


Chlorine Photolysis: A Step Forward in Inactivating Acanthamoeba and Their Endosymbiont Bacteria

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Isolation and FLA Inoculum
2.2. Disinfection Treatments
2.3. Determination of FLA and EB Inactivation
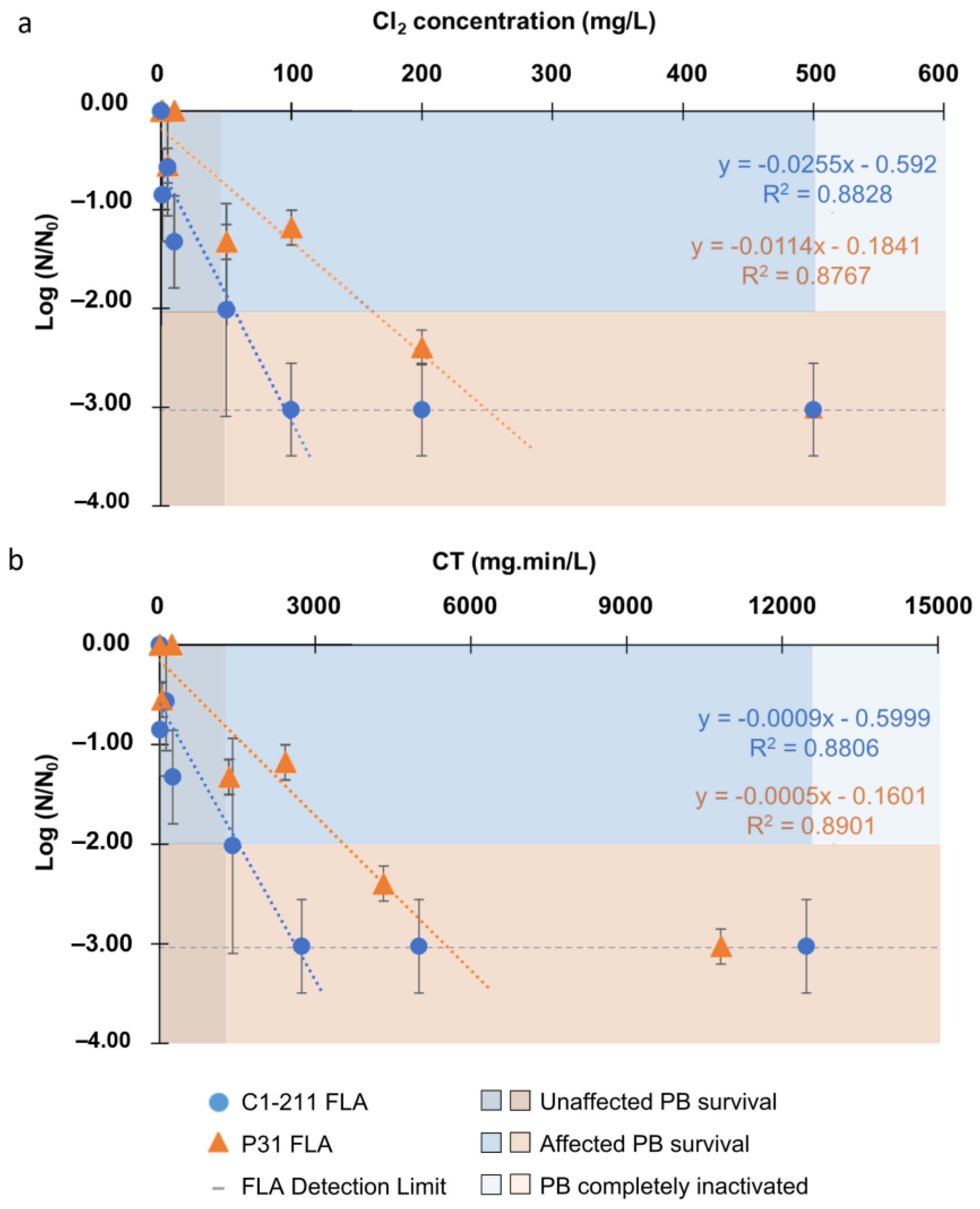
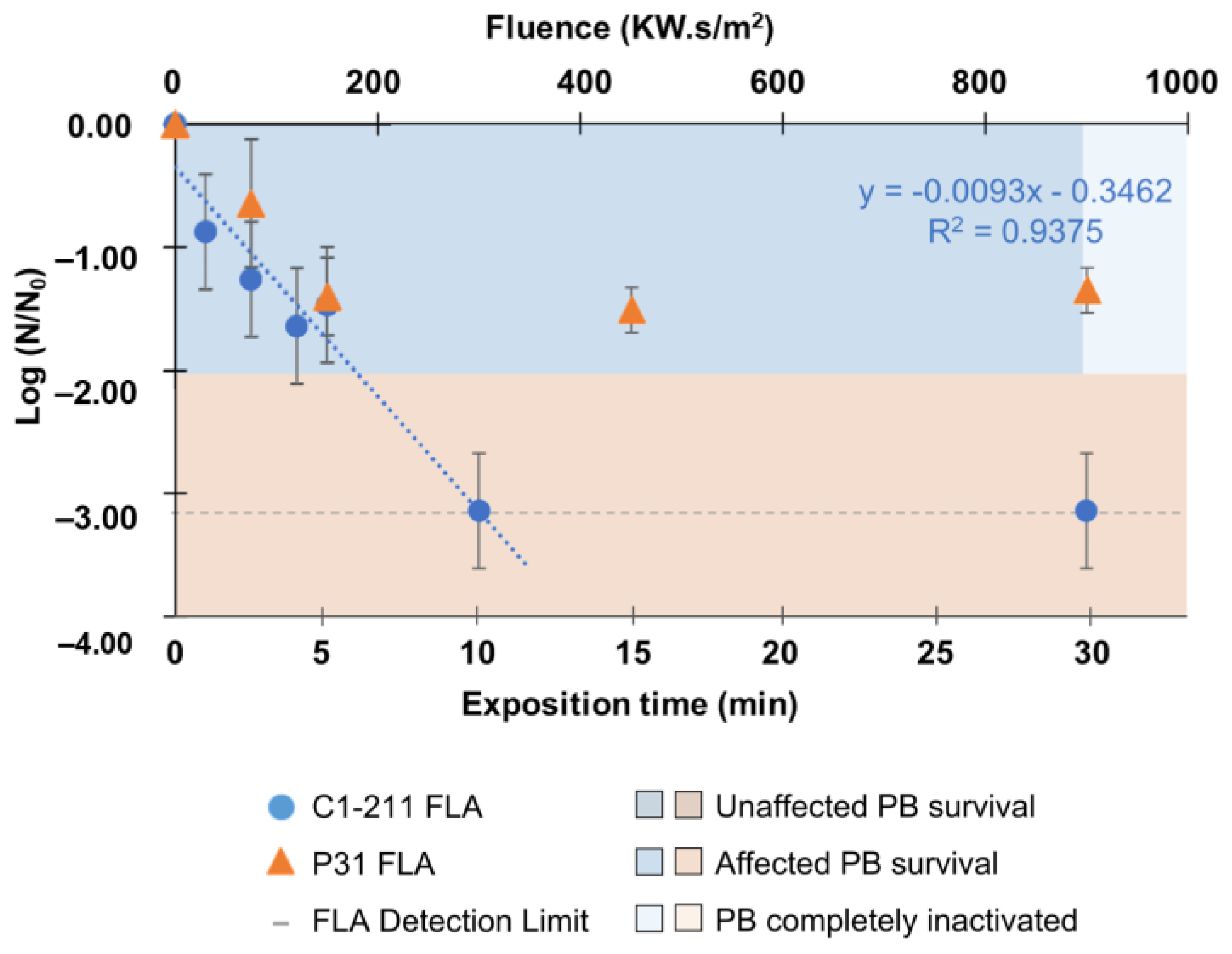
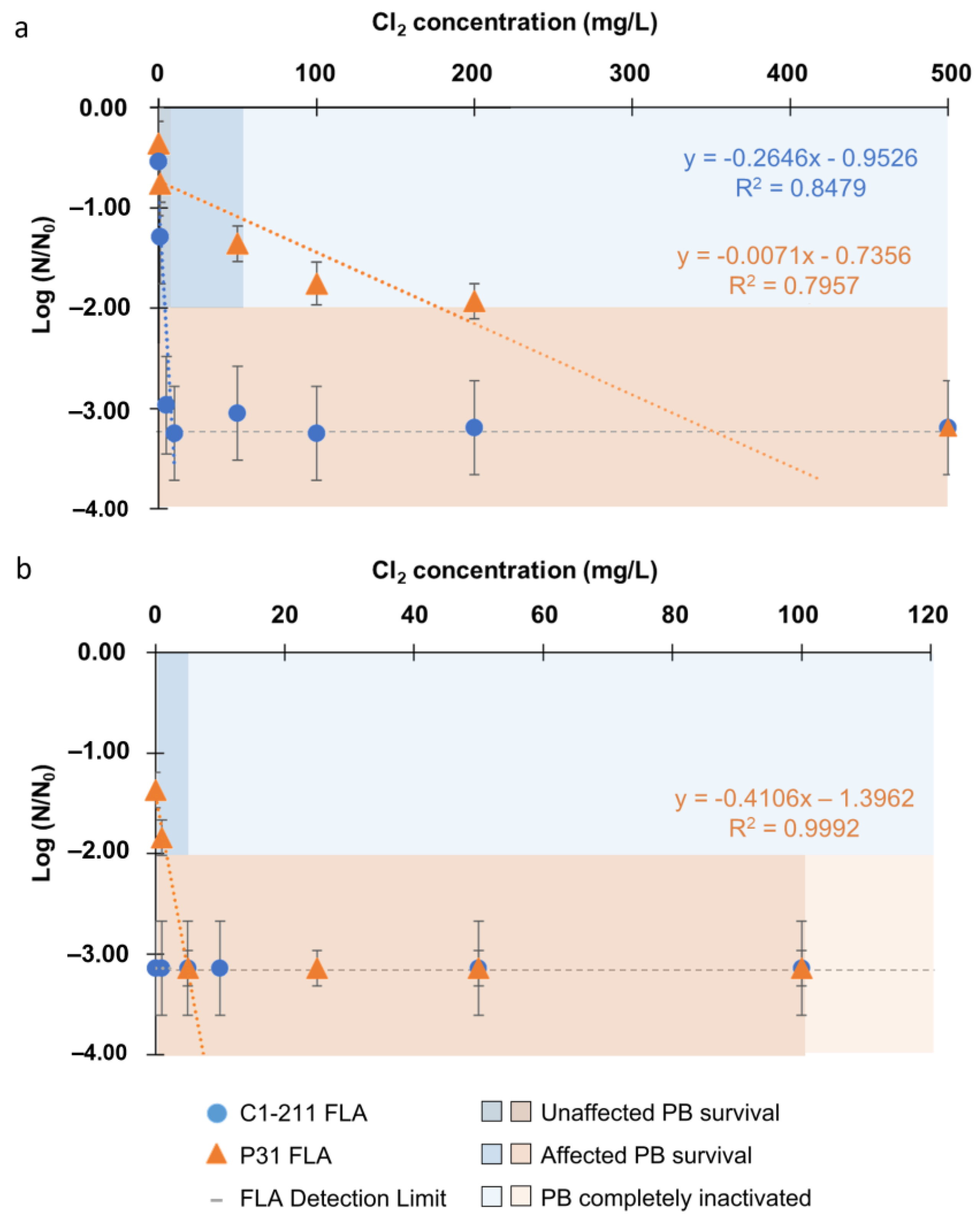
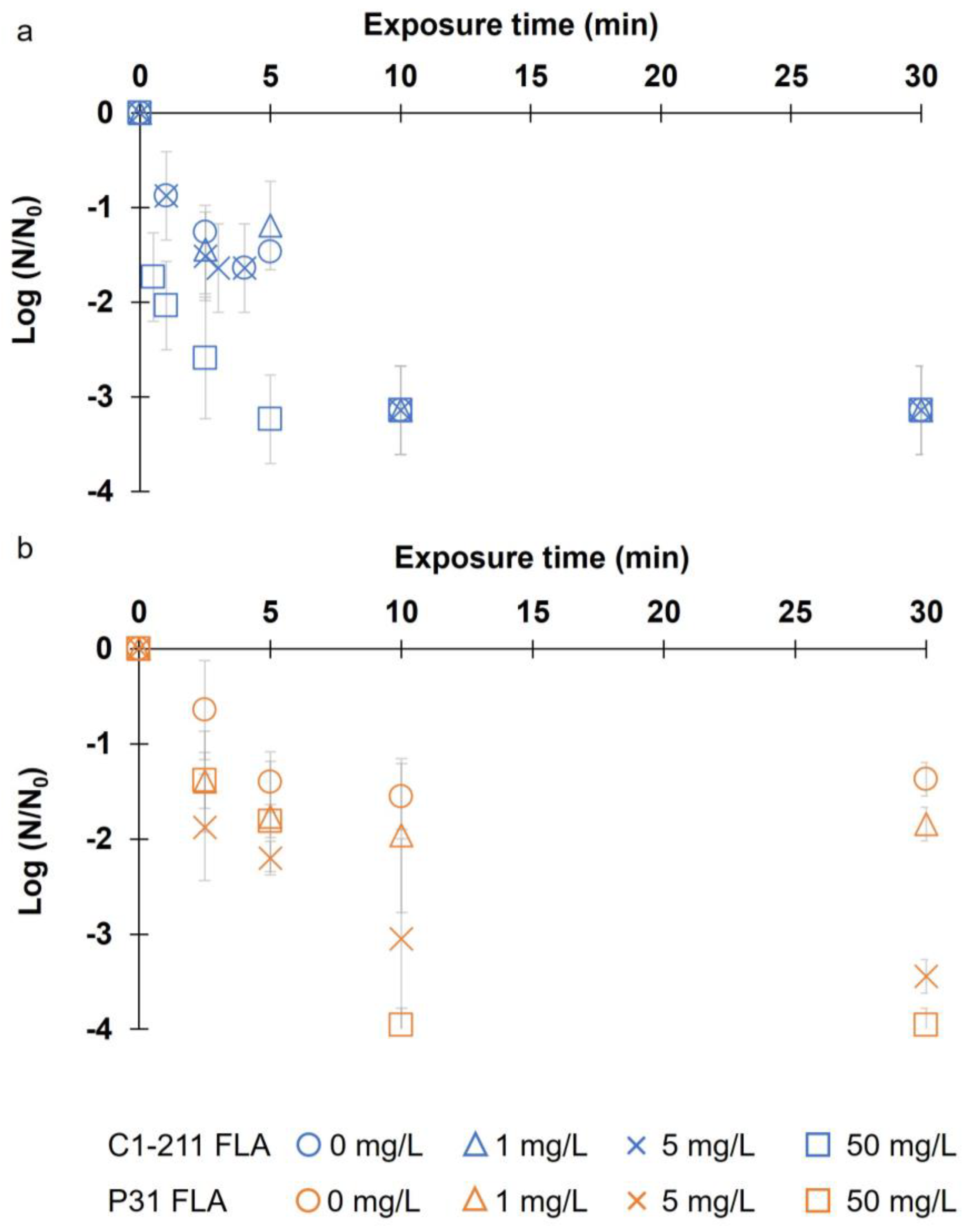
3. Results
