Identification of the Invasive Form of Corbicula Clams in Ireland

Abstract
:1. Introduction
Aims
2. Materials and Methods
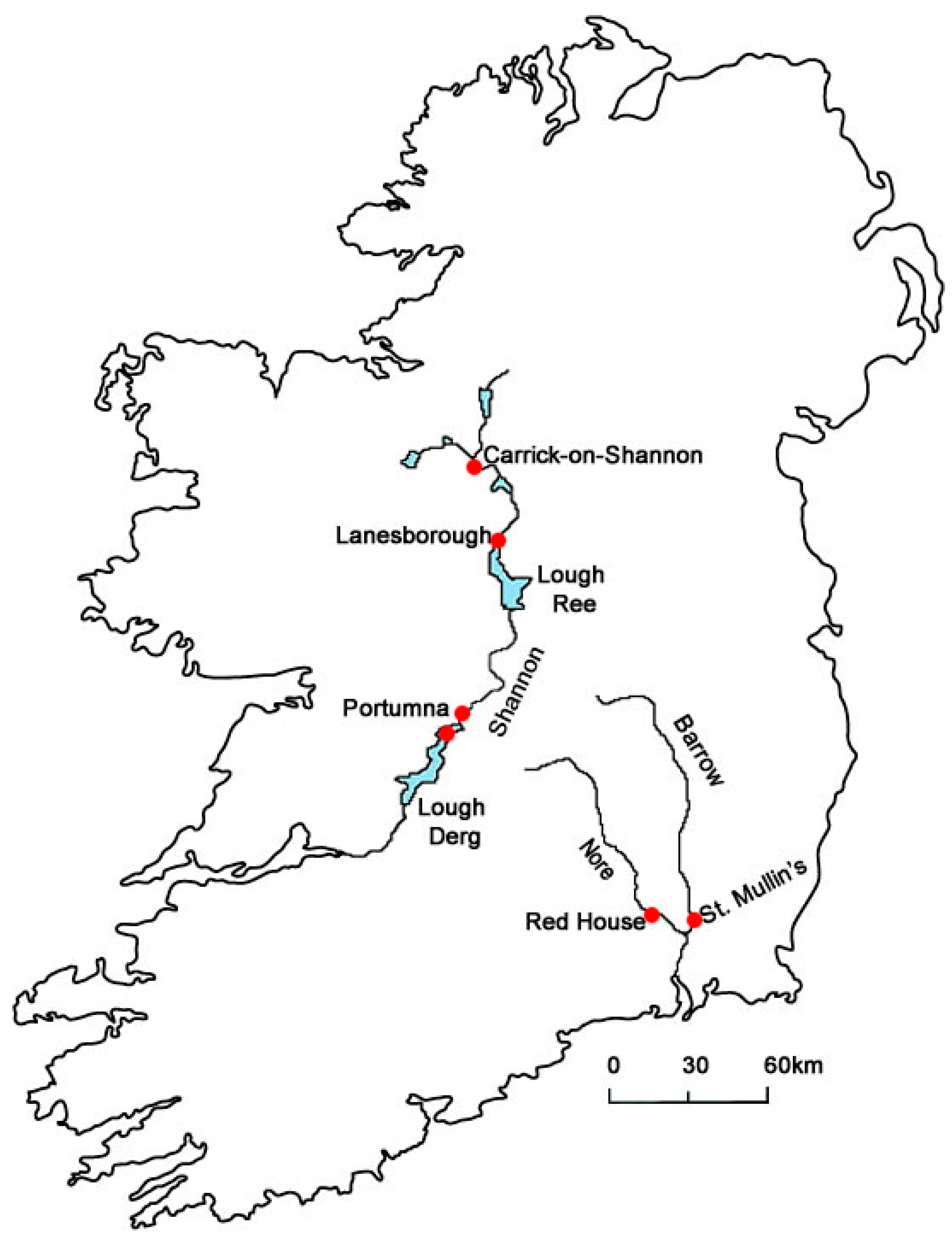
2.1. Specimen Collection
2.2. Morphological Analysis
2.3. DNA Extraction
2.4. Mitochondrial COI Gene Analysis
2.5. Microsatellite Marker Analysis
3. Results
3.1. Morphology
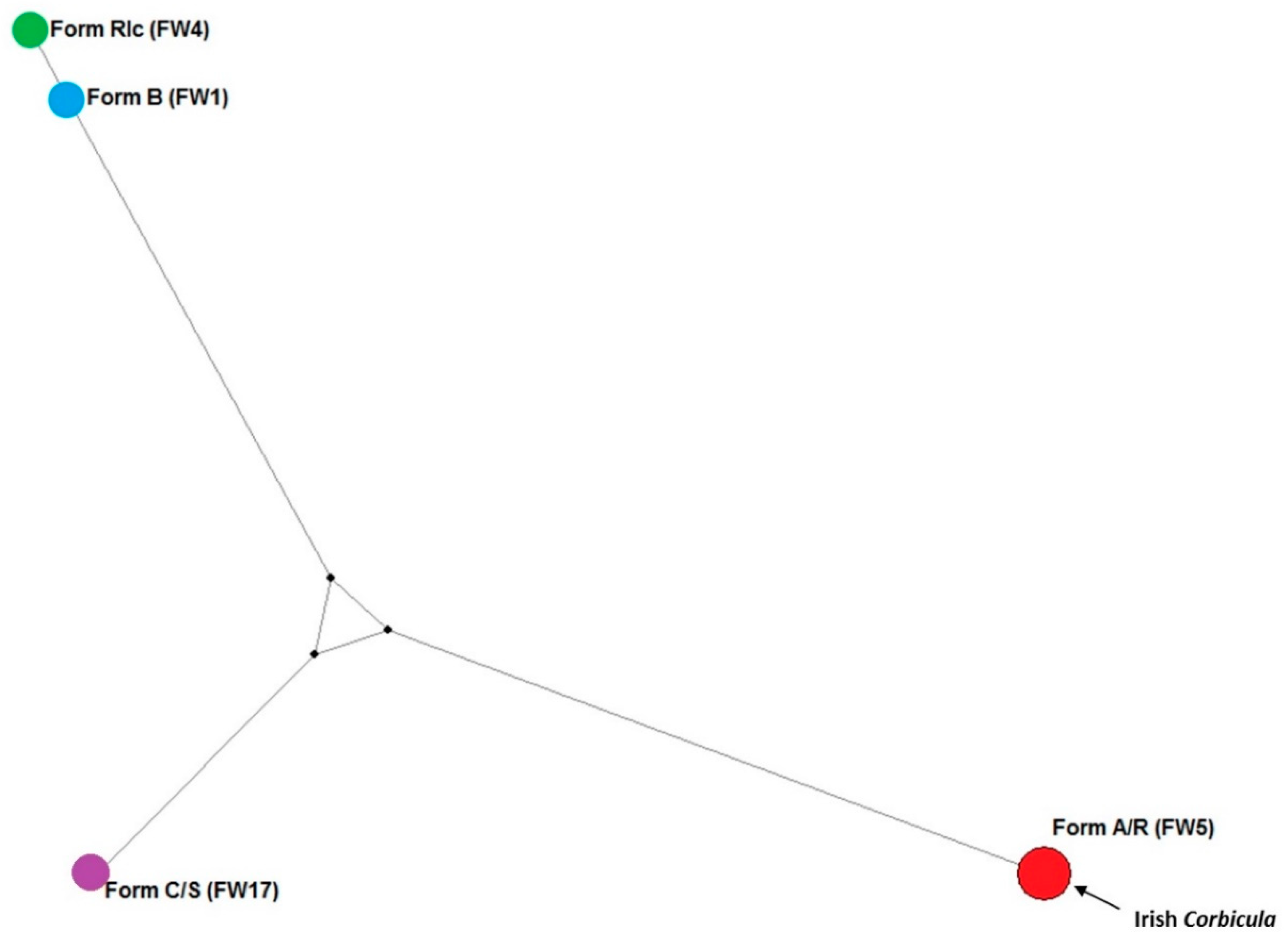
3.2. COI Sequence
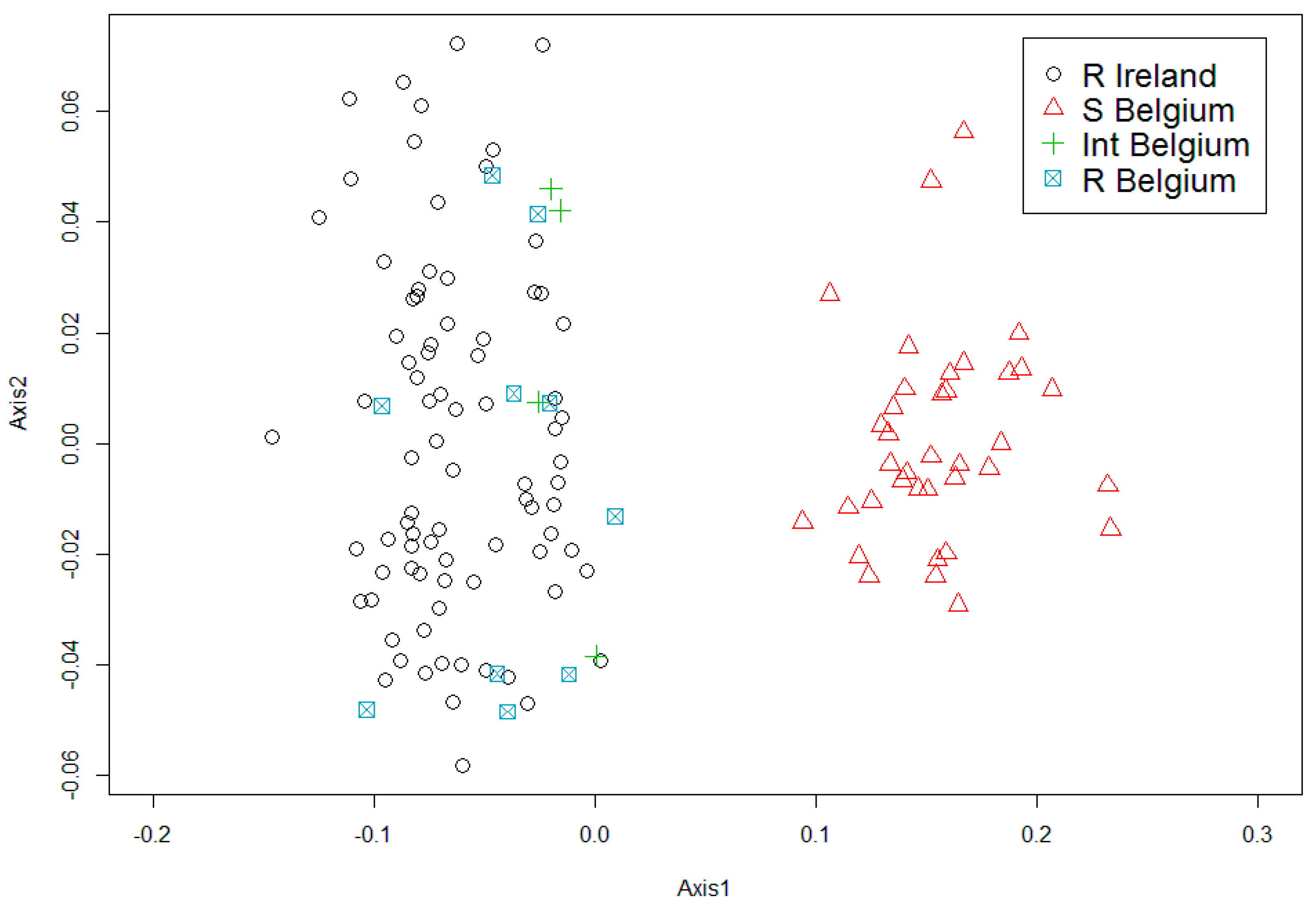
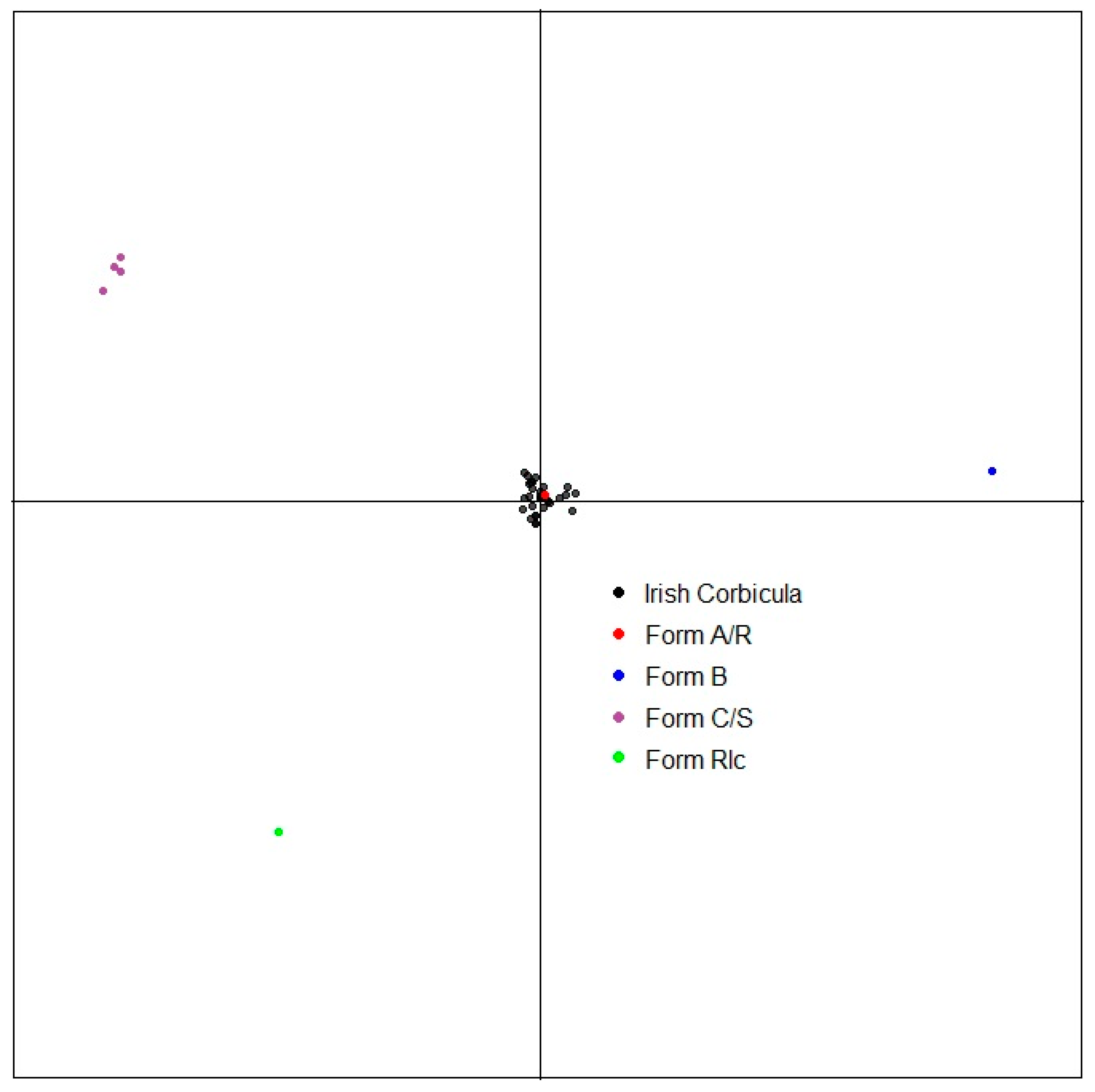
3.3. Microsatellite Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mouthon, J. Sur la présence en France et au Portugal de Corbicula (Bivalvia, Corbiculidae) originaire d’Asie. Basteria 1981, 45, 109–116. [Google Scholar]
- McMahon, R.F. The occurrence and spread of the introduced Asiatic fresh-water clam, Corbicula-fluminea (Müller), in North-America-1924-1982. Nautilus 1982, 96, 134–141. [Google Scholar]
- Kraemer, L.R.; Galloway, M.L. Larval development of Corbicula-fluminea (Muller) (Bivalvia, Corbicula Cea)—An appraisal of its heterochrony. Am. Malacol. Bull. 1986, 4, 61–79. [Google Scholar]
- Araujo, R.; Moreno, D.; Ramos, M.A. The Asiatic clam Corbicula fluminea (Müller, 1774)(Bivalvia: Corbiculidae) in Europe. Am. Malacol. Bull. 1993, 10, 39–49. [Google Scholar]
- Komaru, A.; Kawagishi, T.; Konishi, K. Cytological evidence of spontaneous androgenesis in the freshwater clam Corbicula leana Prime. Dev. Genes Evol. 1998, 208, 46–50. [Google Scholar] [CrossRef]
