Climate Change Impacts on Natural Sulfur Production: Ocean Acidification and Community Shifts

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Experiments
2.1. Natural Sulfur Emissions: Ocean Acidification (ESNa)
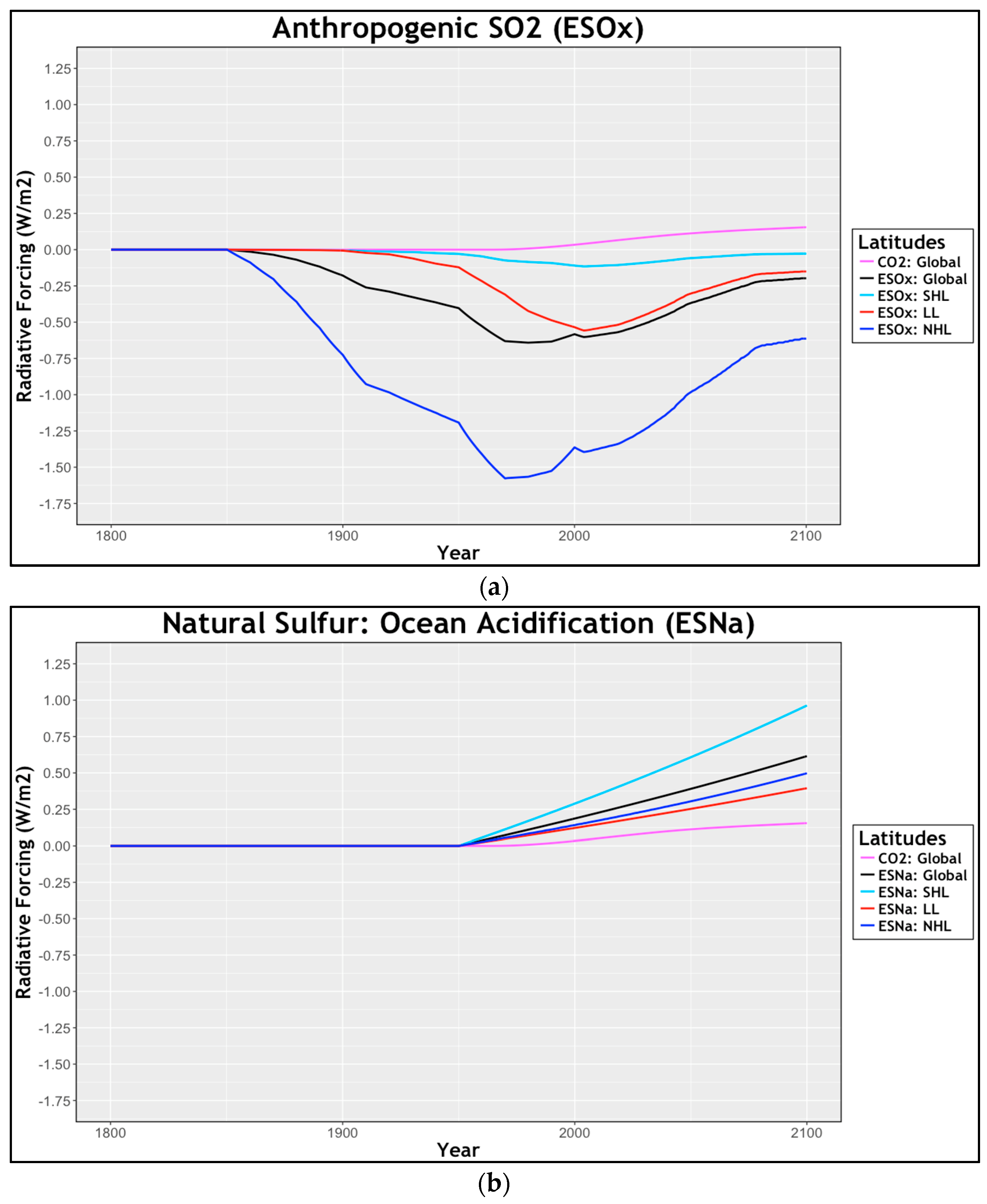
2.1.1. Radiative Forcing: Anthropogenic and ESNa
2.1.2. Anthropogenic Sulfur Emissions
2.1.3. Natural Sulfur Emissions
2.2. Community Shifts (ESNc)
2.2.1. Cyanobacteria and Coccolithophorids
2.2.2. Phaeocystis
2.2.3. Diatoms and Dinoflagellates et al.
2.2.4. Normalization
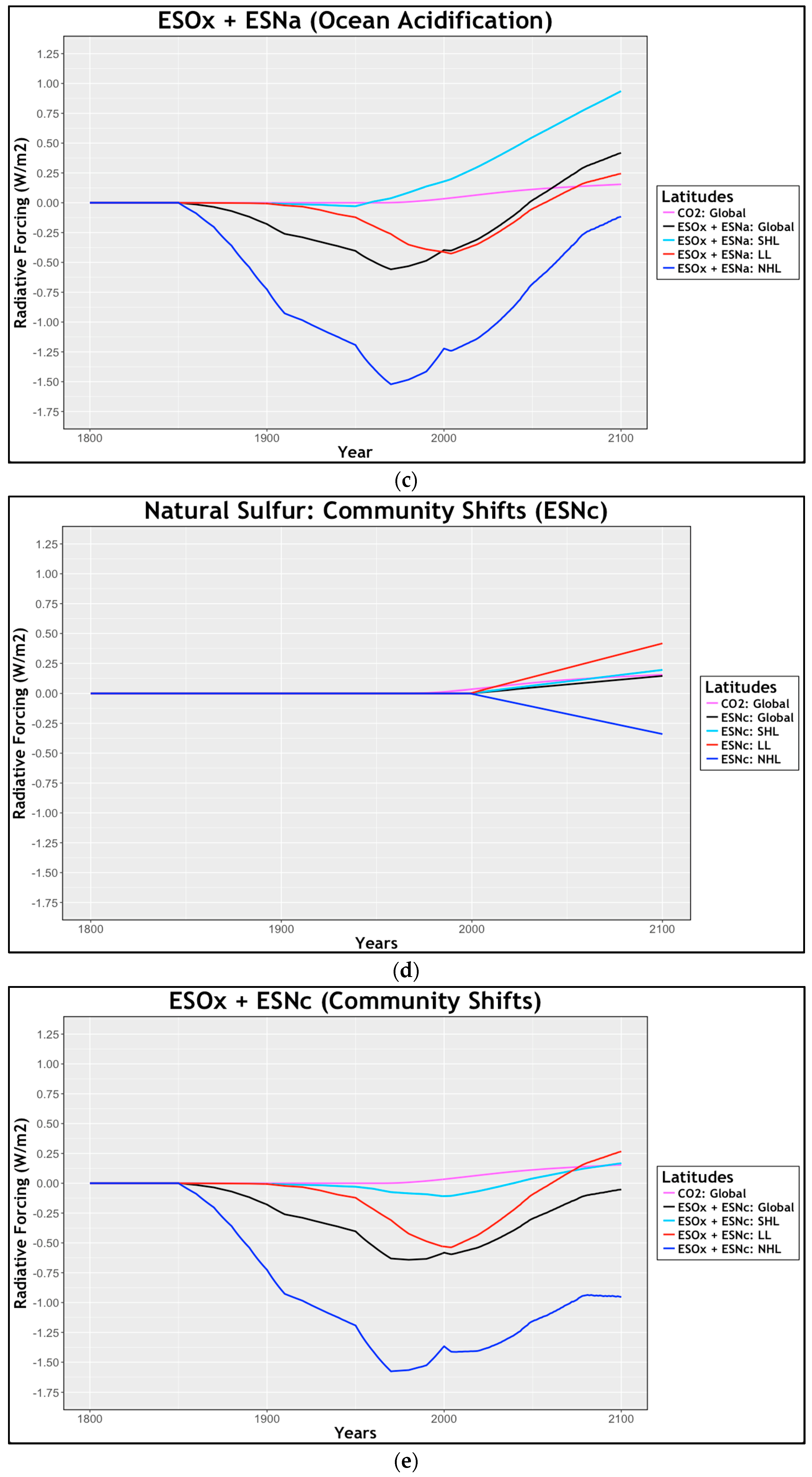
2.2.5. Radiative Forcing: Community Shifts
2.3. CO2 mBGC Feedback
Biological Pump
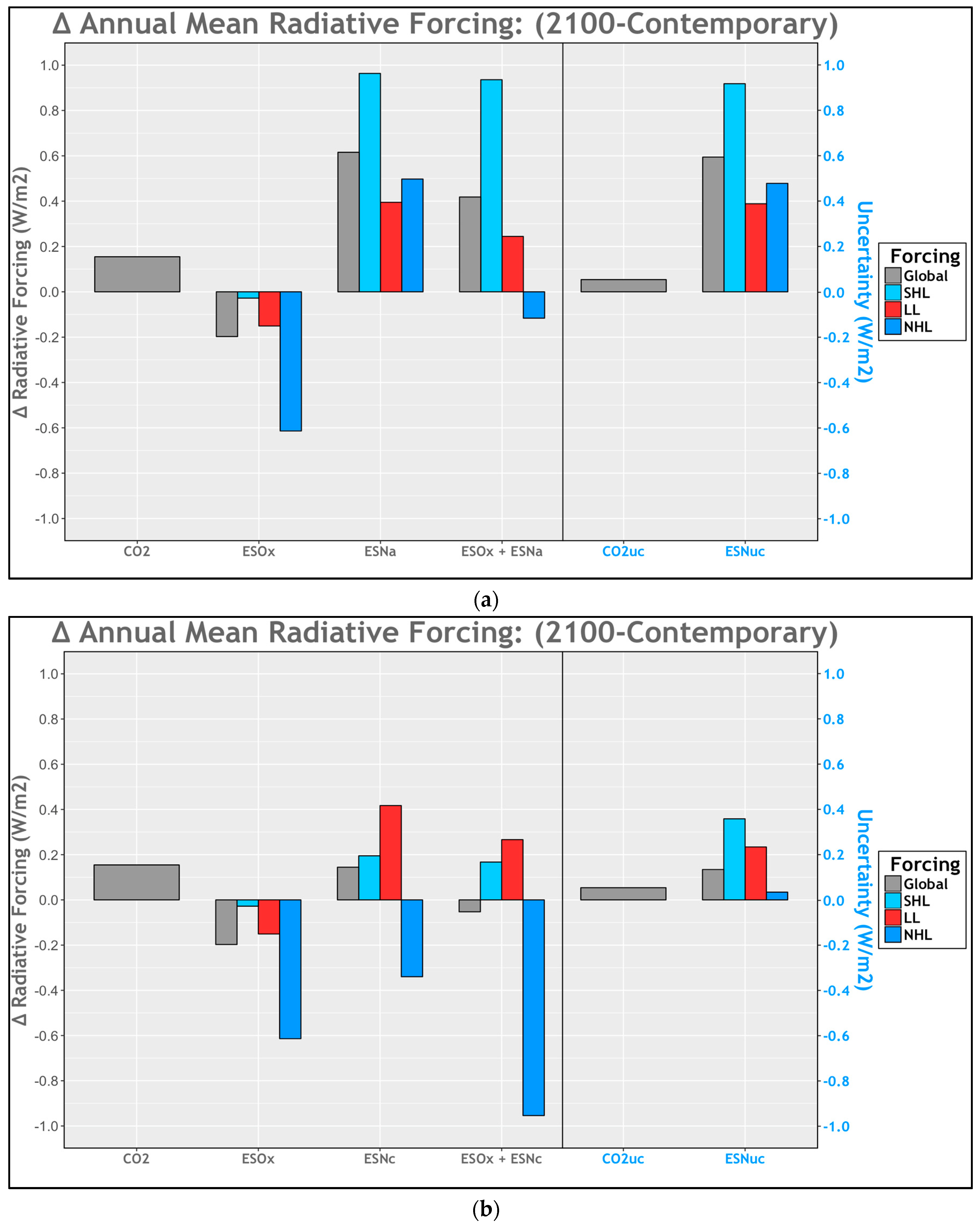
2.4. Sensitivity Tests (ESNuc, CO2uc)
2.4.1. Uncertainty: Ocean Acidification
2.4.2. Uncertainty: Community Shifts
2.4.3. Uncertainty: CO2
3. Results
3.1. Anthropogenic Sulfur Emissions (ESOx)
3.2. Natural Sulfur Emissions: Ocean Acidification (Esna)
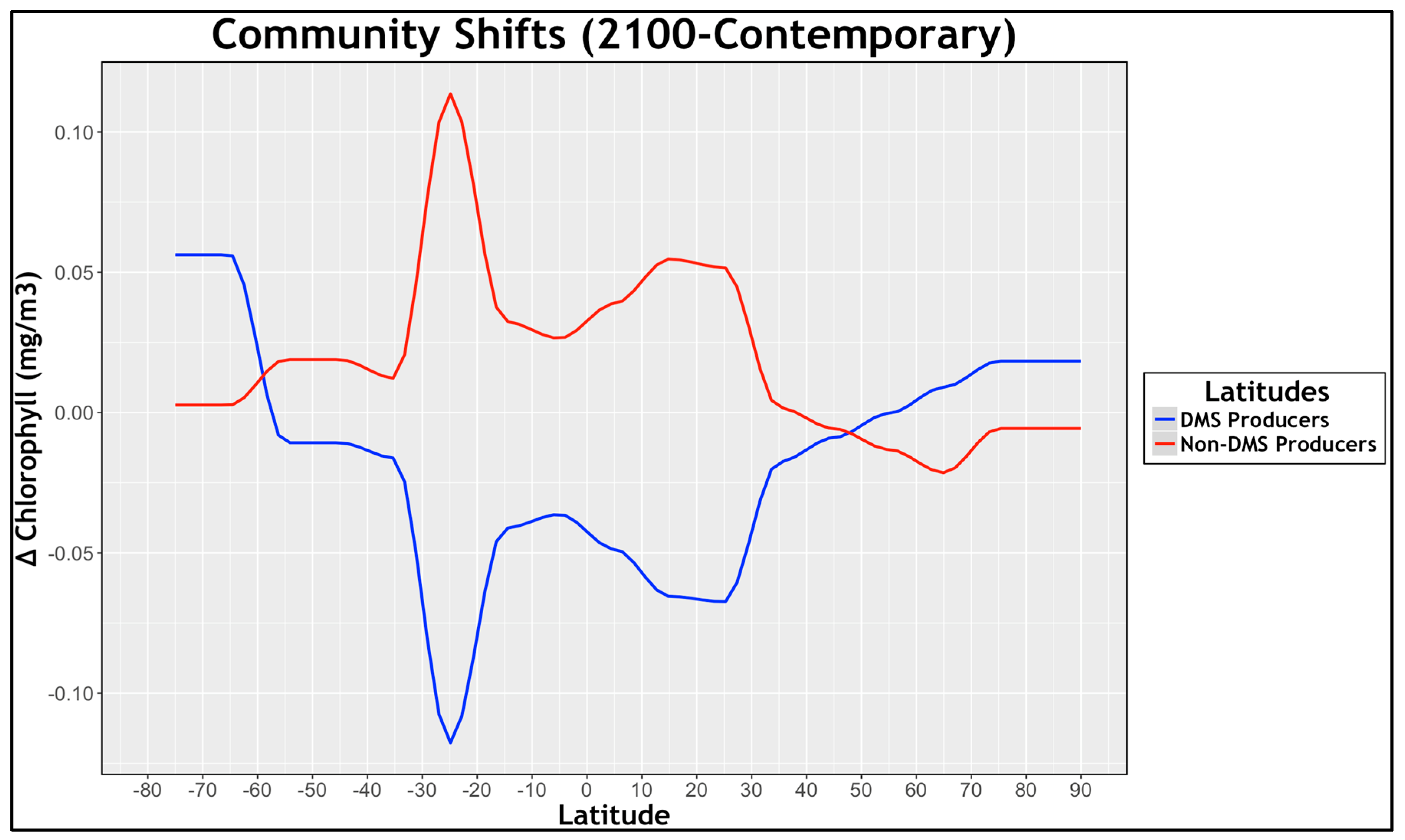
3.3. Natural Sulfur Emissions: Community Shifts (ESNc)
Change in Total Chlorophyll Concentration: (RCP8.5–Contemporary)
3.4. Histograms
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | ESOx (Gg) |
|---|---|
| USA and Canada | 17,054 |
| Western Europe | 7998 |
| Central Europe | 5704 |
| Russia | 6352 |
| Ukraine | 1548 |
| Other Former Soviet Union | 2516 |
| China | 21,393 |
| Japan | 885 |
| Middle East | 5218 |
| Northern High Latitude Total: | 68,668 |
