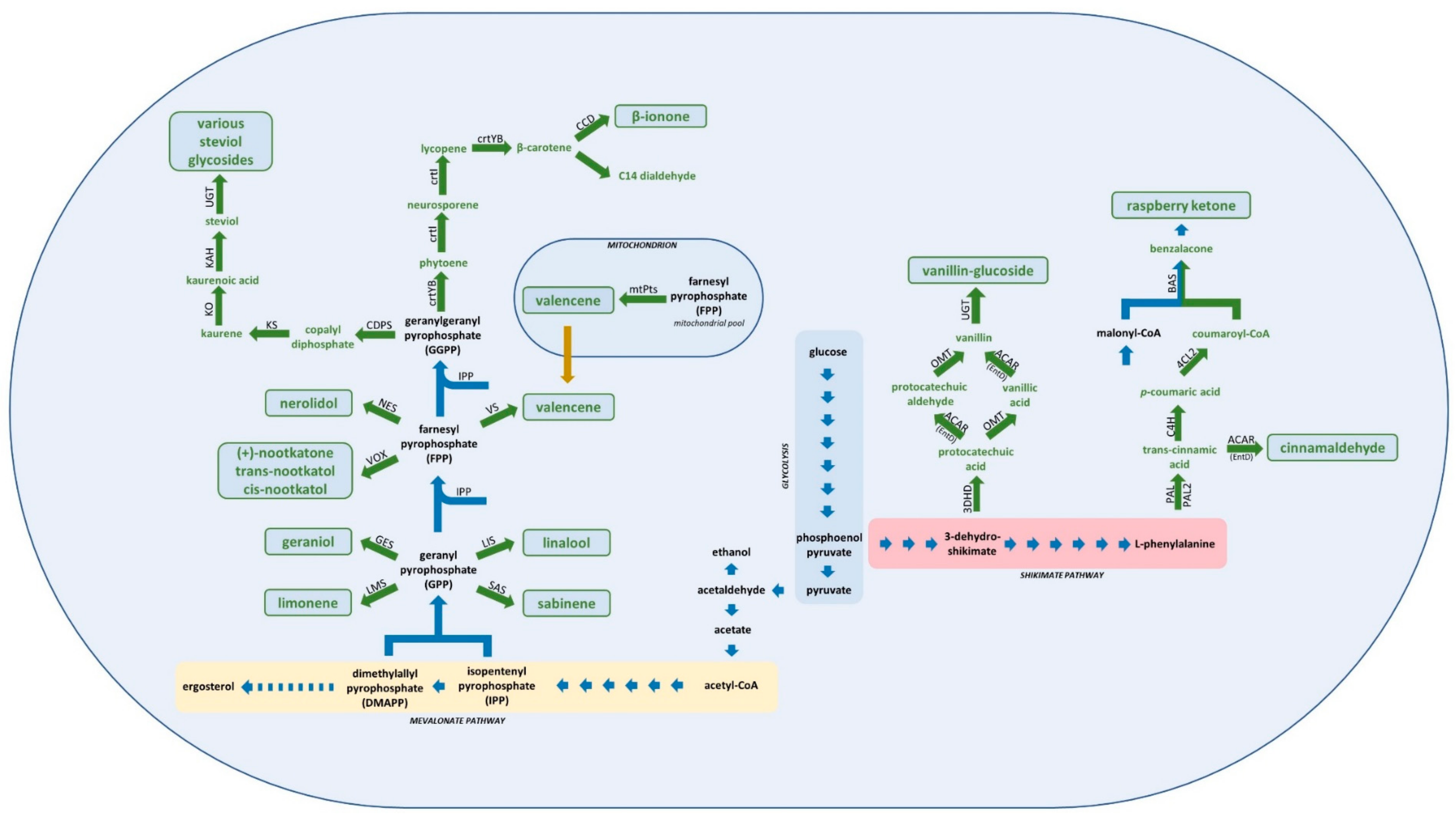
Figure 1.