- Komaru, A.; Ookubo, K.; Kiyomoto, M. All meiotic chromosomes and both centrosomes at spindle pole in the zygotes discarded as two polar bodies in clam Corbicula leana: Unusual polar body formation observed by antitubulin immunofluorescence. Dev. Genes Evol. 2000, 210, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Pigneur, L.M.; Hedtke, S.M.; Etoundi, E.; Van Doninck, K. Androgenesis: A review through the study of the selfish shellfish Corbicula spp. Heredity 2012, 108, 581. [Google Scholar] [CrossRef]
- Counts, C.L. Corbicula fluminea (Bivalvia: Sphaeriacea) in British Columbia. Nautilus 1981, 95, 12–13. [Google Scholar]
- Matthews, M.A.; McMahon, R.F. Effects of temperature and temperature acclimation on survival of zebra mussels (Dreissena polymorpha) and Asian clams (Corbicula fluminea) under extreme hypoxia. J. Molluscan Stud. 1999, 65, 317–325. [Google Scholar] [CrossRef]
- Ituarte, C.F. Corbicula and Neo Corbicula (Bivalvia: Corbiculidae) in the Paraná, Uruguay and Río de La Plata Basins. Nautilus 1994, 107, 129–135. [Google Scholar]
- Pigneur, L.; Etoundi, E.; Aldridge, D.C.; Marescaux, J.; Yasuda, N.; Van Doninck, K. Genetic uniformity and long-distance clonal dispersal in the invasive androgenetic Corbicula clams. Mol. Ecol. 2014, 23, 5102–5116. [Google Scholar] [CrossRef] [PubMed]
- Delivering Alien Invasive Species Inventories for Europe (DAISIE). Available online: http://www.europe-aliens.org/ (accessed on 20 May 2007).
- Aldridge, D.C.; Müller, S.J. The Asiatic clam, Corbicula fluminea, in Britain: Current status and potential impacts. J. Conchol. 2001, 37, 177–183. [Google Scholar]
- Sweeney, P. First record of Asian Clam (Corbicula fluminea (Müller, 1774) in Ireland. Irish Nat. J. 2009, 30, 147–148. [Google Scholar]
