

Water and Sediment Chemistry as Drivers of Macroinvertebrates and Fish Assemblages in Littoral Zones of Subtropical Reservoirs

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Methods and Materials
2.1. Ethical Clearance
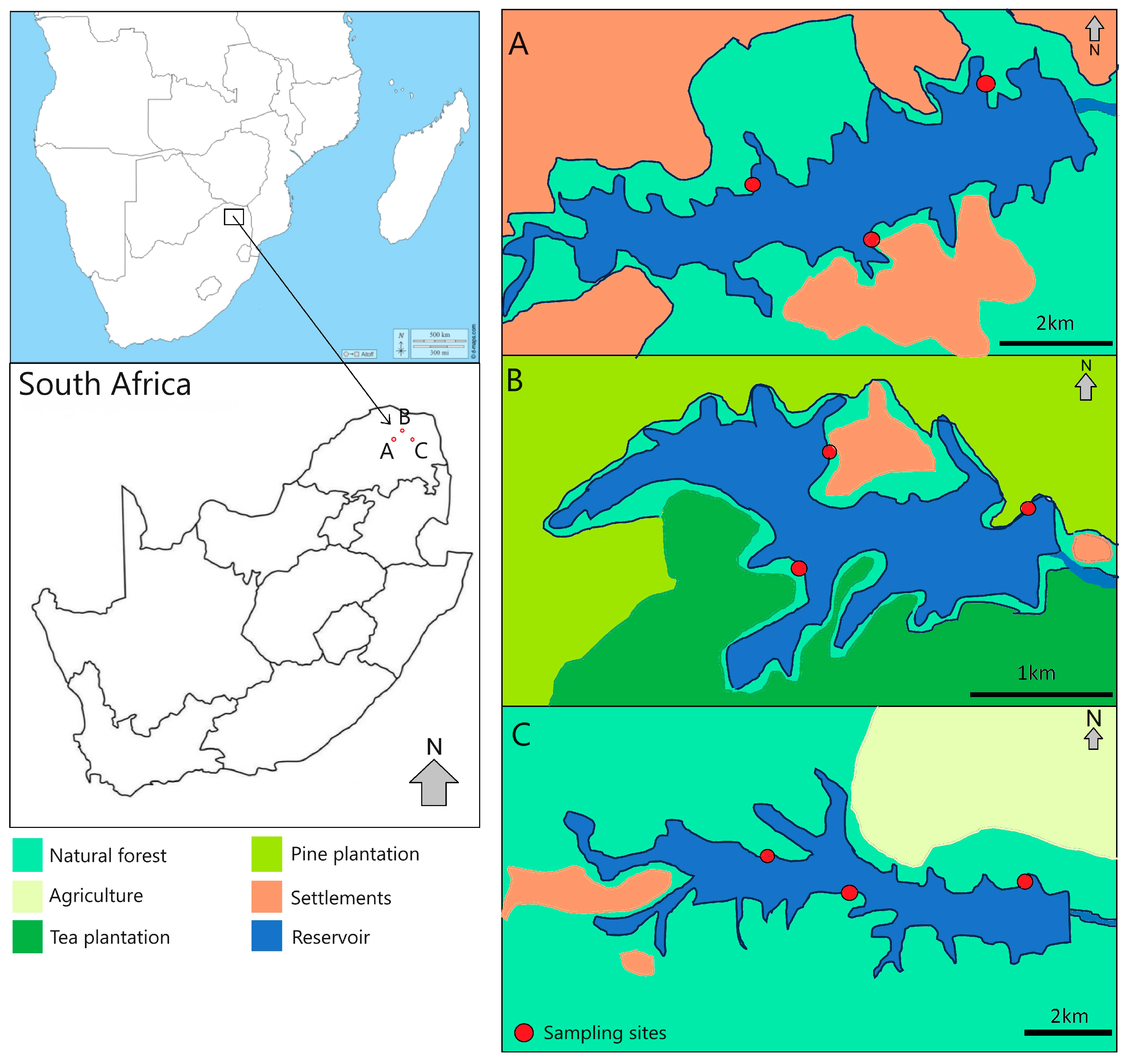
2.2. Study Area
2.3. Determining Environmental Variables
2.3.1. Physicochemical Parameters of Water
2.3.2. Sediment Chemistry Variables
2.4. Littoral Macroinvertebrates Sampling
2.5. Littoral Fish Sampling
2.6. Data Analyses
3. Results
3.1. Environmental Variables (Water and Sediment Chemisty)
3.2. Macroinvertebrate Communities
3.3. Fish Communities
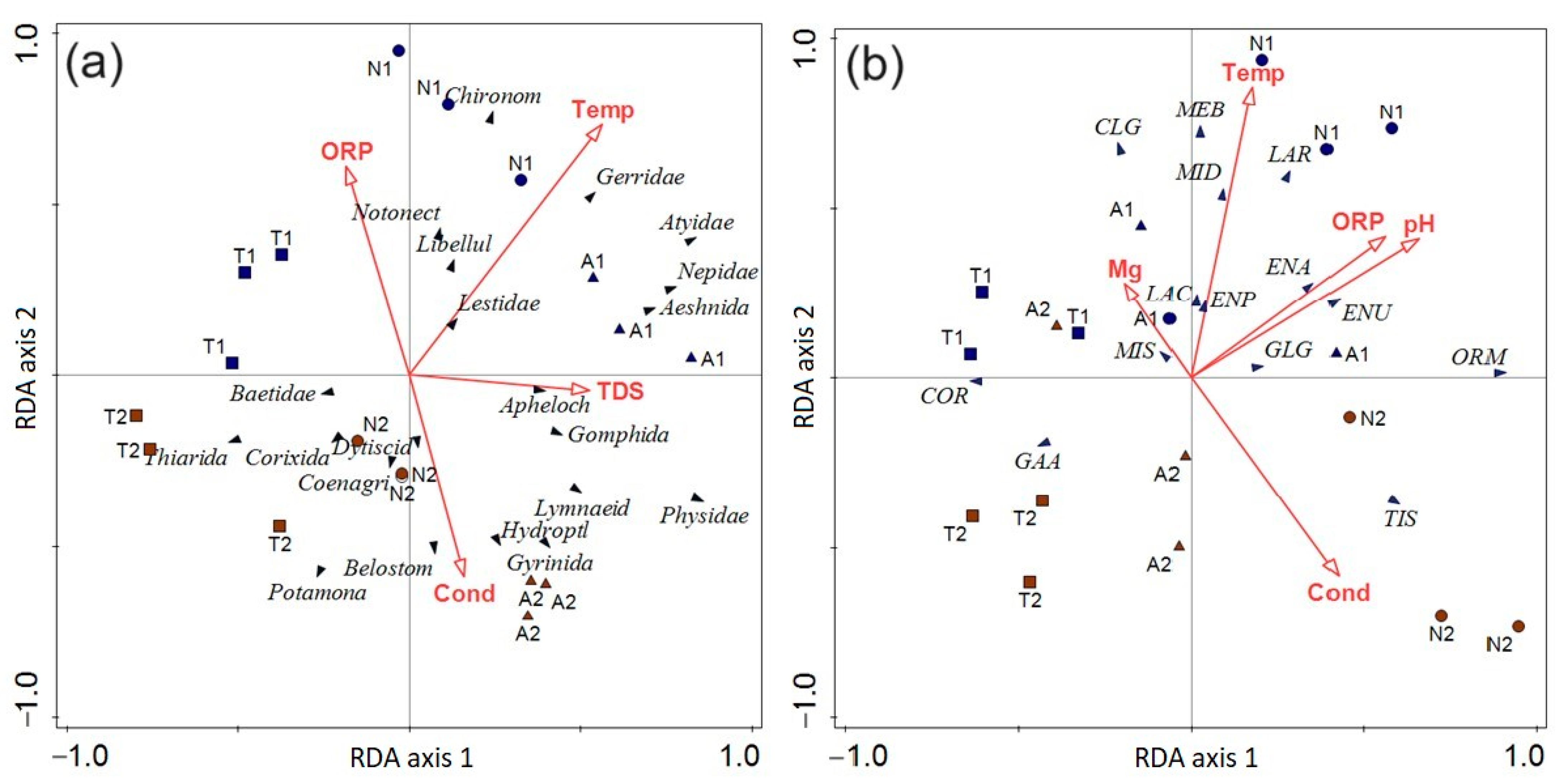
3.4. The Influence of Environmental Parameters on Macroinvertebrate Communities
3.5. The Influence of Environmental Parameters on Fish Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kushawaha, J.; Borra, S.; Kushawaha, A.K.; Singh, G.; Singh, P. Climate change and its impact on natural resources. In Water Conservation in the Era of Global Climate Change; Elsevier Science: Amsterdam, The Netherlands, 2021; pp. 333–346. [Google Scholar]
- Dotaniya, M.L.; Meena, V.D.; Saha, J.K.; Dotaniya, C.K.; Mahmoud, A.E.D.; Meena, B.L.; Meena, M.D.; Sanwal, R.C.; Meena, R.S.; Doutaniya, R.K.; et al. Reuse of poor-quality water for sustainable crop production in the changing scenario of climate. Environ. Dev. Sustain. 2023, 25, 7345–7376. [Google Scholar] [CrossRef] [PubMed]
- Wassie, S.B. Natural resource degradation tendencies in Ethiopia: A review. Environ. Syst. Res. 2020, 9, 1–29. [Google Scholar] [CrossRef]
- Bănăduc, D.; Simić, V.; Cianfaglione, K.; Barinova, S.; Afanasyev, S.; Öktener, A.; McCall, G.; Simić, S.; Curtean-Bănăduc, A. Freshwater as a sustainable resource and generator of secondary resources in the 21st century: Stressors, threats, risks, management and protection strategies, and conservation approaches. Int. J. Environ. Res. Public Health 2022, 19, 16570. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Sinha, D.; Chaudhary, V.; Kar, S.; Singh, A. Water pollution of wetlands: A global threat to inland, wetland, and aquatic phytodiversity. In Handbook of Research on Monitoring and Evaluating the Ecological Health of Wetlands; IGI Global: Hershey, PA, USA, 2022; pp. 27–50. [Google Scholar]
- Jamion, N.A.; Lee, K.E.; Mokhtar, M.; Goh, T.L.; Simon, N.; Goh, C.T.; Bhat, I.U.H. The integration of nature values and services in the nature-based solution assessment framework of constructed wetlands for carbon–water nexus in carbon sequestration and water security. Environ. Geochem. Health 2023, 45, 1201–1230. [Google Scholar] [CrossRef] [PubMed]
- Parris, K. Impact of agriculture on water pollution in OECD countries: Recent trends and future prospects. Int. J. Water Resour. Dev. 2011, 27, 33–52. [Google Scholar] [CrossRef]