| Mexico | 2991 |
| Central America | 867 |
| South America | 4719 |
| Other South and East Asia | 6330 |
| India | 5363 |
| Africa | 3322 |
| Low Latitude Total: | 23,592 |
| South Africa | 2392 |
| Australia and New Zealand | 2438 |
| Southern High Latitude Total: | 4830 |
| International Shipping: | 9779 |
| Global Total: | 106,869 |
Appendix B
| Latitude | ESN2000 | ESN2099–1865 | ESN1865 | ESN2099 | ESN2099–1950 |
|---|---|---|---|---|---|
| 90–80° S: | 0.000 | 0.0000 | 0.000 | 0.000 | 0.0000 |
| 80–70° S: | 0.017 | 0.0003 | −0.020 | 0.046 | 0.0005 |
| 70–60° S: | 0.373 | −0.0001 | 0.386 | 0.363 | −0.0002 |
| 60–50° S: | 1.113 | −0.0024 | 1.430 | 0.867 | −0.0038 |
| 50–40° S: | 2.033 | −0.0033 | 2.445 | 1.707 | −0.0051 |
| 40–30° S: | 2.127 | −0.0052 | 2.784 | 1.606 | −0.0082 |
| 30–20° S: | 2.237 | −0.0047 | 2.822 | 1.773 | −0.0073 |
| 20–10° S: | 2.593 | −0.0035 | 3.027 | 2.249 | −0.0054 |
| 10–0° S: | 2.817 | −0.0041 | 3.328 | 2.412 | −0.0063 |
| 0–10° N: | 3.387 | −0.0035 | 3.821 | 3.042 | −0.0054 |
| 10–20° N: | 2.860 | −0.0040 | 3.359 | 2.464 | −0.0062 |
| 20–30° N: | 1.563 | −0.0029 | 1.925 | 1.276 | −0.0045 |
| 30–40° N: | 1.297 | −0.0030 | 1.678 | 0.994 | −0.0047 |
| 40–50° N: | 0.833 | −0.0037 | 1.295 | 0.467 | −0.0057 |
| 50–60° N: | 0.500 | −0.0020 | 0.757 | 0.296 | −0.0032 |
| 60–70° N: | 0.147 | −0.0006 | 0.227 | 0.083 | −0.0010 |
| 70–80° N: | 0.200 | 0.0001 | 0.186 | 0.211 | 0.0002 |
