
The Diversity of Freshwater Stygobiotic Crustaceans in the Republic of North Ossetia–Alania Provides New Evidence for the Existence of an Ancient Glacial Refugium in the North Caucasus Region

Abstract
:1. Introduction
2. Materials and Methods
3. Results
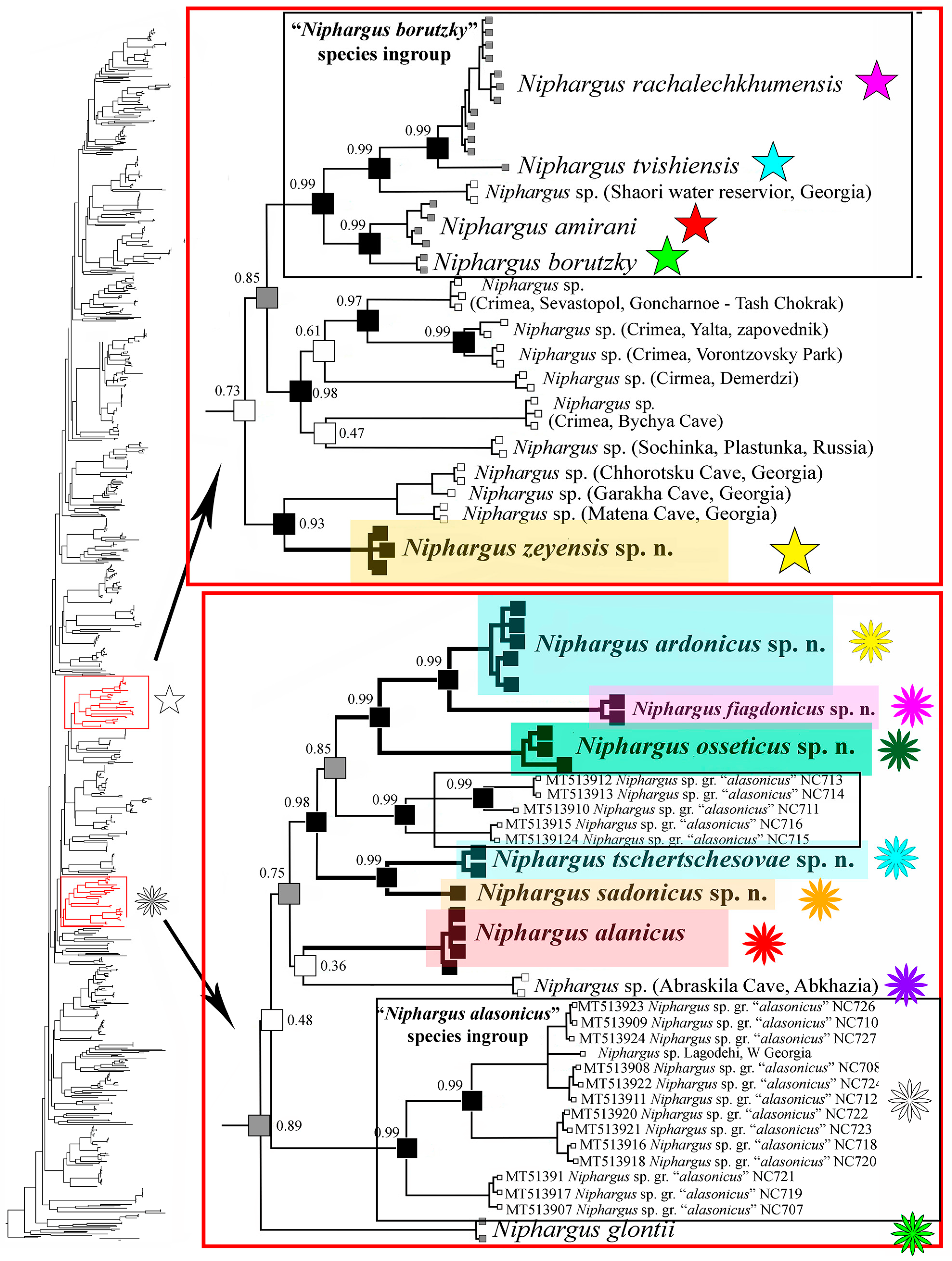
3.1. Phylogenetic Part
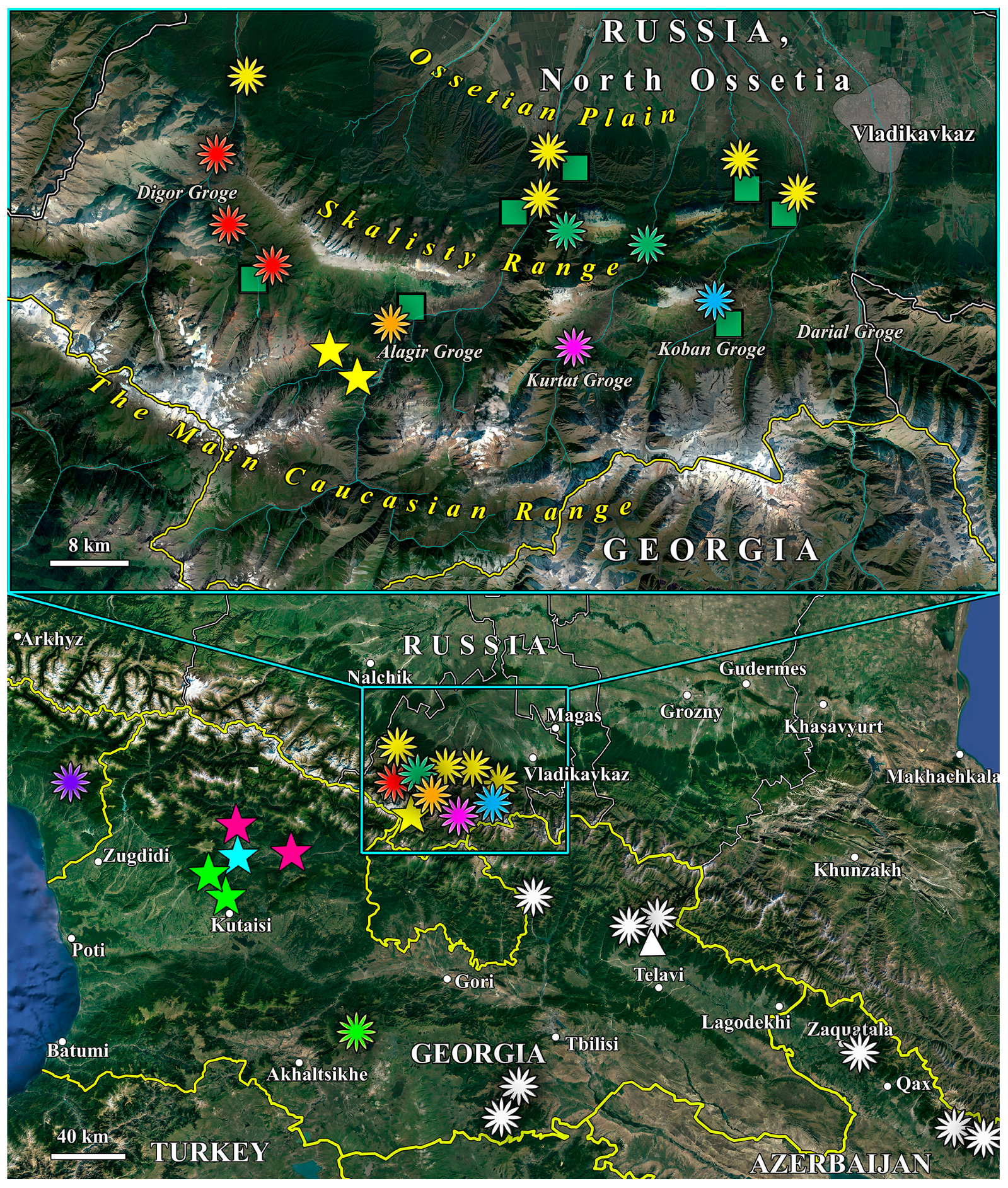
3.2. Geographic Distribution
3.3. Taxonomic Part
 , 4
, 4 (LEMMI)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, a spring in the valley of the Ardon River, about 8 km south of the town of Alagir, 42°55′31.79″ N 44°11′26.14″ E, 823 m asl, coll. D. Palatov, 3 October 2020; 1
(LEMMI)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, a spring in the valley of the Ardon River, about 8 km south of the town of Alagir, 42°55′31.79″ N 44°11′26.14″ E, 823 m asl, coll. D. Palatov, 3 October 2020; 1 , 3 (LEMMI)—a stream in an abandoned mine near the village of Sadon, 42°50′40.28″ N 44°01′16.77″ E, 1155 m asl, coll. D. Palatov, 8 October 2020; 6, 10 (LEMMI)—a spring in the valley of the Mayramadag River, 2 km south of the village, 42°59′47.96″ N 44°29′37.87″ E, 670 m asl, coll. D. Palatov, 9 May 2022; 3, 7 (LEMMI)—Irafsky District, a spring seeping on the banks of the Dargonkom River Valley, 42°53′26.8″ N 43°51′58.3″ E, 1860 m asl, coll. D. Palatov, 3 November 2021; 5, 12 (LEMMI)—Prigorodny District, groundwater of the Gizeldon River near the village of Dargavz, 42°51′08.76″ N 44°26′55.55″ E, 1395 m