- Sheehan, R.; Caffrey, J.M.; Millane, M.; McLoone, P.; Moran, H.; Lucy, F. An investigation into the effectiveness of mechanical dredging to remove Corbicula fluminea (Müller, 1774) from test plots in an Irish river system. Manag. Biol. Invasions 2014, 5, 407–418. [Google Scholar] [CrossRef]
- Caffrey, J.M.; Evers, S.; Millane, M.; Moran, H. Current status of Ireland’s newest invasive species-the Asian clam Corbicula fluminea (Mueller, 1774). Aquat. Invasions 2011, 6, 291–299. [Google Scholar] [CrossRef]
- Hayden, B.; Caffrey, J.M. First recording of the Asian Clam (Corbicula fluminea (Müller, 1774)) from the River Shannon, with preliminary notes on population size and size class distribution. Irish Nat. J. 2013, 32, 29–31. [Google Scholar]
- Minchin, D. The distribution of the Asian clam Corbicula fluminea and its potential to spread in Ireland. Manag Biol Invasions 2014, 5, 165–177. [Google Scholar] [CrossRef]
- Pfenninger, M.; Reinhardt, F.; Streit, B. Evidence for cryptic hybridization between different evolutionary lineages of the invasive clam genus Corbicula (Veneroida, Bivalvia). J. Evol. Biol. 2002, 15, 818–829. [Google Scholar] [CrossRef]
- Park, J.K.; Kim, W. Two Corbicula (Corbiculidae: Bivalvia) mitochondrial lineages are widely distributed in Asian freshwater environment. Mol. Phylogenet. Evol. 2003, 29, 529–539. [Google Scholar] [CrossRef]
- Lee, T.; Siripattrawan, S.; Ituarte, C.F.; Foighil, D.O. Invasion of the clonal clams: Corbicula lineages in the New World. Am. Malacol. Bull. 2005, 20, 113–122. [Google Scholar]