| 80–90° N: | 0.003 | 0.0002 | −0.022 | 0.023 | 0.0003 |
| Global Average | 1.339 | −0.0024 | 1.635 | 1.104 | −0.0037 |
Appendix C
| Functional Group | Contemporary | End of Century | Reference |
|---|---|---|---|
| Cyanobacteria | Gregg et al., 2003 | Gregg et al., 2003 Flombaum et al., 2013 | [7,15] |
| Coccolithophorids | Gregg et al., 2003 | Gregg et al., 2003 Jensen et al., 2017 | [15,16] |
| Phaeocystis | Vogt et al., 2012 | Vogt et al., 2012 Wang et al., 2015 Wang et al., (In Review) | [18,19,20] |
| Diatoms | Marinov et al., 2013 | Marinov et al., 2013 | [21] |
| Dinoflagellates et al. | Marinov et al., 2013 Gregg et al., 2003 Vogt et al., 2012 | Marinov et al., 2013 Gregg et al., 2003 Flombaum et al., 2013 Jensen et al., 2017 Vogt et al., 2012 Wang et al., 2015 Wang et al., (In Review) | [7,15,16,18,19,20,21] |
Appendix D
| Reservoir | Initial (PgC) | PV (m s−1) | Remineralization | Nitrate1745 (mmol m−3) | Nitrate2100 (mmol m−3) |
|---|---|---|---|---|---|
| Atmosphere | 750 | - | - | - | - |
| Surface Low Latitude Ocean | 850 | 2.01 × 10−5 | 79% | 5 | 2 |
| Surface High Latitude Ocean | 200 | 3.12 × 10−5 | 60% | 15 | 15 |
| Intermediate Ocean | 9600 | - | 21% (40%) | - | - |
| Deep Ocean | 26,400 | - | - | - | - |
References
- Kloster, S. DMS cycle in the ocean-atmosphere system and its response to anthropogenic perturbations. Rep. Earth Syst. Sci. 2006, 103, 1–103. [Google Scholar]