- Dalu, T.; Dlamini, P.; Wasserman, R.J.; Mokgoebo, M.J.; Mutshekwa, T.; Dondofema, F.; Cuthbert, R.N. Effects of environmental variables on littoral macroinvertebrate community assemblages in subtropical reservoirs. Chem. Ecol. 2021, 37, 419–436. [Google Scholar] [CrossRef]
- Hojjati-Najafabadi, A.; Mansoorianfar, M.; Liang, T.; Shahin, K.; Karimi-Maleh, H. A review on magnetic sensors for monitoring of hazardous pollutants in water resources. Sci. Total Environ. 2022, 824, 153844. [Google Scholar] [CrossRef] [PubMed]
- Dalu, T.; Wasserman, R.J.; Tonkin, J.D.; Mwedzi, T.; Magoro, M.L.; Weyl, O.L.F. Water or sediment? Partitioning the role of water column and sediment chemistry as drivers of macroinvertebrate communities in an austral South African stream. Sci. Total Environ. 2017, 607, 317–325. [Google Scholar] [CrossRef]
- Dalu, T.; Chauke, R. Assessing macroinvertebrate communities in relation to environmental variables: The case of Sambandou wetlands, Vhembe Biosphere Reserve. Appl. Water Sci. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Massawe, P.I.; Mvena, A.; Nyoki, D.; Chambile, E.L. Effects of anthropogenic activities on availability of clean and safe water: A case of Uluguru forest catchment areas of Morogoro, Tanzania. South. Asian J. Dev. Res. 2019, 1, 114–123. [Google Scholar]
- Polasky, S.; Nelson, E.; Pennington, D.; Johnson, K.A. The impact of land-use change on ecosystem services, biodiversity and returns to landowners: A case study in the state of Minnesota. Environ. Resour. Econ. 2011, 48, 219–242. [Google Scholar] [CrossRef]
- Ayivor, J.S.; Gordon, C. Impact of land use on river systems in Ghana. West. Afr. J. Appl. Ecol. 2012, 20, 83–95. [Google Scholar]
- Hughes, R.M.; Zeigler, M.; Stringer, S.; Linam, G.W.; Flotemersch, J.; Jessup, B.; Joseph, S.; Jacobi, G.; Guevara, L.; Cook, R.; et al. Biological assessment of western USA sandy bottom rivers based on modeling historical and current fish and macroinvertebrate data. River Res. Appl. 2022, 38, 639–656. [Google Scholar] [CrossRef] [PubMed]
- Feio, M.J.; Hughes, R.M.; Serra, S.R.; Nichols, S.J.; Kefford, B.J.; Lintermans, M.; Robinson, W.; Odume, O.N.; Callisto, M.; Macedo, D.R.; et al. Fish and macroinvertebrate assemblages reveal extensive degradation of the world’s rivers. Glob. Change Biol. 2023, 29, 355–374. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.S.B.; Hasan, N.A.; Mondol, M.M.R.; Alam, M.M.; Haque, M.M. Decline in fish species diversity due to climatic and anthropogenic factors in Hakaluki Haor, an ecologically critical wetland in northeast Bangladesh. Heliyon 2021, 7, e05861. [Google Scholar] [CrossRef]
- Sudarso, J.; Suryono, T.; Yoga, G.P.; Samir, O.; Imroatushshoolikhah Ibrahim, A. The impact of anthropogenic activities on benthic macroinvertebrates community in the Ranggeh River. J. Ecol. Eng. 2021, 22, 179–190. [Google Scholar] [CrossRef]