asl, coll. D. Palatov, M. Antipova, 22 May 2023; 1, 2 (LEMMI)—a spring stream in the floodplain of the Gizeldon River, opposite the “Alpina Park” complex, 42°58′39.84″ N 44°34′11.35″ E, 763 m asl, coll. D. Palatov, M. Antipova, 24 May 2023. with weakly expressed basal apophysis, short distal apophysis, and small goulot with developed lips. Endopodite of PlII in , subtriangular, with three short, marginal, simple setae. Lateral and terminal margins of PlIII with 10–11 short simple setae, with endopodites about 1.6 times shorter than exopodites. Lateral margin of exopodite of PlIV without setae. Endopodite of PlIV suboval, about 1.3 times shorter than the exopodite. Exopodite of PlV ovoid, elongated, 2.2 times as long as wide, and with lateral margins without setae; endopodite suboval and about 90% of exopodite. Uropods similar in and ; proto-, endo-, and exopodite length ratio of 1/1.2/1.3, respectively. [20]. However, molecular genetic study indicates that individuals with various morphologies belong to the same biological species (see above). (bl. 10 mm) (ZMMU Mb-1163)—Russian Federation, Republic of North Ossetia–Alania, Irafsky District, Sardidon River Valley, Fansal village, 42°56.0980′ N 43°49.0340′ E, about 1430 m asl, in an abandoned mine, coll. D. Palatov, 31 December 2017; 35 (LEMMI)—same locality, coll. D. Palatov, 3 August 2019; 3 (LEMMI)—Dargonkom River Valley, 42°53’26.0" N 43°52’53.0" E, about 1960 m asl, in helocrene springs, coll. D. Palatov, 30 December 2017.