- Stefels, J.; Steinke, M.; Turner, S.; Malin, G.; Belviso, S. Environmental constraints on the production and removal of the climatically active gas dimethylsulphide (DMS) and implications for ecosystem modeling. Biogeochemistry 2007, 83, 245–275. [Google Scholar] [CrossRef]
- Schwinger, J.; Tjiputra, J.; Goris, N.; Six, K.; Kirkevåg, A.; Seland, Ø.; Heinze, C.; Ilyina, T. Amplification of global warming through pH-dependence of DMS-production simulated with a fully coupled Earth system model. Biogeosci. Discuss. 2017, 20, 1–26. [Google Scholar] [CrossRef]
- Charlson, R.J.; Lovelock, J.E.; Andreae, M.O.; Warren, S.G. Oceanic phytoplankton, atmospheric sulphur, albedo and climate. Nature 1987, 326, 655–661. [Google Scholar] [CrossRef]
- Menzo, Z.M. Web strategy to convey marine biogeochemical feedback concepts to the policy community: Aerosol and sea ice. Atmosphere 2018, 9, 22. [Google Scholar] [CrossRef]
- Six, K.D.; Kloster, S.; Ilyina, T.; Archer, S.D.; Zhang, K.; Maier-Reimer, E. Global warming amplified by reduced sulphur fluxes as a result of ocean acidification. Nat. Clim. Chang. 2013, 3, 975–978. [Google Scholar] [CrossRef]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.; Rincón, J.; Zabala, L.L.; Jiao, N.; Karl, D.; Li, W.K.W.; Lomas, M.; Veneziano, D.; et al. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [PubMed]