3.1. Inactivation of Acanthamoeba by Conventional Treatments
3.2. Inactivation of Acanthamoeba by Chlorine Photolysis
3.3. Endosymbiont Bacteria Inactivation
4. Discussion
4.1. Acanthamoeba Inactivation by Conventional Treatments: Chlorine and Solar Radiation
4.2. Acanthamoeba Inactivation by Advanced Oxidation Process: Combination of Cl2 and SR
4.3. Inactivation of the Endosymbiont Bacteria Protected Inside Acanthamoeba
4.4. Effect of Water Sources: Acanthamoeba Isolated from Natural vs. Artificial Water Sources
5. Conclusions
- Conventional chlorine and solar radiation disinfection treatments required high disinfectant doses and were not effective against the endosymbiont bacteria protected by the amoebae studied.
- The combination of Cl2 and SR320–800nm was effective against the freshwater Acanthamoeba strain and its endosymbiont bacteria, reducing 10 times the required Cl2 dose. Additionally, the Cl2 and SR280–800nm combination was effective against both Acanthamoeba and their endosymbiont bacteria, reducing between 5 and 100 times the necessary Cl2 dose.
- The inactivation of EB requires the prior inactivation of Acanthamoeba cysts, and because of that, the disinfectant doses for efficient disinfection were up to 20 times greater than those used for cyst inactivation.