- Gunkel, G.; Michels, U.; Scheideler, M. Water lice and other macroinvertebrates in drinking water pipes: Diversity, abundance and health risk. Water 2021, 13, 276. [Google Scholar] [CrossRef]
- Jacks, F.; Milošević, D.; Watson, V.; Beazley, K.F.; Medeiros, A.S. Bioassessment of the ecological integrity of freshwater ecosystems using aquatic macroinvertebrates: The case of Sable Island National Park Reserve, Canada. Environ. Monit. Assess. 2021, 193, 1–16. [Google Scholar] [CrossRef]
- Rasifudi, L.; Addo-Bediako, A.; Bal, K.; Swemmer, T.M. Distribution of benthic macroinvertebrates in the Selati River of the Olifants River system, South Africa. Afr. Entomol. 2018, 26, 398–406. [Google Scholar] [CrossRef]
- Passy, S.I.; Bode, R.W.; Carlson, D.M.; Novak, M.A. Comparative environmental assessment in the studies of benthic diatom, macroinvertebrate, and fish communities. Int. Rev. Hydrobiol. 2004, 89, 121–138. [Google Scholar] [CrossRef]
- Dalu, T.; Clegg, B.; Nhiwatiwa, T. Macroinvertebrate communities associated with littoral zone habitats and the influence of environmental factors in Malilangwe Reservoir, Zimbabwe. Knowl. Manag. Aquat. Ecosyst. 2012, 406, 6. [Google Scholar] [CrossRef]
- Ghaleno, O.R. Potential ecological risk assessment of heavy metals in sediments of water reservoir case study: Chah Nimeh of Sistan. Proc. Int. Acad. Ecol. Environ. Sci. 2015, 5, 89. [Google Scholar]
- Moura, D.S.; Neto, I.E.L.; Clemente, A.; Oliveira, S.; Pestana, C.J.; de Melo, M.A.; Capelo-Neto, J. Modeling phosphorus exchange between bottom sediment and water in tropical semiarid reservoirs. Chemosphere 2020, 246, 125686. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, N.C.L.; Garcia-Berthou, E.; Dias, J.D.; Lopes, T.M.; Affonso, I.D.P.; Severi, W.; Gomes, L.C.; Agostinho, A.A. Cumulative ecological effects of a Neotropical reservoir cascade across multiple assemblages. Hydrobiologia 2018, 819, 77–91. [Google Scholar] [CrossRef]
- Liu, S.; Xie, G.; Wang, L.; Cottenie, K.; Liu, D.; Wang, B. Different roles of environmental variables and spatial factors in structuring stream benthic diatom and macroinvertebrate in Yangtze River Delta, China. Ecol. Indic. 2016, 61, 602–611. [Google Scholar] [CrossRef]
- Glibert, P.M.; Fullerton, D.; Burkholder, J.M.; Cornwell, J.C.; Kana, T.M. Ecological stoichiometry, biogeochemical cycling, invasive species, and aquatic food webs: San Francisco Estuary and comparative systems. Rev. Fish. Sci. 2011, 19, 358–417. [Google Scholar] [CrossRef]
- Rettig, J.E.; Smith, G.R. Relative strength of top-down effects of an invasive fish and bottom-up effects of nutrient addition in a simple aquatic food web. Environ. Sci. Pollut. Res. 2021, 28, 5845–5853. [Google Scholar] [CrossRef]