- Cameron-Smith, P.; Elliott, S.; Maltrud, M.; Erickson, D.; Wingenter, O. Changes in dimethyl sulfide oceanic distribution due to climate change. Geophys. Res. Lett. 2011, 38, 1–5. [Google Scholar] [CrossRef]
- Hartin, C.A.; Patel, P.; Schwarber, A.; Link, R.P.; Bond-Lamberty, B.P. A simple object-oriented and open-source model for scientific and policy analyses of the global climate system—Hector v1.0. Geosci. Model Dev. 2015, 8, 939–955. [Google Scholar] [CrossRef]
- Joos, F.; Prentice, C.; Sitch, S.; Meyer, R.; Hooss, G.; Plattner, G.K.; Gerber, S.; Hasselmann, K. Global warming feedbacks on terrestrial carbon uptake under the intergovernmental Panel on Climate Change (IPCC) emission scenarios. Glob. Biogeochem. Cycles 2001, 15, 891–907. [Google Scholar] [CrossRef]
- Smith, S.J.; Van Aardenne, J.; Klimont, Z.; Andres, R.J.; Volke, A.; Delgado Arias, S. Anthropogenic sulfur dioxide emissions: 1850–2005. Atmos. Chem. Phys. 2011, 11, 1101–1116. [Google Scholar] [CrossRef] [Green Version]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Nakicenovic, N.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Simó, R.; Dachs, J. Global ocean emission of dimethylsulfide predicted from biogeophysical data. Glob. Biogeochem. Cycles 2002, 16. [Google Scholar] [CrossRef]
- Fu, W.; Randerson, J.T.; Keith Moore, J. Climate change impacts on net primary production (NPP) and export production (EP) regulated by increasing stratification and phytoplankton community structure in the CMIP5 models. Biogeosciences 2016, 13, 5151–5170. [Google Scholar] [CrossRef]