- The Acanthamoeba strain isolated from pool water (P31) and, consequently, the EB protected inside showed considerable higher resistance to the combination of Cl2 and SR280–800nm compared to the Acanthamoeba strain isolated from freshwater (C1-211), suggesting that higher disinfectant doses may be required to eliminate amoebae previously exposed to chlorine and solar radiation.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Guidelines for Drinking-Water Quality, Fourth ed.: Incorporating the First and Second Addenda; World Health Organization: Geneva, Switzerland, 2022; ISBN 978-92-4-004506-4. Available online: https://www.who.int/publications/i/item/9789240045064 (accessed on 28 December 2023).
- Pichel, N.; Vivar, M.; Fuentes, M. The problem of drinking water access: A review of disinfection technologies with an emphasis of disinfection technologies with an emphasis on solar treatment methods. Chemosphere 2019, 218, 1014–1030. [Google Scholar] [CrossRef]
- Benito, M.; Menacho, C.; Chueca, P.; Ormad, M.P.; Goñi, P. Seeking the reuse of effluents and sludge from conventional wastewater treatment plants: Analysis of the presence of intestinal protozoa and nematode eggs. J. Environ. Manag. 2020, 261, 110268. [Google Scholar] [CrossRef] [PubMed]
- Visvesvara, G.S.; Moura, H.; Schuster, F.L. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 2007, 50, 1–26. [Google Scholar] [CrossRef]
- Garcia, A.; Goñi, P.; Cieloszyk, J.; Fernandez, M.T.; Calvo-Beguería, L.; Rubio, E.; Fillat, M.F.; Peleato, M.L.; Clavel, A. Identification of Free-Living Amoebae and Amoeba-Associated Bacteria from Reservoirs and Water Treatment Plants by Molecular Techniques. Environ. Sci. Technol. 2013, 47, 3132–3140. [Google Scholar] [CrossRef] [PubMed]
- Greub, G.; Raoult, D. Microorganisms Resistant to Free-Living Amoebae. Clin. Microbiol. Rev. 2004, 17, 413–433. [Google Scholar] [CrossRef] [PubMed]
- Goñi, P.; Fernández, M.T.; Rubio, E. Identifying endosymbiont bacteria associated with free-living amoebae. Environ. Microbiol. 2014, 16, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Garajová, M.; Mrva, M.; Vaškovicová, N.; Martinka, M.; Melicherová, J.; Valigurová, A. Cellulose fibrils formation and organisation of cytoskeleton during encystment are essential for Acanthamoeba cyst wall architecture. Sci. Rep. 2019, 9, 4466. [Google Scholar] [CrossRef]
- Marín, I.; Goñi, P.; Lasheras, A.M.; Ormad, M.P. Efficiency of a Spanish wastewater treatment plant for removal potentially pathogens: Characterization of bacteria and protozoa along water and sludge treatment lines. Ecol. Eng. 2015, 74, 28–32. [Google Scholar] [CrossRef]
- Mosteo, R.; Ormad, M.P.; Goñi, P.; Rodríguez-Chueca, J.; García, A.; Clavel, A. Identification of pathogen bacteria and protozoa in treated urban wastewaters discharged in the Ebro River (Spain): Water reuse possibilities. Water Sci. Technol. 2013, 68, 575–583. [Google Scholar] [CrossRef]
- Gomes, T.S.; Vaccaro, L.; Magnet, A.; Izquierdo, F.; Ollero, D.; Martínez-Fernández, C.; Mayo, L.; Moran, M.; Pozuelo, M.J.; Fenoy, S.; et al. Presence and interaction of free-living amoebae and amoeba-resisting bacteria in water from Drinking Water Treatment Plants. Sci. Total Environ. 2020, 719, 137080. [Google Scholar] [CrossRef]
- Fernández, M.T. Molecular Characterization of Free-Living Amoebae and Identification of Other Parasites from the Water Network of Zaragoza District: Association with Other Microorganisms and Risks for Public Health. (Caracterización Molecular de Amebas de Vida Libre e Identificación de Otros Parásitos en Aguas de Red de la Provincia de Zaragoza: Asociación con Otros Microorganismos y Riesgos para la Salud Pública). Ph.D. Thesis, University of Zaragoza, Zaragoza, Spain, 2014. [Google Scholar]