- Kang, M.; Tian, Y.; Peng, S.; Wang, M. Effect of dissolved oxygen and nutrient levels on heavy metal contents and fractions in river surface sediments. Sci. Total Environ. 2019, 648, 861–870. [Google Scholar] [CrossRef]
- Singh, J.; Kalamdhad, A.S. Effects of heavy metals on soil, plants, human health and aquatic life. Int. J. Res. Chem. Environ. 2011, 1, 15–21. [Google Scholar]
- Barnhoorn, I.E.; Van Dyk, J.C.; Pieterse, G.M.; Bornman, M.S. Intersex in feral indigenous freshwater Oreochromis mossambicus, from various parts in the Luvuvhu River, Limpopo Province, South Africa. Ecotoxicol. Environ. Saf. 2010, 73, 1537–1542. [Google Scholar] [CrossRef]
- Odiyo, J.O.; Makungo, R.; Nkuna, T.R. Long-term changes and variability in rainfall and streamflow in Luvuvhu River Catchment, South Africa. South. Afr. J. Sci. 2015, 111, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ramulifho, P.; Ndou, E.; Thifhulufhelwi, R.; Dalu, T. Challenges to implementing an environmental flow regime in the Luvuvhu River Catchment, South Africa. Int. J. Environ. Res. Public Health 2019, 16, 3694. [Google Scholar] [CrossRef] [PubMed]
- Patience, M.T.; Elumalai, V.; Rajmohan, N.; Li, P. Occurrence and distribution of nutrients and trace metals in groundwater in an intensively irrigated region, Luvuvhu catchment, South Africa. Environ. Earth Sci. 2021, 80, 1–15. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Allen, K.; Corre, M.D.; Tjoa, A.; Veldkamp, E. Soil nitrogen-cycling responses to conversion of lowland forests to oil palm and rubber plantations in Sumatra, Indonesia. PLoS ONE 2015, 10, e0133325. [Google Scholar] [CrossRef]
- Dalu, T.; Cuthbert, R.N.; Taylor, J.C.; Magoro, M.L.; Weyl, O.L.; Froneman, P.W.; Wasserman, R.J. Benthic diatom-based indices and isotopic biomonitoring of nitrogen pollution in a warm temperate Austral river system. Sci. Total Environ. 2020, 748, 142452. [Google Scholar] [CrossRef]
- Dalu, T.; Murudi, T.; Dondofema, F.; Wasserman, R.J.; Chari, L.D.; Murungweni, F.M.; Cuthbert, R.N. Balloon milkweed Gomphocarpus physocarpus distribution and drivers in an internationally protected wetland. BioInvasions Rec. 2020, 9, 627–641. [Google Scholar] [CrossRef]
- Fry, C. Field Guide to the Freshwater Macroinvertebrates of South Africa; Jacana Media: Auckland Park, South Africa, 2021. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and Canodraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 5); Cacono: Ithaca, NY, USA, 2012. [Google Scholar]
- Purcell, A.H.; Bressler, D.W.; Paul, M.J.; Barbour, M.T.; Rankin, E.T.; Carter, J.L.; Resh, V.H. Assessment tools for urban catchments: Developing biological indicators based on benthic macroinvertebrates 1. J. Am. Water Resour. Assoc. 2009, 45, 306–319. [Google Scholar] [CrossRef]