Simplified central metabolism of Saccharomyces cerevisiae (blue arrows), and heterologous enzymatic reactions (green arrows) leading to production of various flavour and aroma compounds: vanillin-glucoside (3DSD: 3-dehydroshikimate dehydratase, ACAR: aryl carboxylic acid reductase, OMT: O-methyltransferase, UGT: UDP-glycosyltransferase), raspberry ketone (PAL: phenylalanine/tyrosine ammonia lyase, C4H: cinnamate-4-hydroxlase, 4CL2: coumarate-CoA ligase 2, BAS: benzalacetone synthase), cinnamaldehycde (PAL2: phenylalanine ammonia lyase 2, ACAR: aryl carboxylic acid reductase, EntD: phosphopantetheinyl transferase), geraniol (GES: geraniol synthase), limonene (LMS: limonene synthase), linalool (LIS: linalool synthase), sabinene (SAS: sabinene synthase), valencene (VS: valencene synthase, mtPts: mitochondria targeted valencene synthase), (+)-nootkatone, trans-nootkatol, cis-nootkatol (VOX: (+)-valencene oxidase), nerolidol (NES: nerolidol synthase), β-ionone (crtYB: phytoene synthase/lycopene cyclase, crtI: phytoene desaturase, CCD: carotenoid cleavage dioxygenase), steviol glycosides (CDPS: copalyl diphosphate synthase, KS: kaurene synthase, KO: kaurene oxidase, KAH: kaurenoic acid-13-hydroxylase, UGT: UDP-glycosyltransferase).
Figure 1.
Simplified central metabolism of Saccharomyces cerevisiae (blue arrows), and heterologous enzymatic reactions (green arrows) leading to production of various flavour and aroma compounds: vanillin-glucoside (3DSD: 3-dehydroshikimate dehydratase, ACAR: aryl carboxylic acid reductase, OMT: O-methyltransferase, UGT: UDP-glycosyltransferase), raspberry ketone (PAL: phenylalanine/tyrosine ammonia lyase, C4H: cinnamate-4-hydroxlase, 4CL2: coumarate-CoA ligase 2, BAS: benzalacetone synthase), cinnamaldehycde (PAL2: phenylalanine ammonia lyase 2, ACAR: aryl carboxylic acid reductase, EntD: phosphopantetheinyl transferase), geraniol (GES: geraniol synthase), limonene (LMS: limonene synthase), linalool (LIS: linalool synthase), sabinene (SAS: sabinene synthase), valencene (VS: valencene synthase, mtPts: mitochondria targeted valencene synthase), (+)-nootkatone, trans-nootkatol, cis-nootkatol (VOX: (+)-valencene oxidase), nerolidol (NES: nerolidol synthase), β-ionone (crtYB: phytoene synthase/lycopene cyclase, crtI: phytoene desaturase, CCD: carotenoid cleavage dioxygenase), steviol glycosides (CDPS: copalyl diphosphate synthase, KS: kaurene synthase, KO: kaurene oxidase, KAH: kaurenoic acid-13-hydroxylase, UGT: UDP-glycosyltransferase).

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kutyna, D.R.; Borneman, A.R. Heterologous Production of Flavour and Aroma Compounds in Saccharomyces cerevisiae. Genes 2018, 9, 326. https://doi.org/10.3390/genes9070326
AMA Style
Kutyna DR, Borneman AR. Heterologous Production of Flavour and Aroma Compounds in Saccharomyces cerevisiae. Genes. 2018; 9(7):326. https://doi.org/10.3390/genes9070326
Chicago/Turabian StyleKutyna, Dariusz R., and Anthony R. Borneman. 2018. "Heterologous Production of Flavour and Aroma Compounds in Saccharomyces cerevisiae" Genes 9, no. 7: 326. https://doi.org/10.3390/genes9070326
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.