, 3 (LEMMI)—a stream in an abandoned mine near the village of Sadon, 42°50′40.28″ N 44°01′16.77″ E, 1155 m asl, coll. D. Palatov, 8 October 2020; 6, 10 (LEMMI)—a spring in the valley of the Mayramadag River, 2 km south of the village, 42°59′47.96″ N 44°29′37.87″ E, 670 m asl, coll. D. Palatov, 9 May 2022; 3, 7 (LEMMI)—Irafsky District, a spring seeping on the banks of the Dargonkom River Valley, 42°53′26.8″ N 43°51′58.3″ E, 1860 m asl, coll. D. Palatov, 3 November 2021; 5, 12 (LEMMI)—Prigorodny District, groundwater of the Gizeldon River near the village of Dargavz, 42°51′08.76″ N 44°26′55.55″ E, 1395 m asl, coll. D. Palatov, M. Antipova, 22 May 2023; 1, 2 (LEMMI)—a spring stream in the floodplain of the Gizeldon River, opposite the “Alpina Park” complex, 42°58′39.84″ N 44°34′11.35″ E, 763 m asl, coll. D. Palatov, M. Antipova, 24 May 2023. with weakly expressed basal apophysis, short distal apophysis, and small goulot with developed lips. Endopodite of PlII in , subtriangular, with three short, marginal, simple setae. Lateral and terminal margins of PlIII with 10–11 short simple setae, with endopodites about 1.6 times shorter than exopodites. Lateral margin of exopodite of PlIV without setae. Endopodite of PlIV suboval, about 1.3 times shorter than the exopodite. Exopodite of PlV ovoid, elongated, 2.2 times as long as wide, and with lateral margins without setae; endopodite suboval and about 90% of exopodite. Uropods similar in and ; proto-, endo-, and exopodite length ratio of 1/1.2/1.3, respectively. [20]. However, molecular genetic study indicates that individuals with various morphologies belong to the same biological species (see above). (bl. 10 mm) (ZMMU Mb-1163)—Russian Federation, Republic of North Ossetia–Alania, Irafsky District, Sardidon River Valley, Fansal village, 42°56.0980′ N 43°49.0340′ E, about 1430 m asl, in an abandoned mine, coll. D. Palatov, 31 December 2017; 35 (LEMMI)—same locality, coll. D. Palatov, 3 August 2019; 3 (LEMMI)—Dargonkom River Valley, 42°53’26.0" N 43°52’53.0" E, about 1960 m asl, in helocrene springs, coll. D. Palatov, 30 December 2017. (bl. 10 mm) (LEMMI)—Urukh Cave, coll. S.A. Kapralov, 19 September 2020. has bl. 10.5 mm. (bl. 10 mm) (ZMMU Mb-1269)—Russian Federation, North Caucasus, the Republic of North Ossetia–Alania, Alagirsky District, a spring in the Ardon River Valley, about 8 km south of Alagir, 42°55′31.79″ N 44°11′26.14″ E, 823 m asl, coll. D. Palatov, 3 October 2020. (ZMMU Mb-1270)—same data as for holotype. (LEMMI)—same data as for holotype; 5 (LEMMI)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, a spring in the Ardon River Valley, about 2 km south of Alagir, 42°58′31.8″ N 44°13′11.2″ E, about 670 m asl, coll. D. Palatov, 23 September 2020; 12 (LEMMI)—Alagirsky District, a spring in the Mayramadag River Valley, 2 km south of Mayramadag, 42°59′47.96″ N 44°29′37.87″ E, about 655 m asl, coll. D. Palatov, 9 May 2022; 7 (LEMMI)—Irafsky District, a spring in a damp beech forest, located 2 km south of Kalukh, 43°4′37″ N 43°49′22.7″ E, about 970 m asl, coll. D. Palatov, 5 November 2021; 2 (LEMMI)—Prigorodny District, a spring stream in the floodplain of the Gizeldon River, opposite the “Alpina Park” complex, 42°58′39.84″ N 44°34′11.35″ E, 763 m asl, coll. D. Palatov, M. Antipova, 24 May 2023. had bl. 10.5 mm. (bl. 12 mm) (ZMMU Mb-1271)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, a spring in the Fiagdon River Valley, near the Alan Holy Dormition Monastery in Hidikus, 42°49′18.5″ N 44°16′17.8″ E, 1295 m asl, coll. D. Palatov, 16 May 2023. had bl. 12.0 mm. (bl. 11.5 mm) (ZMMU Mb-1272)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, a spring inside the Agomskaya Cave, Ardon River Basin, bank of Akhshkadon Gorge, 2.9 km northwest from Oldukhankhokh mt., about 1209 m asl, coll. S. Kapralov, 17 September 2020. (ZMMU Mb-1273)—same data as for holotype. (LEMMI)—same data as for holotype; 3 (LEMMI)—Russian Federation, the Republic of North Ossetia–Alania, Prigorodny District, a spring on the right bank of the Fiagdon River opposite the “cascading waterfall”, 42°55′3.7″ N 44°22′10.1″ E, about 1000 m asl, coll. D. Palatov, 18 July 2021. had bl. 11.5 mm. (bl. 10 mm) (ZMMU Mb-1274)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, a stream in an abandoned mine near the village of Sadon, 42°50′40.28″ N 44°01′16.77″ E, about 1155 m asl, coll. D. Palatov, 8 October 2020. (ZMMU Mb-1275)—same data as for holotype. (LEMMI)—same data as for holotype. had bl. 10 mm. (bl. 10 mm) (ZMMU Mb-1276)—Russian Federation, Republic of North Ossetia–Alania, Prigorodny District, groundwater of the Gizeldon River near the village of Dargavz, 42°51′08.76″ N 44°26′55.55″ E, 1395 m asl, coll. D. Palatov, M. Antipova, 22 May 2023. (ZMMU Mb-1277)—same data as for holotype. (LEMMI)—same data as for holotype. has bl. 10.5 mm. (bl. 12.5 mm) (ZMMU Mb-1278)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, Tsey (Zey) Gorge, a stream on the western outskirts of the village of Abaytikau, 42°48′31.7″ N 43°57′18.2″ E, about 1985 m asl, coll. D. Palatov, 28 July 2021. (ZMMU Mb-1279)—same data as holotype. (ZMMU Mb-1279)—same data as holotype; 10—Russian Federation, North Caucasus, the Republic of North Ossetia–Alania, Alagirsky District, a small spring near the road to the Tsey mountain camp (Tsey Gorge), 42°48′11.51″ N 43°57′21.56″ E, about 1722 m asl, coll. D. Palatov, 7 October 2020. had bl. 12.5 mm. (vs. distal article about 1/3 of the proximal one), (2) epimeral plates II–III armed with paired spines along the ventral margins (vs. single spines in a row), (3) dactyli of gnathopods I–II with several tufts of setae along the outer margins (vs. only one seta), (4) telson with small and thin setae on the dorsal surface and inner margins of lobes of the telson (vs. absent), (5) 6 apical spines on each lobe of the telson (vs. 3–5 spines, except N. alanicus, which also has 6 spines), and (6) 5–6 (usually 6) hooks in the retinacula of pleopods (vs. maximum 5 hooks, usually less)., which is almost equal to the basal article; (3) the almost subrectangular basis of pereiopod VII; (4) the presence of small additional setae on the dactyli of ambulatory pereiopods ventrally (vs. well-marked strong spines); and (5) the presence of paired ventral spines on epimeral plate II (vs. single ventral spines).