- Nisar, M.A.; Ross, K.E.; Brown, M.H.; Bentham, R.; Hinds, J.; Whiley, H. Molecular screening and characterization of Legionella pneumophila associated free-living amoebae in domestic and hospital water systems. Water Res. 2022, 226, 119238. [Google Scholar] [CrossRef]
- Di Filippo, M.M.; Santoro, M.; Lovreglio, P.; Monno, R.; Capolongo, C.; Calia, C.; Fumarola, L.; D’Alfonso, R.; Berrilli, F.; Di Cave, D. Isolation and molecular characterization of freeliving amoebae from different water sources in Italy. Int. J. Environ. Res. Public Health 2015, 12, 3417–3427. [Google Scholar] [CrossRef]
- Loret, J.F.; Greub, G. Free-living amoebae: Biological by-passes in water treatment. Int. J. Hyg. Environ. Health 2010, 213, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Thomas, V. Sensitivity and Resistance of Protozoa to Microbicides. In Principles and Practice of Disinfection, Preservation and Sterilization; Fraise, A.P., Maillard, J.Y., Sattar, S.A., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2013; pp. 155–177. [Google Scholar]
- Dalrymple, K.; Stefanakos, W.; Trotz, M.A.; Goswami, D.Y. A review of the mechanisms and modeling of photocatalytic disinfection. Appl. Catal. B Environ. 2010, 98, 27–38. [Google Scholar] [CrossRef]
- Gabriel, A.A.; Panaligan, D.C. Heat and chlorine resistance of a soil Acanthamoeba sp. cysts in water. J. Appl. Microbiol. 2020, 129, 453–464. [Google Scholar] [CrossRef]
- Lonnen, J.; Kilvington, S.; Kehoe, S.C.; Al-Touatic, F.; McGuiganc, K.G. Solar and photocatalytic disinfection of protozoan, fungal and bacterial microbes in drinking water. Water Res. 2005, 39, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Kilvington, S.; Price, J. Survival of Legionella pneumophila within cysts of Acanthamoeba polyphaga following chlorine exposure. J. Appl. Bacteriol. 1990, 68, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.T.; Jones, S.; Pelaz, C.; Millar, R.D.; Kwaik, Y.A. Acanthamoeba polyphaga resuscitates viable non-culturable Legionella pneumophila after disinfection. Environ. Microbiol. 2007, 9, 1267–1277. [Google Scholar] [CrossRef]
- He, Z.; Wang, L.; Ge, Y.; Zhang, S.; Tian, Y.; Yang, X.; Shu, L. Both viable and inactivated amoeba spores protect their intracellular bacteria from drinking water disinfection. J. Hazard. Mater. 2021, 47, 126006. [Google Scholar] [CrossRef] [PubMed]
- Muchesa, P.; Bartie, C.; Barnard, T.G. Free-Living Amoebae and Associated Drug-Resistant Bacteria in Surface Waters: A Public Health Concern? In Parasite and Disease Spread by Major Rivers on Earth. Parasitology Research Monographs; Mehlhorn, H., Klimpel, S., Eds.; Springer: Cham, Switzerland, 2019; Volume 12, pp. 221–236. [Google Scholar] [CrossRef]
- Zhou, P.; Di Giovanni, G.D.D.; Meschke, J.S.; Dodd, M.C. Enhanced Inactivation of Cryptosporidium parvum Oocysts during Solar Photolysis of Free Available Chlorine. Environ. Sci. Technol. Lett. 2014, 1, 453–458. [Google Scholar] [CrossRef]
- Remucal, C.K.; Manley, D. Emerging investigators series: The efficacy of chlorine photolysis as an advanced oxidation process for drinking water treatment. Environ. Sci. Water Res. Technol. 2016, 2, 565–579. [Google Scholar] [CrossRef]
- Khajouei, G.; Finkle, H.O.; Lin, L.S. UV/chlorine advanced oxidation processes for degradation of contaminants in water and wastewater: A comprehensive review. J. Environ. Chem. Eng. 2022, 10, 107508. [Google Scholar] [CrossRef]