- Gregg, W.W.; Ginoux, P.; Schopf, P.S.; Casey, N.W. Phytoplankton and iron: Validation of a global three-dimensional ocean biogeochemical model. Deep-Sea Res. Part II Top. Stud. Oceanogr. 2003, 50, 3143–3169. [Google Scholar] [CrossRef]
- Jensen, L.Ø.; Mousing, E.A.; Richardson, K. Using species distribution modelling to predict future distributions of phytoplankton: Case study using species important for the biological pump. Mar. Ecol. 2017, 38. [Google Scholar] [CrossRef]
- NOAA Climate Model: Temperature Change (RCP 8.5)—2006–2100. Available online: https://sos.noaa.gov/datasets/climate-model-temperature-change-rcp-85-2006-2100 (accessed on 15 August 2017).
- Vogt, M.; O’Brien, C.; Peloquin, J.; Schoemann, V.; Breton, E.; Estrada, M.; Gibson, J.; Karentz, D.; Van Leeuwe, M.A.; Stefels, J.; et al. Global marine plankton functional type biomass distributions: Phaeocystis spp. Earth Syst. Sci. Data 2012, 4, 107–120. [Google Scholar] [CrossRef]
- Wang, S.; Elliott, S.; Maltrud, M.; Cameron-Smith, P. Influence of explicit Phaeocystis parameterizations on the global distribution of marine dimethyl sulfide. J. Geophys. Res. G Biogeosci. 2015, 120, 2158–2177. [Google Scholar] [CrossRef]
- Wang, S.; Maltrud, M.; Burrows, S.; Elliott, S.; Cameron-Smith, P. Impacts of shifts in phytoplankton community on clouds and climate via the sulfur cycle. Glob. Biogeochem. Cycles. in review.