- Halliwell, D.B.; Langdon, R.W.; Daniels, R.A.; Kurtenbach, J.P.; Jacobson, R.A. Classification of freshwater fish species of the northeastern United States for use in the development of indices of biological integrity, with regional applications. In Assessing the Sustainability and Biological Integrity of Water Resources Using Fish Communities; CRC Press: Boca Raton, FL, USA, 2020; pp. 301–337. [Google Scholar]
- Kolpin, D.W.; Blazer, V.S.; Gray, J.L.; Focazio, M.J.; Young, J.A.; Alvarez, D.A.; Iwanowicz, L.R.; Foreman, W.T.; Furlong, E.T.; Speiran, G.K.; et al. Chemical contaminants in water and sediment near fish nesting sites in the Potomac River basin: Determining potential exposures to smallmouth bass (Micropterus dolomieu). Sci. Total Environ. 2013, 443, 700–716. [Google Scholar] [CrossRef]
- Mofu, L.; Dalu, T.; Wasserman, R.J.; Woodford, D.J.; Khosa, D.; Weyl, O.L.F. Seasonal variation and drivers of zooplankton, macroinvertebrate and littoral fish communities from irrigation ponds in a semi-arid region in the Eastern Cape (South Africa). Afr. J. Aquat. Sci. 2021, 46, 452–463. [Google Scholar] [CrossRef]
- Arkia, S.; Siahkalroudi, S.Y.; Kheradpir, N. Chironomidae (Insecta: Diptera) biodiversity at generic level in Lar river, Tehran province with introducing two new genera for Iranian fauna. J. Wildl. Biodivers. 2019, 3, 31–39. [Google Scholar]
- Al-Shami, S.A.; Rawi, C.S.M.; HassanAhmad, A.; Nor, S.A.M. Distribution of Chironomidae (Insecta: Diptera) in polluted rivers of the Juru River Basin, Penang, Malaysia. J. Environ. Sci. 2010, 22, 1718–1727. [Google Scholar] [CrossRef] [PubMed]
- Trottier, G.; Embke, H.; Turgeon, K.; Solomon, C.; Nozais, C.; Gregory-Eaves, I. Macroinvertebrate abundance is lower in temperate reservoirs with higher winter drawdown. Hydrobiologia 2019, 834, 199–211. [Google Scholar] [CrossRef]
- Tickner, D.; Opperman, J.J.; Abell, R.; Acreman, M.; Arthington, A.H.; Bunn, S.E.; Cooke, S.J.; Dalton, J.; Darwall, W.; Edwards, G.; et al. Bending the curve of global freshwater biodiversity loss: An emergency recovery plan. BioScience 2020, 70, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Sanganyado, E.; Gwenzi, W. Antibiotic resistance in drinking water systems: Occurrence, removal, and human health risks. Sci. Total Environ. 2019, 669, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Häder, D.P.; Banaszak, A.T.; Villafañe, V.E.; Narvarte, M.A.; González, R.A.; Helbling, E.W. Anthropogenic pollution of aquatic ecosystems: Emerging problems with global implications. Sci. Total Environ. 2020, 713, 136586. [Google Scholar] [CrossRef]
- Huang, W.; Song, B.; Liang, J.; Niu, Q.; Zeng, G.; Shen, M.; Deng, J.; Luo, Y.; Wen, X.; Zhang, Y. Microplastics and associated contaminants in the aquatic environment: A review on their ecotoxicological effects, trophic transfer, and potential impacts to human health. J. Hazard. Mater. 2021, 405, 124187. [Google Scholar] [CrossRef]