(bl. 10 mm) (LEMMI)—Urukh Cave, coll. S.A. Kapralov, 19 September 2020. has bl. 10.5 mm. (bl. 10 mm) (ZMMU Mb-1269)—Russian Federation, North Caucasus, the Republic of North Ossetia–Alania, Alagirsky District, a spring in the Ardon River Valley, about 8 km south of Alagir, 42°55′31.79″ N 44°11′26.14″ E, 823 m asl, coll. D. Palatov, 3 October 2020. (ZMMU Mb-1270)—same data as for holotype. (LEMMI)—same data as for holotype; 5 (LEMMI)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, a spring in the Ardon River Valley, about 2 km south of Alagir, 42°58′31.8″ N 44°13′11.2″ E, about 670 m asl, coll. D. Palatov, 23 September 2020; 12 (LEMMI)—Alagirsky District, a spring in the Mayramadag River Valley, 2 km south of Mayramadag, 42°59′47.96″ N 44°29′37.87″ E, about 655 m asl, coll. D. Palatov, 9 May 2022; 7 (LEMMI)—Irafsky District, a spring in a damp beech forest, located 2 km south of Kalukh, 43°4′37″ N 43°49′22.7″ E, about 970 m asl, coll. D. Palatov, 5 November 2021; 2 (LEMMI)—Prigorodny District, a spring stream in the floodplain of the Gizeldon River, opposite the “Alpina Park” complex, 42°58′39.84″ N 44°34′11.35″ E, 763 m asl, coll. D. Palatov, M. Antipova, 24 May 2023. had bl. 10.5 mm. (bl. 12 mm) (ZMMU Mb-1271)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, a spring in the Fiagdon River Valley, near the Alan Holy Dormition Monastery in Hidikus, 42°49′18.5″ N 44°16′17.8″ E, 1295 m asl, coll. D. Palatov, 16 May 2023. had bl. 12.0 mm. (bl. 11.5 mm) (ZMMU Mb-1272)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, a spring inside the Agomskaya Cave, Ardon River Basin, bank of Akhshkadon Gorge, 2.9 km northwest from Oldukhankhokh mt., about 1209 m asl, coll. S. Kapralov, 17 September 2020. (ZMMU Mb-1273)—same data as for holotype. (LEMMI)—same data as for holotype; 3 (LEMMI)—Russian Federation, the Republic of North Ossetia–Alania, Prigorodny District, a spring on the right bank of the Fiagdon River opposite the “cascading waterfall”, 42°55′3.7″ N 44°22′10.1″ E, about 1000 m asl, coll. D. Palatov, 18 July 2021. had bl. 11.5 mm. (bl. 10 mm) (ZMMU Mb-1274)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, a stream in an abandoned mine near the village of Sadon, 42°50′40.28″ N 44°01′16.77″ E, about 1155 m asl, coll. D. Palatov, 8 October 2020. (ZMMU Mb-1275)—same data as for holotype. (LEMMI)—same data as for holotype. had bl. 10 mm. (bl. 10 mm) (ZMMU Mb-1276)—Russian Federation, Republic of North Ossetia–Alania, Prigorodny District, groundwater of the Gizeldon River near the village of Dargavz, 42°51′08.76″ N 44°26′55.55″ E, 1395 m asl, coll. D. Palatov, M. Antipova, 22 May 2023. (ZMMU Mb-1277)—same data as for holotype. (LEMMI)—same data as for holotype. has bl. 10.5 mm. (bl. 12.5 mm) (ZMMU Mb-1278)—Russian Federation, Republic of North Ossetia–Alania, Alagirsky District, Tsey (Zey) Gorge, a stream on the western outskirts of the village of Abaytikau, 42°48′31.7″ N 43°57′18.2″ E, about 1985 m asl, coll. D. Palatov, 28 July 2021. (ZMMU Mb-1279)—same data as holotype. (ZMMU Mb-1279)—same data as holotype; 10—Russian Federation, North Caucasus, the Republic of North Ossetia–Alania, Alagirsky District, a small spring near the road to the Tsey mountain camp (Tsey Gorge), 42°48′11.51″ N 43°57′21.56″ E, about 1722 m asl, coll. D. Palatov, 7 October 2020. had bl. 12.5 mm. (vs. distal article about 1/3 of the proximal one), (2) epimeral plates II–III armed with paired spines along the ventral margins (vs. single spines in a row), (3) dactyli of gnathopods I–II with several tufts of setae along the outer margins (vs. only one seta), (4) telson with small and thin setae on the dorsal surface and inner margins of lobes of the telson (vs. absent), (5) 6 apical spines on each lobe of the telson (vs. 3–5 spines, except N. alanicus, which also has 6 spines), and (6) 5–6 (usually 6) hooks in the retinacula of pleopods (vs. maximum 5 hooks, usually less)., which is almost equal to the basal article; (3) the almost subrectangular basis of pereiopod VII; (4) the presence of small additional setae on the dactyli of ambulatory pereiopods ventrally (vs. well-marked strong spines); and (5) the presence of paired ventral spines on epimeral plate II (vs. single ventral spines).- Niphargus ardonicus sp. nov.: urn:lsid:zoobank.org:pub:CB1BB16F-2225-4A04-8438-5E37F2B2EDE8
- Niphargus fiagdonicus sp. nov.: urn:lsid:zoobank.org:pub:CB1BB16F-2225-4A04-8438-5E37F2B2EDE8
- Niphargus osseticus sp. nov.: urn:lsid:zoobank.org:pub:CB1BB16F-2225-4A04-8438-5E37F2B2EDE8