- Marinov, I.; Doney, S.C.; Lima, I.D.; Lindsay, K.; Moore, J.K.; Mahowald, N. North-South asymmetry in the modeled phytoplankton community response to climate change over the 21st century. Glob. Biogeochem. Cycles 2013, 27, 1274–1290. [Google Scholar] [CrossRef]
- Elliott, S. Dependence of DMS global sea-air flux distribution on transfer velocity and concentration field type. J. Geophys. Res. Biogeosci. 2009, 114, 1–18. [Google Scholar] [CrossRef]
- Sarmiento, J.L.; Slater, R.D.; Fasham, M.J.R.; Ducklow, H.W.; Toggweiler, J.R.; Evans, G.T. A seasonal three-dimensional ecosystem model of nitrogen cycling in the North Atlantic Euphotic zone. Glob. Biogeochem. Cycles 1993, 7, 417–450. [Google Scholar] [CrossRef]
- Huebert, B.J.; Blomquist, B.W.; Hare, J.E.; Fairall, C.W.; Johnson, J.E.; Bates, T.S. Measurement of the sea-air DMS flux and transfer velocity using eddy correlation. Geophys. Res. Lett. 2004, 31. [Google Scholar] [CrossRef]
- Lana, A.; Bell, T.G.; Simó, R.; Vallina, S.M.; Ballabrera-Poy, J.; Kettle, A.J.; Dachs, J.; Bopp, L.; Saltzman, S.; Stefels, J.; et al. An updated climatology of surface dimethlysulfide concentrations and emission fluxes in the global ocean. Glob. Biogeochem. Cycles 2011, 25. [Google Scholar] [CrossRef]
- Watson, A.J.; Liss, P.S. Marine biological controls on climate via the carbon and sulphur geochemical cycles. Philos. Trans. R. Soc. B Biol. Sci. 1998, 353, 41–51. [Google Scholar] [CrossRef]
- Sarmiento, J.L.; Gruber, N. Ocean Biogeochemical Dynamics, 1st ed.; Princeton University Press: Princeton, NJ, USA, 2006; pp. 103–326. ISBN 0-691-01707-7. [Google Scholar]
- Byrne, B.; Goldblatt, C. Radiative forcing at high concentrations of well-mixed greenhouse gases. Geophys. Res. Lett. 2014, 41, 152–160. [Google Scholar] [CrossRef]
- Henson, S.A.; Sanders, R.; Madsen, E. Global patterns in efficiency of particulate organic carbon export and transfer to the deep ocean. Glob. Biogeochem. Cycles 2012, 26. [Google Scholar] [CrossRef]
- Marine Biota Exchange—The Biological Pump. Available online: https://www.e-education.psu.edu/earth103/node/1022 (accessed on 20 September 2017).
- Crueger, T.; Roeckner, E.; Raddatz, T.; Schnur, R.; Wetzel, P. Ocean dynamics determine the response of oceanic CO2 uptake to climate change. Clim. Dyn. 2008, 31, 151–168. [Google Scholar] [CrossRef]
- Friedlingstein, P.; Cox, P.; Betts, R.; Bopp, L.; von Bloh, W.; Brovkin, V.; Cadule, P.; Doney, S.; Eby, M.; Fung, I.; et al. Climate–Carbon Cycle Feedback Analysis: Results from the C4MIP Model Intercomparison. J. Clim. 2006, 19, 3337–3353. [Google Scholar] [CrossRef]
- Myhre, G.; Shindell, D.; Bréon, F.-M.; Collins, W.; Fuglestvedt, J.; Huang, J.; Koch, D.; Lamarque, J.-F.; Lee, D.; Mendoza, B.; et al. Anthropogenic and Natural Radiative Forcing. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 659–740. ISBN 978-1-107-05799-1. [Google Scholar]




| Location | ESOx (Tg) | Percent Total (ESOx) | ||
|---|---|---|---|---|
| Southern High Latitude: | 4.830 | 4.52% | −0.110 | 0.963 |