- Odume, O.N.; Muller, W.J.; Arimoro, F.O.; Palmer, C.G. The impact of water quality deterioration on macroinvertebrate communities in the Swartkops River, South Africa: A multimetric approach. Afr. J. Aquat. Sci. 2012, 37, 191–200. [Google Scholar] [CrossRef]
- Gething, K.J. Physicochemical drivers of managed river and agricultural drainage channel macroinvertebrate communities. River Res. Appl. 2021, 37, 675–680. [Google Scholar] [CrossRef]
- Sylvain, F.É.; Holland, A.; Audet-Gilbert, É.; Luis Val, A.; Derome, N. Amazon fish bacterial communities show structural convergence along widespread hydrochemical gradients. Mol. Ecol. 2019, 28, 3612–3626. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Guo, L.; Deng, Z.; Wang, D.; Liu, L. Assessment of heavy metal pollution and water quality characteristics of the reservoir control reaches in the middle Han River, China. Sci. Total Environ. 2021, 799, 149472. [Google Scholar] [CrossRef] [PubMed]
- Tamiru, S.M. Impacts of human activities on selected physico-chemical parameters and macroinvertebretes of Lake Tana, Northwestern Ethiopia. Ethiop. Renaiss. J. Social. Sci. Humanit. 2021, 8, 146–159. [Google Scholar]

{kind=link}
{kind=link}
| Variables | Unit | Seasons | Reservoirs | ||
|---|---|---|---|---|---|
| Albasini | Nandoni | Thathe | |||
| Temperature | °C | Cool-dry | 22.4 ± 10.3 | 20.8 ± 9.5 | 18.7 ± 8.5 |
| Hot-wet | 27.6 ± 12.9 | 28.4 ± 13.3 | 25.5 ± 11.9 | ||
| pH | Cool-dry | 7.5 ± 2.9 | 8.3 ± 3.3 | 7.5 ± 2.9 | |
| Hot-wet | 8.3 ± 3.3 | 8.4 ± 3.3 | 8.1 ± 3.2 | ||
| Conductivity | µS cm−1 | Cool-dry | 128.1 ± 148.2 | 421.2 ± 209.8 | 316.0 ± 157.1 |
| Hot-wet | 205.7 ± 102.3 | 169.4 ± 85.0 | 78.3 ± 39.6 | ||
| TDS | mg L–1 | Cool-dry | 106.8 ± 52.5 | 132.5 ± 65.4 | 113.3 ± 55.8 |
| Hot-wet | 142.0 ± 70.2 | 123.2 ± 60.8 | 58.1 ± 29.7 | ||
| Salinity | ppm | Cool-dry | 481.5 ± 331.0 | 1902.4 ± 1417.6 | 2246.1 ± 1749.6 |
| Hot-wet | 462.6 ± 232.6 | 686.7 ± 501.6 | 688.3 ± 504.9 | ||
| Resistivity | ohm | Cool-dry | 1541.5 ± 950.2 | 778.0 ± 743.4 | 720.3 ± 787.9 |
| Hot-wet | 1665.0 ± 842.3 | 1397.0 ± 835.5 | 1304.6 ± 810.1 | ||
| ORP | mV | Cool-dry | 43.3 ± 20.8 | 50.1 ± 24.3 | 43.5 ± 20.9 |
| Hot-wet | 41.8 ± 20.1 | 54.6 ± 26.5 | 46.5 ± 22.4 | ||
| Variables | Reservoirs | Seasons | Reservoirs × Seasons | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| Water chemistry variables | ||||||
| Temperature | 145.270 | <0.001 | 1862.535 | <0.001 | 22.449 | <0.001 |
| pH | 9.631 | 0.003 | 21.715 | 0.001 | 3.915 | 0.049 |
| Conductivity | 148.528 | <0.001 | 1732.485 | <0.001 | 119.397 | <0.001 |
| TDS | 127.415 | <0.001 | 16.652 | 0.002 | 199.144 | <0.001 |