- Niphargus sadonicus sp. nov.: urn:lsid:zoobank.org:act:CC4DD058-25E6-4C11-92BB-27CC3FBEE368
- Niphargus zeyensis sp. nov.: urn:lsid:zoobank.org:pub:CB1BB16F-2225-4A04-8438-5E37F2B2EDE8
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Krever, V.; Zazanashvili, N.; Jungius, H.; Williams, L.; Petelin, D. Biodiversity of the Caucasus Ecoregion; World Wide Fund for Nature: Moscow, Russia, 2001. [Google Scholar]
- Tarkhnishvili, D. Historical Biogeography of the Caucasus; Nova Science Publishers: New York, NY, USA, 2014. [Google Scholar]
- Shatberashvili, N.; Rucevska, I.; Jørstad, H.; Artsivadze, K.; Mehdiyev, B.; Aliyev, M.; Fayvush, G.; Dzneladze, M.; Jurek, M.; Kirkfeldt, T.; et al. Outlook on Climate Change Adaptation in the South Caucasus Mountains; United Nations Environment Programme, GRID-Arendal and Sustainable Caucasus: Nairobi, Kenya; Arendal, Norway; Tbilisi, Georgia, 2016. [Google Scholar]
- Hewitt, G.M. The structure of biodiversity insights from molecular phylogeography. Front. Zool. 2004, 1, 4. [Google Scholar] [CrossRef]
- Cane, M.A.; Braconnot, P.; Clement, A.; Gildor, H.; Joussaume, S.; Kageyama, M.; Khodri, M.; Paillard, D.; Tett, S.; Zorita, E. Progress in paleoclimate modeling. J. Clim. 2006, 19, 5031–5057. [Google Scholar] [CrossRef]
- Schmitt, T. Molecular biogeography of Europe: Pleistocene cycles and postglacial trends. Front. Zool. 2007, 4, 11. [Google Scholar] [CrossRef]
- Schmitt, T.; Varga, Z. Extra-Mediterranean refugia: The rule and not the exception? Front. Zool. 2012, 9, 22. [Google Scholar] [CrossRef]
- Keppel, G.; Van Niel, K.P.; Wardell-Johnson, G.W.; Yates, C.J.; Byrne, M.; Mucina, L.; Schut, A.G.; Hopper, S.D.; Franklin, S.E. Refugia: Identifying and understanding safe havens for biodiversityunder climate change. Glob. Ecol. Biogeogr. 2012, 21, 393–404. [Google Scholar] [CrossRef]
- Keppel, G.; Wardell-Johnson, G.W. Refugia: Keys to climatechange management. Glob. Chang. Biol. 2012, 18, 2389–2391. [Google Scholar] [CrossRef]
- Gavin, D.G.; Fitzpatrick, M.C.; Gugger, P.F.; Heath, K.D.; Rodríguez-Sánchez, F.; Dobrowski, S.Z.; Hampe, A.; Hu, F.S.; Ashcroft, M.B.; Bartlein, P.J.; et al. Climate refugia: Joint inference from fossil records, species distribution models and phylogeography. New Phytol. 2014, 204, 37–54. [Google Scholar] [CrossRef]
- Stewart, J.R.; Lister, A.M. Cryptic northern refugia and the origins of the modern biota. Trends Ecol. Evol. 2001, 16, 608–613. [Google Scholar] [CrossRef]
- Stewart, J.R.; Lister, A.M.; Barnes, I.; Dalen, L. Refugia revisited: Individualistic responses of species in space and time. Proc. R. Soc. Lond. Ser. B 2010, 277, 661–671. [Google Scholar] [CrossRef]
- Bennett, K.D.; Provan, J. What do we mean by ’refugia’? Quat. Sci. Rev. 2008, 27, 2449–2455. [Google Scholar] [CrossRef]
- Kolakovsky, A.A. The Flora and Vegetation of Colchida; The Publishing House of Moscow University: Moscow, Russia, 1991; 215p. [Google Scholar]
- Shatilova, I.; Mchedlishvili, N.; Rukhadze, L.; Kvavadze, E. The History of the Flora and Vegetation of Georgia (South Caucasus); Georgian National Museum: Tbilisi, Georgia, 2011; 345p. [Google Scholar]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global biodiversity conservation: The critical role of hotspots. In Biodiversity Hotspots: Distribution and Protection of Conservation Priority Areas; Zachos, F.E., Habel, J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–22. [Google Scholar] [CrossRef]
- Nieto, G.F. Southern European glacial refugia: A tale of tales. Taxon 2011, 60, 365–372. [Google Scholar] [CrossRef]
- Marin, I.; Palatov, D. A new species of the genus Niphargus (Crustacea: Amphipoda: Niphargidae) from the south-western part of the North Caucasus. Zool. Middle East 2019, 65, 336–346. [Google Scholar] [CrossRef]
- Palatov, D.M.; Sokolova, A.M. Two new stygobiotic species of the genus Proasellus (Crustacea: Isopoda: Asellidae) from the North Caucasus. Invertebr. Zool. 2021, 18, 481–501. [Google Scholar] [CrossRef]
- Marin, I.; Palatov, D. Insights on the Existence of Ancient Glacial Refugee in the Northern Black/Azov Sea Lowland, with the Description of the First Stygobiotic Microcrustacean Species of the Genus Niphargus Schiödte, 1849 from the Mouth of the Don River. Diversity 2023, 15, 682. [Google Scholar] [CrossRef]
- Trontelj, P.; Douady, C.J.; Fišer, C.; Gibert, J.; Gorički, S.; Lefébure, T.; Sket, B.; Zakšek, V. A molecular test for cryptic diversity in ground water: How large are the ranges of macro-stygobionts? Freshw. Biol. 2009, 54, 727–744. [Google Scholar] [CrossRef]