| Low Latitude: | 23.592 | 22.08% | −0.538 | 0.395 |
| Northern High Latitude: | 68.668 | 64.25% | −1.567 | 0.498 |
| International Shipping: | 9.779 | 9.15% | −0.223 | - |
| Global Total: | 106.869 | - | −0.610 | 0.615 |
| Latitude | Biomes | Diatoms | Cyano. | Dino. | Cocco. | Phaeo. |
|---|---|---|---|---|---|---|
| 90–80° S: | - | 0.0 (0.0) | 0.0 (0.0) | 0.0 (0.0) | 0.0 (0.0) | 0.0 (0.0) |
| 80–70° S: | Ice SH | 96.1 (98.8) | 0.0 (0.0) | 0.0 (114.0) | 4.6 (13.9) | 427.8 (360.7) |
| 70–60° S: | Ice SH | 96.1 (98.8) | 0.0 (0.0) | 0.0 (184.4) | 105.6 (115.5) | 326.8 (188.7) |
| 60–50° S: | Subpolar SH | 120.1 (139.0) | 0.0 (0.0) | 104.7 (94.4) | 10.6 (10.2) | 0.0 (0.0) |
| 50–40° S: | Subpolar SH | 120.1 (139.0) | 0.0 (0.0) | 101.1 (82.7) | 0.4 (0.3) | 13.9 (21.6) |
| 40–30° S: | Subtropical SH | 88.9 (89.7) | 0.0 (11.4) | 141.3 (126.0) | 5.2 (4.4) | 0.0 (0.0) |
| 30–20° S: | Subtropical SH | 88.9 (89.7) | 17.3 (130.1) | 112.0 (0.0) | 17.3 (11.6) | 0.0 (0.0) |
| 20–10° S: | LLU SH | 88.9 (85.2) | 41.9 (78.1) | 87.0 (46.3) | 0.8 (0.3) | 0.0 (0.0) |
| 10–0° S: | Equatorial | 108.1 (103.6) | 39.3 (70.3) | 82.9 (46.8) | 0.3 (0.2) | 0.0 (0.0) |
| 0–10° N: | Equatorial | 108.1 (103.6) | 38.1 (81.4) | 81.4 (32.8) | 3.1 (2.9) | 0.0 (0.0) |
| 10–20° N: | Upwelling NH | 84.1 (78.7) | 47.6 (107.7) | 66.7 (6.8) | 8.1 (3.8) | 4.9 (3.7) |
| 20–30° N: | Subtropical NH | 62.5 (53.0) | 63.8 (124.9) | 0.0 (0.0) | 14.5 (2.7) | 63.3 (7.8) |
| 30–40° N: | Subtropical NH | 62.5 (53.0) | 55.8 (67.0) | 49.1 (40.4) | 4.1 (3.6) | 32.7 (24.3) |
| 40–50° N: | - | 86.5 (75.4) | 28.1 (33.5) | 24.6 (43.2) | 16.1 (12.1) | 63.4 (39.9) |
| 50–60° N: | Subpolar NH | 110.5 (97.7) | 0.3 (0.0) | 0.0 (46.0) | 28.1 (20.5) | 94.1 (55.4) |
| 60–70° N: | - | 96.1 (74.8) | 0.2 (0.0) | 0.0 (33.6) | 14.1 (10.3) | 71.1 (50.3) |
| 70–80° N: | Ice NH | 57.7 (52.0) | 0.0 (0.0) | 0.0 (21.1) | 0.0 (0.0) | 48.0 (45.2) |
| 80–90° N: | Ice NH | 57.7 (52.0) | 0.0 (0.0) | 0.0 (34.3) | 0.0 (0.0) | 48.0 (32.1) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menzo, Z.M.; Elliott, S.; Hartin, C.A.; Hoffman, F.M.; Wang, S. Climate Change Impacts on Natural Sulfur Production: Ocean Acidification and Community Shifts. Atmosphere 2018, 9, 167. https://doi.org/10.3390/atmos9050167
Menzo ZM, Elliott S, Hartin CA, Hoffman FM, Wang S. Climate Change Impacts on Natural Sulfur Production: Ocean Acidification and Community Shifts. Atmosphere. 2018; 9(5):167. https://doi.org/10.3390/atmos9050167
Chicago/Turabian StyleMenzo, Zachary M., Scott Elliott, Corinne A. Hartin, Forrest M. Hoffman, and Shanlin Wang. 2018. "Climate Change Impacts on Natural Sulfur Production: Ocean Acidification and Community Shifts" Atmosphere 9, no. 5: 167. https://doi.org/10.3390/atmos9050167
APA StyleMenzo, Z. M., Elliott, S., Hartin, C. A., Hoffman, F. M., & Wang, S. (2018). Climate Change Impacts on Natural Sulfur Production: Ocean Acidification and Community Shifts. Atmosphere, 9(5), 167. https://doi.org/10.3390/atmos9050167