- Sousa, R.; Freire, R.; Rufino, M.; Méndez, J.; Gaspar, M.; Antunes, C.; Guilhermino, L. Genetic and shell morphological variability of the invasive bivalve Corbicula fluminea (Müller, 1774) in two Portuguese estuaries. Estuar. Coast. Shelf Sci. 2007, 74, 166–174. [Google Scholar] [CrossRef]
- Pigneur, L.M.; Marescaux, J.; Roland, K.; Etoundi, E.; Descy, J.P.; Van Doninck, K. Phylogeny and androgenesis in the invasive Corbicula clams (Bivalvia, Corbiculidae) in Western Europe. BMC Evol. Biol. 2011, 11, 147. [Google Scholar] [CrossRef] [PubMed]
- Pigneur, L.M.; Risterucci, A.M.; Dauchot, N.; Li, X.; Van Doninck, K. Development of novel microsatellite markers to identify the different invasive lineages in the Corbicula complex and to assess androgenesis. Mol. Ecol. Resour. 2011, 11, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Hillis, D.M.; Patton, J.C. Morphological and electrophoretic evidence for two species of Corbicula (Bivalvia: Corbiculidae) in North America. Am. Midl. Nat. 1982, 108, 74–80. [Google Scholar] [CrossRef]
- Marescaux, J.; Pigneur, L.M.; Van Doninck, K. New records of Corbicula clams in French rivers. Aquat. Invasions 2010, 5, S35–S39. [Google Scholar] [CrossRef]
- Etoundi, E.; Marescaux, J.; Vastrade, M.; Debortoli, N.; Hedtke, S.M.; Pigneur, L.M.; Virgo, J.; Flot, J.F.; Van Doninck, K. Distinct biogeographic origins of androgenetic Corbicula lineages followed by genetic captures. bioRxiv 2019, 590836. [Google Scholar] [CrossRef]
- Hedtke, S.M.; Stanger-Hall, K.; Baker, R.J.; Hillis, D.M. All-male asexuality: Arigin and maintenance ofandrogenesis in the Asian clam Corbicula. Evolution 2008, 62, 1119–1136. [Google Scholar]