| Salinity | 2.100 | 0.165 | 4.825 | 0.048 | 1.212 | 0.332 |
| Resistivity | 1.491 | 0.264 | 2.115 | 0.172 | 0.276 | 0.764 |
| ORP | 48.958 | <0.001 | 5.620 | 0.035 | 4.543 | 0.034 |
| Sediment chemistry variables | ||||||
| pH | 0.76 | 0.927 | 0.757 | 0.401 | 4.882 | 0.028 |
| P | 2.655 | 0.111 | 0.207 | 0.657 | 2.038 | 0.176 |
| NH4+ | 0.681 | 0.525 | 0.531 | 0.480 | 0.567 | 0.582 |
| K | 1.690 | 0.226 | 1.625 | 0.227 | 0.482 | 0.629 |
| Ca | 0.312 | 0.738 | 0.285 | 0.603 | 0.303 | 0.744 |
| Mg | 1.242 | 0.323 | 0.571 | 0.465 | 1.841 | 0.201 |
| Cu | 0.403 | 0.677 | 0.001 | 1.000 | 0.244 | 0.787 |
| Zn | 0.131 | 0.878 | 0.001 | 0.971 | 3.154 | 0.079 |
| Mn | 0.582 | 0.574 | 0.096 | 0.763 | 2.639 | 0.112 |
| B | 1.165 | 0.317 | 1.559 | 0.236 | 0.324 | 0.730 |
| Fe | 0.087 | 0.917 | 0.048 | 0.830 | 1.024 | 0.388 |
| C | 0.956 | 0.412 | 3.119 | 0.103 | 0.719 | 0.507 |
| S | 1.165 | 0.345 | 0.235 | 0.637 | 5.459 | 0.021 |
| Macroinvertebrate diversity metrics | ||||||
| Evenness | 49.408 | <0.001 | 1.353 | 0.267 | 4.228 | 0.041 |
| Margalef’s diversity | 40.745 | <0.001 | 0.267 | 0.615 | 18.737 | <0.001 |
| Taxa richness | 52.125 | <0.001 | 0.042 | 0.842 | 7.292 | 0.008 |
| Shannon–Wiener diversity | 15.887 | <0.001 | 3.650 | 0.080 | 3.399 | 0.068 |
| Simpson’s diversity | 7.549 | 0.008 | 5.985 | 0.031 | 2.046 | 0.172 |
| Total abundance | 51.128 | <0.001 | 0.139 | 0.716 | 4.794 | 0.029 |
| Fish diversity metrics | ||||||
| Evenness | 29.031 | <0.001 | 0.008 | 0.928 | 10.045 | 0.003 |
| Margalef’s diversity | 0.309 | 0.740 | 8.112 | 0.015 | 4.409 | 0.037 |
| Taxa richness | 3.040 | 0.085 | 16.000 | 0.002 | 2.560 | 0.119 |
| Shannon–Wiener diversity | 1.487 | 0.265 | 20.071 | 0.001 | 9.163 | 0.004 |
| Simpson’s diversity | 9.824 | 0.003 | 17.173 | 0.001 | 15.023 | 0.001 |
| Total abundance | 15.031 | 0.001 | 6.870 | 0.022 | 0.046 | 0.956 |
| Order | Taxa | Cool-Dry | Hot-Wet | ||||
|---|---|---|---|---|---|---|---|
| Albasini | Nandoni | Thathe | Albasini | Nandoni | Thathe | ||
| Mollusca | Thiaridae | 26.9 | 15.7 | 5.7 | |||
| Mollusca | Lymnaeidae | 3.6 | 2.3 | ||||
| Mollusca | Physidae | 57.1 | 15.7 | 37.9 | 5.6 | ||
| Crustacea | Atyidae | 3.6 | 5.9 | 23.5 | 21.3 | 2.9 | |
| Crustacea | Potamonautidae | 3.6 | 19.2 | 5.9 | 8.6 | ||
| Odonata | Aeshnidae | 2.4 | 5.9 | 3.0 | 6.7 | ||
| Odonata | Lestidae | 3.9 | 4.5 | ||||
| Odonata | Libellulidae | 3.0 | 7.8 | 0.8 | 10.1 | 14.3 | |
| Odonata | Coenagrionidae | 1.8 | 15.4 | 3.9 | 3.8 | 8.6 | |
| Odonata | Gomphidae | 2.4 | 15.4 | 2.0 | 7.6 | 1.1 | |
| Coleoptera | Dytiscidae | 4.8 | 19.2 | 7.8 | 5.6 | ||
| Coleoptera | Gyrinidae | 6.0 | 0.8 | ||||
| Coleoptera | Hydroptilidae | 3.0 | |||||
| Hemiptera | Nepidae | 3.6 | 5.9 | 6.1 | 10.1 | ||
| Hemiptera | Notonectidae | 9.8 | 9.0 | ||||
| Hemiptera | Gerridae | 3.0 | 7.9 | ||||