- Forsyth, J.E.; Zhou, P.; Mao, Q.; Asato, S.S.; Meschke, J.S.; Dodd, M.C. Enhanced inactivation of Bacillus subtilis spores during solar photolysis of free available chlorine. Environ. Sci. Technol. 2013, 47, 12976–12984. [Google Scholar] [CrossRef]
- Chauque, B.J.M.; Rott, M.B. Photolysis of sodium chloride and sodium hypochlorite by ultraviolet light inactivates the trophozoites and cysts of Acanthamoeba castellanii in the water matrix. J. Water Health 2021, 19, 190–202. [Google Scholar] [CrossRef]
- Wang, L.; Mai, Y.; Li, S.; Shu, L.; Fang, J. Efficient inactivation of amoeba spores and their intraspore bacteria by solar/chlorine: Kinetics and mechanisms. Water Res. 2023, 242, 120288. [Google Scholar] [CrossRef]
- Chauque, B.J.M.; Benetti, A.D.; Corçao, G.; Silva, C.E.; Gonçalves, R.F.; Rott, M.B. A new continuous-flow solar water disinfection system inactivating cysts of Acanthamoeba castellanii, and bacteria. Photochem. Photobiol. Sci. 2021, 20, 123–137. [Google Scholar] [CrossRef]
- Ortillés, A.; Goñi, P.; Rubio, E.; Sierra, M.; Gámez, E.; Fernandez, M.T.; Benito, M.; Cristóbal, J.A.; Calvo, B. A rabbit model of Acanthamoeba keratitis: Use of infected soft contact lenses after corneal epithelium debridement with a diamond burr. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.M.; Booton, G.C.; Hay, J.; Niszl, I.A.; Seal, D.V.; Markus, M.B.; Fuerst, P.A.; Byers, T.J. Use of subgenic 18S ribosomal DNA PCR and sequencing for genus and genotype identification of Acanthamoebae from humans with keratitis and from sewage sludge. J. Clin. Microbiol. 2001, 39, 1903–1911. [Google Scholar] [CrossRef] [PubMed]
- Claassen, J.A.H.R. The gold standard: Not a golden standard. BMJ 2005, 330, 1121. [Google Scholar] [CrossRef]
- Yuranova, T.; Enea, O.; Mielczarski, E.; Mielczarski, J.; Albers, P.; Kiwi, J. Fenton immobilized photo-assisted catalysis through a Fe/C structured fabric. Appl. Catal. B. Environ. 2004, 49, 39–50. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. Alternative Disinfectants and Oxidants Guidance Manual; Office of Water: Washington, DC, USA, 1999; EPA 815-R-99-014. Available online: https://www.epa.gov/ (accessed on 20 December 2023).
- Kahler, A.M.; Cromeans, T.L.; Roberts, J.M.; Hill, V.R. Effects of source water quality on chlorine inactivation of adenovirus, coxsackievirus, echovirus and murine norovirus. Appl. Environ. Microbiol. 2010, 76, 5159–5164. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005; ISBN 08-7553-047-8. [Google Scholar]
- Beattie, T.K.; Seal, D.V.; Tomlinson, A.; McFadyen, A.K.; Grimason, A.M. Determination of amoebicidal activities of multipurpose contact lens solutions by using a most probable number enumeration technique. J. Clin. Microbiol. 2003, 41, 2992–3000. [Google Scholar] [CrossRef]
- Storey, M.V.; Winiecka-Krusnelli, J.; Ashbolt, N.J.; Stenström, T.A. The Efficacy of Heat and Chlorine Treatment against Thermotolerant Acanthamoebae and Legionellae. Scand. J. Infect. Dis. 2004, 36, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Coulon, C.; Collignon, A.; McDonnell, G.; Thomas, V. Resistance of Acanthamoeba cysts to disinfection treatments used in health care settings. J. Clin. Microbiol. 2010, 48, 2689–2697. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, M.; Berne, F.; Herbelin, P.; Binet, M.; Berthelot, N.; Rodier, M.H.; Soreau, S.; Héchard, Y. Sensitivity of free-living amoeba trophozoites and cysts to water disinfectants. Int. J. Hyg. Environ. Health 2014, 217, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.C.; Ortega, Y.R. Review: Inactivation of protozoan parasites in food, water and environmental systems. J. Food Prot. 2006, 69, 2786–2808. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, A.; Corno, G.; Manaia, C.M.; Rizzo, L. Impact of disinfection processes on bacterial community in urban wastewater: Should we rethink microbial assessment methods? J. Environ. Chem. Eng. 2020, 8, 104393. [Google Scholar] [CrossRef]