- Eme, D.; Malard, F.; Konecny-Dupre, L.; Lefébure, T.; Douady, C.J. Bayesian phylogeographic inferences reveal contrasting colonization dynamics among European groundwater isopods. Mol. Ecol. 2013, 22, 5685–5699. [Google Scholar] [CrossRef]
- McInerney, C.E.; Maurice, L.; Robertson, A.L.; Lee, R.F.D.K.; Arnscheidt, J.; Venditti, C.; Dooley, J.S.G.; Mathers, T.; Matthijs, S.; Eriksson, K.; et al. The ancient Britons: Groundwater fauna survived extreme climate change over tens of millions of years across NW Europe. Mol. Ecol. 2014, 23, 1153–1166. [Google Scholar] [CrossRef]
- Rendoša, M.; Delić, T.; Copilaș-Ciocianu, D.; Fišer, C. First insight into cryptic diversity of a Caucasian subterranean amphipod of the genus Niphargus (Crustacea: Amphipoda: Niphargidae). Zool. Anz. 2021, 290, 1–11. [Google Scholar] [CrossRef]
- Palatov, D.M.; Marin, I.N. Epigean (pond-dwelling) species of the genus Niphargus Schiödte, 1849 (Crustacea: Amphipoda: Niphargidae) from the coastal plains of the Black and Azov seas of the north- and south-western Caucasus. Invertebr. Zool. 2021, 18, 105–151. [Google Scholar] [CrossRef]
- Marin, I.; Krylenko, S.; Palatov, D. Euxinian relict amphipods of the Eastern Paratethys in the subterranean fauna of coastal habitats of the Northern Black Sea region. Invertebr. Zool. 2021, 18, 247–320. [Google Scholar] [CrossRef]
- Palatov, D.M.; Dzhamirzoev, G.S.; Sokolova, A.M. A new stygobiotic species of the genus Proasellus (Crustacea: Isopoda: Asellidae) from South Dagestan, Russia. Invertebr. Zool. 2023, 20, 295–306. [Google Scholar] [CrossRef]
- Marin, I.; Krylenko, S.; Palatov, D. The Caucasian relicts: A new species of the genus Niphargus (Crustacea: Amphipoda: Niphargidae) from the Gelendzhik–Tuapse area of the Russian southwestern Caucasus. Zootaxa 2021, 4963, 483–504. [Google Scholar] [CrossRef]
- Marin, I.; Barjadze, S.; Marghradge, E.; Palatov, D. Diversity, taxonomy and phylogenetic relationships of the “Niphargus borutzkyi” ingroup (Crustacea: Amphipoda: Niphargidae) in Western Georgia, SW Caucasus. Zootaxa 2023, 5352, 477–500. [Google Scholar] [CrossRef] [PubMed]
- Birštein, Y.A. Malacostraca der Kutais-Hohlen am Rion (Transkaukasus, Georgien). Zool. Anzeig. 1933, 104, 143–156. [Google Scholar]
- Birstein, J.A. K faune pešernjih Amphipoda Abhazii (Über die Fauna der Höhlen Amphipoden Abchasiens). Bull. Soc. Nat. Moscou 1940, 49, 47–55. (In Russian) [Google Scholar]
- Birštein, J.A. Sovjetska biospeologi. XII Podzemnije bokoplavlji raniona Hosta-Gudauta [Les Amphipodes souterraines de la region de Khosta-Goudauta (Ouest-Transkaukasia)]. Bull. Soc. Nat. Moscou 1952, 57, 26–39. [Google Scholar]
- Birštein, J.A. Subterranean Waterlouses (Crustacea: Isopoda: Asellota) of Transcaucasia. Zool. Zhurnal 1967, 46, 856–865. (In Russian) [Google Scholar]
- Marin, I.N. Crustacean “cave fishes” from the Arabika karst massif (Abkhazia, Western Caucasus): New species of stygobiotic crustacean genera Xiphocaridinella and Niphargus from the Gegskaya Cave and adjacent area. Arthr. Sel. 2019, 28, 225–245. [Google Scholar] [CrossRef]
- Marin, I.N. The Quaternary speciation in the Caucasus: A new cryptic species of stygobiotic amphipod of the genus Niphargus (Crustacea: Amphipoda: Niphargidae) from the Kumistavi (Prometheus) Cave, Western Georgia. Arthr. Sel. 2020, 29, 419–432. [Google Scholar] [CrossRef]
- Marin, I.; Palatov, D. Cryptic refugee on the northern slope of the Greater Caucasian Ridge: Discovery of Niphargus (Crustacea: Amphipoda: Niphargidae) in the North Ossetia–Alania, North Caucasus, separated from its relatives in the late Miocene. Zool. Anzeig. 2021, 292, 163–183. [Google Scholar] [CrossRef]
- Palatov, D.M.; Chertoprud, M.V. Macrozoobenthos communities of springs and streams of the Eastern Circum-Pontic region. Inland Water Biol. 2020, 13, 583–596. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit 1 from diverse metazoan. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA 7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Avise, J.C. Molecular Markers, Natural History and Evolution; Chapman and Hall: New York, NY, USA, 1994; 511p. [Google Scholar] [CrossRef]
- Hebert, P.D.; Ratnasingham, S.; de Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 96–99. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Guy-Haim, T.; Simon-Blecher, N.; Frumkin, A.; Naaman, I.; Achituv, Y. Multiple transgressions and slow evolution shape the phylogeographic pattern of the blind cave-dwelling shrimp Typhlocaris. PeerJ 2018, 6, e5268. [Google Scholar] [CrossRef]