- Park, J.K.; Lee, J.S.; Kim, W. A single mitochondrial lineage is shared by morphologically and allozymatically distinct freshwater Corbicula clones. Mol. Cells 2002, 14, 318–322. [Google Scholar]
- Tiemann, J.S.; Haponski, A.E.; Douglass, S.A.; Lee, T.; Cummings, K.S.; Davis, M.A.; Foighil, D.Ó. First record of a putative novel invasive Corbicula lineage discovered in the Illinois River, Illinois, USA. BioInvasions Rec. 2017, 6, 159–166. [Google Scholar] [CrossRef]
- Komaru, A.; Yamada, M.; Houki, S. Relationship between two androgenetic clam species, Corbicula leana and Corbicula fluminea, inferred from mitochondrial cytochrome b and nuclear 28S rRNA markers. Zoolog. Sci. 2013, 30, 360–366. [Google Scholar] [CrossRef]
- Lucy, F.E.; Karatayev, A.Y.; Burlakova, L.E. Predictions for the spread, population density, and impacts of Corbicula fluminea in Ireland. Aquat. Invasions 2012, 7, 465–474. [Google Scholar] [CrossRef]
- Minchin, D.; Boelens, R. Natural dispersal of the introduced Asian clam Corbicula fluminea (Müller, 1774) (Cyrenidae) within two temperate lakes. Bioinvasions Rec. 2018, 7, 259–268. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Series 1999, 41, 95–98. [Google Scholar]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [PubMed]
- Legendre, L.; Legrendre, L. Numerical Ecology. Developments in Environmental Modelling; Elsevier Science & Technology: Amsterdam, The Nederland, 1998; ISBN 1281021598. [Google Scholar]
- Renard, E.; Bachmann, V.; Cariou, M.L.; Moreteau, J.C. Morphological and molecular differentiation of invasive freshwater species of the genus Corbicula (Bivalvia,Corbiculidea) suggest the presence of three taxa in French rivers. Mol. Ecol. 2000, 9, 2009–2016. [Google Scholar] [CrossRef]