| Hemiptera | Belostomatidae | 4.2 | 2.0 | 8.6 | |||
| Hemiptera | Aphelocheiridae | 3.9 | 2.3 | ||||
| Hemiptera | Corixidae | 3.9 | 3.9 | ||||
| Ephemeroptera | Baetidae | 1.2 | 17.1 | ||||
| Diptera | Chironomidae | 9.1 | 18.0 | 34.3 | |||
| Diversity metrics | |||||||
| Evenness | 0.4 | 0.9 | 0.9 | 0.6 | 0.9 | 0.9 | |
| Margalef’s diversity | 2.8 | 3.2 | 1.1 | 2.4 | 2.5 | 2.1 | |
| Taxa richness | 14 | 6 | 15 | 12 | 11 | 8 | |
| Shannon–Wiener diversity | 1.7 | 2.2 | 1.0 | 1.8 | 2.1 | 1.6 | |
| Simpson’s diversity | 0.7 | 0.9 | 0.6 | 0.8 | 0.9 | 0.8 | |
| Species | Cool-Dry | Hot-Wet | ||||
|---|---|---|---|---|---|---|
| Albasini | Nandoni | Thathe | Albasini | Nandoni | Thathe | |
| Clarias gariepinus (Burchell, 1822) | 13.3 | 12.2 | 11.9 | 16.1 | ||
| Micropterus dolomieu (Lacépède, 1802) | 7.9 | 39.0 | 18.6 | |||
| Micropterus salmoides (Lacépède, 1802) | 23.7 | 5.9 | 6.8 | 16.1 | ||
| Oreochromis mossambicus (Peters, 1852) | 15.8 | 52.9 | 26.7 | 7.3 | 23.7 | 6.5 |
| Coptodon rendalli (Boulenger, 1897) | 13.2 | 5.9 | 26.7 | 7.3 | 5.1 | 9.7 |
| Tilapia sparrmanii (Smith. 1840) | 18.5 | 14.7 | 4.9 | 5.1 | 6.5 | |
| Enteromius afrohamiltoni (Crass, 1960) | 10.5 | 3.4 | ||||
| Gambusia affinis (Baird and Girard, 1853) | 33.3 | 6.5 | ||||
| Enteromius paludinosus (Peters, 1852) | 14.7 | 12.2 | 13.6 | 16.1 | ||
| Enteromius unitaeniatus (Günther, 1866) | 13.2 | 5.9 | 8.5 | 6.5 | ||
| Labeo rosae (Steindachner, 1894) | 3.4 | |||||
| Mesobola brevianalis (Boulenger, 1908) | 7.3 | 16.1 | ||||
| Glossogobius guiris (Hamilton, 1822) | 4.9 | |||||
| Labeo cylindricus (Peters, 1852) | 4.9 | |||||
| Diversity metrics | ||||||
| Evenness | 0.8 | 0.5 | 0.5 | 0.7 | 0.7 | 0.8 |
| Margalef’s diversity | 1.9 | 1.3 | 1.3 | 2.1 | 2.4 | 2.0 |
| Taxa richness | 7 | 6 | 3 | 9 | 10 | 9 |
| Shannon–Wiener diversity | 1.6 | 1.0 | 1.0 | 1.7 | 1.9 | 1.7 |
| Simpson’s diversity | 0.8 | 0.5 | 0.5 | 0.8 | 0.8 | 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munyai, L.F.; Liphadzi, T.; Mutshekwa, T.; Mutoti, M.I.; Mofu, L.; Murungweni, F.M. Water and Sediment Chemistry as Drivers of Macroinvertebrates and Fish Assemblages in Littoral Zones of Subtropical Reservoirs. Water 2024, 16, 42. https://doi.org/10.3390/w16010042
Munyai LF, Liphadzi T, Mutshekwa T, Mutoti MI, Mofu L, Murungweni FM. Water and Sediment Chemistry as Drivers of Macroinvertebrates and Fish Assemblages in Littoral Zones of Subtropical Reservoirs. Water. 2024; 16(1):42. https://doi.org/10.3390/w16010042
Chicago/Turabian StyleMunyai, Linton F., Thendo Liphadzi, Thendo Mutshekwa, Mulalo I. Mutoti, Lubabalo Mofu, and Florence M. Murungweni. 2024. "Water and Sediment Chemistry as Drivers of Macroinvertebrates and Fish Assemblages in Littoral Zones of Subtropical Reservoirs" Water 16, no. 1: 42. https://doi.org/10.3390/w16010042