- Lefébure, T.; Douady, C.J.; Gouy, M.; Trontelj, P.; Briolay, J.; Gibert, J. Phylo-geography of a subterranean amphipod reveals cryptic diversity and dynamic evolution in extreme environments. Mol. Ecol. 2006, 15, 1797–1806. [Google Scholar] [CrossRef]
- Copilaş-Ciocianu, D.; Petrusek, A. The southwestern Carpathians as an ancient centre of diversity of freshwater gammarid amphipods: Insights from the Gammarus fossarum species complex. Mol. Ecol. 2015, 24, 3980–3992. [Google Scholar] [CrossRef]
- Derzhavin, A.N. The subterranean Amphipoda of Transcaucasus. Bull. Acad. Sci. Azerb. SSR 1945, 8, 27–43. (In Russian) [Google Scholar]
- Saccò, M.; Mammola, S.; Altermatt, F.; Alther, R.; Bolpagni, R.; Brancelj, A.; Brankovits, D.; Fišer, C.; Gerovasileiou, V.; Griebler, C.; et al. Groundwater is a hidden global keystone ecosystem. Authorea 2023, 30, e17066. [Google Scholar] [CrossRef]
- Marin, I.; Turbanov, I. Molecular genetic analysis of stygobiotic shrimps of the genus Xiphocaridinella (Crustacea: Decapoda: Atyidae) reveals a connection between distant caves in Central Abkhazia, southwestern Caucasus. Inter. J. Speleol. 2021, 50, 301–311. [Google Scholar] [CrossRef]
- Duque, C.; Rosenberry, D.O. Advances in the Study and Understanding of Groundwater Discharge to Surface Water. Water 2022, 14, 1698. [Google Scholar] [CrossRef]
- Tarkhnishvili, D.; Gavashelishvili, A.; Mumladze, L. Palaeoclimatic models help to understand current distribution of Caucasian forest species. Biol. J. Linn. Soc. 2012, 105, 231–248. [Google Scholar] [CrossRef]
- Seddon, J.M.; Santucci, F.; Reeve, N.; Hewitt, G.M. Caucasus Mountains divide postulated postglacial colonization routes in the white-breasted hedgehog, Erinaceus concolor. J. Evol. Biol. 2002, 15, 463–467. [Google Scholar] [CrossRef]
- Parvizi, E.; Keikhosravi, A.; Naderloo, R.; Solhjouy-Fard, S.; Sheibak, F.; Schubart, C.D. Phylogeography of Potamon ibericum (Brachyura: Potamidae) identifies quaternary glacial refugia within the Caucasus biodiversity hot spot. Ecol. Evol. 2019, 9, 4749–4759. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, D.; Kukushkin, O.V.; Avci, A.; Bunyatova, S.; Kumlutas, Y.; Ilgaz, C.; Polyakova, E.; Shiryaev, K.; Tuniyev, B.; Jandzik, D. The biogeography of Elaphe sauromates (Pallas, 1814), with a description of a new rat snake species. PeerJ 2019, 7, e6944. [Google Scholar] [CrossRef] [PubMed]
- Levin, B.A.; Gandlin, A.A.; Simonov, E.S.; Levina, M.A.; Barmintseva, A.E.; Japoshvili, B.; Mugue, N.S.; Mumladze, L.; Mustafayev, N.J.; Pashkov, A.N.; et al. Phylogeny, phylogeography and hybridization of Caucasian barbels of the genus Barbus (Actinopterygii, Cyprinidae). Mol. Phylogenet. Evol. 2019, 135, 31–44. [Google Scholar] [CrossRef]
- Taberlet, P.; Fumagalli, L.; Wust-Saucy, A.G.; Cosson, J.F. Comparative phylogeography and postglacial colonization routes in Europe. Mol. Ecol. 1998, 7, 453–464. [Google Scholar] [CrossRef]
- Waltari, E.; Hijmans, R.J.; Peterson, A.T.; Nyari, A.S.; Perkins, S.L.; Guralnick, R.P. Locating Pleistocene refugia: Comparing phylogeographic and ecological niche model predictions. PLoS ONE 2007, 2, e563. [Google Scholar] [CrossRef]
- Tarasov, P.E.; Volkova, V.S.; Webb, T.I.I.I.; Guiot, J.; Andreev, A.A.; Bezusko, L.G.; Bezusko, T.V.; Bykova, G.V.; Dorofeyuk, N.I.; Kvavadze, E.V.; et al. Last glacial maximum biomes reconstructed from pollen and plant macrofossil data from northern Eurasia. J. Biogeogr. 2000, 27, 609–620. [Google Scholar] [CrossRef]
- Popov, S.V.; Shcherba, I.G.; Ilyina, L.B.; Nevesskaya, L.A.; Paramonova, N.P.; Khondkarian, S.O.; Magyar, I. Late Miocene to Pliocene paleogeography of the Paratethys and its relation to the Mediterranean. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 238, 91–106. [Google Scholar] [CrossRef]
- Philip, H.; Cisternas, A.; Gvishiani, A.; Gorshkov, A. The Caucasus: An actual example of the initial stages of continental collision. Tectonophysics 1989, 161, 1–21. [Google Scholar] [CrossRef]
- Avdeev, B. Tectonics of the Greater Caucasus and the Arabia-Eurasia Orogen. Ph.D. Thesis, University of Michigan, Ann Arbor, MI, USA, 2011; 137p. [Google Scholar]
- Cowgill, E.; Forte, A.M.; Niemi, N.; Avdeev, B.; Tye, A.; Trexler, C.; Javakhishvili, Z.; Elashvili, M.; Godoladze, T. Relict basin closure and crustal shortening budgets during continental collision: An example from Caucasus sediment provenance. Tectonics 2016, 35, 2918–2947. [Google Scholar] [CrossRef]