- Minchin, D. The Asian Clam (Corbicula fluminea (OF Müller))(Cyrenidae) arrives in the Erne River. Irish Nat. J. 2017, 35, 99–104. [Google Scholar]
- Caffrey, J.M.; Dick, J.T.A.; Lucy, F.E.; Davis, E.; Niven, A.; Coughlan, N.E. First record of the Asian clam Corbicula fluminea (Müller, 1774)(Bivalvia, Cyrenidae) in Northern Ireland. BioInvasions Rec. 2016, 5, 239–244. [Google Scholar] [CrossRef]
- Peñarrubia, L.; Viñas, J.; Sanz, N.; Smith, B.L.; Bremer, J.R.A.; Pla, C.; Vidal, O. SNP identification and validation in two invasive species: Zebra mussel (Dreissena polymorpha) and Asian clam (Corbicula fluminea). Anim. Biodivers. Conserv. 2019, 42, 65–68. [Google Scholar] [CrossRef]
- Schmidlin, S.; Schmera, D.; Ursenbacher, S.; Baur, B. Separate introductions but lack of genetic variability in the invasive clam Corbicula spp. in Swiss lakes. Aquat. Invasions 2012, 7, 73–80. [Google Scholar] [CrossRef]
- Gherardi, F.; Aquiloni, L.; Diéguez-Uribeondo, J.; Tricarico, E. Managing invasive crayfish: Is there a hope? Aquat. Sci. 2011, 73, 185–200. [Google Scholar] [CrossRef]
- González-Moreno, P.; Lazzaro, L.; Vilà, M.; Preda, C.; Adriaens, T.; Bacher, S.; Brundu, G.; Copp, G.H.; Essl, F.; García-Berthou, E. Consistency of impact assessment protocols for non-native species. NeoBiota 2019, 44, 1–25. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | No | Site | Sampling Method | Grid Reference |
|---|---|---|---|---|
| Shannon (Carrick-on-Shannon) | 1 | 1 km above town | Diving | 53.951402, −8.109267 |
| Shannon (Lanesborough) | 2 | Below bridge | Grab + dredge | 53.669457, −7.998963 |