- Giannakis, S.; Polo, M.I.; Spuhler, D.; Sánchez, J.A.; Fernández, P.; Pulgarin, C. Solar disinfection is an augmentable, in situ-generated photo-Fenton reaction—Part 1: A review of the mechanisms and the fundamental aspects of the process. Appl. Catal. B Environ. 2016, 199, 199–223. [Google Scholar] [CrossRef]
- Adan, C.; Magnet, A.; Fenoy, S.; Pablos, C.; Águila, C.; Marugán, J. Concomitant inactivation of Acanthamoeba spp. and Escherichia coli using suspended and immobilized TiO2. Water Res. 2018, 144, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Heaselgrave, W.; Kilvington, S. The efficacy of simulated solar disinfection (SODIS) against Ascaris, Giardia, Acanthamoeba, Naegleria, Entamoeba and Cryptosporidium. Acta Trop. 2011, 119, 138–143. [Google Scholar] [CrossRef]
- Dupuy, M.; Mazoua, S.; Berne, F.; Bodet, C.; Garrec, N.; Herbelin, P.; Ménard-Szczebara, F.; Oberti, S.; Rodier, M.H.; Soreau, S.; et al. Efficiency of water disinfectants against Legionella pneumophila and Acanthamoeba. Water Res. 2011, 45, 1087–1094. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Zhang, B.; Queller, D.C.; Strassmann, J.E. Burkholderia bacteria use chemotaxis to find social amoeba Dictyostelium discoideum hosts. ISME J. 2018, 12, 1977–1993. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A. Pathogenesis of Acanthamoeba infections. Microb. Pathog. 2003, 34, 277–285. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C1-211 | P31 | |||||
|---|---|---|---|---|---|---|
| Treatment | [Cl2]99% | [Cl2]99.9% | [Cl2]EB | [Cl2]99% | [Cl2]99.9% | [Cl2]EB |
| Cl2 | 55 | 94 | 500 | 159 | 247 | >500 |
| Cl2/SR320–800nm | 4 | 8 | 50 | 178 | 319 | 500 |
| Cl2/SR280–800nm | - | - | 5 | 1 | 4 | 100 |
| Treatment | CT99% | CT99.9% | CTEB | CT99% | CT99.9% | CTEB |
| Cl2 | 1556 | 2667 | 12,471 | 3680 | 5680 | >10,820 |
| Treatment | F99% | F99.9% | FEB | F99% | F99.9% | FEB |
| SR320–800nm | >900 | >900 | >900 | >900 | >900 | >900 |
| SR280–800nm | 178 | 285 | 900 | - | - | >900 |
| Initial Concentration (mg Cl2/L) | Cl2 | Cl2/SR280–800nm | Cl2/SR320–800nm | ||||
|---|---|---|---|---|---|---|---|
| 0 min | 30 min | 0 min | 5 min | 10 min | 0 min | 30 min | |
| 0 | 7.0 | 7.0 | 7.0 | 7.0 | 7.1 | 7.0 | 7.4 |
| 1 | 7.0 | 7.1 | nd | nd | nd | 7.1 | 7.4 |
| 5 | 8.4 | 8.2 | 8.4 | 8.6 | 9.5 | 8.6 | 7.4 |
| 10 | 8.6 | 7.6 | nd | nd | nd | 8.7 | 8.2 |
| 50 | 9.9 | 9.0 | 9.9 | 10.0 | 10.3 | 9.9 | 9.0 |
| 100 | 10.3 | 9.6 | nd | nd | nd | 10.3 | 9.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menacho, C.; Soler, M.; Chueca, P.; Ormad, M.P.; Goñi, P. Chlorine Photolysis: A Step Forward in Inactivating Acanthamoeba and Their Endosymbiont Bacteria. Water 2024, 16, 668. https://doi.org/10.3390/w16050668
Menacho C, Soler M, Chueca P, Ormad MP, Goñi P. Chlorine Photolysis: A Step Forward in Inactivating Acanthamoeba and Their Endosymbiont Bacteria. Water. 2024; 16(5):668. https://doi.org/10.3390/w16050668
Chicago/Turabian StyleMenacho, Carmen, Maria Soler, Patricia Chueca, Maria P. Ormad, and Pilar Goñi. 2024. "Chlorine Photolysis: A Step Forward in Inactivating Acanthamoeba and Their Endosymbiont Bacteria" Water 16, no. 5: 668. https://doi.org/10.3390/w16050668
APA StyleMenacho, C., Soler, M., Chueca, P., Ormad, M. P., & Goñi, P. (2024). Chlorine Photolysis: A Step Forward in Inactivating Acanthamoeba and Their Endosymbiont Bacteria. Water, 16(5), 668. https://doi.org/10.3390/w16050668