- Adamia, S.; Zakariadze, G.; Chkhotua, T.; Sadradze, N.; Tsereteli, N.; Chabukiani, A.; Gventsadze, A. Geology of the Caucasus: A Review. Turk. J. Earth Sci. 2011, 20, 489–544. [Google Scholar] [CrossRef]
- Artamonova, V.S.; Bolotov, I.N.; Vinarski, M.V.; Makhrov, A.A. Fresh- and Brackish-Water Cold-Tolerant Species of Southern Europe: Migrants from the Paratethys That Colonized the Arctic. Water 2021, 13, 1161. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Niphargus alanicus | N. sadonicus sp. nov. | N. osseticus sp. nov. | N. ardonicus sp. nov. | N. tschertschesovae sp. nov. | N. zeyensis sp. nov. | |
|---|---|---|---|---|---|---|
| N. sadonicus sp. nov. | 0.130 ± 0.016 | |||||
| N. osseticus sp. nov. | 0.154 ± 0.019 | 0.158 ± 0.018 | ||||
| N. ardonicus sp. nov. | 0.158 ± 0.020 | 0.127 ± 0.016 | 0.133 ± 0.017 | |||
| N. tschertschesovae sp. nov. | 0.163 ± 0.018 | 0.086 ± 0.013 | 0.162 ± 0.020 | 0.122 ± 0.015 | ||
| N. zeyensis sp. nov. | 0.160 ± 0.019 | 0.159 ± 0.019 | 0.178 ± 0.019 | 0.170 ± 0.019 | 0.176 ± 0.017 | |
| N. fiagdonicus sp. nov. | 0.191 ± 0.022 | 0.167 ± 0.021 | 0.180 ± 0.021 | 0.118 ± 0.015 | 0.150 ± 0.017 | 0.186 ± 0.019 |
| Species | p-Distance | Estimated Divergence Time (Mya) | ||
|---|---|---|---|---|
| Min. (0.0516/Mya−1) | Average (0.025/Mya−1) | Max. (0.0077/Mya−1) | ||
| Niphargus zeyensis sp. nov. | ||||
| Chchortsku/Matena/Garakha | 0.1495 ± 0.0156 | 2.89 ± 0.30 | 5.98 ± 0.624 | 19.41 ± 2.02 |
| Crimea_Buchya_Cave | 0.1839 ± 0.0178 | 3.56 ± 0.34 | 7.35 ± 0.71 | 23.88 ± 2.31 |
| Niphargus clade (Crimea) | 0.1946 ± 0.0144 | 3.77 ± 0.27 | 7.78 ± 0.57 | 25.27 ± 1.87 |
| Sochi, Plastunka | 0.1949 ± 0.0187 | 3.77 ± 0.36 | 7.79 ± 0.74 | 25.31 ± 2.42 |
| “borutzkyi” species group | 0.2029 ± 0.0167 | 3.93 ± 0.32 | 8.11 ± 0.66 | 26.35 ± 2.16 |
| Niphargus alanicus | ||||
| Niphargus glontii | 0.1856 ± 0.0176 | 3.59 ± 0.34 | 7.42 ± 0.70 | 24.10 ± 2.28 |
| Abraskila Cave, Abkhazia | 0.1971 ± 0.0184 | 3.81 ± 0.35 | 7.88 ± 0.73 | 25.59 ± 2.38 |
| “alasonicus” species group | 0.2112 ± 0.0161 | 4.09 ± 0.31 | 8.44 ± 0.64 | 27.42 ± 2.09 |
| N. ardonicus sp. nov., N. fiagdonicus sp. nov. and N. osseticus sp. nov. (Clade 2) | 0.1790 ± 0.0144 | 3.46 ± 0.28 | 7.16 ± 0.57 | 23.24 ± 1.87 |
| Niphargus sadonicus sp. nov. and N. tschertschesovae sp. nov. (Clade 3) | 0.1828 ± 0.0190 | 3.54 ± 0.37 | 7.31 ± 0.76 | 23.74 ± 2.46 |
| Niphargus ardonicus sp. nov., N. fiagdonicus sp. nov. and N. osseticus sp. nov. (Clade 2) | ||||
| Niphargus glontii | 0.1939 ± 0.0155 | 3.75 ± 0.30 | 7.75 ± 0.62 | 25.18 ± 2.01 |
| Abraskila Cave, Abkhazia | 0.2075 ± 0.0169 | 4.02 ± 0.32 | 8.30 ± 0.67 | 26.94 ± 2.19 |
| “alasonicus” species group | 0.2011 ± 0.0140 | 3.89 ± 0.27 | 8.04 ± 0.56 | 26.11 ± 1.81 |
| Niphargus alanicus | 0.1790 ± 0.0144 | 3.46 ± 0.28 | 7.16 ± 0.57 | 23.24 ± 1.87 |
| N.sadonicus sp. nov. and N. tschertschesovae sp. nov. (Clade 3) | 0.1703 ± 0.0134 | 3,.30 ± 0.26 | 6.81 ± 0.53 | 22.11 ± 1.74 |
| Niphargus sadonicus sp. nov. and N. tschertschesovae sp. nov. (Clade 3) | ||||
| Niphargus glontii | 0.1748 ± 0.0165 | 3.38 ± 0.32 | 6.99 ± 0.66 | 22.70 ± 2.14 |
| Abraskila Cave, Abkhazia | 0.1867 ± 0.0174 | 3.61 ± 0.33 | 7.46 ± 0.69 | 24.24 ± 2.26 |
| “alasonicus” species group | 0.1961 ± 0.0148 | 3.80 ± 0.28 | 7.84 ± 0.59 | 25.46 ± 1.92 |
| N. ardonicus sp. nov., N. fiagdonicus sp. nov. and N. osseticus sp. nov. (Clade 2) | 0.1703 ± 0.0134 | 3.30 ± 0.26 | 6.81 ± 0.53 | 22.11 ± 1.74 |
| Niphargus alanicus | 0.1828 ± 0.0190 | 3.54 ± 0.37 | 7.31 ± 0.76 | 23.74 ± 2.46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marin, I.N.; Palatov, D.M. The Diversity of Freshwater Stygobiotic Crustaceans in the Republic of North Ossetia–Alania Provides New Evidence for the Existence of an Ancient Glacial Refugium in the North Caucasus Region. Water 2024, 16, 1212. https://doi.org/10.3390/w16091212
Marin IN, Palatov DM. The Diversity of Freshwater Stygobiotic Crustaceans in the Republic of North Ossetia–Alania Provides New Evidence for the Existence of an Ancient Glacial Refugium in the North Caucasus Region. Water. 2024; 16(9):1212. https://doi.org/10.3390/w16091212
Chicago/Turabian StyleMarin, Ivan N., and Dmitry M. Palatov. 2024. "The Diversity of Freshwater Stygobiotic Crustaceans in the Republic of North Ossetia–Alania Provides New Evidence for the Existence of an Ancient Glacial Refugium in the North Caucasus Region" Water 16, no. 9: 1212. https://doi.org/10.3390/w16091212