| Shannon (Portumna) | 3 | Below bridge | Grab + dredge | 53.091139, −8.195643 |
| Lough Derg | 4 | Upper lake | Grab + dredge | 52.928985, −8.280803 |
| River Barrow | 5 | St Mullin’s | Diving | 52.487025, −6.926753 |
| River Nore | 6 | Redhouse | Kick-net | 52.469403, −7.044622 |
| River Meuse | 7 | Meuse River, Petit Lanaye, Belgium | Kick-net | 50.810898, −5.692482 |
| Site | Described Form | Morphology | mtCOI | Microsatellites |
|---|---|---|---|---|
| Carrick-on-Shannon | R | 11 | 5 | 9 |
| Lanesborough | R | 9 | 5 | 9 |
| Lough Derg | R | 39 | 5 | 14 |
| River Barrow | R | 10 | 4 | 9 |
| Portumna | R | 10 | 1 | 2 |
| River Nore | R | 5 | 5 | 5 |
| Meuse River | R | 10 | N/A | N/A |
| S | 37 | N/A | N/A | |
| Int | 4 | N/A | N/A |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheehan, R.; Etoundi, E.; Minchin, D.; Doninck, K.V.; Lucy, F. Identification of the Invasive Form of Corbicula Clams in Ireland. Water 2019, 11, 1652. https://doi.org/10.3390/w11081652
Sheehan R, Etoundi E, Minchin D, Doninck KV, Lucy F. Identification of the Invasive Form of Corbicula Clams in Ireland. Water. 2019; 11(8):1652. https://doi.org/10.3390/w11081652
Chicago/Turabian StyleSheehan, Rory, Emilie Etoundi, Dan Minchin, Karine Van Doninck, and Frances Lucy. 2019. "Identification of the Invasive Form of Corbicula Clams in Ireland" Water 11, no. 8: 1652. https://doi.org/10.3390/w11081652
APA StyleSheehan, R., Etoundi, E., Minchin, D., Doninck, K. V., & Lucy, F. (2019). Identification of the Invasive Form of Corbicula Clams in Ireland. Water, 11(8), 1652. https://doi.org/10.3390/w11081652